THE LIVING WORLD
Unit Three. The Continuity of Life
10. Foundations of Genetics
10.7. Chromosomes Are the Vehicles of Mendelian Inheritance
The Chromosomal Theory of Inheritance
In the early twentieth century it was by no means obvious that chromosomes were the vehicles of hereditary information. A central role for chromosomes in heredity was first suggested in 1900 by the German geneticist Karl Correns, in one of the papers announcing the rediscovery of Mendel’s work. Soon observations that similar chromosomes paired with one another during meiosis led to the chromosomal theory of inheritance, first formulated by American Walter Sutton in 1902.
Several pieces of evidence supported Sutton’s theory. One was that reproduction involves the initial union of only two cells, egg and sperm. If Mendel’s model was correct, then these two gametes must make equal hereditary contributions. Sperm, however, contain little cytoplasm, suggesting that the hereditary material must reside within the nuclei of the gametes. Furthermore, while diploid individuals have two copies of each pair of homologous chromosomes, gametes have only one. This observation was consistent with Mendel’s model, in which diploid individuals have two copies of each heritable gene and gametes have one. Finally, chromosomes segregate during meiosis, and each pair of homologues orients on the metaphase plate independently of every other pair. Segregation and independent assortment were two characteristics of the genes in Mendel’s model.
Problems with the Chromosomal Theory
Investigators soon pointed out one problem with this theory, however. If Mendelian traits are determined by genes located on the chromosomes, and if the independent assortment of Mendelian traits reflects the independent assortment of chromosomes in meiosis, why does the number of traits that assort independently in a given kind of organism often greatly exceed the number of chromosome pairs the organism possesses? This seemed a fatal objection, and it led many early researchers to have serious reservations about Sutton’s theory.
Morgan's White-Eyed Fly
The essential correctness of the chromosomal theory of heredity was demonstrated by a single small fly. In 1910 Thomas Hunt Morgan, studying the fruit fly Drosophila melanogaster, detected a mutant male fly that differed strikingly from normal fruit flies: Its eyes were white instead of red (figure 10.20).
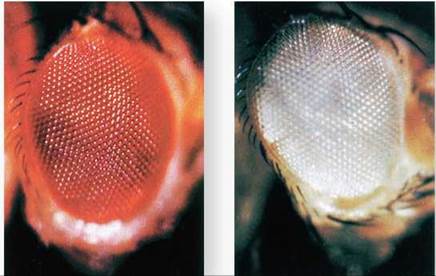
Figure 10.20. Red-eyed (wild type) and white-eyed (mutant) Drosophila.
The white-eye defect is hereditary, the result of a mutation in a gene located on the X chromosome. By studying this mutation, Morgan first demonstrated that genes are on chromosomes.
Morgan immediately set out to determine if this new trait would be inherited in a Mendelian fashion. He first crossed the mutant male with a normal female to see if red or white eyes were dominant. All of the F1 progeny had red eyes, so Morgan concluded that red eye color was dominant over white. Following the experimental procedure that Mendel had established long ago, Morgan then crossed the red-eyed flies from the F1 generation with each other. Of the 4,252 F2 progeny Morgan examined, 782 (18%) had white eyes. Although the ratio of red eyes to white eyes in the F2 progeny was greater than 3:1, the results of the cross nevertheless provided clear evidence that eye color segregates. However, there was something about the outcome that was strange and totally unpredicted by Mendel’s theory—all of the white-eyed F2 flies were males!
How could this result be explained? Perhaps it was impossible for a white-eyed female fly to exist; such individuals might not be viable for some unknown reason. To test this idea, Morgan testcrossed the female F1 progeny with the original white-eyed male. He obtained white-eyed and red-eyed males and females in a 1:1:1:1 ratio, just as Mendelian theory predicted. Hence, a female could have white eyes. Why, then, were there no white-eyed females among the progeny of the original cross?
Sex Linkage Confirms the Chromosomal Theory
The solution to this puzzle involved sex. In Drosophila, the sex of an individual is determined by the number of copies of a particular chromosome, the X chromosome, that an individual possesses. A fly with two X chromosomes is a female, and a fly with only one X chromosome is a male. In males, the single X chromosome pairs in meiosis with a large, dissimilar partner called the Y chromosome. The female thus produces only X gametes, while the male produces both X and Y gametes. When fertilization involves an X sperm, the result is an XX zygote, which develops into a female; when fertilization involves a Y sperm, the result is an XY zygote, which develops into a male.
The solution to Morgan’s puzzle is that the gene causing the white-eye trait in Drosophila resides only on the X chromosome—it is absent from the Y chromosome. (We now know that the Y chromosome in flies carries almost no functional genes.) A trait determined by a gene on the sex chromosome is said to be sex-linked. Knowing the white-eye trait is recessive to the red-eye trait, we can now see that Morgan’s result was a natural consequence of the Mendelian assortment of chromosomes. Figure 10.21 steps you through Morgan’s experiment, showing both the eye color alleles and the sex chromosomes. In this experiment, the Fj generation all had red eyes, while the F2 generation contained flies with white eyes—but they were all males. This at-first-surprising result happens because the segregation of the white-eye trait has a one-to-one correspondence with the segregation of the X chromosome. In other words, the white-eye gene is on the X chromosome. In humans, traits such as color-blindness (see page 208 and chapter 29) and hemophilia (a bloodclotting disease discussed later in this chapter) are sex-linked.
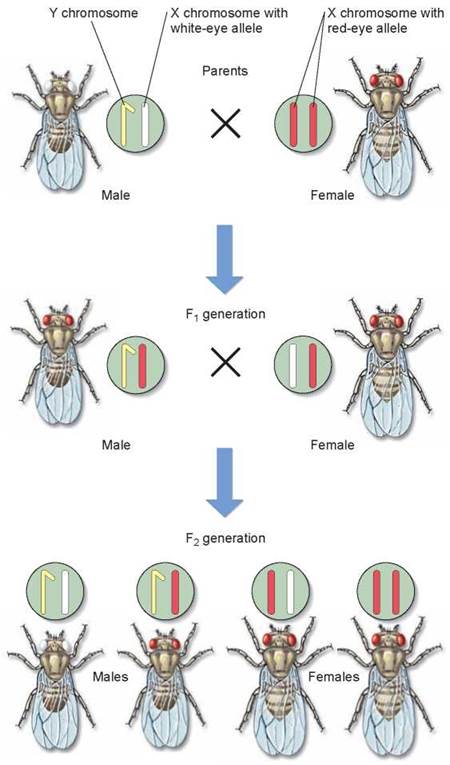
Figure 10.21. Morgan's experiment demonstrating the chromosomal basis of sex linkage.
The white-eyed mutant male fly was crossed with a normal female. The F1 generation flies all exhibited red eyes, as expected for flies heterozygous for a recessive white-eye allele. In the F2 generation, all of the white-eyed flies were male.
Morgan’s experiment presented the first clear evidence that the genes determining Mendelian traits reside on chromosomes, just as Sutton had proposed. Now we can see that the reason Mendelian traits assort independently is because chromosomes assort independently. When Mendel observed the segregation of alternative traits in pea plants, he was observing a reflection of the meiotic segregation of the chromosomes, which contained the characters he was observing.
If genes are located on chromosomes, you might expect that two genes on the same chromosome would segregate together. However, if the two genes are located far from each other on the chromosome, like genes A and I in figure 10.22, the likelihood of crossing over occurring between them is very high, leading to independent segregation. Conversely, the closer two genes are to each other on a chromosome, like genes I and T, the less likely it is that a cross-over event will occur between them. Genes that are located quite close to each other almost always segregate together, meaning that they are inherited together. The tendency of close-together genes to segregate together is called linkage.
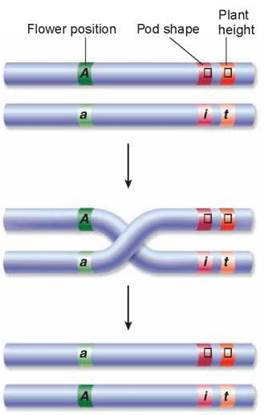
Figure 10.22. Linkage.
Genes that are located farther apart on a chromosome, like the genes for flower position (A) and pod shape (I) in Mendel's peas, will assort independently because crossing over results in recombination of these alleles. Pod shape (I) and plant height (T), however, are positioned very near each other, such that crossing over usually would not occur. These genes are said to be linked and do not undergo independent assortment.
Key Learning Outcome 10.7. Mendelian traits assort independently because they are determined by genes located on chromosomes that assort independently in meiosis.