MCAT Biology Review
Chapter 5: The Endocrine System
Introduction
Human beings consist of many trillions of cells that must work together to sustain life. Fuel resources must be conserved or used appropriately, such as during responses to stressful situations. In addition, organs must be able to communicate with the brain to cause a change in behavior and physiology to maintain homeostasis. Hormones, as the messengers in the endocrine system, play an essential role in this communication. For example, the pancreas produces both insulin and glucagon. Insulin favors the transport of glucose into organs as well as the storage of excess glucose when blood glucose concentrations are high. Conversely, glucagon triggers the release of sugar stores and raises blood glucose concentration. Working together, these hormones ensure that there is enough glucose available for organ function, but that glucose levels are not so high as to cause damage to organ systems.
This is no inconsequential fact. Diabetes mellitus is one of the most common diagnoses in the United States and a major cause of morbidity and mortality. Type 1 diabetes mellitus is an autoimmune disease in which insulin-producing cells in the islets of Langerhans are destroyed; type 2 diabetes mellitus is caused by end-organ insensitivity to insulin. In both cases, blood glucose concentrations rise to dangerous levels (sometimes up to ten times the normal concentration) and can cause significant damage to multiple organs, including the retina of the eye, the glomeruli of the kidneys, the coronary vessels of the heart and cerebral vessels of the brain, and nerves in the extremities. Left untreated (or, to be frank, even if treated in many cases), diabetes can lead to blindness, kidney failure, heart attacks, strokes, and amputation. Regardless of the field you enter, you will spend a significant amount of time working with diabetic patients and will have to think about the effects of this diagnosis on other diagnoses and their treatment.
In this chapter, we will explore the different types of hormones and how they work. We’ll survey the various endocrine organs and discuss the hormones each one produces. This is an extremely high-yield chapter: the MCAT frequently tests not only the content of the endocrine system (the hormones and their functions), but also the reasoning of the endocrine system (feedback loops and their regulation). Return to this chapter frequently during studying; a thorough knowledge of this system will assuredly pay off in points on Test Day.
5.1 Mechanisms of Hormone Action
The endocrine system consists of organs, known as glands, that secrete hormones. Hormones are signaling molecules that are secreted directly into the bloodstream to travel to a distant target tissue. At that tissue, hormones bind to receptors, inducing a change in gene expression or cellular functioning. Not all hormones share the same structure and function. In order to understand how each hormone functions, it is first important to understand basic hormone structure.
CLASSIFICATION OF HORMONES BY CHEMICAL STRUCTURE
Hormones can be subdivided into categories based on different criteria. First, hormones can be classified by their chemical identities. Hormones can be peptides, steroids, or amino acid derivatives.
Peptide Hormones
Peptide hormones are made up of amino acids, ranging in size from quite small (such as ADH) to relative large (such as insulin). Peptide hormones are all derived from larger precursor polypeptides that are cleaved during posttranslational modification. These smaller units are transported to the Golgi apparatus for further modifications that activate the hormone and direct it to the correct location in the cell. Such hormones are released by exocytosis after being packaged into vesicles.
Because peptide hormones are charged and cannot pass through the plasma membrane, these hormones must bind to an extracellular receptor. The peptide hormone is considered the first messenger; it binds to the receptor and triggers the transmission of a second signal, known as thesecond messenger. There are many different receptor subtypes, and the type of receptor determines what happens once the hormone has stimulated the receptor.
The connection between the hormone at the surface and the effect brought about by second messengers within the cell is known as a signaling cascade. At each step, there is the possibility of amplification. For example, one hormone molecule may bind to multiple receptors before it is degraded. Also, each receptor may activate multiple enzymes, each of which will trigger the production of large quantities of second messengers. Thus, each step can result in an increase in the intensity of the signal. Some common second messengers are cyclic adenosine monophosphate (cAMP), inositol triphosphate (IP3), and calcium. The activation of a G protein-coupled receptor is shown in Figure 5.1. In this system, the binding of a peptide hormone triggers the receptor to either activate or inhibit an enzyme called adenylate cyclase, raising or lowering the levels of cAMP accordingly. cAMP can bind to intracellular targets, such as protein kinase A, which phosphorylates transcription factors like cAMP response element-binding protein (CREB) to exert the hormone’s ultimate effect. Keep in mind that protein kinase A can modify other enzymes as well as transcription factors, thus it can have a rapid or slow effect on the cell.
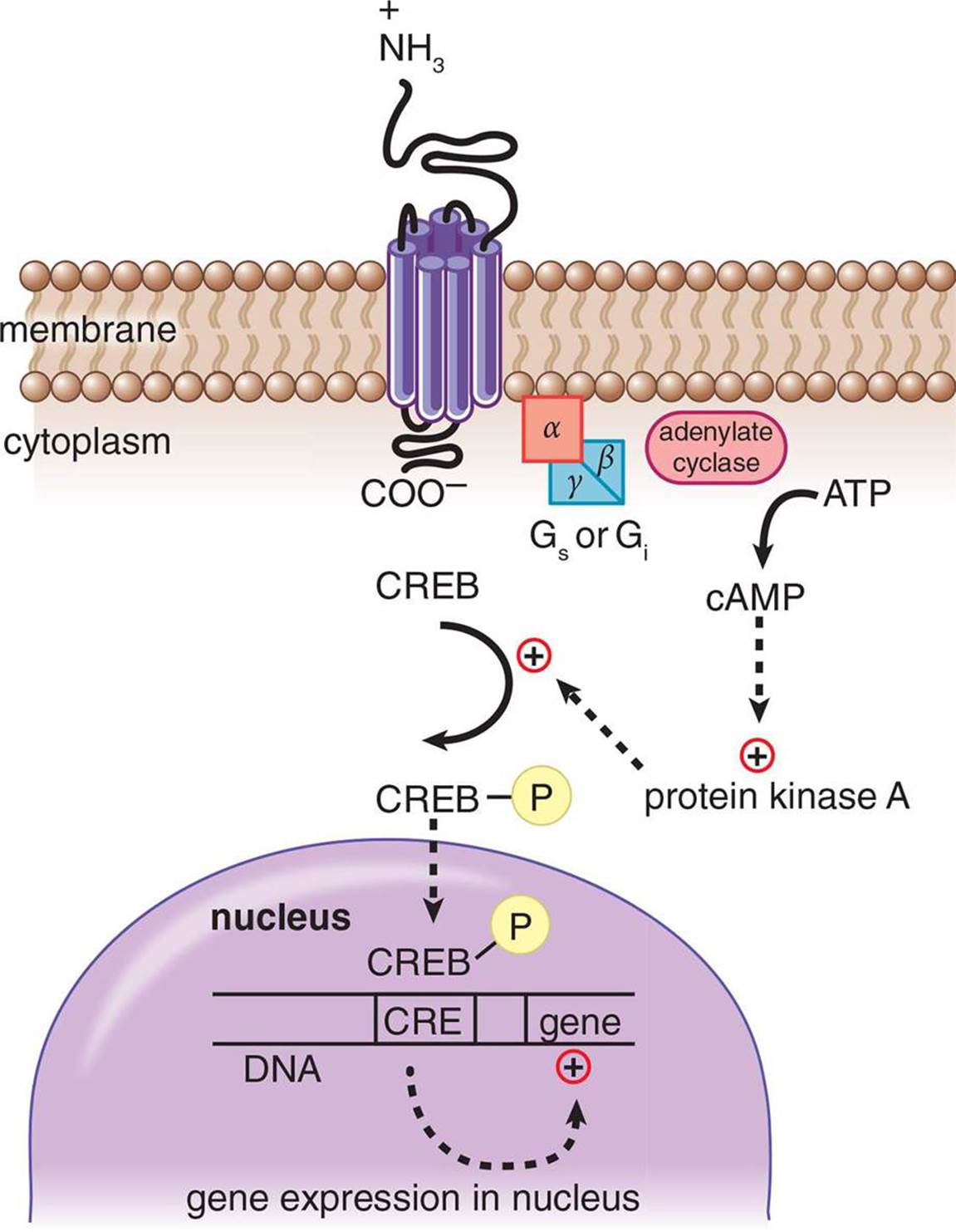 Figure 5.1. Mechanism of Action of a Peptide Hormone Peptide hormones bind to membrane-bound receptors to intiate a signal cascade, using second messengers like cAMP.
Figure 5.1. Mechanism of Action of a Peptide Hormone Peptide hormones bind to membrane-bound receptors to intiate a signal cascade, using second messengers like cAMP.
The effects of peptide hormones are usually rapid but short-lived because these hormones act through transient second messenger systems. It is quicker to turn them on and off, compared with steroid hormones, but their effects do not last without relatively constant stimulation.
Because peptides are generally water-soluble, peptide hormones can travel freely in the bloodstream and usually do not require carriers. This is in stark contrast to steroid hormones, as we will explore in the next section.
Steroid Hormones
Steroid hormones are derived from cholesterol and are produced primarily by the gonads and adrenal cortex. Because steroid hormones are derived from nonpolar molecules, they can easily cross the cell membrane. In fact, their receptors are usually intracellular (in the cytosol) or intranuclear (in the nucleus). Upon binding to the receptor, steroid hormone–receptor complexes undergo conformational changes. The receptor can then bind directly to DNA, resulting in either increased or decreased transcription of particular genes, depending on the identity of the hormone, as shown in Figure 5.2. One common form of conformational change is dimerization, or pairing of two receptor–hormone complexes. The effects of steroid hormones are slower but longer-lived than peptide hormones because steroid hormones cause alterations in the amount of mRNA and protein present in a cell.
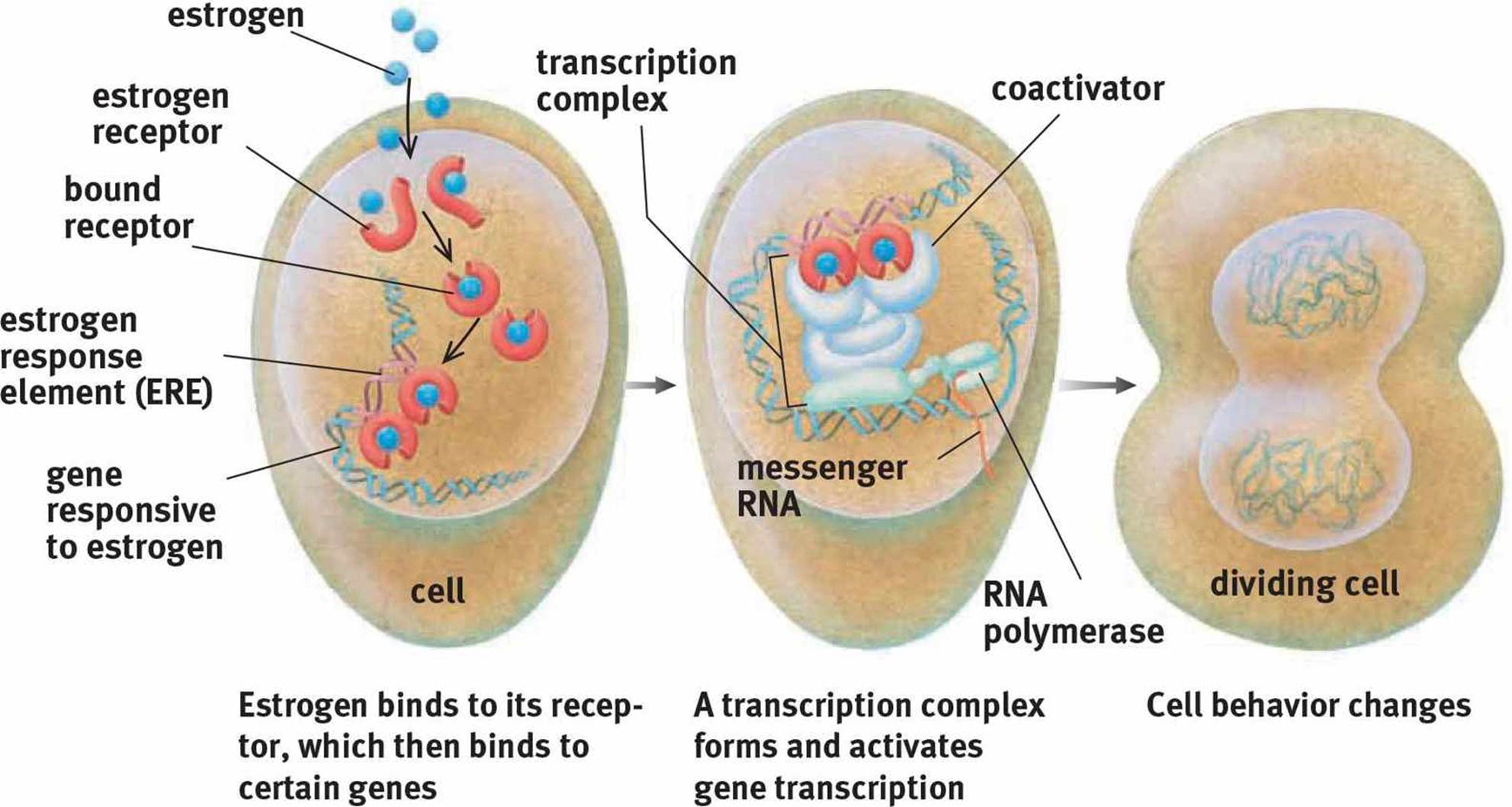 Figure 5.2. Mechanism of Action of a Steroid Hormone Estrogen, like all steroid hormones, influences cell behavior by modifying transcription.
Figure 5.2. Mechanism of Action of a Steroid Hormone Estrogen, like all steroid hormones, influences cell behavior by modifying transcription.
KEY CONCEPT
Peptide hormones have surface receptors and act via second messenger systems. Steroid hormones bind to intracellular receptors and function by binding to DNA to alter gene transcription.
MNEMONIC
Insulin is a peptide hormone, and it has to be released at every meal in order to be active. Thus, it has fast onset but is short-acting (as most peptide hormones are). Estrogen and testosterone are steroid hormones that promote sexual maturation. This is a slower, but longer-lasting change (as is true for most steroid hormones).
Steroid hormones are not water-soluble and, thus, must be carried by proteins in the bloodstream to be able to travel around the body. Some of these proteins are very specific, and carry only one hormone (such as sex hormone-binding globulin), while other proteins are nonspecific (such asalbumin). Note that hormones are generally inactive while attached to a carrier protein and must dissociate from the carrier to function. Levels of carrier proteins can change the levels of active hormone. For example, some conditions increase the quantity of the protein that carries thyroid hormones, thyroxine-binding globulin (TBG). This causes the body to perceive a lower level of thyroid hormone because the increased quantity of TBG binds a larger proportion of the hormone, meaning there is less free hormone available.
REAL WORLD
During pregnancy, high levels of estrogen and progesterone cause increased production of TBG, thyroxine-binding globulin. In order to compensate, pregnant women secrete much higher levels of the thyroid hormones. Thus, in order to diagnose thyroid disease in a pregnant woman, different reference values must be used.
Amino Acid-Derivative Hormones
Finally, amino acid-derivative hormones are less common than peptide and steroid hormones, but include some of the most important hormones discussed in this chapter, including epinephrine, norepinephrine, triiodothyronine, and thyroxine. These hormones are derived from one or two amino acids, usually with a few additional modifications. For example, thyroid hormones are made from tyrosine with the addition of several iodine atoms.
This chemistry of this family of hormones is considerably less predictable and is one of the few instances where overt memorization may be the best strategy. The catecholamines (epinephrine and norepinephrine) bind to G protein-coupled receptors while the thyroid hormones bind intracellularly.
MCAT EXPERTISE
The mechanism of action of the amino acid-derivative hormones should be memorized because it is so unpredictable. Epinephrine and norepinephrine have extremely fast onset but are short-lived, like peptide hormones—think of an adrenaline rush. Thyroxine and triiodothyronine, on the other hand, have slower onset but a longer duration, like steroid hormones—they regulate metabolic rate over a long period of time.
CLASSIFICATION OF HORMONES BY TARGET TISSUE
Some hormones, known as direct hormones, are secreted and then act directly on a target tissue. For example, insulin released by the pancreas causes increased uptake of glucose by muscles. Other hormones, known as tropic hormones, require an intermediary to act. For example, as discussed in Chapter 2 of MCAT Biology Review, gonadotropin-releasing hormone (GnRH) from the hypothalamus stimulates the release of luteinizing hormone (LH) and follicle-stimulating hormone (FSH). LH then acts on the gonads to stimulate testosterone production in the male and estrogen production in the female. GnRH and LH do not cause direct changes in the physiology of muscle, bone, and hair follicles; rather, they stimulate the production of another hormone by another endocrine gland that acts on these target tissues. Tropic hormones usually originate in the brain and anterior pituitary gland, as these structures are involved in coordination of multiple processes within the body.
MNEMONIC
Most peptide and amino acid-derivative hormones have names that end in –in or –ine (insulin, vasopressin, thyroxine, triiodothyronine, and so on). Most steroid hormones have names that end in –one, –ol, or –oid (testosterone, aldosterone and other mineralocorticoids, cortisol and other glucocorticoids, and so on). This is not exhaustive, but may help you identify the chemistry of a hormone on Test Day.
MCAT Concept Check 5.1:
Before you move on, assess your understanding of the material with these questions.
1. Compare and contrast peptide and steroid hormones based on the following criteria:
|
Criterion |
Peptide Hormones |
Steroid Hormones |
|
Chemical precursor |
||
|
Location of receptor |
||
|
Mechanism of action |
||
|
Method of travel in the bloodstream |
||
|
Rapidness of onset |
||
|
Duration of action |
2. How are amino acid-derivative hormones synthesized?
3. What is the difference between a direct and a tropic hormone?
· Direct hormone:
· Tropic hormone: