Biology For Dummies
Part IV Systems Galore! Animal Structure and Function
Chapter 19
Reproduction 101: Making More Animals
In This Chapter
Becoming aware of how asexual reproduction occurs in animals
Getting an up-close-and-personal look at sexual reproduction
Seeing how the birds and bees (and other animals) do it
Figuring out how a cell “knows” where to go and what to do
Exploring the complexities of development and aging
This chapter is your chance to find out all about how babies are made. It covers the nuts and bolts of how animals reproduce (we explain how plants reproduce in Chapter 20). Get ready to discover what happens inside a female’s body to prepare for reproduction, how different types of animals (including humans) actually mate, how offspring develop prior to birth, and how gender is determined in humans.
This Budding’s for You: Asexual Reproduction
Asexual reproduction allows organisms to reproduce rapidly and without a partner, which makes asexual organisms essentially just fresher, younger versions of their original selves. Also, asexual organisms don’t really die; instead, they just bud off into new versions of themselves and continue on.
 The basic cellular process that makes asexual reproduction possible is mitosis, the type of cell division that produces exact copies of parent cells (see Chapter 6 for details).
The basic cellular process that makes asexual reproduction possible is mitosis, the type of cell division that produces exact copies of parent cells (see Chapter 6 for details).
Asexual reproduction occurs by several different methods in a variety of animals:
Budding happens when a small outgrowth begins on the original organism. That outgrowth gradually becomes larger and eventually separates to create a new individual. Several species of invertebrates, including the hydra, produce offspring by budding.
Fission occurs when the original organism grows larger and then splits in two. Sea anemones are an example of an invertebrate that reproduces asexually by fission.
Fragmentation happens when small pieces of the original organism break off and then grow into complete individuals. Starfish are among the animals that use fragmentation to reproduce.
For organisms that are widely separated from others of their kind and for organisms that are doing well in a particular environment, asexual reproduction poses quite the advantage. Yet what makes asexual reproduction an asset for some species — the fact that it doesn’t allow for change — also makes it a disadvantage. If a disease strikes or the environment changes and all the organisms are identical, they’ll all be affected equally. If the disease can easily kill the organisms, for example, they’ll all die. If they were the only organisms of their kind, then an entire species would be wiped out in one fell swoop. Species ultimately have a better chance of surviving changes if their members have some differences from one another.
The Ins and Outs of Sexual Reproduction
Whereas asexual reproduction involves the entire organism (one whole organism splits into other whole organisms), sexual reproduction starts at the cellular level and involves two parents. The new organisms that result from sexual reproduction develop and grow over time.
In animals, sexual reproduction begins with an egg and a sperm, each of which is a single cell. Mating combines the two single cells to produce an entirely new organism that contains either eggs or sperm. Therefore, the new organism can continue the life cycle and contribute its genetic material to yet another generation of the species.
The following sections delve into the details of sexual reproduction, from the cells and mating rituals that make it possible to how humans go about creating new life. (We focus on humans rather than other species because that’s obviously what’s most applicable to you.)
Getting to know gametes
Gametes are the sex cells of sexually reproducing organisms. Two kinds of gametes exist: sperm and eggs. Each sperm and each egg contain half the number of chromosomes that are normally present in the whole organism. When a sperm and an egg join, the organism has all the chromosomes it needs.
Gametogenesis is the process that produces gametes. During gametogenesis, meiosis occurs to reduce the number of chromosomes in the cells by half (see Chapter 6 for a breakdown of meiosis). Human cells, for example, have 46 chromosomes, so human gametes have only 23. The entire process of gametogenesis is controlled by hormones, those protein-containing substances that start, stop, and alter many metabolic processes (flip to Chapter 18 for more on hormones). We explain exactly how gametogenesis works in the sections that follow.
Note: We focus our explanation on humans, but the process of gametogenesis works pretty much the same in all animals that undergo sexual reproduction. The differences lie in how the animals mate and transfer their gametes.
Spermatogenesis: Making little swimmers
Sperm are the male gametes, and spermatogenesis is the metabolic process that produces them. Beginning at puberty, human males start producing millions of sperm each day. The sperm only survive inside the male for a short time, which is why they need to be produced continually.
The process of spermatogenesis begins in spermatogonia, cells containing 46 chromosomes. These cells line the walls of the seminiferous tubules. When a human male’s hormones pull the trigger to initiate spermatogenesis, the spermatogonia go through mitosis to produce cells calledprimary spermatocytes that also contain 46 chromosomes. Here’s how the rest of the process goes down from there:
1. Each primary spermatocyte undergoes meiosis to produce two secondary spermatocytes.
Secondary spermatocytes are cells that contain 23 replicated chromosomes (we cover replication in Chapter 6).
2. Each secondary spermatocyte completes meiosis, producing four spermatids, each of which contains 23 chromosomes.
3. Spermatids develop into spermatozoa.
Spermatozoa is just the technical name for what you know as a sperm. To become a full-fledged sperm, the spermatid must mature to the point where it has a tail, middle piece, and a head. The tail is a flagellum that moves the sperm through body fluids — in other words, it’s what allows the sperm to swim. The middle piece contains many mitochondria, which supply the energy for the sperm’s tail to move, and the head contains the cell’s 23 chromosomes inside its nucleus.
Oh, oh, oogenesis: Making eggs
Oogenesis is how human females produce eggs, the female gamete. A human female is born with all the eggs she’ll ever have (which means oogenesis occurs when the female is a developing fetus). These eggs lie dormant from birth until puberty, at which time the female’s hormones kick-start the eggs into the menstrual cycle, which continues monthly from puberty until the woman begins menopause.
The process of oogenesis begins in cells called oogonia (singular: oogonium) that contain all 46 human chromosomes. These cells grow in size and mature into primary oocytes. Here’s a breakdown of the rest of the process:
1. Primary oocytes begin going through meiosis but pause early in the first half of the process.
Each primary oocyte remains paused in meiosis until the female enters puberty.
2. After the hormones of sexual development start flooding the female’s ovaries during puberty, the menstrual cycle begins, triggering ovulation and the completion of meiosis in the primary oocyte.
Ovulation, the release of an egg from an ovary, occurs at the midpoint of the menstrual cycle. To prepare for release and possible fertilization, the joining of egg and sperm, the primary oocytes continue on through meiosis, completing the first meiotic division to produce two unequal cells: a secondary oocyte (also called a daughter cell), which receives most of the cytoplasm (so it’s large), and the first polar body, which has minimal cytoplasm (so it’s tiny). Both the secondary oocyte and the polar body contain 23 replicated chromosomes.
3. The secondary oocyte and first polar body complete the second division of meiosis.
The secondary oocyte undergoes another unequal division of cytoplasm, producing a larger egg and a smaller second polar body. The first polar body divides equally into two polar bodies. The egg and the three polar bodies each contain 23 chromosomes.
The splitting of the cytoplasm, called cytokinesis, is unequal so that the egg ends up full of cytoplasm. The egg gets most of the cytoplasm so it can hold plenty of the nutrients and organelles necessary for a developing embryo. Meiosis in one original oogonium produces just one functional egg. The three polar bodies also produced just wither away.
 In human females, the meiotic division that pauses in the oocyte can remain paused for 40 years or more! From before a woman is born until her ovulation ends during menopause, oocytes are in “hang time,” just waiting to develop into an egg and get fertilized. Of the thousands of oocytes that a female is born with, only about 500 develop into eggs during the woman’s lifetime.
In human females, the meiotic division that pauses in the oocyte can remain paused for 40 years or more! From before a woman is born until her ovulation ends during menopause, oocytes are in “hang time,” just waiting to develop into an egg and get fertilized. Of the thousands of oocytes that a female is born with, only about 500 develop into eggs during the woman’s lifetime.
Mating rituals and other preparations for the big event
Mating in humans can take place whenever a man and a woman are in the mood to do so. Most other animals, however, must follow more rigid reproductive cycles.
Pretend for a moment that you’re a female oyster living in the ocean. Every season you release 60 million eggs into the water (no, we’re not exaggerating). With that many eggs, your mating process is really left to chance. You just have to hope that one of your eggs happens to bump into a male oyster’s sperm so fertilization can occur. Leaving the continuation of the oyster species to chance is one reason why oysters release such huge numbers of eggs in the first place. Obviously, most of the eggs never bump into Mr. Right; if they did, the ocean would be overflowing with oysters, and pearls wouldn’t be so expensive. But what happens if you release all those eggs just because you feel like it and the male oysters in the area aren’t in the mood and don’t release any sperm? Then you’re left alone in your shell with nothing to show for it. Animals have reproductive cycles and specific mating seasons to ensure this type of scenario doesn’t happen.
In the next few sections, we explain how animals decide when it’s time to start looking for a mate and how they attract one. We also fill you in on the details of human reproductive cycles because they play a huge part in the continuation of mankind.
Syncing up for mating season
Animals of the same species need to be in sync to have successful sexual reproduction. In other words, they need to be attempting to mate at the same time.
Most species mate when the conditions are optimal. Often that’s the time of year when the offspring has the best chance for survival. For example, in deer, the gestation period (the length of time the fetus is developing inside a female mammal) is approximately five or six months. The best time for a fawn (a baby deer) to be born is in the spring because food is plentiful, temperatures are a bit warmer, leaves are on the trees, and shrubs can provide cover. A spring birth also gives the fawn the longest period of time to develop before conditions get harsh the next winter. So, backing up six months from spring puts the mating season for deer around October or November. And that’s exactly when you can see bucks competing for does. The strongest buck, which is supposedly the one with the strongest genes, gets to mate and pass on its genetic material to continue the species.
Sometimes, however, mating is triggered by a specific environmental signal. For instance, aquatic (water-living) animals that live in the desert reproduce only when the scarce desert rainfall produces a temporary pond. During the dry season, these animals are in diapause, which is a dormant state. In diapause, the metabolism of the animal is very low, and extreme heat and dry weather don’t affect it. When rain does come, desert-dwelling aquatic animals become active immediately, breed quickly, and have offspring that develop as fast as they can before the pond dries up. Then, the new generation gets to sit in the desert in diapause waiting for the next thunderclouds to appear in the sky.
Attracting a partner
The birds and bees may reproduce sexually, but they don’t fall in love. Nor do they suffer angst over whether the other bee will remain committed or worry about whether their partner will be faithful. Bees “do it” solely for the purpose of creating more bees.
Although love may not be a requirement for sexual reproduction, attraction most certainly is. Animals generally rely on one or both of the following to help them attract a mate:
Mating rituals: Mating rituals are behaviors that animals use to attract a mate. During mating rituals, animals (usually males) “show off” their best features in the hope of proving they’d be good partners. One male may fight other males to show that he’s the strongest; another male may show that he controls access to lots of resources such as food or nesting space. Consider doves. When a male dove is trying to win a mate, he struts around bowing and cooing to the female, trying to win her over. But before they mate, they build a home (sound familiar?). The male and female doves work together to first choose a place for their nest and then build it. During the period of time that they’re building the nest, they take a break and copulate (that’s the formal way of saying they had sex). A few days later, the female lays two eggs in the new nest, and when the chicks hatch, both parents feed them. When the chicks are old enough to start feeding themselves (in about two to three weeks), the adults repeat the reproductive cycle and start courting all over again.
Secondary sex characteristics: These develop as an animal matures, and they’re far less obvious than primary sex characteristics, which are the male and female reproductive organs. Secondary sex characteristics in humans include hair growth and distribution (beards in males), deepening of the voice (in males), increase in muscle mass (males), increase in amount and distribution of fat (in females), and development of breasts (in females). Male deer grow antlers, male lions grow manes, and male peacocks develop a fan of beautiful tail feathers.
Human reproductive cycles
Even though humans are capable of reproducing year-round, reproductive cycles are still involved. Sperm can fertilize an egg any given day of the week, but eggs can only be fertilized a few days out of every month. Human sexual reproduction is therefore controlled by the monthlyovarian cycle (the development of the egg in the ovary) and menstrual cycle (the periodic series of changes associated with menstruation), both of which are controlled by hormones (for more on hormones, see Chapter 18).
As described in the earlier “Oh, oh, oogenesis: Making eggs” section, an oocyte needs to complete meiosis and mature into an egg before it can be released by the ovary. Believe it or not, the brain runs this process.
The hypothalamus, which is found at the center of the brain, keeps a check on how much of the hormones estrogen and progesterone are floating through the bloodstream. When the levels decline, the hypothalamus secretes a hormone called gonadotropin-releasing hormone (GnRH) that heads straight for the pituitary gland (which is also found in the center of the brain) and stimulates part of the pituitary to secrete follicle-stimulating hormone (FSH) and luteinizing hormone (LH). FSH and LH begin the ovarian cycle by kick-starting meiosis again and continuing development of the follicles (another name for the oocytes that are suspended in meiosis) so they can release an egg. As the follicles grow, they release the hormone estrogen.
When the hypothalamus detects rising levels of estrogen in the bloodstream, it releases even more GnRH, causing the pituitary gland to release a large amount of LH about midway through the ovarian cycle. LH stimulates the release of the egg from the follicle in the ovary — in other words, LH triggers ovulation. It also triggers the remaining follicle cells to develop into a mass of cells called the corpus luteum, which secretes estrogen and progesterone for the rest of the ovarian cycle (about two more weeks). These hormones prepare the body for a possible pregnancy by spurring the tissues lining the uterus to develop thicker blood vessels, which brings more nutrients into the uterus.
The hypothalamus can detect when the levels of estrogen and progesterone have reached the point where the lining of the uterus is ready for implantation. If fertilization
Has occurred: The fertilized egg implants in the lining of the uterus and an embryo begins developing. That embryo immediately starts to secrete the hormone human chorionic gonadotropin (hCG). The presence of hCG ensures that estrogen and progesterone production continue so the lining of the uterus remains nourished by larger blood vessels. After the placenta (a blood-filled, nutrient-rich temporary organ) has formed, the embryo gets its nutrients and blood supply through the umbilical cord connecting the embryo to the placenta, which is connected to the mother’s blood supply. Therefore, the production of hCG by the embryo declines after the placenta is up and running.
Pregnancy tests detect the presence of hCG. Because hCG is produced solely by fertilized eggs, only women with a fertilized egg in their bodies should have detectable levels of hCG.
Hasn’t occured: No hCG is produced, and the hypothalamus tells the pituitary gland to stop producing FSH and LH. The lack of FSH and LH stops the production of estrogen and progesterone, which causes the lining of the uterus, sometimes referred to as the endometrium, to stop receiving all that extra nourishment. The endometrium then starts to disintegrate and eventually sloughs off and is carried out of the body by the menstrual flow. The first day of menstrual flow is the first day of the menstrual cycle (pictured in Figure 19-1). The drop in LH also causes the corpeus luteum to disintegrate, ending the ovarian cycle.
The ovarian cycle and the menstrual cycle occur simultaneously and are synchronized to each other by hormones, but each cycle consists of different events.
The ovarian cycle includes the development of the follicle, the secretion of hormones by the follicle, ovulation, and the formation of the corpus luteum. It occurs in the ovary, takes about 28 days to go from beginning to end, and is controlled by GnRH, FSH, LH, and estrogen.
The menstrual cycle includes the thickening of the endometrium to prepare for possible implantation and the shedding of the endometrium if there’s no implanted embryo. It occurs in the uterus, takes about 28 days, and is controlled by the levels of progesterone and estrogen.
Many more eggs are fertilized in a woman’s lifetime than she may realize. Not every fertilized egg results in a bouncing baby boy or girl. If the hormone levels aren’t right from the start, a fertilized egg may never implant or may implant but not secrete enough hormones to maintain the pregnancy. If an embryo doesn’t produce a sufficient amount of hCG, the pregnancy fails to continue, and the embryo aborts itself (a spontaneous abortion is another term for a miscarriage). An unusually heavy menstrual period that started a few days late is often really the spontaneous abortion of a fertilized egg that didn’t work out.
Figure 19-1: The menstrual cycle.
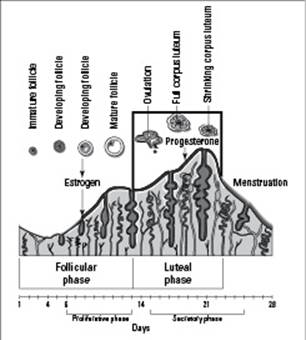
From LifeART®, Super Anatomy 3, © 2002, Lippincott Williams & Wilkins
Talk about overstaying your welcome
After a follicle develops and releases an egg, the empty follicle is called a corpus luteum, which is Latin for “yellow body.” If the egg becomes fertilized and implants in the uterus, the corpus luteum hangs around to help out with the beginning stages of pregnancy. It secretes progesterone for a few weeks until the placenta is fully developed and can secrete progesterone on its own. (The progesterone helps keep the lining of the uterus rich with blood and nutrients for the developing embryo.) Sometimes the corpus luteum sticks around for a few months, but in most cases, it eventually shrinks and withers away during the pregnancy.
Notice how we said “in most cases”? About 10 percent of the time, the corpus luteum hangs out in the ovary far longer than it should. Sometimes it remains even if the woman isn’t pregnant. When that happens, the corpus luteum can turn into a cyst, which is aptly called a corpus luteum cyst. Usually, this cyst on the ovary isn’t a problem, unless it continues to grow, twist, or rupture. Only then does it need to be removed surgically.
How humans mate
The most important thing you should know about mating is that members of different species can’t successfully reproduce with each other. After all, the whole point of sexual reproduction is to create a new generation that contains the genetic information from the previous generations. Interspecies sexual reproduction doesn’t work because different species contain different numbers of chromosomes, and those chromosomes contain differing genes. For instance, humans carry 46 chromosomes in each cell, whereas chimpanzees have 48 per cell. If a human and a chimp were to mate, the cell divisions wouldn’t be equal, and a theoretical offspring (we won’t even go there) probably wouldn’t be able to survive.
Eggs are actually surrounded by a layer of proteins on top of the plasma membrane that contains receptor molecules made solely for receiving sperm of the same species. In human eggs, the zone that prevents fertilization by a different species is called the zona pellucida. Only human sperm can crack the code to get into the egg.
In the following sections, we stick to how humans make babies. (If your parents have yet to have this conversation with you, you can let them know they’re off the hook.)
Human reproductive systems
Before you dive into the details of human sexual reproduction, it helps to know a little bit about the organ systems involved. As you can see in Figure 19-2, the male reproductive system is made up of the penis, the testes, and the seminiferous tubules. Sperm is made in these tubules through the process of spermatogenesis (which we explain earlier in this chapter).
The female reproductive system consists of the vagina, the uterus, the ovaries, and the fallopian tubes (see Figure 19-3). A woman’s egg cells are produced in her ovaries through the process of oogenesis (which we walk you through earlier in this chapter).
Sexual intercourse
In order to prepare the body for sexual intercourse (the insertion of the man’s penis into the woman’s vagina), men and women engage in activities that increase arousal (responsiveness to sensory stimulation). When a man is sexually aroused, his penis becomes erect because the erectile tissue within the penis fills with blood. This erection allows the penis to stiffen so it can remain inside a woman’s vagina during intercourse. In females, the clitoris is the sexually sensitive organ. It has erectile tissue and a glans tip, just like a penis does. When a woman becomes aroused, the erectile tissue within the clitoris (which is located toward the front end of the labia just under the pubic bone) fills with blood, and the increased pressure causes drops of fluid to be squeezed out of the tissue. This lubrication prepares the vagina for sexual intercourse so the erect penis can be inserted into it easily.
When the penis is fully inside the vagina, the tip of the penis is as close as possible to the woman’s cervix. The cervix is the bottom end of the uterus, which extends down into the vagina. Sperm must travel through the cervix to enter the uterus. During sexual intercourse, sperm travel from the man’s epididymides (tubules in the scrotum that store produced sperm) to his vas deferens, tubes that carry sperm from the scrotum to the urethra so they can be ejaculated.
The actions that occur during sexual intercourse serve to bring the man and woman to the climax of stimulation, which is followed shortly by orgasm.
Figure 19-2: The male reproductive system.
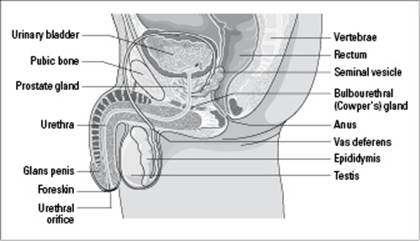
From LifeART®, Super Anatomy 1, © 2002, Lippincott Williams & Wilkins
Figure 19-3: The female reproductive system.
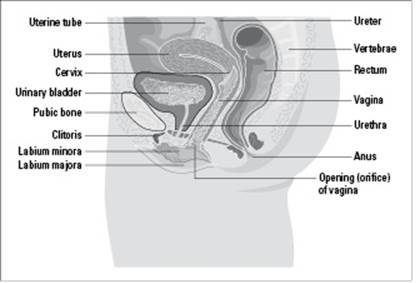
From LifeART®, Super Anatomy 1, © 2002, Lippincott Williams & Wilkins
Orgasm
Believe it or not, orgasm, the highly pleasurable climax of sexual intercourse, serves a physiological purpose. As the sexual stimulation of a male intensifies, the sperm move from the vas deferens into the urethra and secretions from three glands — the seminal vesicles, the prostate gland, and the bulbourethral gland — all add their fluids to create semen (seminal fluid). The semen contains the following “ingredients” that help promote fertilization:
Fructose: This sugar gives the sperm energy to swim upstream.
Prostaglandins: These hormones cause contractions of the uterus that help propel the sperm upward.
A pH of 7.5: This relatively high pH provides the basic solution in which sperm can live and helps neutralize the acidic conditions of the vagina, which would otherwise kill the sperm.
Orgasm occurs at the height of sexual stimulation and is signaled by muscular contractions and a pleasurable feeling of release. The muscular contractions cause semen to be expelled from the penis, which is called ejaculation. The average amount of semen expelled during one ejaculation is less than 1 teaspoon, but it contains more than 400 million sperm.
When orgasm occurs in a male, a sphincter muscle closes off the bladder to prevent urine from entering the urethra. Shutting out urine allows the urethra to be used solely for ejaculation at that time. (In males, the urethra is shared by both the urinary tract and the reproductive tract; in females, the urethra is solely part of the urinary tract.)
In females, the height of sexual stimulation also causes intense muscular contractions and a pleasurable feeling of release. The fluid released inside the vagina helps create a watery environment that the sperm can swim in. The muscular contractions of the uterus slightly open the cervix, which allows sperm to get inside the uterus and also helps “pull” sperm upward toward the fallopian tubes.
Fertilization
After ejaculation, sperm have quite a bit of swimming to do before they find the egg. They have to travel from wherever they’re deposited in the vagina, through the muscular cervix, up through the entire uterus, and finally into the fallopian tubes, which is where fertilization (the joining of sperm and egg) actually occurs.
Because a human egg lives no longer than 24 hours after ovulation and human sperm live no longer than 72 hours, intercourse that occurs in the three-day period prior to ovulation or within the day after ovulation is the only chance of fertilization during a given month.
If the sperm does find its way to the egg, it must penetrate the egg in order to supply it with its 23 chromosomes. However, human eggs have several layers of cells and a thick membrane surrounding them. To get through all that, the sperm produces enzymes in a structure near its nucleus called the acrosome. The acrosomal enzymes digest the protective layers of the egg (so the sperm essentially “eats” its way into the egg). But the sperm isn’t alone in its efforts. The egg helps the sperm get inside by going through physical and biochemical changes. After the sperm has successfully joined with the egg, the two gametes create a cell that contains the full human chromosome count of 46.
How Other Animals Do It
Humans obviously aren’t the only animals that mate and undergo sexual reproduction. All kinds of animals reproduce sexually. The question is: How do they do it?
Following is a look at the mating styles of birds, bees, worms, and sea urchins, as well as explanations of how the species’ zygotes form (a zygote is the fertilized cell that results when two gametes join together). Note: We highlight these animals’ mating styles and zygote formation because they give an overview of the different strategies for sexual reproduction that occur in the animal kingdom.
Bees: The process by which bees reproduce is called parthenogenesis, meaning “virgin production.” (Partheno- is Greek for “virgin” [as in the Parthenon], and genesis means “production.”) A queen bee receives all the sperm she’ll ever be impregnated with during her nuptial flight when she mates with the drones (male bees that never have a full chromosome count). The queen never mates with the drones again because she doesn’t have to — she has stored their sperm cells inside her body, leaving her in total control of when fertilization occurs.
When the queen lays eggs and releases the sperm, the eggs become fertilized. Those fertilized eggs develop into females, many of which are worker bees (these bees are born diploid but never produce gametes, which means they can’t reproduce). The other fertilized eggs develop into new queens. When the queen lays eggs but withholds sperm and prevents fertilization, the unfertilized eggs develop into male drones. The cells in drones’ testes develop into gametes that are received by the new queen, which technically is one of their sisters. (If you’re confused by what haploid and diploid mean, see Chapter 6.)
Birds: Birds copulate, and the male bird deposits his sperm inside the female bird. The egg becomes fertilized and is deposited outside the female bird’s body to continue developing until it’s time to hatch. The yolk of an egg (the yellow part) is where the developing embryo resides, and the albumen (the white part) serves to nourish the embryo throughout its development.
Just after fertilization, one spot on the yolk of an egg goes through a series of divisions called cleavage. At the end of the cleavage divisions, an embryonic disk called a blastoderm is created on one side of the yolk. The blastoderm is the initial cell tissue that begins to develop into a baby bird; it separates into an epiblast, which is the top layer, and a hypoblast, which is the bottom layer. The epiblast cells migrate down into the hypoblast along a line in the yolk, called a primitive streak, to create the mesoderm, which goes on to develop the rest of the bird.
Earthworms: Earthworms are hermaphrodites, meaning they have both female and male reproductive parts (specifically ovaries, testes, seminal vesicles, vas deferens, and seminal receptacles). When earthworms copulate, they face in opposite directions and put their citella together. Citella are the external, smooth, nonsegmented parts of an earthworm; their job is to secrete mucus and help the sperm get from the vas deferens of one worm to the seminal receptacle of the other worm. Cocoons form and are protected by a mucus sheath created by the citella. The sperm and eggs are fertilized inside the cocoon, and the zygotes stay enclosed in it until they hatch in the soil.
Planarian worms: These freshwater flatworms are fairly unique because they can reproduce both asexually (by constricting their bodies and literally splitting in two) and sexually. All planarian worms are hermaphrodites. When two of these worms reproduce sexually, they use their male organs to secrete and exchange sperm with each other. Then they use their female organs to create zygotes. The zygotes develop into small planarian worms that later hatch and mature into adults.
Sea urchins: Male and female sea urchins look exactly the same on the outside — they both have a ring of genital pores at the center of their bodies. Males discharge sperm through their pores into the water, and females discharge eggs through their pores into the water. Fertilization is left to chance, but it’s helped by the fact that sea urchins live in close contact, their eggs have a sticky coat to which sperm adhere, and ejaculation by any one sea urchin signals the other males to ejaculate too.
Developing New Humans
Sexual reproduction involves the production of gametes and the act of mating to join the gametes so fertilization can occur. After fertilization takes place, the term development describes how the fertilized egg becomes another new organism that possesses a mix of its parents’ DNA.
In the sections that follow, we walk you through the process of development, from zygote to newborn. Note: We focus on the development of human offspring because that’s most relevant to your life. However, many similarities exist between the development of humans and other animals.
From single cells to blastocyst
After the nucleus of a sperm and the nucleus of an egg fuse, fertilization is complete, and the new, diploid cell is referred to as a zygote.
The zygote begins traveling down the fallopian tube, heading for the uterus so it can implant in the uterine lining. As it travels, the zygote undergoes cleavage, a rapid series of mitotic divisions that result in a multicellular embryo (see Step 3 in Figure 19-4). The zygote divides into 2 cells, which divide into 4 cells, which divide into 8 cells, which divide into 16 cells. At this point, the zygote is a solid ball of cells called a morula.
Figure 19-4:Fertilization, conception, and early embryonic and fetal development of humans.
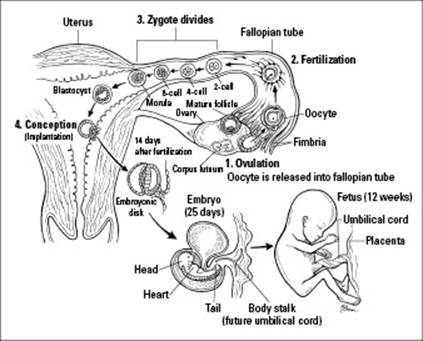
Cell division continues, but the morula becomes filled with liquid, which pushes the increasing number of cells out toward the periphery of the embryo’s membrane, forming a hollow ball of cells called a blastula. In humans, a group of cells inside the blastula become specialized to form the embryo, and the blastula becomes a blastocyst. Different layers of cells become specialized within the blastocyst, taking the first step toward forming specialized tissues:
The flattened cells along the edge of the blastocyst form the trophoblast.
The fluid-filled cavity is called the blastocoel.
The sphere of larger, rounded cells that’s destined to become the embryo is called the embryoblast.
The trophoblast cells secrete an enzyme that helps degrade the lining of the uterus. After the blastocyst “eats” its way into the wall of the uterus, it sinks into the wall and implants itself. Conception occurs when the blastocyst successfully implants itself in the wall of the uterus (see Step 4 in Figure 19-4); if the blastocyst doesn’t implant in the wall of the uterus, there’s no pregnancy. If conception occurs, the trophoblast cells of the blastocyst go on to form the chorion, which becomes part of the placenta.
After implanting itself, the developing mass of cells moves inward, forming a ball of cells called a gastrula that has three layers; this process is referred to as gastrulation. Each layer of cells in the gastrula eventually becomes a different type of tissue:
The outer layer of the gastrula is the ectoderm, which develops into the skin and nervous systems.
The middle layer of the gastrula is the mesoderm, which develops into the muscular, skeletal, and circulatory systems.
The innermost layer is the endoderm, which gives rise to the linings of your digestive and respiratory tracts, as well as organs such as the liver and pancreas.
Conception isn’t the same as fertilization. An egg can be fertilized, but a woman isn’t pregnant until the blastocyst is rooted in her uterine wall where it can develop further.
Go, go, embryo
You get more done in the 12 weeks or so that you’re an embryo than at any other time in your life. That’s because every organ in your body forms during the embryonic period — essentially the first trimester of pregnancy.
After gastrulation (see the preceding section), the specialized cells of the ectoderm, mesoderm, and endoderm begin migrating toward other cells with the same specialty. This cellular migration is referred to as morphogenesis because it gives the embryo a shape (morph- is Greek for form or structure).
Outside the embryo, specialized membranes develop. The chorion combines with tissues created by the mother to become the placenta, an organ that’s filled with blood vessels and provides a large surface area for the exchange of gases, nutrients, and wastes.
A tubular structure called the allantois forms off of the center cavity of the blastocyst. In humans, the allantois eventually becomes the body stalk and then the umbilical cord, which connects the fetus to the placenta (both of which are shown in Figure 19-4).
The amnion surrounds the amniotic cavity, which fills with amniotic fluid that protects the developing embryo. Amniotic fluid cushions movements created by the mother and protects the developing organism from bumps. It’s also thought to contain surfactant, a substance that coats the internal surfaces of the fetus’s lungs so its lung tissues don’t stick together after birth, which would prevent the infant from breathing.
The genetic material in the cells of amniotic fluid matches that of the developing embryo, so doctors can use amniocentesis (a procedure that uses a needle to remove a small amount of amniotic fluid for genetic testing) to see whether the embryo will have a genetic defect.
Throughout embryonic development, all other systems and structures of the body form as well. Cells that split off the neural tube and form the neural crest become the teeth, bones, skin pigments, and muscles of the skull, for example. At the end of the embryonic period, a human embryo is about an inch and a half long, and it starts to look less like a lizard and more like a human.
Fetal development and birth
In humans, the fetal period encompasses the last six months of pregnancy, or the second and third trimesters. Fetuses are completely differentiated, meaning their cells have migrated and formed organ systems. All fetuses do in the uterus is continue to grow and develop features such as hair and nails. As the fetus gets stronger, longer, and heavier, it looks more and more like a newborn baby.
Prostaglandins and the hormone oxytocin cause the uterus to contract, but the initial production of these hormones is thought to be triggered by an as-yet-unknown chemical produced by the fetus. So, the fetus prompts its mother to start producing the hormones that initiate labor. If labor doesn’t begin naturally, medical personnel give the mother prostaglandin suppositories and/or synthetic oxytocin — called pitocin — to induce labor.
When the fetus is finally born, the organism is called a neonate, meaning newborn. A life begins, and development continues. For more information about what happens during pregnancy, check out Pregnancy For Dummies, 3rd Edition (Wiley).
Differentiation, Development, and Determination
We bet you never realized that a large part of who you are is controlled by three D’s: differentiation, development, and determination. Differentiation is the specialization of cells that occurs during development; it determines what the structural and functional aspects of the cell will be.Development is the overall process of an organism going through stages of differentiation; over time, the changes occurring at the cellular level during development become visible. Determination occurs when a cell commits to developing in a certain manner — that is, when it’s determined to become a particular type of cell, such as a brain cell or a bone cell.
Think about this: You started out as one tiny cell. When the nucleus of your father’s sperm fused with the nucleus of your mother’s egg, a single cell was created containing all the genetic information you’d ever have for the rest of your life. As that first cell divided, some of its descendants became determined to be heart cells, skin cells, brain cells, and liver cells. Each of these cell types looks different and behaves differently in the body, but they all have the exact same set of genetic information. What makes each cell type different from the other isn’t what kind of genetic information it has but how it uses that genetic information.
Cells become differentiated through the process of gene regulation, a process that controls which set of genes a particular cell uses at any given time.
 If you think of the different types of cells in your body as workers that each have a different job to do, then it makes sense to think about each worker needing a different set of tools. To a cell, that means a different set of proteins. And to get proteins, cells access the genes in the DNA that contain the blueprints for those proteins. So, your heart cells use some of the genes in your DNA to build the proteins they need for contraction, whereas your skin cells use other genes in your DNA to build the proteins they need to protect you from infection (see Chapter 8 for more on gene regulation).
If you think of the different types of cells in your body as workers that each have a different job to do, then it makes sense to think about each worker needing a different set of tools. To a cell, that means a different set of proteins. And to get proteins, cells access the genes in the DNA that contain the blueprints for those proteins. So, your heart cells use some of the genes in your DNA to build the proteins they need for contraction, whereas your skin cells use other genes in your DNA to build the proteins they need to protect you from infection (see Chapter 8 for more on gene regulation).
In the next few sections, we look at the signals that direct cells to become specialized for certain tasks in the body. We also look at some of the experiments that scientists are doing to try and figure out how to reset the programming of a cell.
The ability to become any type of cell
Initially, each and every cell in your body (and the bodies of many other animals) has totipotency — the ability to develop into any kind of cell or even a whole organism. As the cells differentiate during development, they lose the ability to express all the genes they contain, which means that under normal conditions, a cell that becomes a muscle cell won’t later change and become a skin cell.
Differentiation, then, occurs as a result of signals that cause cells to use only some of the genetic information they contain. Scientists are very interested in understanding these signals and discovering how to reprogram cells so they can be used to heal traumatic injury and disease. Following are just a couple of the experiments scientists have conducted to try and figure out how to reset the programming of a cell:
Two researchers, Robert Briggs and T.J. King, tested tadpole cells to see whether they lost the ability to be reprogrammed and, if so, when they did. They found that up until the blastula stage, at which point the organism contains 8,000 to 16,000 cells, a single tadpole cell retained the capacity to develop into an entire organism. In other words, one cell from a blastula could develop into a whole new tadpole. However, when they used cells from a later period in development, they weren’t successful.
Another researcher, J.B. Gurdon, had success in “growing” normal frog embryos from adult frog skin cells. Gurdon used nuclear transplantation to move the nucleus from an adult frog skin cell into an egg cell from which he’d removed the nucleus. When the nucleus from the differentiated skin cell was placed into the environment of the egg cell cytoplasm, the nucleus directed the growth and development of a frog tadpole that was genetically identical to the frog that had donated the nucleus from its skin cell. As long as the egg into which the cell was transplanted wasn’t past the critical point in development, a tadpole developed.
Gurdon’s experiment was the first successful attempt to clone an organism — produce an organism that’s genetically identical to the organism that donates the transplanted nucleus. Since Gurdon’s experiment, other types of animals have been cloned, the most famous of which is Dolly the sheep.
Animal cloning experiments demonstrate that the nuclei of differentiated cells retain all the genetic information necessary to become other cell types.
Although animals seem to have a point at which cells become determined to differentiate into certain types, the totipotency in many plant cells remains intact. You know how you can grow a whole plant from a cutting of another plant? The cutting doesn’t contain roots, but it’s able to grow them because it can still access the genes necessary for the function of root cells. It’s also possible to take a few plant cells and grow an entire new plant from them.
The factors that affect differentiation and development
When embryonic development begins, all the cells in the embryo are totipotent until signals around the cells make the cells turn into nervous system cells, muscle tissue cells, heart cells, lung cells . . . you get the idea. Most developmental changes depend upon signals in the environment of embryonic cells that tell them exactly what to do and when to do it. Here’s a rundown of those signals:
Embryonic induction: The influence of one group of cells on another group of cells is called embryonic induction, and it causes the recipients of the influence to change their course in development. Cells that wield this power are called organizers, and they exert their influence by secreting certain chemicals or by interacting directly with target cells through cell-to-cell contact. Embryonic induction occurs when the lens of the eye is developing. The eyes start out as bulging outgrowths (or optic vesicles) on the sides of the early brain. When the optic vesicles touch the ectoderm, the ectoderm thickens into the lens placode, which then develops into the curved lens of the eye.
Cell migration: When cells move to new locations during development, that’s considered cell migration. Cells may follow chemical trails to find their new destination. Once there, they attach to similar cells and differentiate to become a particular kind of tissue. In the developing brain, for example, primitive neural cells migrate out of the neural tube to establish the parts of the brain and then begin growing and forming connections with their target cells.
Homeotic genes: Special genes that turn other genes on or off are homeotic genes. They work kind of like a master switch. The proteins produced from homeotic genes interact with DNA, affecting the expression of other genes. When certain genes are turned on, certain proteins are produced that contribute to development. When certain genes are turned off, the protein normally created is withheld so it can’t affect development. These actions control what substances are present or absent in a developing embryo, thereby regulating the development of that embryo. The powerful effect of homeotic genes can be seen in genetic studies of fruit flies. If the homeotic genes in fruits flies are mutated, body parts end up in the wrong places — for example, legs can appear where antennae should be.
Several years ago, a stretch of DNA about 180 nucleotides long (that’s not very big in the DNA world) was found in most of the homeotic genes in many species. This short segment of genes is called a homeobox, and yes, even humans have homeoboxes. A homeobox is the sequence in the homeotic gene that remains unchanged generation after generation. In fact, even after so many generations, similarities exist in the homeoboxes of all living things on Earth.
Programmed cell death: Also called apoptosis, programmed cell death causes cells to commit suicide at particular times during development in order to remove cells from the developing organism. In humans, for example, fingers are created when cells between each finger commit suicide, creating the spaces between the fingers.
Cytoplasmic factors: These can create different local environments in the cytoplasm of a cell, leading to different developmental fates for the descendents of the cell. During cell division early in development, the cytoplasm is often divided unequally among secondary oocytes. So, some cells may get more of certain cytoplasmic factors than other cells. These cytoplasmic factors may cause the cell to turn on certain genes that determine its fate. For instance, differences in the cytoplasm of cells early in development establish the anterior-posterior axis in the embryo, which ultimately leads to the differentiation of the head from the rest of the body.
Hormones: Hormones play a role in development after most of the vital organs (heart, lungs, kidneys, and liver) are formed and limbs and other appendages are where they belong. Then they take control of the actual appearance of the body. The production of testosterone in a male fetus, for example, leads to formation of male primary sex characteristics.
Gender differentiation in humans
Humans are no strangers to the effects of hormones during development. In fact, human males and females are identical organisms until the time sexual differentiation occurs.
In the very early stages of development, human fetuses have two sets of ducts: one for the female reproductive system, and one for the male reproductive system. When both sets of ducts are present, the stage of development is called the indifferent stage (because there’s no difference yet between male and female). Humans remain in this stage until about seven weeks after fertilization (about the end of the second month of pregnancy), which is why an ultrasound done any earlier than this time can’t tell the sex of the developing embryo. (Note: The first ultrasound of a normal pregnancy is usually performed at 16 weeks of gestation, allowing visualization of the sex of a developing fetus.)
The two sets of ducts are the Wolffian ducts, which eventually become the male vas deferens, epididymis (on the testes), and seminal vesicles, and the Müllerian ducts, which eventually become the oviducts, uterus, and vagina.
Inside the cells, the chromosomes determine whether the embryo will develop into a male or a female. Of the 46 human chromosomes, the last pair — the two Chromosomes 23 — are either two X chromosomes or an X and a Y chromosome. Two X chromosomes indicate female; an X and a Y indicate male. If two X chromosomes are inside the cells of the developing reproductive system, the female ducts develop, and the male ducts disintegrate. The reverse happens if an X chromosome and a Y chromosome are in the cells.
The following sections provide the specifics on how primary sex characteristics develop and list some of the ways that can happen incorrectly.
How boys become boys
A gene called SRY (for sex-determining region Y chromosome) is the specific gene that determines maleness. SRY contains the blueprint for a protein called testes determining factor (TDF), which is a transcription factor that interacts with DNA to turn on the transcription of the genes necessary for testes development. (Transcription factors are proteins that turn genes on and off; turn to Chapter 8 to find out more.)
After the testes are formed, they begin to secrete the hormone testosterone (in the form dihydrotestosterone, or DHT). Testosterone supports the development of the male reproductive system and directs development of external genitalia. The tubules necessary for ejaculation of semen are complete at about 14 weeks of gestation (which is the beginning of the second trimester of pregnancy). At about that time, the penis, testes, and scrotum develop from the urogenital tubercle, urogenital swellings, and urogenital folds. The urogenital tubercle becomes the glans penis in the male, the urogenital folds become the shaft of the penis, and the urogenital swellings become the scrotum.
If SRY is absent, the primary gonad develops into ovaries, but two X chromosomes are required for the ovaries to be maintained.
How girls become girls
The absence of DHT in a fetus is what spurs the development of female external genitalia. Without DHT, the urogenital tubercle becomes the clitoris (which is equivalent to the glans penis), the urogenital swellings become the labia majora, and the urogenital folds become the labia minora.
A female’s external genitalia develop even if the internal genitalia fail to develop. External female structures are completed between 14 and 16 weeks of gestation.
Problems with sexual development
The complex process of sexual differentiation involving genes and hormones isn’t without error. Following are the problems that can occur in the hormonal stimulation of genitalia:
Androgen insensitivity: A male who can’t develop external male genitalia has androgen insensitivity. Embryos with an abnormal androgen receptor can’t bind the DHT necessary to produce male genitalia. Therefore, they may be male genetically (XY), but they have female external genitalia.
Hermaphrodites: People with some male and some female characteristics are hermaphrodites. This condition can result from hormonal imbalances. In embryos that oversecrete adrenal androgens (hormones that are involved in the normal synthesis of DHT and testosterone), a genetic female may have masculinized external genitalia complete with a penis but have normal ovaries and other female internal reproductive structures. Or, a genetic male may be undermasculinized.
Klinefelter’s syndrome: Males with Klinefelter’s syndrome have two X chromosomes and one Y chromosome (XXY). They usually have small testes that don’t produce enough testosterone. As a result, male secondary sex characteristics, such as facial hair, may not develop completely, and the male is usually infertile. Males with Klinefelter’s syndrome tend to be taller; they may also have feminizing characteristics such as enlarged breasts. Treatment with hormone therapy can greatly reduce these effects and allow males to develop more normally and have normal sex lives.
Turner syndrome: This genetic disorder, which leaves females infertile, can occur in two ways. First, a genetically female (XX) individual may be missing part or all of one of the X chromosomes, resulting in an XO individual (a person with only one sex chromosome, an X) that’s neither completely female nor male. Second, an embryo may have an X chromosome and a Y chromosome, which normally indicates male, but a deletion occurs in the region of the sex-determining gene on the Y chromosome. This deletion prevents development of testes, so no DHT is produced. Female internal and external genitalia develop, but the ovaries fail prematurely. Women with Turner syndrome are often shorter than other women, may have extra folds of skin around the neck, and may fail to enter puberty. However, recent advances in hormone therapy have significantly reduced these effects.