CONCEPTS IN BIOLOGY
PART IV. EVOLUTION AND ECOLOGY
13. Evolution and Natural Selection
13.6. The Processes. That Drive Selection
Several mechanisms allow for the selection of certain individuals for successful reproduction. If predators must pursue swift prey organisms, the faster predators will be selected for, and the selecting agent is the swiftness of available prey. If predators must find prey that are slow but hard to see, the selecting agent is the camouflage coloration of the prey, and keen eyesight is selected for. If plants are eaten by insects, the production of toxic materials in the leaves is selected for. All selecting agents influence the likelihood that certain characteristics will be passed on to subsequent generations.
Differential Survival
As stated previously, the phrase “survival of the fittest” is often associated with the theory of natural selection. Although this is recognized as an oversimplification of the concept, survival is an important factor in influencing the flow of genes to subsequent generations. If a population consists of a large number of genetically and phenotypically different individuals it is likely that some of them will possess characteristics that make their survival difficult. Therefore, they are likely to die early in life and not have an opportunity to pass on their genes to the next generation.
Charles Darwin described several species of ground finches on the Galapagos Islands (figure 13.7), and scientists have often used these birds in scientific studies of evolution. On one of the islands, scientists studied one of the species of seed-eating ground finches, Geospiza fortis. They measured the size of the animals and the size of their bills and related these characteristics to their survival. They found the following: During a drought, the birds ate the smaller, softer seeds more readily than the larger, harder seeds. As the birds consumed the more easily eaten seeds, only the larger, harder seeds remained. During the drought, finch mortality was extremely high. When scientists looked at ground finch mortality, they found that the larger birds with stronger, deeper bills survived better than the smaller birds with weaker, narrower bills. They also showed that the offspring of the survivors tended to show larger body and bill size as well. The lack of small, easily eaten seeds resulted in selection for larger birds with stronger bills, which could crack open larger, tougher seeds. Table 13.1 shows data on two of the parameters measured in this study.

FIGURE 13.7. A Sample of Darwin's Finches
Ten species of ground and tree finches were described by Darwin. This figure shows six members of the genus Geospiza. The four species of ground finches are primarily seedeaters and use their bills to crush seeds. The two species of cactus finch primarily feed on the fruit and flowers of cactus plants.
TABLE 13.1. Changes in Body Structure of Geospiza fortis
|
Before Drought |
After Drought |
Average Body Weight |
16.06 g |
17.13 g |
Average Bill Depth |
9.21 mm |
9.70 mm |
Body Structure
As another example of how differential survival can lead to changed gene frequencies, consider what has happened to many insect populations as humans have subjected them to a variety of insecticides. Because there is genetic diversity within all species of insects, an insecticide that is used for the first time on a particular species kills all the exposed individuals that are genetically susceptible. However, individuals with slightly different genetic compositions and those not exposed may not be killed by the insecticide.
Suppose that, in a population of a particular species of insect, 5% of the individuals have genes that make them resistant to a specific insecticide. The first application of the insecticide could, therefore, kill a majority of those exposed. However, tolerant individuals and those that escaped exposure would then constitute the remaining breeding population. This would mean that many more insects in the second generation would be tolerant. The second use of the insecticide on this population would not be as effective as the first. With continued use of the same insecticide, each generation would become more tolerant, because the individuals that were not tolerant were being eliminated and those that could tolerate the toxin passed on their genes for tolerance to their offspring.
Many species of insects produce a new generation each month. In organisms with a short generation time, 99% of the population could become resistant to an insecticide in just 5 years. As a result, the insecticide would no longer be useful in controlling the species. As a new selecting agent (the insecticide) is introduced into the insect’s environment, natural selection results in a change in the gene frequency of a population, so that most individuals are tolerant of the insecticide.
The same kind of selection process has occurred with herbicides. Within the past 50 years, many kinds of herbicides have been developed to control weeds in agricultural fields. After several years of use, a familiar pattern develops as more and more species of weeds show resistance to the herbicide. Figure 13.8 shows several kinds of herbicides and the number of weed species that have become resistant over time. In each weed species, there has been selection for the individuals that have the genetic information that allows them to tolerate the presence of the herbicide.
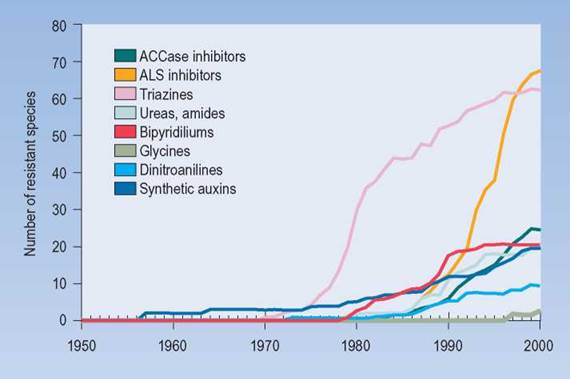
FIGURE 13.8. Evolutionary Change
Populations of weed plants that have been subjected repeatedly to herbicides often develop resistant populations. The individual weed plants that have been able to resist the effects of the herbicide have lived to reproduce and pass on their genes for resistance to their offspring; thus, resistant populations of weeds have developed.
Differential Reproductive Rates
Survival alone does not always ensure reproductive success. For a variety of reasons, some organisms are better able to use the available resources to produce offspring. If an individual leaves 100 offspring and another leaves only 2, the first organism has passed on more copies of its genetic information to the next generation than has the second. If we assume that all 102 offspring have similar survival rates, the first organism has been selected for, and its genes have become more common in the subsequent population.
Scientists have studied the gene frequencies for the height of clover plants. Two i dentical fields of clover were planted and cows were allowed to graze in one of them. The cows acted as a selecting agent by eating the taller plants first. These tall plants rarely got a chance to reproduce. Only the shorter plants flowered and produced seeds. After some time, seeds were collected from both the grazed and the ungrazed fields and grown in a greenhouse under identical conditions. The average height of the plants from the ungrazed field was compared with that of the plants from the grazed field. The seeds from the ungrazed field produced some tall, some short, but mostly medium-sized plants. However, the seeds from the grazed field produced many more short plants than medium or tall ones. The cows had selectively eaten the tall plants. Because the flowers are at the tip of the plant, the tall plants were less likely to successfully reproduce, even though they were able to survive grazing by cows.
Differential Mate Choice—Sexual Selection
Sexual selection occurs within animal populations when some individuals are more likely to be chosen as mates than others. Obviously, those that are frequently chosen have more opportunities to pass on more copies of their genetic information than those that are rarely chosen. The characteristics of the more frequently chosen individuals may involve general characteristics, such as body size or aggressiveness, or specific, conspicuous characteristics attractive to the opposite sex.
For example, male red-winged blackbirds establish territories in cattail marshes, where females build their nests. A male will chase out all other males, but not females. Some blackbird territories are large and others are small; some males have none. Although it is possible for any male to mate, those that have no territory are least likely to mate. Those that defend large territories may have two or more females nesting in their territories and are very likely to mate with those females. It is unclear exactly why females choose one male’s territory over another, but the fact is that some males are chosen as mates and others are not.

In other cases, it appears that females select males that display specific, conspicuous characteristics. Certain male birds, such as peacocks, have very conspicuous tail feathers (figure 13.9). Those with spectacular tails are more likely to mate and have offspring. Darwin was puzzled by such cases, because the large, conspicuous tail should have been a disadvantage to the bird. Long tails require energy to produce, make it more difficult to fly, and make it more likely that predators will capture the individual. The current theory that seeks to explain this paradox involves female choice. If the females have an innate (genetic) tendency to choose the most elaborately decorated males, genes that favor such plumage will be regularly passed on to the next generation.

FIGURE 13.9. Mate Selection
In many animal species the males display very conspicuous characteristics that are attractive to females. Because the females choose the males they will mate with, those males with the most attractive characteristics will have more offspring and, in future generations, there will be a tendency to enhance the characteristic. With peacocks, those individuals with large colorful displays are more likely to mate.
13.6. CONCEPT REVIEW
12. List three factors that can lead to changed gene frequencies from one generation to the next.
13. Give two examples of selecting agents and explain how they operate.