THE LIVING WORLD
Unit Three. The Continuity of Life
8.4. Cell Division
Interphase
When cell division begins in interphase, chromosomes first replicate, and then begin to wind up tightly, a process called condensation. Sister chromatids are held together by a complex of proteins called cohesin. Chromosomes are not usually visible during interphase, but to clarify what is happening, they are shown in panel 1 of figure 8.5 as if they were.
Mitosis
Interphase is not a phase of mitosis, but it sets the stage for cell division. It is followed by nuclear division, called mitosis. Although the process of mitosis is continuous, with the stages flowing smoothly one into another, for ease of study, mitosis is traditionally subdivided into four stages: prophase, metaphase, anaphase, and telophase. We will be referring to the panels in figure 8.5 in the following descriptions.
Prophase: Mitosis Begins. In prophase, the individual condensed chromosomes, the blue structures in the photo of panel 2, first become visible with a light microscope. As the replicated chromosomes condense, the nucleolus disappears and the cell dismantles the nuclear envelope and begins to assemble the apparatus it will use to pull the replicated sister chromatids to opposite ends (“poles”) of the cell. In the center of an animal cell, the centrioles have replicated, and the two pairs of centrioles move apart toward opposite poles of the cell, forming between them as they move apart a network of protein cables called the spindle. In panel 2, the centrioles are positioned at the poles; the red structures in the drawing and photo are the protein cables that make up the spindle. Each cable is called a spindle fiber and is made of microtubules, which are long, hollow tubes of protein. Plant cells lack centrioles and instead brace the ends of the spindle toward the poles.
As condensation of the chromosomes continues, a second group of microtubules extends out from the poles toward the centromeres of the chromosomes. Each set of microtubules continues to grow longer until it makes contact with a disk of protein, called a kinetochore, associated with each side of the centromere. When the process is complete, one sister chromatid of each pair is attached by microtubules to one pole and the other sister chromatid to the other pole.
Metaphase: Alignment of the Chromosomes. The second phase of mitosis, metaphase, begins when the chromosomes, each consisting of a pair of sister chromatids, align in the center of the cell along an imaginary plane that divides the cell in half, referred to as the equatorial plane. Panel 3 shows the chromosomes beginning to align along the equatorial plane. Microtubules attached to the kinetochores of the centromeres are fully extended back toward the opposite poles of the cell.
Anaphase: Separation of the Chromatids. In anaphase, enzymes cleave the cohesin link holding sister chromatids together, the kinetochores split, and the sister chromatids are freed from each other. Cell division is now simply a matter of reeling in the microtubules, dragging to the poles the sister chromatids, now referred to as daughter chromosomes. In panel 4 you see the daughter chromosomes being pulled by their centromeres, the arms of the chromosomes dangling behind. The ends of the microtubules are dismantled, one bit after another, making the tubes shorter and shorter and so drawing the chromosome attached to the far end closer and closer to the opposite poles of the cell. When they finally arrive, each pole has one complete set of chromosomes.
Telophase: Re-formation of the Nuclei. The only tasks that remain in telophase are the dismantling of the stage and the removal of the props. The mitotic spindle is disassembled, and a nuclear envelope forms around each set of chromosomes while they begin to uncoil, as shown in panel 5, and the nucleolus reappears.
Cytokinesis
At the end of telophase, mitosis is complete. The cell has divided its replicated chromosomes into two nuclei, which are positioned at opposite ends of the cell. Mitosis is also referred to as karyokinesis. You may recall from chapter 4 that the nucleus is also referred to as karyon (Latin for “kernel”); therefore, karyokinesis is the division of the nucleus. Toward the end of mitosis, cytokinesis, the division of the cytoplasm, occurs, and the cell is cleaved into roughly equal halves. Cytoplasmic organelles have already been replicated and resorted to the areas that will separate and become the daughter cells. Cytokinesis, shown in panel 6 of figure 8.5, signals the end of cell division.
In animal cells, which lack cell walls, cytokinesis is achieved by pinching the cell in two with a contracting belt of actin filaments. As contraction proceeds, a cleavage furrow becomes evident around the cell’s circumference, where the cytoplasm is being progressively pinched inward by the decreasing diameter of the actin belt. In figure 8.6a you see an animal cell pinching in half during cytokinesis. Imagine the cleavage furrow deepening further, until the cell is literally pinched in two.
Plant cells have rigid walls that are far too strong to be deformed by actin filament contraction. A different approach to cytokinesis has therefore evolved in plants. Plant cells assemble membrane components in their interior, at right angles to the mitotic spindle. In figure 8.6b, you can see how membrane is deposited between the daughter cells by vesicles that fuse together. This expanding partition, called a cell plate, grows outward until it reaches the interior surface of the plasma membrane and fuses with it, at which point it has effectively divided the cell in two. Cellulose is then laid down over the new membranes, forming the cell walls of the two new cells.
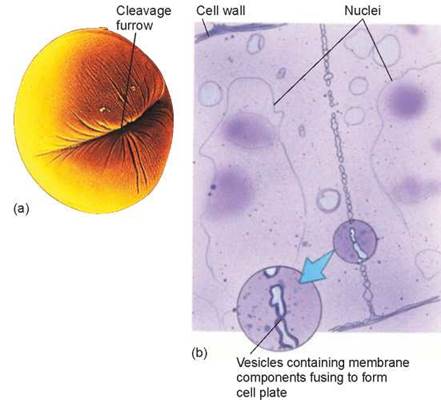
Figure 8.6. Cytokinesis.
The division of cytoplasm that occurs after mitosis is called cytokinesis and cleaves the cell into roughly equal halves. (a) In an animal cell, such as this sea urchin egg, a cleavage furrow forms around the dividing cell. (b) In this dividing plant cell, a cell plate is forming between the two newly forming daughter cells.
Cell Death
Despite the ability to divide, no cell lives forever. The ravages of living slowly tear away at a cell’s machinery. To some degree, damaged parts can be replaced, but no replacement process is perfect. And sometimes the environment intervenes. If food supplies are cut off, for example, animal cells cannot obtain the energy necessary to maintain their lysosome membranes. The cells die, digested from within by their own enzymes.
During fetal development, many cells are programmed to die. In human embryos, hands and feet appear first as “paddles,” but the skin cells between bones die on schedule to form the separated toes and fingers. Figure 8.7 shows a developing human hand looking like a paddle. The cells in the tissue between the bones will later die, leaving behind a set of fingers. In ducks, this cell death is not part of the developmental program, which is why ducks have webbed feet and you don’t.
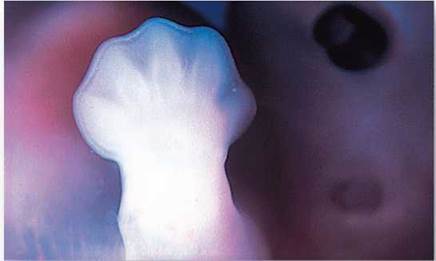
Figure 8.7. Programmed cell death.
In the human embryo, programmed cell death results in the formation of fingers and toes from paddlelike hands and feet.
Human cells appear to be programmed to undergo only so many cell divisions and then die, following a plan written into the genes. In tissue culture, cell lines divide about 50 times, and then the entire population of cells dies off. Even if some of the cells are frozen for years, when they are thawed they simply resume where they left off and die on schedule.
Only cancer cells appear to thwart these instructions, dividing endlessly. All other cells in your body contain a hidden clock that keeps time by counting cell divisions, and when the alarm goes off the cells die.
Key Learning Outcome 8.4. The eukaryotic cell cycle starts in interphase with the condensation of replicated chromosomes; in mitosis, these chromosomes are drawn by microtubules to opposite ends of the cell; in cytokinesis, the cell is split into two daughter cells.