THE LIVING WORLD
Unit Six. Animal Life
29. The Senses
29.5. Sensing Light: Vision
No other stimulus provides as much detailed information about the environment as light. Vision, the perception of light, is carried out by a special sensory apparatus called an eye. All the sensory receptors described to this point have been chemical or mechanical ones. Eyes contain sensory receptors called rods and cones that respond to photons of light. The light energy is absorbed by pigments in the rods and cones, which respond by triggering nerve impulses in sensory neurons.
Evolution of the Eye
Vision begins with the capture of light energy by photoreceptors. Because light travels in a straight line and arrives virtually instantaneously, visual information can be used to determine both the direction and the distance of an object. No other stimulus provides as much detailed information.
Many invertebrates have simple visual systems with photoreceptors clustered in an eyespot. The flatworm in figure 29.10 has an eyespot, consisting of pigment molecules that are stimulated by light, triggering a nerve impulse in photoreceptor cells. Although an eyespot can perceive the direction of light, it cannot be used to construct a visual image. The members of four phyla—annelids, mollusks, arthropods, and vertebrates—have evolved well-developed, image-forming eyes. True image-forming eyes in these phyla, though they at first seem similar (compare the eyes of an arthropod, mollusk, and vertebrate in figure 29.11), are believed to have evolved independently. Interestingly, the photoreceptors in all of them use the same light-capturing molecule, suggesting that not many alternative molecules are able to play this role.
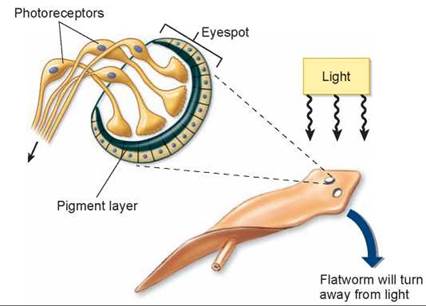
Figure 29.10. Simple eyespots in the flatworm.
Eyespots will detect the direction of light because a pigmented layer on one side of the eyespot screens out light coming from the back of the animal. Light is thus detected more readily coming from the front of the animal; flatworms will respond by turning away from the light.
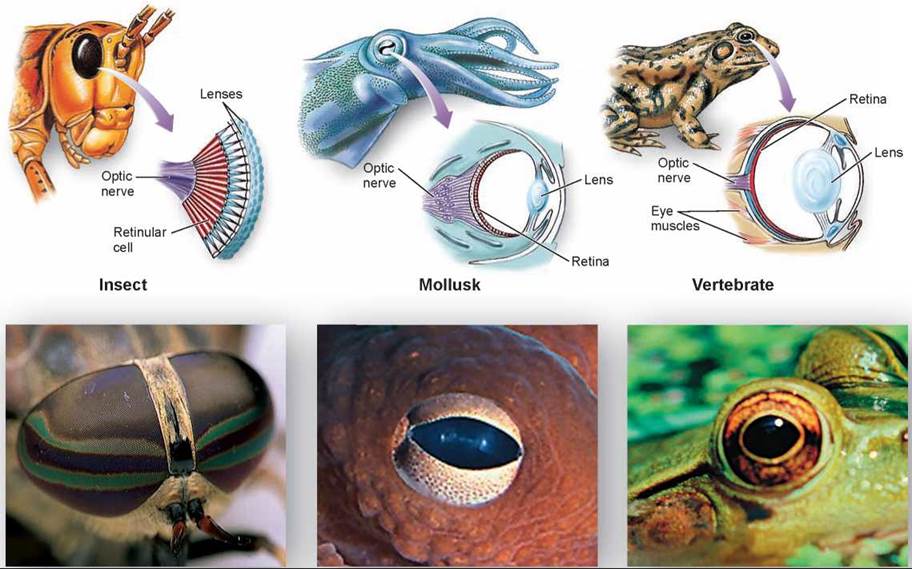
Figure 29.11. Eyes in three phyla of animals.
Although they are superficially similar, these eyes differ greatly in structure and are not homologous. Each has evolved separately and, despite the apparent structural complexity, has done so from simpler structures.
Structure of the Vertebrate Eye
The vertebrate eye works like a lens-focused camera. Light first passes through a transparent protective covering called the cornea (the light blue layer in figure 29.12), which begins to focus the light onto the rear of the eye. The beam of light then passes through the lens, which completes the focusing. The lens is attached by stringlike suspensory ligaments to ciliary muscles. When these muscles contract and relax, they change the shape of the lens and thus allow the eye to view objects that are near or far. The amount of light entering the eye is controlled by a shutter, called the iris (the colored part of your eye), between the cornea and the lens. The transparent zone in the middle of the iris, the pupil, gets larger in dim light and smaller in bright light.
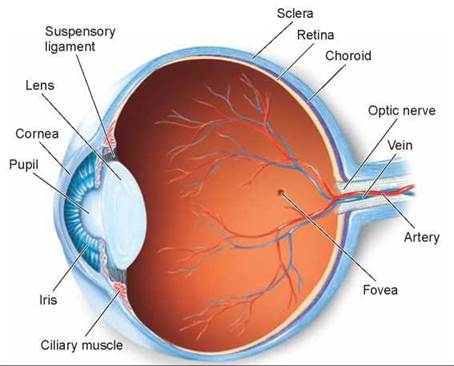
Figure 29.12. The structure of the human eye.
Light passes through the transparent cornea and is focused by the lens on the rear surface of the eye, the retina. The retina is rich in photoreceptors, with a high concentration in an area called the fovea.
The light that passes through the pupil is focused by the lens onto the back of the eye. An array of light-sensitive receptor cells lines the back surface of the eye, called the retina. The retina is the light-sensing portion of the eye. The vertebrate retina contains two kinds of photoreceptors, called rods and cones, which, when stimulated by light, generate nerve impulses that travel to the brain along a short, thick nerve pathway called the optic nerve. Rods, the taller, flat-topped cell in figure 29.13, are receptor cells that are extremely sensitive to light, and they can detect various shades of gray even in dim light. However, they cannot distinguish colors, and because they do not detect edges well, they produce poorly defined images. Cones, the pointed-topped cells, are receptor cells that detect color and are sensitive to edges so that they produce sharp images. The center of the vertebrate retina contains a tiny pit, called the fovea, densely packed with some 3 million cones. This area produces the sharpest image, which is why we tend to move our eyes so that the image of an object we want to see clearly falls on this area.
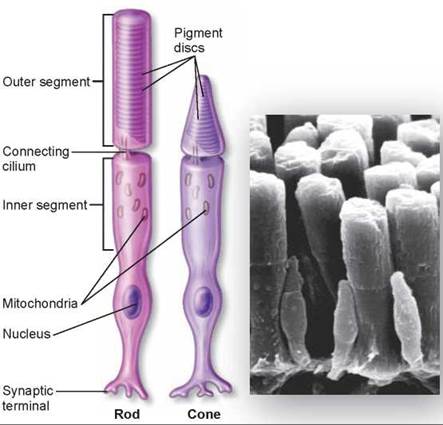
Figure 29.13. Rods and cones.
The broad tubular cell on the left is a rod. The shorter, tapered cell next to it is a cone. The electron micrograph of human rods and cones illustrates that rods are typically larger.
The lens of the vertebrate eye is constructed to filter out short-wavelength light. This solves a difficult optical problem: Any uniform lens bends short wavelengths more than it does longer ones, a phenomenon known as chromatic aberration. Consequently, these short wavelengths cannot be brought into focus simultaneously with longer wavelengths. Unable to focus the short wavelengths, the vertebrate eye eliminates them. Insects, whose eyes do not focus light, are able to see these lower, ultraviolet wavelengths quite well and often use them to locate food or mates.
A rod or cone cell in the eye is able to detect a single photon of light. How can it be so sensitive? The primary sensing event of vision is the absorption of a photon of light by a pigment. The pigments in rods and cones are made from plant pigments called carotenoids. That is why eating carrots is said to be good for night vision—the orange color of carrots is due to the presence of carotenoids called carotenes. The visual pigment in the human eye is a fragment of carotene called c/s-retinal. The pigment is attached to a protein called opsin to form a light-detecting complex called rhodopsin.
When it receives a photon of light, the pigment undergoes a change in shape. This change in shape must be large enough to alter the shape of the opsin protein attached to it. When light is absorbed by the cis-retinal pigment (the upper molecule in figure 29.14), the linear end of the molecule rotates sharply upward, straightening out that end of the molecule. The new form of the pigment is referred to as trans-retinal and the dashed outline in the figure shows the shape before it was stimulated by light. This radical change in the pigment’s shape induces a change in the shape of the protein opsin to which the pigment is bound, initiating a chain of events that leads to the generation of a nerve impulse.
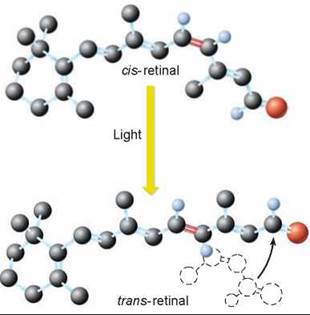
Figure 29.14. Absorption of light.
When light is absorbed by cis- retinal, the pigment undergoes a change in shape and becomes trans-retinal.
Each rhodopsin activates several hundred molecules of a protein called transducin. Each of these activates several hundred molecules of an enzyme whose product stimulates sodium channels in the photoreceptor membrane at a rate of about 1,000 per second. This cascade of events allows a single photon to have a large effect on the receptor.
Color Vision
Three kinds of cone cells provide us with color vision. Each possesses a different version of the opsin protein (that is, one with a distinctive amino acid sequence and thus a different shape). These differences in shape affect the flexibility of the attached retinal pigment, shifting the wavelength at which it absorbs light. The absorption spectrum in figure 29.15 shows the wavelength of light that is absorbed by each cone and rod cell. In rods, light is absorbed at 500 nanometers. In cones, the three versions of opsin absorb light at 420 nanometers (blue-absorbing), 530 nanometers (green-absorbing), or 560 nanometers (red-absorbing). By comparing the relative intensities of the signals from the three cones, the brain can calculate the intensity of other colors.
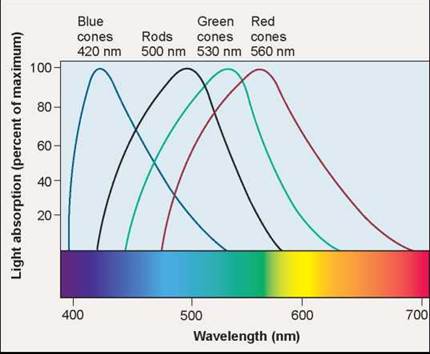
Figure 29.15. Color vision.
The absorption spectrum of cis-retinal is shifted in cone cells from the 500 nanometers characteristic of rod cells. The amount of the shift determines what color the cone absorbs: 420 nanometers yields blue absorption; 530 nanometers yields green absorption; and 560 nanometers yields red absorption. Red cones do not peak in the red part of the spectrum, but they are the only cones that absorb red light.
Some people are not able to see all three colors, a condition referred to as color blindness. Color blindness is typically due to an inherited lack of one or more types of cones. People with normal vision have all three types of cones. People with only two types of cones lack the ability to detect the third color. For example, people with red-green color blindness lack red cones and have difficulty distinguishing red from green (figure 29.16). Color blindness is a sex-linked trait (see chapter 10), and so men are far more likely to be color blind than women.

Figure 29.16. Test for color blindness.
People with normal color vision see the number 16, but people that are red-green color blind see just spots and no discernible number.
Source: This image has been reproduced from Ishihara's Tests for Color Deficiency published by KANEHARA TRADING INC., located in Tokyo, Japan. But tests for color deficiency cannot be conducted with this material. For accurate testing, the original plates should be used.
Most vertebrates, particularly those that are diurnal (active during the day), have color vision, as do many insects. Indeed, honeybees can see light in the nearultraviolet range, which is invisible to the human eye. Color vision requires the presence of more than one photopigment in different receptor cells, but not all animals with color vision have the three-cone system characteristic of humans and other primates. Fish, turtles, and birds, for example, have four or five kinds of cones; the “extra” cones enable these animals to see near-ultraviolet light. Many mammals such as squirrels have only two types of cones.
Conveying the Light Information to the Brain
The path of light through each eye is the reverse of what you might expect. The rods and cones are at the rear of the retina, not the front. If you track the path that light would take in figure 29.17, you will see that light passes through several layers of ganglion and bipolar cells before it reaches the rods and cones. Once the photoreceptors are activated, they stimulate bipolar cells, which in turn stimulate ganglion cells. The direction of nerve impulses in the retina is thus opposite to the direction of light.
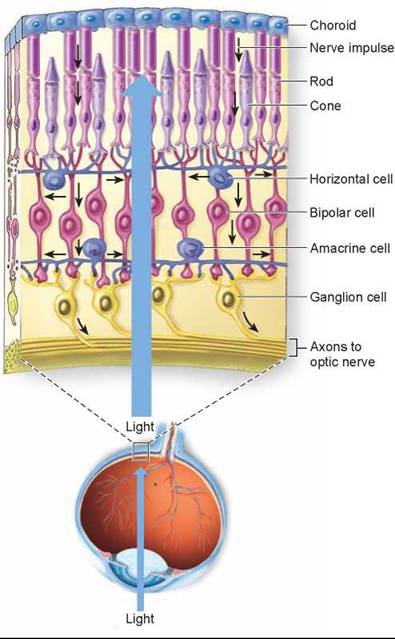
Figure 29.17. Structure of the retina.
The rods and cones are at the rear of the retina. Light passes over four other types of cells in the retina before it reaches the rods and cones. The black arrows indicate how nerve impulses travel through the bipolar cells to the ganglion cells and on to the optic nerve.
Action potentials propagated along the axons of ganglion cells are relayed through structures called the lateral geniculate nuclei of the thalamus and projected to the occipital lobe of the cerebral cortex. There the brain interprets this information as light in a specific region of the eye’s receptive field. The pattern of activity among the ganglion cells across the retina encodes a point-to-point map of the receptive field, allowing the retina and brain to image objects in visual space. In addition, the frequency of impulses in each ganglion cell provides information about the light intensity at each point, while the relative activity of ganglion cells connected (through bipolar cells) with the three types of cones provides color information.
Binocular Vision
Primates (including humans) and most predators have two eyes, one located on each side of the face. When both eyes are trained on the same object, the image that each sees is slightly different because each eye views the object from a different angle. This slight displacement of the images permits binocular vision, the ability to perceive three-dimensional images and to sense depth or the distance to an object. Having their eyes facing forward maximizes the field of overlap in which this stereoscopic vision occurs, as seen by the overlapping blue triangles in the human in figure 29.18. The triangles are the field of view for each eye.

Figure 29.18. Binocular vision.
When the eyes are located on the sides of the head (as on the left), the two vision fields do not overlap and binocular vision does not occur. When both eyes are located toward the front of the head (as on the right) so that the two fields of vision overlap, depth can be perceived.
In contrast, prey animals generally have eyes located to the sides of the head, preventing binocular vision but enlarging the overall receptive field. Depth perception is less important to prey than detection of potential enemies from any angle. The eyes of the American woodcock, for example, are located at exactly opposite sides of its skull so that it has a 360-degree field of view without turning its head! Most birds have laterally placed eyes and, as an adaptation, have two foveas in each retina. One fovea provides sharp frontal vision, like the single fovea in the retina of mammals, and the other fovea provides sharper lateral vision.
Key Learning Outcome 29.5. Vision receptors detect reflected light; binocular vision allows the brain to form three-dimensional images of objects.