MCAT Biology Review
Chapter 1: The Cell
1.2 Eukaryotic Cells
The first major distinction we can make between living organisms is whether they are composed of prokaryotic or eukaryotic cells. Eukaryotic organisms can be unicellular or multicellular. Whereas eukaryotic cells contain a true nucleus enclosed in a membrane, prokaryotic cells do not contain a nucleus. The major organelles are identified in the eukaryotic cell in Figure 1.1.
 Figure 1.1. Eukaryotic Cell Numerous membrane-bound organelles are found in the cytoplasm of a eukaryotic cell.
Figure 1.1. Eukaryotic Cell Numerous membrane-bound organelles are found in the cytoplasm of a eukaryotic cell.
MEMBRANE-BOUND ORGANELLES
Each cell has a cell membrane enclosing a semifluid cytosol in which the organelles are suspended. In eukaryotic cells, most organelles are membrane bound, allowing for compartmentalization of functions. Membranes of eukaryotic cells consist of a phospholipid bilayer. This membrane is unique in that its surfaces are hydrophilic, electrostatically interacting with the aqueous environments inside and outside of the cell, while its inner portion is hydrophobic, which helps to provide a highly selective barrier between the interior of the cell and the external environment. The cell membrane is such an important topic on the MCAT that an entire chapter—Chapter 8 of MCAT Biochemistry Review—is devoted solely to discussing the structure and physiology of biological membranes. The cytosol allows for the diffusion of molecules throughout the cell. Within thenucleus, genetic material is encoded in deoxyribonucleic acid (DNA), which is organized into chromosomes. Eukaryotic cells reproduce by mitosis, allowing for the formation of two identical daughter cells.
The Nucleus
As the control center of the cell, the nucleus is the most heavily tested organelle on the MCAT. It contains all of the genetic material necessary for replication of the cell. The nucleus is surrounded by the nuclear membrane or envelope, a double membrane that maintains a nuclear environment separate and distinct from the cytoplasm. Nuclear pores in the nuclear membrane allow for selective two-way exchange of material between the cytoplasm and the nucleus.
BRIDGE
The nuclear envelope creates two distinct environments within the cell because it separates the nucleus from the cytoplasm. This allows for compartmentalization of transcription (the formation of hnRNA from DNA, which is subsequently processed to form mRNA) and translation (the formation of a peptide from mRNA). These processes are discussed in Chapter 7 of MCAT Biochemistry Review.
The genetic material (DNA) contains coding regions called genes. Linear DNA is wound around organizing proteins known as histones, and is then further wound into linear strands called chromosomes. The location of DNA in the nucleus allows for the compartmentalization of DNA transcription separate from RNA translation. Finally, there is a subsection of the nucleus known as the nucleolus, where the ribosomal RNA (rRNA) is synthesized. The nucleolus actually takes up approximately 25 percent of the volume of the entire nucleus and can often be identified as a darker spot in the nucleus.
Mitochondria
Mitochondria, shown in Figure 1.2, are often called the power plants of the cell, in reference to their important metabolic functions. The mitochondrion contains two layers: the outer and inner membranes. The outer membrane serves as a barrier between the cytosol and the inner environment of the mitochondrion. The inner membrane, which is thrown into numerous infoldings called cristae, contains the molecules and enzymes necessary for the electron transport chain. The cristae are highly convoluted structures that increase the surface area available for electron transport chain enzymes. The space between the inner and outer membranes is called the intermembrane space; the space inside the inner membrane is called the mitochondrial matrix. As described in Chapter 10 of MCAT Biochemistry Review, the pumping of protons from the mitochondrial matrix to the intermembrane space establishes the proton-motive force; ultimately, these protons flow through ATP synthase to generate ATP during oxidative phosphorylation.
REAL WORLD
The serial endosymbiosis theory attempts to explain the formation of some of the membrane-bound organelles; it posits that these organelles formed by the engulfing of one prokaryote by another and the establishment of a symbiotic relationship between the two. In addition to mitochondria, chloroplasts in plant cells and organelles of motility (such as flagella) are believed to have evolved through this process.
Mitochondria are different from other parts of the cell in that they are semi-autonomous. They contain some of their own genes and replicate independently of the nucleus via binary fission. Mitochondria are thought to have evolved from an anaerobic prokaryote engulfing an aerobic prokaryote and establishing a symbiotic relationship.
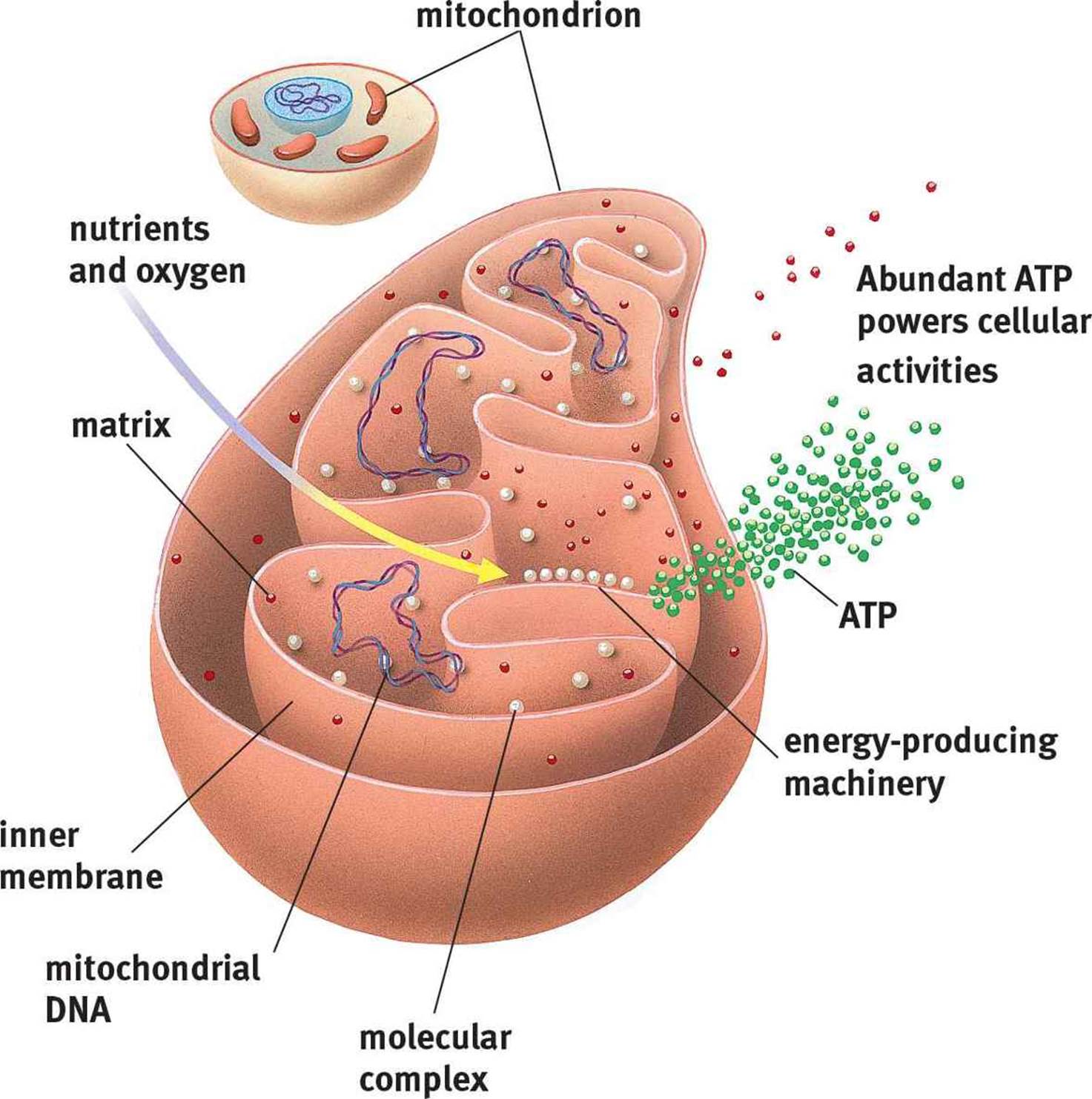 Figure 1.2. Mitochondrial Structure
Figure 1.2. Mitochondrial Structure
In addition to keeping the cell alive by providing energy, the mitochondria are also capable of killing the cell by release of enzymes from the electron transport chain. This release kick-starts a process known as apoptosis, or programmed cell death.
Lysosomes
Lysosomes are membrane-bound structures containing hydrolytic enzymes that are capable of breaking down many different substrates, including substances ingested by endocytosis and cellular waste products. The lysosomal membrane sequesters these enzymes to prevent damage to the cell. However, release of these enzymes can occur in a process known as autolysis. Like mitochondria, when lysosomes release their hydrolytic enzymes, it results in apoptosis. In this case, the released enzymes directly lead to the degradation of cellular components.
Endoplasmic Reticulum
The endoplasmic reticulum (ER) is a series of interconnected membranes that are actually contiguous with the nuclear envelope. The single membrane of the endoplasmic reticulum is folded into numerous invaginations, creating complex structures with a central lumen. There are two varieties of ER: smooth and rough. The rough ER (RER) is studded with ribosomes, which permit the translation of proteins destined for secretion directly into its lumen. On the other hand, the smooth ER (SER) lacks ribosomes and is utilized primarily for lipid synthesis and the detoxification of certain drugs and poisons. The SER also transports proteins from the RER to the Golgi apparatus.
Golgi Apparatus
The Golgi apparatus consists of stacked membrane-bound sacs. Materials from the ER are transferred to the Golgi apparatus in vesicles. Once in the Golgi apparatus, these cellular products may be modified by the addition of various groups, including carbohydrates, phosphates, and sulfates. The Golgi apparatus may also modify cellular products through the introduction of signal sequences, which direct the delivery of the product to a specific cellular location. After modification and sorting in the Golgi apparatus, cellular products are repackaged in vesicles, which are subsequently transferred to the correct cellular location. If the product is destined for secretion, then the secretory vesicle merges with the cell membrane and its contents are released via exocytosis. The relationships between lysosomes, the ER, and the Golgi apparatus are shown in Figure 1.3.
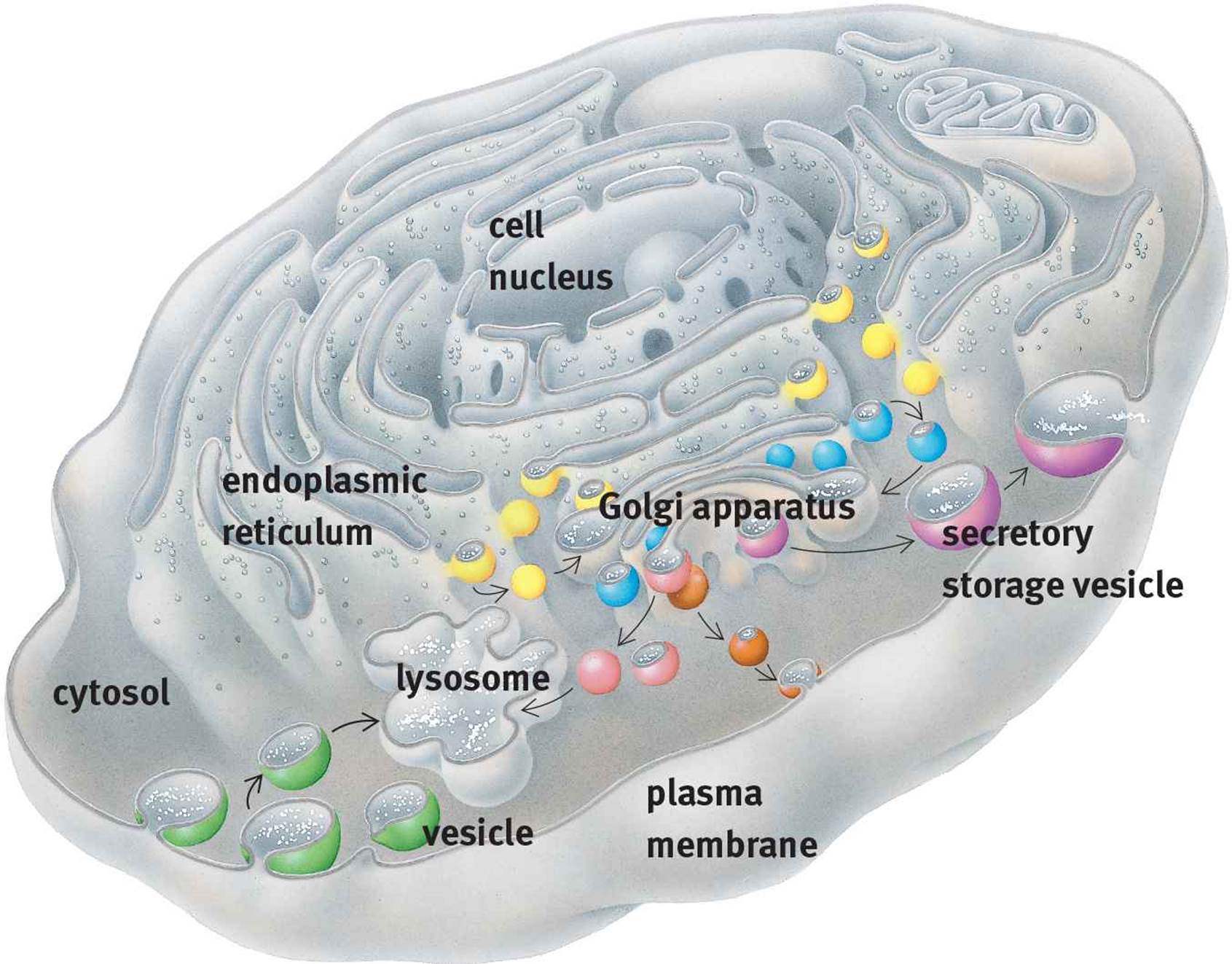 Figure 1.3. Lysosomes, the Endoplasmic Reticulum, and the Golgi Apparatus
Figure 1.3. Lysosomes, the Endoplasmic Reticulum, and the Golgi Apparatus
KEY CONCEPT
Not all cells have the same relative distribution of organelles. Form will follow function. Cells that require a lot of energy for locomotion (such as sperm cells) have high concentrations of mitochondria. Cells involved in secretion (such as pancreatic islet cells and other endocrine tissues) have high concentrations of RER and Golgi apparatuses. Other cells, such as red blood cells, which primarily serve a transport function, have no organelles at all.
Peroxisomes
Peroxisomes contain hydrogen peroxide. One of the primary functions of peroxisomes is the breakdown of very long chain fatty acids via β-oxidation. Peroxisomes participate in the synthesis of phospholipids and contain some of the enzymes involved in the pentose phosphate pathway, discussed in Chapter 9 of MCAT Biochemistry Review.
THE CYTOSKELETON
The cytoskeleton, shown in Figure 1.4, provides structure to the cell and helps it to maintain its shape. In addition, the cytoskeleton provides a conduit for the transport of materials around the cell. There are three components of the cytoskeleton: microfilaments, microtubules, and intermediate filaments.
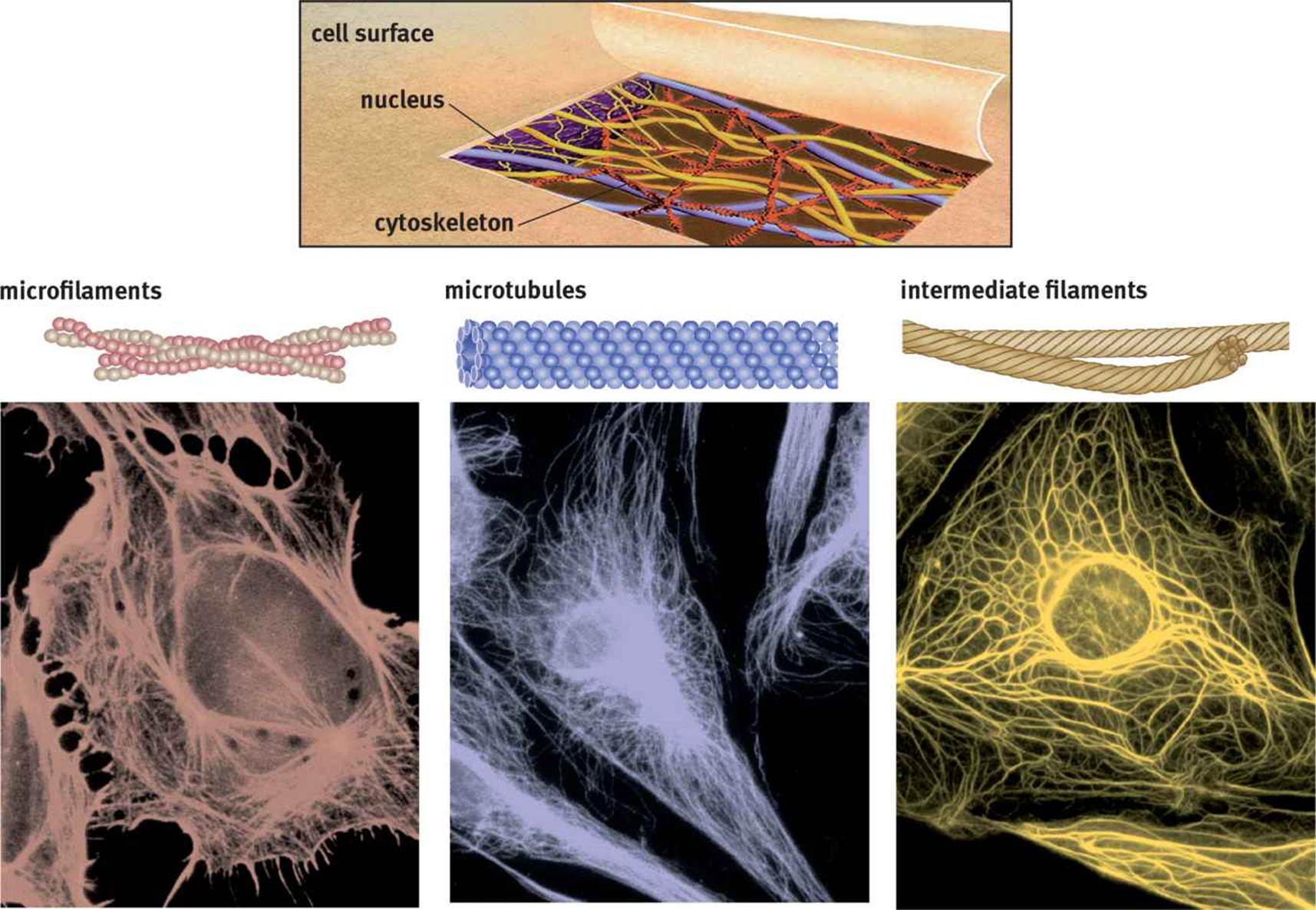 Figure 1.4. Cytoskeletal Elements The rounded shape near the center in each of these photographs is the nucleus.
Figure 1.4. Cytoskeletal Elements The rounded shape near the center in each of these photographs is the nucleus.
Microfilaments
Microfilaments are made up of solid polymerized rods of actin. The actin filaments are organized into bundles and networks and are resistant to both compression and fracture, providing protection for the cell. Actin filaments can also use ATP to generate force for movement by interacting with myosin, such as in muscle contraction.
Microfilaments also play a role in cytokinesis, or the division of materials between daughter cells. During mitosis, the cleavage furrow is formed from microfilaments, which organize as a ring at the site of division between the two new daughter cells. As the actin filaments within this ring contract, the ring becomes smaller, eventually pinching off the connection between the two daughter cells.
Microtubules
Unlike microfilaments, microtubules are hollow polymers of tubulin proteins. Microtubules radiate throughout the cell, providing the primary pathways along which motor proteins like kinesin and dynein carry vesicles.
BRIDGE
Motor proteins like kinesin and dynein are classic examples of nonenzymatic proteins, along with binding proteins, cell adhesion molecules, immunoglobulins, and ion channels. Motor proteins often travel along cytoskeletal structures to accomplish their functions. Nonenzymatic proteins are discussed in Chapter 3 of MCAT Biochemistry Review.
Cilia and flagella are motile structures composed of microtubules. Cilia are projections from a cell that are primarily involved in movement of materials along the surface of the cell; for example, cilia line the respiratory tract and are involved in movement of mucus. Flagella are structures involved in movement of the cell itself, such as the movement of sperm cells through the reproductive tract. Cilia and flagella share the same structure, composed of nine pairs of microtubules forming an outer ring, with two microtubules in the center, as shown in Figure 1.5. This is known as a 9 + 2 structure and is seen only in eukaryotic organelles of motility. Bacterial flagella have a different structure with a different chemical composition.
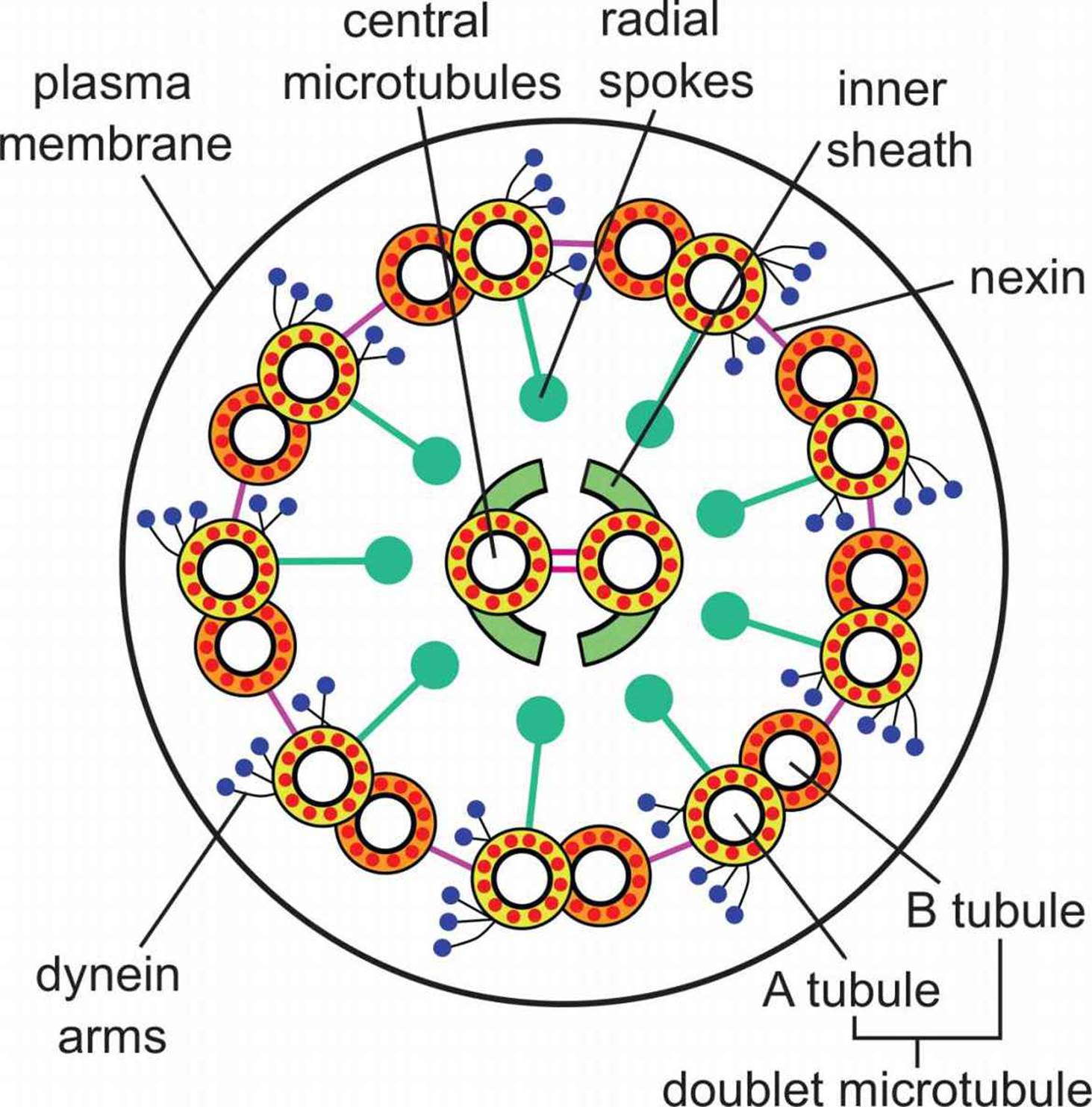 Figure 1.5. Cilium and Flagellum Structure Microtubules are organized into a ring of 9 doublets with 2 central microtubules.
Figure 1.5. Cilium and Flagellum Structure Microtubules are organized into a ring of 9 doublets with 2 central microtubules.
Centrioles are found in a region of the cell called the centrosome. They are the organizing centers for microtubules and are structured as nine triplets of microtubules with a hollow center. During mitosis, the centrioles migrate to opposite poles of the dividing cell and organize the mitotic spindle. The microtubules emanating from the centrioles attach to the chromosomes via complexes called kinetochores and can exert force on the sister chromatids, pulling them apart.
Intermediate Filaments
Intermediate filaments are a diverse group of filamentous proteins, including keratin and desmin. Many intermediate filaments are involved in cell–cell adhesion or maintenance of the overall integrity of the cytoskeleton. Intermediate filaments are able to withstand a tremendous amount of tension, making the cell structure more rigid. In addition, intermediate filaments help anchor other organelles, including the nucleus. The identity of the intermediate filament proteins within a cell is specific to the cell and tissue type.
TISSUE FORMATION
One of the unique characteristics of eukaryotic cells is the formation of tissues with division of labor, as different cells in a tissue may carry out different functions. For example, in the heart, some cells participate in the conduction pathways while others cause contraction; still others serve a supportive role, maintaining structural integrity of the organ. There are four tissue types: epithelial tissue, connective tissue, muscle, and nervous tissue. While muscle and nervous tissue are considered more extensively in subsequent chapters, we explore epithelial and connective tissues below.
Epithelial Tissue
Epithelial tissues cover the body and line its cavities, providing a means for protection against pathogen invasion and desiccation. In certain organs, epithelial cells are involved in absorption, secretion, and sensation. To remain one cohesive unit, epithelial cells are tightly joined to each other and to an underlying layer of connective tissue known as the basement membrane. Epithelial cells are highly diverse and serve numerous functions depending on the identity of the organ in which they are found; in most organs, epithelial cells constitute the parenchyma, or the functional parts of the organ. For example, nephrons in the kidney, hepatocytes in the liver, and acid-producing cells of the stomach are all composed of epithelial cells.
Epithelial cells are often polarized, meaning that one side faces a lumen (the hollow inside of an organ or tube) or the outside world, while the other side interacts with blood vessels and structural cells. For example, in the small intestine, one side of the cell will be involved in absorption of nutrients from the lumen, while the other side will be involved in releasing those nutrients into circulation for use in the rest of the body.
We can classify different epithelia according to the number of layers they have and the shape of their cells. Simple epithelia have one layer of cells; stratified epithelia have multiple layers; and pseudostratified epithelia appear to have multiple layers due to differences in cell height, but are, in reality, only one layer. Turning to shape, cells may be classified as cuboidal, columnar, or squamous. As their names imply, cuboidal cells are cube-shaped and columnar cells are long and thin. Squamous cells are flat and scalelike.
Connective Tissue
Connective tissue supports the body and provides a framework for the epithelial cells to carry out their functions. Whereas epithelial cells contribute to the parenchyma of an organ, connective tissues are the main contributors to the stroma or support structure. Bone, cartilage, tendons, ligaments, adipose tissue, and blood are all examples of connective tissues. Most cells in connective tissues produce and secrete materials such as collagen and elastin to form the extracellular matrix.
MCAT Concept Check 1.2:
Before you move on, assess your understanding of the material with these questions.
1. Briefly describe the functions of each of the organelles listed below:
· Nucleus:
· Mitochondrion:
· Lysosome:
· Rough endoplasmic reticulum:
· Smooth endoplasmic reticulum:
· Golgi apparatus:
· Peroxisome:
2. A child is diagnosed with an enzyme deficiency that prevents the production of hydrogen peroxide. What would the likely outcome be of such a deficiency?
3. What are the predominant proteins in each cytoskeletal element?
· Microfilaments:
· Microtubules:
· Intermediate filaments:
4. How do the cytoskeletal structures of centrioles and flagella differ?
5. Classify each of the following cells as epithelial cells or connective tissue:
· Fibroblasts, which produce collagen in a number of organs:
· Endothelial cells, which line blood vessels:
· α-cells, which produce glucagon in the pancreas:
· Osteoblasts, which produce osteoid, the material that hardens into bone:
· Chondroblasts, which produce cartilage: