CONCEPTS IN BIOLOGY
PART IV. EVOLUTION AND ECOLOGY
14. The Formation of Species and Evolutionary Change
14.4. Evolutionary Patterns Above the Species Level
A species is the smallest irreversible unit of evolution. Because the exact conditions present when a species came into being will never exist again, it is highly unlikely that it will evolve back into an earlier stage in its development. Furthermore, because species are reproductively isolated from one another, they usually do not combine with other species to make something new; they can only diverge further. When life in the past is compared with our current diversity of life, many different evolutionary patterns emerge.
Divergent Evolution
Divergent evolution is a basic evolutionary pattern in which individual speciation events cause successive branches in the evolution of a group of organisms. This basic pattern is well illustrated by the evolution of the horse, shown in figure 14.6. Each of the many branches of the evolutionary history of the horse began with a speciation event that separated one species into two species as each separately adapted to local conditions. Changes in the environment from moist forests to drier grasslands would have set the stage for change. The modern horse, with its large size, single toe on each foot, and teeth designed for grinding grasses, is thought to be the result of accumulated genetic changes beginning from a small, dog-sized animal with four toes on its front feet, three toes on its hind feet, and teeth designed for chewing leaves and small twigs. Even though we know much about evolution, there are still many gaps that need to be filled before we have a complete evolutionary history of any organism (Outlooks 14.1).
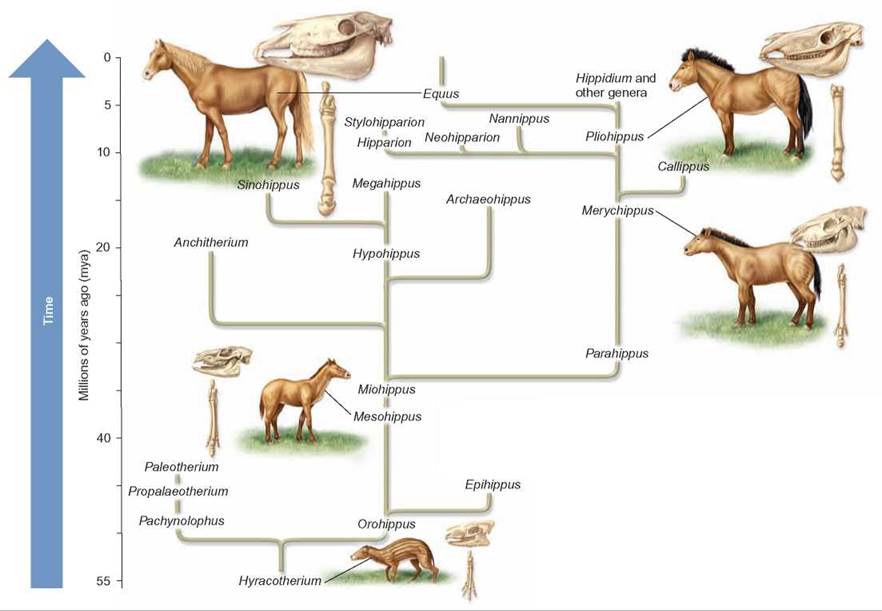
FIGURE 14.6. Divergent Evolution
In the evolution of the horse, many speciation events occurred. What began as a small, leaf-eating, fourtoed animal of the forest evolved into a large, grasseating, single-toed animal of the plains. There are many related species alive today (e.g., horse, donkey, zebra), but early ancestral types are extinct. An analysis of fossilized bones provided the phylogenetic tree described here.
OUTLOOKS 14.1
Evolution and Domesticated Cats
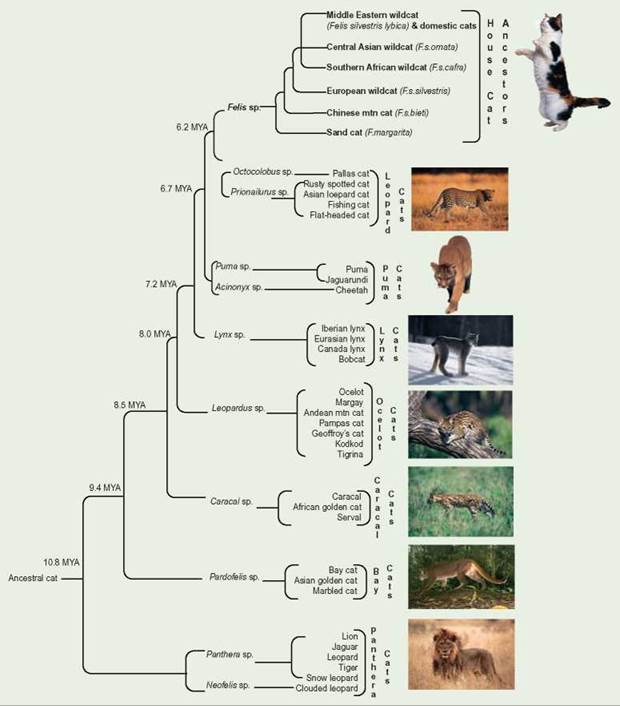
The evolutionary history of the domestic cat (Felis silvestris lybica) has been unclear because of incomplete fossil records. However, an international team sampling both mitochondrial DNA and the DNA from both X and Y sex chromosomes has finally come up with an evolutionary tree for felines. The group proposes that about 11 million years ago, a single, ancestral feline-like species migrated from Asia throughout the world except Australia.
Researchers believe that 10 to 3 million years ago (MYA) land bridges between continents were created when sea levels fell. The common ancestor to all of today's cats probably migrated south to Africa from Asia. The cats also moved north crossing the Bering land bridge (as wide as 1,000 miles) to North America and migrated to South America by the Panamanian land bridge. When sea levels rose, they covered the land bridges and cut off cat species from their original groups. These isolated subpopulations genetically drifted apart, each adapting to its unique environment. When the subpopulations had the chance of coming back together, they were no longer able to interbreed and, at that point, found themselves to be different species. Ancestral felines, originally a Eurasian genus, successfully migrated throughout the globe because they encountered little or no competition from other carnivores. They continue to be one of the most successful of carnivore families.
Traditionally, domestication was thought to have occurred about 3,600 years ago in Egypt. Archeological evidence in Egyptian hieroglyphics portrays cats, and bones of cats have been found buried with humans in tombs. However, more recent archeological and genetic evidence strongly suggests that cats were domesticated about 9,500 years ago in the Fertile Crescent, the Middle East. Today, this region includes Egypt, Israel, the West Bank, Gaza strip, and Lebanon; and parts of Jordan, Syria, Iraq, southeastern Turkey, and southwestern Iran and Kuwait.
Extinction
Extinction is the loss of a species. It is a common pattern in the evolution of organisms. Notice in figure 14.6 that most of the species that developed during the evolution of the horse are extinct. Only members of the genus Equus remain. This is typical. Most of the species that have ever existed are extinct. Estimates of extinction are around 99%; that is, 99% or more of all the species that ever existed are extinct. Given this high rate of extinction, we can picture current species of organisms as the product of much evolutionary experimentation. This is not the complete picture, though. Recall from chapter 13 that organisms are continually being subjected to selection pressures that lead to a high degree of adaptation to a particular set of environmental conditions. Organisms become more and more specialized. However, the environment does not remain constant; it often changes in such a way that the species that were originally present are unable to adapt to the new set of conditions. The early ancestors of the modern horse were well adapted to a moist tropical environment, but, when the climate became drier, most were no longer able to survive. Only some kinds had the genes necessary to lead to the development of modern horses and their relatives.
Furthermore, many extinct species were very successful organisms for millions of years. They were not failures for their time but simply did not survive to the present. It is also important to realize that many currently existing organisms will eventually become extinct—perhaps even Homo sapiens. Thus, the basic evolutionary pattern is one of divergence with a great deal of extinction. Although divergence and extinction are dominant themes in the evolution of life, adaptive radiation and convergent evolution are two other important evolutionary patterns.
Adaptive Radiation
Adaptive radiation is an evolutionary pattern characterized by a rapid increase in the number of kinds of closely related species. Adaptive radiation results in an evolutionary explosion of new species from a common ancestor. There are basically two situations thought to favor adaptive radiation. One is a condition in which an organism invades a previously unexploited environment. For example, at one time, there were no animals on the landmasses of the Earth. The amphibians were the first vertebrate animals able to spend part of their lives on land. Fossil evidence shows that a variety of amphibians evolved rapidly and exploited several kinds of lifestyles.
Another good example of adaptive radiation is found among the finches of the Galapagos Islands, located 1,000 kilometers west of Ecuador in the Pacific Ocean. These birds were first studied by Charles Darwin. Because these islands are volcanic and arose from the ocean floor, it is assumed that they have always been isolated from South America and originally lacked finches and other land-based birds. It is thought that one kind of finch arrived from South America to colonize the islands and that adaptive radiation from the common ancestor resulted in the many kinds of finches found on the islands today (figure 14.7). Although the islands are close to one another, they are quite diverse. Some are dry and treeless, some have moist forests, and others have intermediate conditions. Conditions were ideal for several speciation events. Because the islands were separated from one another, the element of geographic isolation was present. Because environmental conditions on the islands were quite different, particular characteristics in the resident birds would have been favored. Furthermore, the absence of other kinds of birds meant that there were many lifestyles that had not been exploited.
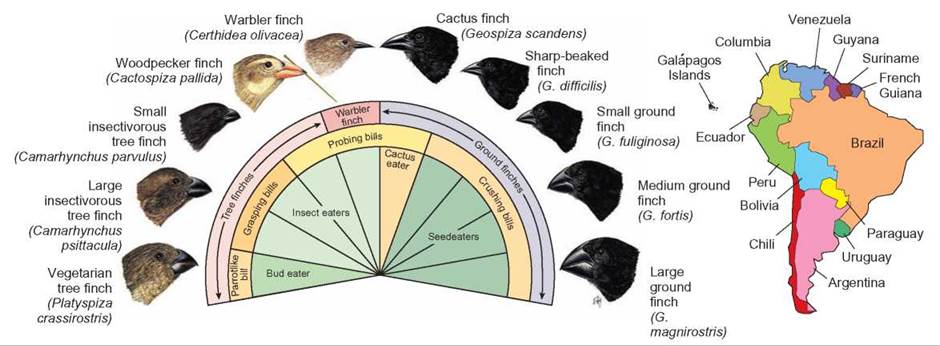
FIGURE 14.7. Adaptive Radiation
When Darwin discovered the finches of the Galapagos Islands, he thought they might all have derived from one common ancestor that arrived on these relatively isolated islands. If they were the only birds to inhabit the islands, they could have evolved very rapidly into the many types shown here. The drawings show the specializations of beaks for different kinds of food.
In the absence of competition, some of these finches took roles normally filled by other kinds of birds elsewhere in the world. Although finches are normally seed-eating birds, some of the Galapagos finches became warblerlike insect-eaters, others became leaf-eaters, and one uses a cactus spine as a tool to probe for insects.
The second situation that can favor adaptive radiation is one in which a type of organism evolves a new set of characteristics that enables it to displace organisms that previously filled specific roles in the environment. For example, although amphibians were the first vertebrates to occupy land, they lived only near freshwater, where they would not dry out and could lay eggs, which developed in the water. They were replaced by reptiles with such characteristics as dry skin, which prevented the loss of water, and an egg that could develop on land. The adaptive radiation of reptiles was extensive. They invaded most terrestrial settings and even evolved forms that flew and lived in the sea. With the extinction of dinosaurs and many other reptiles, birds and mammals went through a similar radiation. Perhaps the development of homeothermism (the ability to maintain a constant body temperature) had something to do with the success of birds and mammals. Figure 14.8 shows the radiations that occurred within the vertebrate group. The number of species of bony fishes has increased as the number of other kinds of fishes (jawless fishes, placoderms, cartilaginous fishes) declined and the number of species of birds and mammals has increased as the number of reptiles and amphibians decreased.
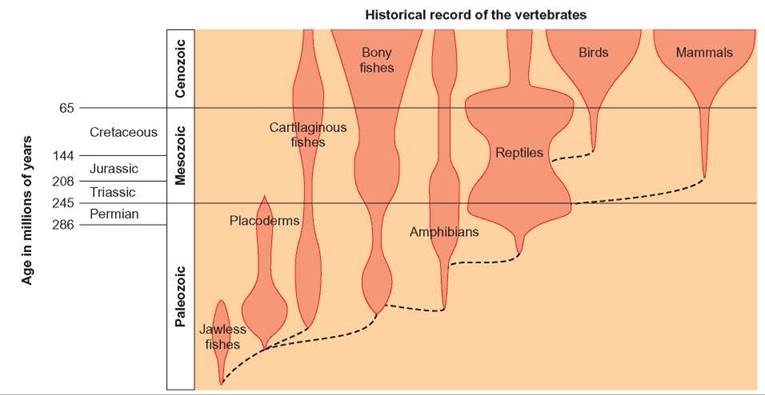
FIGURE 14.8. Adaptive Radiation in Terrestrial Vertebrates
The amphibians were the first vertebrates to live on land. They were replaced by the reptiles, which were better adapted to land. The reptiles, in turn, were replaced by the adaptive radiation of birds and mammals. (Note: The width of the colored bars indicates the number of species present.)
Convergent Evolution
Convergent evolution is an interesting evolutionary pattern that involves the development of similar characteristics in organisms of widely different evolutionary backgrounds. This pattern often leads people to misinterpret the evolutionary history of organisms. For example, many kinds of plants that live in desert situations have sharp, pointed structures, such as spines or thorns, and lack leaves during much of the year. Superficially, the sharp, pointed structures may resemble one another to a remarkable degree but may have a completely different evolutionary history. Some of these structures are modified twigs, others are modified leaves, and still others are modifications of the surface of the stem. The presence of sharp, pointed structures and the absence of leaves are adaptations to a desert type of environment: The thorns and spines discourage herbivores and the absence of leaves reduces water loss.
Another example is animals that survive by catching insects while flying. Bats, swallows, and dragonflies all obtain food in this manner. They have wings, good eyesight or hearing to locate flying insects, and great agility and speed in flight, but they are evolved from quite different ancestors (figure 14.9). At first glance, they may appear very similar and perhaps closely related, but a detailed study of their wings and other structures shows that they are different kinds of animals. They have simply converged in structure, the type of food they eat, and their method of obtaining food. Likewise, whales, sharks, and the barracuda appear to be similar—they all have a streamlined shape, which aids in rapid movement through the water; a dorsal fin, which helps prevent rolling; fins or flippers for steering; and a large tail, which provides power for swimming. However, they are quite different kinds of animals that happen to live in the open ocean, where they pursue other animals as prey. Their structural similarities are adaptations to being fast-swimming predators.
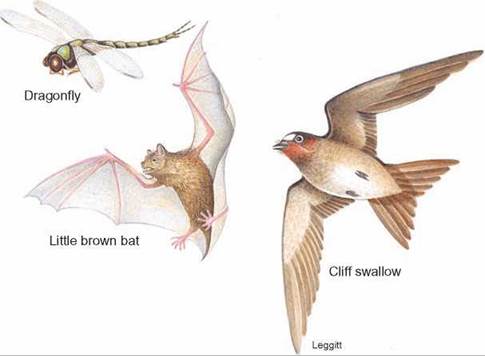
FIGURE 14.9. Convergent Evolution
All of these animals have evolved wings as a method of movement and capture insects for food as they fly. However, flight originated independently in each of them.
Homologous or Analogous Structures
To help evolutionary biologists distinguish between structures that are the result of convergent evolution and those that are not, they try to determine if the structures evolved from a common ancestor. Homologous structures are structures in different species that have been derived from a common ancestral structure. Thus, the wing of a bat, the front leg of a horse, and the arm of a human show the same basic pattern of bones, but they are extreme modifications of the same basic evolutionary structure (figure 14.10). On the other hand, structures that have the same function (such as the wing of a butterfly and the wing of a bird) but different evolutionary backgrounds are called analogous structures and are the result of convergent evolution (review figure 14.9).
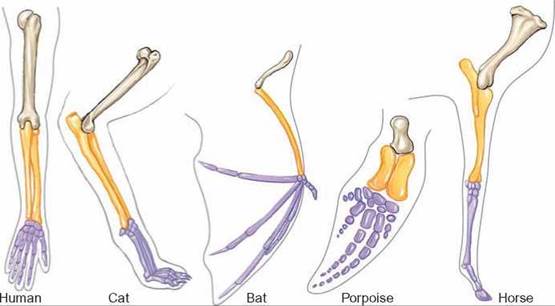
FIGURE 14.10. Maintaining Traits Through Time—Homologous Structures
Although these body parts are considerably different in structure and function, the same bones are present in the forelimbs of humans, cats, bats, porpoises, and horses.
14.4. CONCEPT REVIEW
10. Describe convergent evolution and adaptive radiation.
11. What are the two dominant evolutionary patterns?
12. How are analogous and homologous structures different?