CONCEPTS IN BIOLOGY
PART V. THE ORIGIN AND CLASSIFICATION OF LIFE
20. The Classification and Evolution of Organisms
20.2. A Brief Survey of the Domains of Life
Members of the domains Bacteria and Archaea are all tiny, prokaryotic cells that are difficult to distinguish from one another. Because of this, in the past it was assumed that the members of these groups were closely related. However, recent evidence gained from studying DNA and RNA nucleotide sequences and a comparison of the amino acid sequences of proteins indicate that there are major differences between the Bacteria and Archaea. The Bacteria evolved first and then gave rise to the Archaea; finally, the Eucarya evolved.

The Domain Bacteria
The Bacteria are small, prokaryotic, single-celled organisms ranging in size from 1 to 10 micrometers (pm). Their cell walls contain a complex organic molecule known as peptidoglycan. Peptidoglycan is found only in the Bacteria and is composed of two kinds of sugars linked together by amino acids. One of these sugars, muramic acid, is found only in the cell walls of Bacteria. The most common shapes of the cells are spheres, rods, and spirals.
Because they are prokaryotic, Bacteria have no nucleus. Their genetic material consists of a single loop of DNA. When Bacteria reproduce, the loop of DNA replicates to form two loops, and the cell divides into two, each having one copy of the DNA loop. This method of reproduction is known as binary fission (figure 20.9). Since the two cells have the same DNA, this is a form of asexual reproduction. Many Bacteria are also known to show a kind of sexual reproduction, in which two bacteria exchange pieces of DNA. Although no additional individuals were formed, both individuals have new combinations of genes.
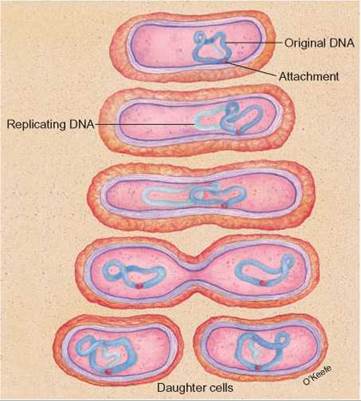
FIGURE 20.9. Asexual Reproduction (Binary Fission) in Bacteria and Archaea
When Bacteria and Archaea reproduce, the loop of DNA replicates and the cell divides, with each cell having one of the two loops of DNA. This form of asexual reproduction is known as binary fission.
Some Bacteria move by secreting a slime that glides over their surface, causing them to move through the environment. Others move by means of a kind of flagellum. The structure of the flagellum is different from the flagellum found in eukaryotic organisms.
Because the early atmosphere is thought to have been a reducing atmosphere, the first Bacteria were probably anaerobic organisms. Today, there are both anaerobic and aerobic Bacteria. Many heterotrophic Bacteria are saprophytes, organisms that obtain energy by the decomposition of dead organic material; others are parasites that obtain energy and nutrients from living hosts and cause disease (Outlooks 20.1); still others are mutualistic and have a mutually beneficial relationship with their host; finally, some are commensalistic and derive benefit from a host without harming it. Several kinds of Bacteria are autotrophic. Many are called cyanobacteria because they contain a blue-green pigment, which allows them to capture sunlight and carry on photosynthesis. They can become extremely numerous in some polluted waters where nutrients are abundant. Others use inorganic chemical reactions for their energy sources and are called chemosynthetic.
OUTLOOKS 20.1
A Bacterium that Controls Animal Reproduction
Wolbachia is a small kind of bacterium called a rickettsia. It is an intracellular parasite that only lives in the cells of its hosts— many kinds of insects, other arthropods, and nematode worms. Wolbachia is extremely common—at least 20% of all insects are infected with this parasite. Because it is an intracellular parasite, it is most easily transmitted from one generation of its host to the next during reproduction. Since the eggs of the host species carry Wolbachia but the sperm do not, Wolbachia is passed from one generation of its host to the next only in the egg of the host as if it were an organelle in the egg. Wolbachia manipulates the reproductive biology of its host in several ways that enhance the likelihood that Wolbachia-infected female hosts will produce large numbers of Wolbachia-infected female offspring.
1. Egg/sperm incompatibility
When a female insect uninfected with Wolbachia mates with a male infected with Wolbachia, some or all of her fertilized eggs die. This means that uninfected female insects produce fewer offspring than infected females. Thus, there is a bias toward producing Wolbachia-infected offspring.
2. Killing of host males
In some cases, infected male hosts are killed by the presence of the parasite. The killing of male hosts means that only infected females are produced.
3. Changing the sex of genetic male hosts
In some cases, genetic male host offspring that are infected with Wolbachia are converted to females. If they are fertile females, they will produce Wolbachia-containing eggs. If they are infertile pseudofemales, they do not reproduce.
4. Parthenogenetic development
In certain species of wasps, it appears that infection with Wolbachia has resulted in the complete elimination of males. The female wasps reproduce by parthenogenesis, which is a form of reproduction in which unfertilized eggs develop into only female offspring. Thus, all of the female wasps offspring are infected with Wolbachia.
5. Mutualism
Some species of nematode worms are unable to reproduce without the presence of Wolbachia.
These examples suggest that the Wolbachia DNA has taken over some of the functions of its host. In fact, examination of the DNA of the eukaryotic host species shows that they contain Wolbachia genes in their nucleus.
The Domain Archaea
The term archaea comes from the Greek word archaios, meaning ancient. This is a little misleading, because the Archaea are thought to have branched off from the Bacteria between 2 and 3 billion years ago.
The Archaea are similar to the Bacteria in that they both have a prokaryotic cell structure. However, the Archaea differ from the Bacteria in several fundamental ways. The Archaea do not have peptidoglycan in their cell walls, and the structure of their DNA is different from that of the Bacteria. Although the DNA of the Archaea is a loop, like that of the Bacteria, the DNA of the Archaea appears to have a large proportion of genes that are different from either the Bacteria or the Eucarya. Archaea reproduce asexually by binary fission, as do the Bacteria. They also exchange pieces of DNA between individuals, as do Bacteria. Also, the cell membranes of the Archaea have a unique chemical structure, found in neither the Bacteria nor the Eucarya. Members of the Archaea exist in many shapes, including rods, spheres, spirals, filaments, and flat plates.
Because many members of the Archaea are found in extreme environments, they have become known as extremophiles. However, as more species are discovered and organisms that were once thought to be Bacteria are reclassified as Archaea, it is becoming clear there are many that do not live in extreme environments. Archaea use a variety of ways of obtaining energy. Many are autotrophs that use inorganic chemical reactions (chemoauto- trophs) or light (photoautotrophs) as sources of energy and carbon dioxide as a source of carbon. Some are heterotrophs and use organic molecules as a source of energy and carbon.
Members of the Archaea are extremely diverse. Based on the particular habitats they occupy and the kind of metabolism they display, Archaea are divided into several functional groups:
1. Methanogens are anaerobic, methane-producing organisms. They can be found in sewage, swamps, and the i ntestinal tracts of termites and ruminant animals, such as cows, sheep, and goats. They are even found in the intestines of humans.
2. Halobacteria (halo = salt) live in very salty environments, such as the Great Salt Lake (Utah), salt ponds, and brine solutions. Many have a reddish pigment and can be present in such high numbers that they color the water red. Some contain a special kind of chlorophyll and are therefore capable of generating their ATP by a kind of photosynthesis but they do not release oxygen.
3. Thermophilic Archaea live in environments that normally have very high temperatures and high concentrations of sulfur (e.g., hot sulfur springs and around deep-sea hydrothermal vents). Over 500 species of thermophiles have been identified at the openings of hydrothermal vents in the open oceans. One such thermophile, Pyrolobus fumarii, grows in a hot spring in Yellowstone National Park (figure 20.10). It grows best at 106°C and can grow at temperatures up to 113°C but will not grow below 90°C. Another species that survives at 122°C has been discovered. Some of these heat-loving Archaea also live in extremely acid conditions.

FIGURE 20.10. Habitat for Thermophilic Archaea
Thermophilic Archaea are found in hot springs like these at Yellowstone National Park and around thermal vents on the ocean floor.
4. Marine, freshwater, and soil Archaea have recently been discovered to be extremely abundant, but little is yet known about their role in these habitats.
5. Recently an archeon has been discovered that appears to be parasitic on another archeon.
The Domain Eucarya
Most biologists now believe that eukaryotic cells evolved from prokaryotic cells through endosymbiosis (see chapter 19). This hypothesis proposes that structures such as mitochondria, chloroplasts, and several other membranous organelles found in eukaryotic cells were originally separate living organisms that were incorporated into another cell. Once inside another cell, these structures and their functions became integrated with the host cell and ultimately became essential to its survival. This new type of cell was the forerunner of present-day eukaryotic cells.
Eukaryotic cells are usually much larger than prokaryotic cells, typically having more than a thousand times the volume of prokaryotic cells. Their larger size was made possible by the presence of specialized membranous organelles, such as the endoplasmic reticulum, mitochondria, chloroplasts, and nuclei.
Kingdom Protista
The changes in cell structure that led to eukaryotic organisms probably gave rise to single-celled organisms similar to those currently grouped in the kingdom Protista.
There is a great deal of diversity among the approximately 60,000 known species of Protista. Many species live in freshwater; others are found in marine or terrestrial habitats, and some are parasitic, commensalistic, or mutualistic. All species can undergo mitosis, resulting in asexual reproduction. Some species can also undergo meiosis and reproduce sexually. Many are autotrophs that have chloroplasts and carry on photosynthesis. These are commonly called algae. Others are heterotrophs that require organic molecules as sources of energy. These are commonly called protozoa. Both autotrophs and heterotrophs have mitochondria and respire aerobically.
Because members of this kingdom are so diverse with respect to details of cell structure, metabolism, and reproductive methods, most biologists do not think that the Protista form a valid phylogenetic unit and should be divided into several distinct kingdoms. However, it is still a convenient taxonomic grouping. By placing these organisms together in this group, it is possible to gain a useful perspective on how they relate to other kinds of organisms. After the origin of eukaryotic organisms, evolution proceeded along several pathways. Three major lines of evolution within the Protista can be seen. There are plantlike autotrophs (algae), animal-like heterotrophs (protozoa), and funguslike heterotrophs (slime molds). Amoeba and Paramecium are commonly encountered examples of protozoa. Many seaweeds and pond scums are collections of large numbers of algal cells. Slime molds are seen less frequently, because they live in and on the soil in moist habitats; they are most often encountered as slimy masses on decaying logs. Figure 20.11 shows some examples of this diverse group of organisms.

FIGURE 20.11. A Diversity of Protista
The kingdom Protista includes a wide variety of organisms that are simple in structure. They are not a phylogenetic group.
Kingdom Fungi
Fungus is the common name for members of the kingdom Fungi. The ancestors of fungi were probably related to some of the heterotrophic protozoa. They have evolved several distinct characteristics. Most fungi are nonmotile. They have a rigid, thin cell wall, which contains chitin, a complex carbohydrate containing nitrogen. The members of the kingdom Fungi are nonphotosynthetic, eukaryotic organisms. Most of the approximately 70,000 species (mushrooms and molds) are multicellular, but a few, such as yeasts, are single-celled. In the multicellular fungi, the basic structural unit is a network of multicellular filaments.
Because all of these organisms are heterotrophs, they must obtain nutrients from organic sources. Most are saprophytes and secrete enzymes that digest large molecules into smaller units, which are absorbed as food for the organism. They are very important as decomposers in all ecosystems. They feed on a variety of nutrients ranging from dead organisms to such products as shoes, foodstuffs, and clothing. Most synthetic organic molecules are not attacked as readily by fungi; this is why plastic bags and foam cups are slow to decompose.
Some fungi are parasitic. Many of the parasitic fungi are important plant pests. Some (e.g., chestnut blight, Dutch elm disease) attack and kill plants; others injure the fruit, leaves, roots, or stems. In crop plants, fungi are important pests that kill or weaken plants and reduce yields. The fungi that are human parasites are responsible for athlete’s foot, vaginal yeast infections, valley fever, “ringworm,” and other diseases.
Other fungi are mutualistic. Lichens are common organisms that consist of a mutualistic relationship between fungal and algal cells. Another important group of mutualistic fungi form an association with the roots of plants and improve the nutrient and water capturing capacity of plants. Indeed many species of plants will not grow without their fungal associates. Figure 20.12 shows examples of several kinds of fungi.

FIGURE 20.12. Examples of Fungi
Molds, mushrooms, and puffballs are common examples of this kingdom.
Kingdom Plantae
The members of the kingdom Plantae are nonmotile, terrestrial (mostly), multicellular organisms that contain chlorophyll and produce their own organic compounds by photosynthesis. All plant cells have a cellulose cell wall.
Most biologists believe that the evolution of this kingdom began nearly 500 million years ago, when the green algae gave rise to nonvascular plants, such as mosses. The development of vascular (water-transporting) tissue was a major step in the evolution of plants, because it allowed plants to increase in size and to live in drier environmental conditions. Ferns, cone-bearing plants, and flowering plants all have vascular tissue. Some of the vascular plants evolved into seed-producing plants, which today are the cone-bearing and flowering plants, whereas the ferns never developed seeds (figure 20.13). Over 300,000 species of plants have been classified; about 85% are flowering plants, 14% are mosses and ferns, and the remaining 1% are cone-bearers and several other small groups within the kingdom.
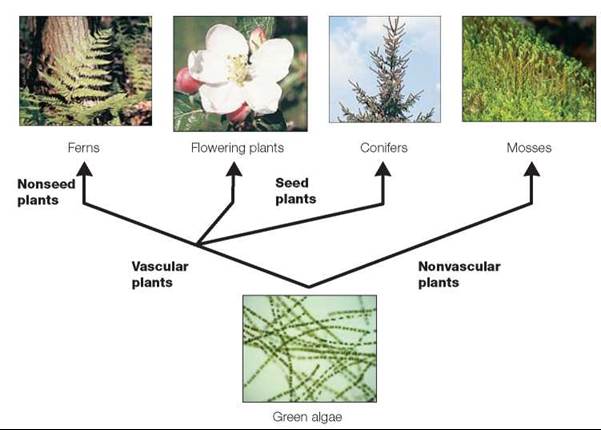
FIGURE 20.13. Plant Evolution
Two lines of plants are thought to have evolved from the green algae of the kingdom Protista. The nonvascular mosses evolved as one type of plant. The second type, the vascular plants, evolved into more complex plants. The early vascular plants did not produce seeds, like present-day ferns. Eventually, the evolution of the ability to produce seeds resulted in two distinct lines: the cone-bearing plants and the flowering plants.
A major unifying theme in the plants and their green alga ancestors is the presence of a unique life cycle with two distinct generation or stages. They have a haploid gametophyte generation, which produces haploid sex cells by mitosis. The sex cells unite to form a diploid zygote that grows into a diploid sporophyte generation. The sporophyte generation produces haploid spores by meiosis and the haploid spores germinate and grow into the gametophyte generation. This kind of life cycle is referred to as alternation of generations. In addition to sexual reproduction, plants are able to reproduce asexually.
Kingdom Animalia
Over a million species of animals have been classified, ranging from microscopic types, such as mites and the aquatic larvae of marine animals, to huge animals, such as elephants and whales. All animals have some common traits. All are composed of eukaryotic cells and all species are heterotrophic and multicellular. All animals are motile, at least during some portion of their lives, although some, such as the sponges, barnacles, mussels, and corals, are sessile (i.e., nonmotile) as adults. All animals are capable of sexual reproduction, but many of the less complex animals also reproduce asexually.
The ancestors of the kingdom Animalia are thought to be certain kinds of flagellated protozoa. It is thought that colonies of flagellated protozoa gave rise to simple, multicellular forms of animals such as the ancestors of present-day sponges. These first animals lacked specialized tissues and organs. As cells became more specialized, organisms developed special tissues, organs, and systems of organs, and the variety of kinds of animals increased (figure 20.14).

FIGURE 20.14. Animal Diversity
Animals range in complexity from simple, marine sponges and sea anemones to complex, terrestrial insects, birds, and mammals.
20.2. CONCEPT REVIEW
6. List two ways that Bacteria and Archaea differ.
7. Describe two distinctly different ways that Bacteria obtain energy.
8. Describe two distinctly different ways that Archaea obtain energy.
9. List the four kingdoms of the domain Eucarya and give two distinguishing characteristics for each.
10. Which of the following groups of organisms contain members that are autotrophic: Bacteria, Archaea, Protista, Fungi, Plantae, Animalia?