Medical Microbiology
Section 4 Clinical manifestation and diagnosis of infections by body system
22 Gastrointestinal tract infections
Introduction
Ingested pathogens may cause disease confined to the gut or involve other parts of the body
Ingestion of pathogens can cause many different infections. These may be confined to the gastrointestinal tract or are initiated in the gut before spreading to other parts of the body. In this chapter, we consider the important bacterial causes of diarrheal disease and summarize the other bacterial causes of food-associated infection and food poisoning. Viral and parasitic causes of diarrheal disease are discussed, as well as infections acquired via the gastrointestinal tract and causing disease in other body systems, including typhoid and paratyphoid fevers, listeriosis and some forms of viral hepatitis. For clarity, all types of viral hepatitis are included in this chapter, despite the fact that some are transmitted by other routes of infection. Infections of the liver can also result in liver abscesses, and several parasitic infections cause liver disease. Peritonitis and intra-abdominal abscesses can arise from seeding of the abdominal cavity by organisms from the gastrointestinal tract. Several different terms are used to describe infections of the gastrointestinal tract; those in common use are shown in Box 22.1.
![]()
Box 22.1  Terms Used to Describe Gastrointestinal Tract Infections
Terms Used to Describe Gastrointestinal Tract Infections
As well as many colloquial expressions, several different clinical terms are used to describe infections of the gastrointestinal tract. Diarrhea without blood and pus is usually the result of enterotoxin production, whereas the presence of blood and/or pus cells in the faeces indicates an invasive infection with mucosal destruction.
• Gastroenteritis
• A syndrome characterized by gastrointestinal symptoms including nausea, vomiting, diarrhea and abdominal discomfort
• Diarrhea
• Abnormal faecal discharge characterized by frequent and/or fluid stool; usually resulting from disease of the small intestine and involving increased fluid and electrolyte loss
• Dysentery
• An inflammatory disorder of the gastrointestinal tract often associated with blood and pus in the faeces and accompanied by symptoms of pain, fever, abdominal cramps; usually resulting from disease of the large intestine
• Enterocolitis
• Inflammation involving the mucosa of both the small and large intestine
![]()
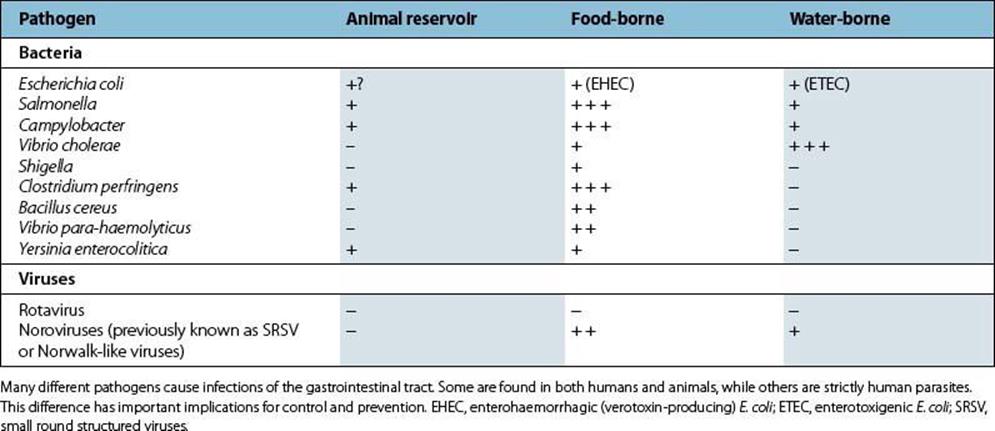
A wide range of microbial pathogens is capable of infecting the gastrointestinal tract, and the important bacterial and viral pathogens are listed in Table 22.1. They are acquired by the faecal–oral route, from faecally contaminated food, fluids or fingers.
Table 22.1 Important bacterial and viral pathogens of the gastrointestinal tract
For an infection to occur, the pathogen must be ingested in sufficient numbers or possess attributes to elude the host defences of the upper gastrointestinal tract and reach the intestine (Fig. 22.1; see also Ch. 13). Here they remain localized and cause disease as a result of multiplication and/or toxin production, or they may invade through the intestinal mucosa to reach the lymphatics or the bloodstream (Fig. 22.2). The damaging effects resulting from infection of the gastrointestinal tract are summarized in Box 22.2.
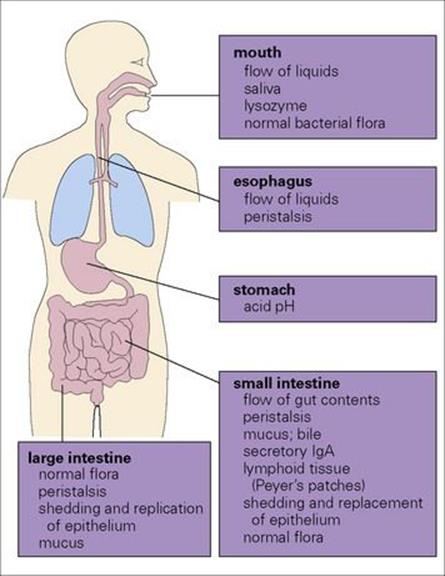
Figure 22.1 Every day we swallow large numbers of microorganisms. Because of the body’s defence mechanisms, however, they rarely succeed in surviving the passage to the intestine in sufficient numbers to cause infection.
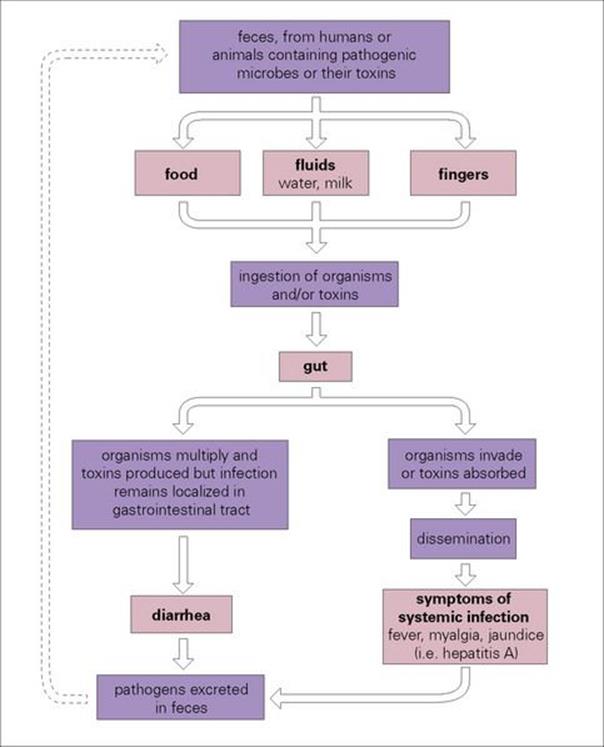
Figure 22.2 Infections of the gastrointestinal tract can be grouped into those that remain localized in the gut and those that invade beyond the gut to cause infection in other sites in the body. In order to spread to a new host, pathogens are excreted in large numbers in the faeces and must survive in the environment for long enough to infect another person directly or indirectly through contaminated food or fluids.
![]()
Box 22.2 Damage Resulting from Infection of The Gastrointestinal Tract
• Pharmacologic action of bacterial toxins, local or distant to site of infection, e.g. cholera, staphylococcal food poisoning
• Local inflammation in response to superficial microbial invasion, e.g. shigellosis, amoebiasis
• Deep invasion to blood or lymphatics; dissemination to other body sites, e.g. Hepatitis A, enteric fevers
• Perforation of mucosal epithelium after infection, surgery or accidental trauma, e.g. peritonitis, intra-abdominal abscesses.
Infection of the gastrointestinal tract can cause damage locally or at distant sites.
![]()
Food-associated infection versus food poisoning
Infection associated with consumption of contaminated food is often termed ‘food poisoning’, but ‘food-associated infection’ is a better term. True food poisoning occurs after consumption of food containing toxins, which may be chemical (e.g. heavy metals) or bacterial in origin (e.g. fromClostridium botulinum or Staphylococcus aureus). The bacteria multiply and produce toxins within contaminated food. The organisms may be destroyed during food preparation, but the toxin is unaffected, consumed and acts within hours. In food-associated infections, the food may simply act as a vehicle for the pathogen (e.g. Campylobacter) or provide conditions in which the pathogen can multiply to produce numbers large enough to cause disease (e.g. Salmonella).
Diarrheal diseases caused by bacterial or viral infection
Diarrhea is the most common outcome of gastrointestinal tract infection
Infections of the gastrointestinal tract range in their effects from a mild self-limiting attack of ‘the runs’ to severe, sometimes fatal, diarrhea. There may be associated vomiting, fever and malaise. Diarrhea is the result of an increase in fluid and electrolyte loss into the gut lumen, leading to the production of unformed or liquid faeces and can be thought of as the method by which the host forcibly expels the pathogen (and in doing so, aids its dissemination). However, diarrhea also occurs in many non-infectious conditions, and an infectious cause should not be assumed.
In the resource-poor world, diarrheal disease is a major cause of mortality in children
In the resource-poor world, diarrheal disease is a major cause of morbidity and mortality, particularly in young children. In the resource-rich world, it remains a very common complaint, but is usually mild and self-limiting except in the very young, the elderly and immunocompromised patients. Most of the pathogens listed in Table 22.1 are found throughout the world, but some, such as Vibrio cholerae, have a more limited geographic distribution. However, such infections can be acquired by travellers to these areas and imported into their home countries.
Many cases of diarrheal disease are not diagnosed, either because they are mild and self-limiting and the patient does not seek medical attention, or because medical and laboratory facilities are unavailable, particularly in resource-poor countries. It is generally impossible to distinguish on clinical grounds between infections caused by the different pathogens. However, information about the patient’s recent food and travel history, and macroscopic and microscopic examination of the faeces for blood and pus can provide helpful clues. A precise diagnosis can only be achieved by laboratory investigations. This is especially important in outbreaks, because of the need to instigate appropriate epidemiologic investigations and control measures.
Bacterial causes of diarrhea
Escherichia coli
E. coli is one of the most versatile of all bacterial pathogens. Some strains are important members of the normal gut flora in humans and animals (see Ch. 2), whereas others possess virulence factors that enable them to cause infections in the intestinal tract or at other sites, particularly the urinary tract (see Ch. 20). Strains that cause diarrheal disease do so by several distinct pathogenic mechanisms and differ in their epidemiology (Table 22.2).
Table 22.2 Characteristics of Escherichia coli strains causing gastrointestinal infections
|
Pathogenic group |
Epidemiology |
Laboratory diagnosis* |
|
Enteropathogenic E. coli (EPEC) |
EPEC strains belong to particular O serotypes |
Isolate organisms from faeces |
|
Enterotoxigenic E. coli (ETEC) |
Most important bacterial cause of diarrhea in children in resource-poor countries |
Isolate organisms from faeces |
|
Enterohaemorrhagic(verotoxin-producing) E. coli (EHEC) |
Serotype O157 most important EHEC in human infections |
Isolate organisms from faeces |
|
Enteroinvasive E. coli (EIEC) |
Important cause of diarrhea in areas of poor hygiene |
Isolate organisms from faeces |
|
Enteroaggregative E. coli (EAEC) |
Characteristic attachment to tissue culture cells |
Tissue culture assays for aggregative or diffuse adherence |
E. coli is a major cause of gastrointestinal infection, particularly in resource-poor countries and in travellers. There is a range of pathogenic mechanisms within the species, resulting in more or less invasive disease.
* Specialized tests are given in italics. LT, heat-labile enterotoxin; ST, heat-stable enterotoxin.
There are six distinct groups of E. coli with different pathogenetic mechanisms
Initially, all diarrhea-associated Escherichia coli were termed enteropathogenic E. coli (EPEC). However, greater insight into mechanisms of pathogenicity has led to specific group designations: enteropathogenic E. coli (EPEC), enterotoxigenic E. coli (ETEC), enterohaemorrhagic E. coli(EHEC), enteroinvasive E. coli (EIEC), enteroaggregative E. coli (EAEC), and diffuse-aggregative E. coli (DAEC).
Enteropathogenic E. coli (EPEC) do not appear to make any toxins
They do produce bundle-forming pili (Bfp), intimin (an adhesin) and an associated protein (translocated intimin receptor, Tir). These virulence factors allow bacterial attachment to epithelial cells of the small intestine, leading to disruption of the microvillus (an ‘attaching–effacing’ mechanism of action; Table 22.2; Fig. 22.3) leading to diarrhea (Table 22.3).
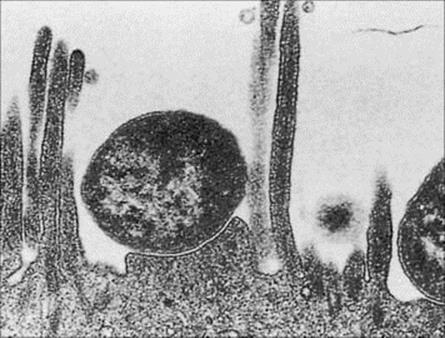
Figure 22.3 Electron micrograph of enteropathogenic E. coli adhering to the brush border of intestinal mucosal cells with localized destruction of microvilli.
(Courtesy of S. Knutton.)
Table 22.3 The clinical features of bacterial diarrheal infection
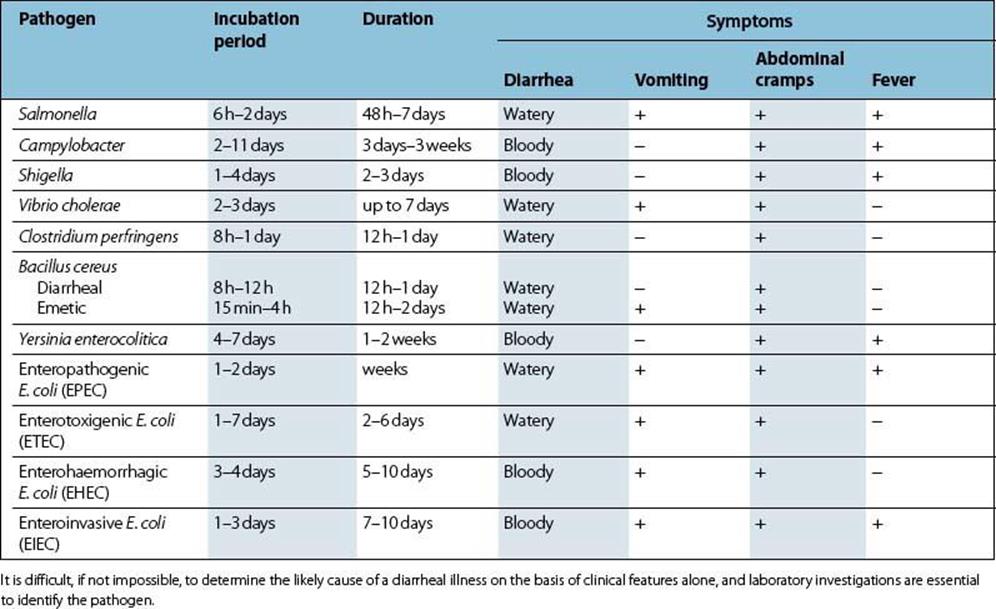
Enterotoxigenic E. coli (ETEC) possess colonization factors (fimbrial adhesins)
These bind the bacteria to specific receptors on the cell membrane of the small intestine (Table 22.2; Fig. 22.4). These organisms produce powerful plasmid-associated enterotoxins which are characterized as being either heat labile (LT) or heat stable (ST):
• Heat-labile enterotoxin LT-I is very similar in structure and mode of action to cholera toxin produced by V. cholerae, and infections with strains producing LT-I can mimic cholera, particularly in young and malnourished children (Table 22.3).
• Other ETEC strains produce heat-stable enterotoxins (STs) in addition to or instead of LT. STs have a similar but distinct mode of action to that of LT. STA activates guanylate cyclase activity, causing an increase in cyclic guanosine monophosphate, which results in increased fluid secretion. Immunoassays are commercially available for the identification of ETEC (Table 22.2).
Figure 22.4 Electron micrograph of enterotoxin E. coli, showing pili necessary for adherence to mucosal epithelial cells.
(Courtesy of S. Knutton.)
Enterohaemorrhagic E. coli (EHEC) isolates produce a verotoxin
The verotoxin (i.e. toxic to tissue cultures of ‘vero’ cells) is essentially identical to Shiga (Shigella) toxin. After attachment to the mucosa of the large intestine (by the ‘attaching– effacing’ mechanism also seen in EPEC), the produced toxin has a direct effect on intestinal epithelium, resulting in diarrhea (Table 22.3). EHEC cause haemorrhagic colitis (HC) and haemolytic-uraemic syndrome (HUS). In HC, there is destruction of the mucosa and consequent haemorrhage; this may be followed by HUS. Verotoxin receptors have been identified on renal epithelium and may account for kidney involvement. While there are many serotypes of EHEC, the most common is O157:H7.
Enteroinvasive E. coli (EIEC) attach specifically to the mucosa of the large intestine
They invade the cells by endocytosis by using plasmid-associated genes. Inside the cell, they lyse the endocytic vacuole, multiply and spread to adjacent cells, causing tissue destruction, inflammation, necrosis and ulceration, resulting in blood and mucus in stools (Tables 22.2, 22.3).
Enteroaggregative E. coli (EAEC) derive their name from their characteristic attachment pattern to tissue culture cells
The pattern is an aggregative or ‘stacked brick’ formation. These organisms act in the small intestine to cause persistent diarrhea, especially in children in resource-poor countries. Their aggregative adherence ability is due to plasmid-associated fimbriae. EAEC also produce heat-labile toxins (an enterotoxin and a toxin related to E. coli haemolysin) but their role in diarrheal disease is uncertain.
Diffuse-aggregative E. coli (DAEC) produce an alpha haemolysin and cytotoxic necrotizing factor 1
They are also known as diffuse-adherent or cell-detaching E. coli. Their role in diarrheal disease, especially in young children, is incompletely understood and somewhat controversial, with some studies reporting no association.
EPEC and ETEC are the most important contributors to global incidence of diarrhea, while EHEC is more important in resource-rich countries
The diarrhea produced by E. coli varies from mild to severe, depending upon the strain and the underlying health of the host. ETEC diarrhea in children in resource-poor countries may be clinically indistinguishable from cholera. EIEC and EHEC strains both cause bloody diarrhea (Table 22.3). Following EHEC infection, HUS is characterized by acute renal failure (Fig. 22.5), anaemia and thrombocytopenia, and there may be neurologic complications. HUS is the most common cause of acute renal failure in children in the UK and USA. Although E. coli O157:H7 is the most commonly recognized serotype involved in HUS, E. coli 0104:H4, that had not been reported as causing an outbreak previously, caused a significant outbreak of HUS and bloody diarrhea in 15 countries across Europe in 2011. Over several months starting in May 2011, 860 individuals with HUS and over 3000 with bloody diarrhea were reported in Germany, many of whom had laboratory confirmed E. coli 0104:H4 infection. More than 50 people died and the likely vehicle was sprouted beans imported from the Middle East.
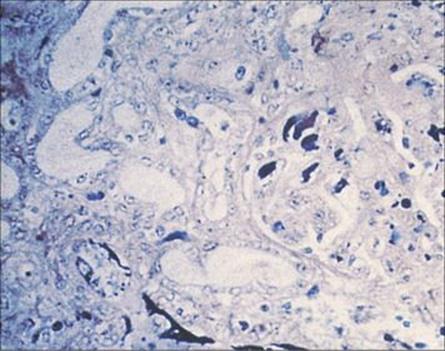
Figure 22.5 Verotoxin-producing E. coli infection, showing fibrin ‘thrombi’ in glomerular capillaries in haemolytic–uraemic syndrome. (Weigert stain.)
(Courtesy of H.R. Powell.)
Specific tests are needed to identify strains of pathogenic E. coli
Because E. coli is a member of the normal gastrointestinal flora, specific tests are required to identify strains that may be responsible for diarrheal disease. These are summarized in Table 22.2. Infections are more common in children and are also often travel-associated, and these factors should be considered when samples are received in the laboratory. It is important to note that specialized tests beyond routine stool cultures are required to identify specific diarrhea-associated E. coli types. Such tests are not ordinarily performed with uncomplicated diarrhea, which is usually self-limiting. However, concern regarding EHEC (e.g. bloody diarrhea) has led most laboratories in resource-rich countries to screen for E. coli O157:H7.
Antibacterial therapy is not indicated for E. coli diarrhea
Specific antibacterial therapy is not indicated. Fluid replacement may be necessary, especially in young children. Treatment of HUS is urgent and may involve dialysis.
Provision of a clean water supply and adequate systems for sewage disposal are fundamental to the prevention of disease. Food and unpasteurized milk can be important vehicles of infection, especially for EIEC and EHEC, but there is no evidence of an animal or environmental reservoir.
Salmonella
Salmonellae are the most common cause of food-associated diarrhea in many resource-rich countries
However, in some countries such as the USA and UK, they have been relegated to second place by Campylobacter. Like E. coli, the salmonellae belong to the family Enterobacteriaceae. Historically, salmonella nomenclature has been somewhat confusing, with more than 2000 serotypes defined on the basis of differences in the cell wall (O) and flagellar (H) antigens (Kauffmann–White scheme). However, DNA hybridization studies indicate that there are only two species, the most important of which, for human infection, is Salmonella enterica. To simplify discussion and comparison, past convention has been to replace this species name with the serotype designation. While technically incorrect (the serotype is not a species), this practice is helpful when discussing interrelationships between different isolates, e.g. in epidemiologic analysis when tracing the source of an outbreak. This convention is thus followed here to maintain continuity with other scientific literature.
All salmonellae except for Salmonella typhi and S. paratyphi are found in animals as well as humans. There is a large animal reservoir of infection, which is transmitted to humans via contaminated food, especially poultry and dairy products (Fig. 22.6). Water-borne infection is less frequent. Salmonella infection is also transmitted from person to person, and secondary spread can therefore occur, for example, within a family after one member has become infected after consuming contaminated food.
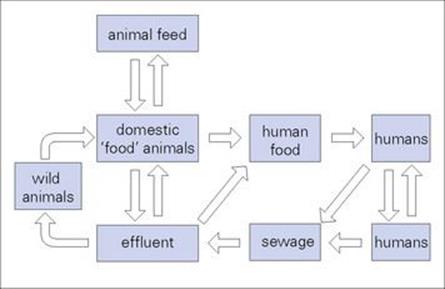
Figure 22.6 The recycling of salmonellae. With the exception of Salmonella typhi, salmonellae are widely distributed in animals, providing a constant source of infection for humans. Excretion of large numbers of salmonellae from infected individuals and carriers allows the organisms to be ‘recycled’.
Salmonellae are almost always acquired orally in food or drink that is contaminated
Diarrhea is produced as a result of invasion by the salmonellae of epithelial cells in the terminal portion of the small intestine (Fig. 22.7). Initial entry is probably through uptake by M cells (the ‘antigenic samplers’ of the bowel) with subsequent spread to epithelial cells. A similar route of invasion occurs in Shigella, Yersinia and reovirus infections. The bacteria migrate to the lamina propria layer of the ileocaecal region, where their multiplication stimulates an inflammatory response, which both confines the infection to the gastrointestinal tract and mediates the release of prostaglandins. These in turn activate cyclic adenosine monophosphate (cAMP) and fluid secretion, resulting in diarrhea.
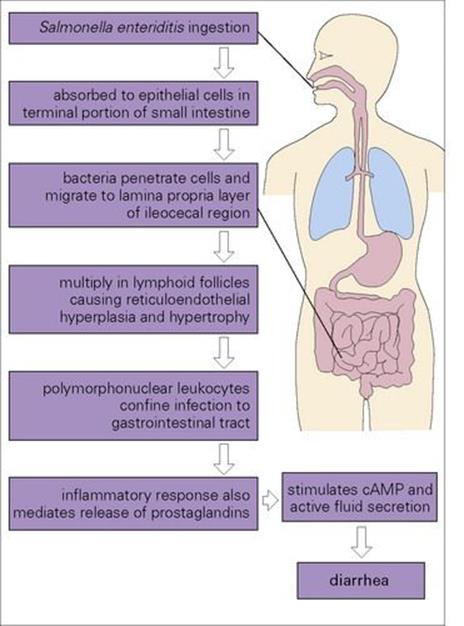
Figure 22.7 The passage of salmonellae through the body. The vast majority of salmonellae cause infection localized to the gastrointestinal tract and do not invade beyond the gut mucosa. cAMP, cyclic adenosine monophosphate.
Species of Salmonella that normally cause diarrhea (e.g. S. enteritidis, S. choleraesuis) may become invasive in patients with particular predispositions (e.g. children, immunocompromised patients or those with sickle cell anaemia). The organisms are not contained within the gastrointestinal tract, but invade the body to cause septicaemia; consequently, many organs become seeded with salmonellae, sometimes leading to osteomyelitis, pneumonia or meningitis.
In the vast majority of cases, Salmonella spp. cause an acute but self-limiting diarrhea, though in the young and the elderly the symptoms may be more severe. Vomiting is also common with enterocolitis, while fever is usually a sign of invasive disease (Table 22.3). S. typhi and S. paratyphiinvade the body from the gastrointestinal tract to cause systemic illness and are discussed in a later section.
Salmonella diarrhea can be diagnosed by culture on selective media
The methods for culturing faecal specimens on selective media are summarized in the Appendix. The organisms are not fastidious and can usually be isolated within 24 h, although small numbers may require enrichment in selenite broth before culture. Preliminary identification can be made rapidly, but the complete result, including serotype, takes at least 48 h.
Fluid and electrolyte replacement may be needed for salmonella diarrhea
Diarrhea is usually self-limiting and resolves without treatment. Fluid and electrolyte replacement may be required, particularly in the very young and the elderly. Unless there is evidence of invasion and septicaemia, antibiotics should be positively discouraged because they do not reduce the symptoms or shorten the illness, and may prolong excretion of salmonellae in the faeces. There is some evidence that symptomatic treatment with drugs that reduce diarrhea has the same adverse effect.
Salmonellae may be excreted in the faeces for several weeks after a salmonella infection
Figure 22.6 illustrates the problems associated with the prevention of salmonella infections. The large animal reservoir makes it impossible to eliminate the organisms, and preventive measures must therefore be aimed at ‘breaking the chain’ between animals and humans, and from person to person. Such measures include:
• maintaining adequate standards of public health (clean drinking water and proper sewage disposal)
• education programmes on hygienic food preparation.
Following an episode of salmonella diarrhea, an individual can continue to carry and excrete organisms in the faeces for several weeks. Although in the absence of symptoms, the organisms will not be dispersed so liberally into the environment, thorough handwashing before food handling is essential. People employed as food handlers are excluded from work until three specimens of faeces have failed to grow salmonella.
Campylobacter
Campylobacter infections are among the most common causes of diarrhea
Campylobacter spp. are curved or S-shaped Gram-negative rods (Fig. 22.8). They have long been known to cause diarrheal disease in animals, but are also one of the most common causes of diarrhea in humans. The delay in recognizing the importance of these organisms was due to their cultural requirements, which differ from those of the enterobacteria as they are microaerophilic and thermophilic (growing well at 42°C); they do not therefore grow on the media used for isolating E. coli and salmonellae. Several species of the genus Campylobacter are associated with human disease, but Campylobacter jejuni is by far the most common. Helicobacter pylori, previously classified as Campylobacter pylori, is an important cause of gastritis and gastric ulcers (see below).
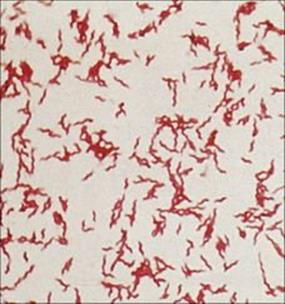
Figure 22.8 Campylobacter jejuni infection. Gram stain showing Gram-negative, S-shaped bacilli.
(Courtesy of I. Farrell.)
As with salmonellae, there is a large animal reservoir of Campylobacter in cattle, sheep, rodents, poultry and wild birds. Infections are acquired by consumption of contaminated food, especially poultry, milk or water. Studies have shown an association between infection and consumption of milk from bottles with tops that have been pecked by wild birds. Household pets such as dogs and cats can become infected and provide a source for human infection, particularly for young children. Person-to-person spread by the faecal–oral route is rare, as is transmission from food handlers.
Campylobacter diarrhea is clinically similar to that caused by other bacteria such as salmonella and shigella
The gross pathology and histologic appearances of ulceration and inflamed bleeding mucosal surfaces in the jejunum, ileum and colon (Fig. 22.9) are compatible with invasion of the bacteria, but the production of cytotoxins by C. jejuni has also been demonstrated. Invasion and bacteraemia are not uncommon, particularly in neonates and debilitated adults.
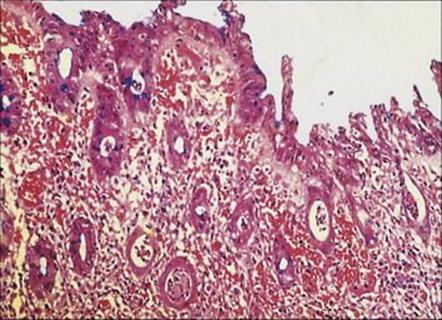
Figure 22.9 Inflammatory enteritis caused by Campylobacter jejuni, involving the entire mucosa, with flattened atrophic villi, necrotic debris in the crypts and thickening of the basement membrane. (Cresyl-fast violet stain.)
(Courtesy of J. Newman.)
The clinical presentation is similar to that of diarrhea caused by salmonellae and shigella, although the disease may have a longer incubation period and a longer duration. The key features are summarized in Table 22.3.
Cultures for Campylobacter should be set up routinely in every investigation of a diarrheal illness
The methods are described in the Appendix, but it is important to note that the media and conditions for growth differ from those required for the enterobacteria. Growth is often somewhat slow compared with that of the enterobacteria, but a presumptive identification should be available within 48 h of culture.
Erythromycin is used for severe Campylobacter diarrhea
Macrolide antibiotics such as erythromycin can be used in diarrheal disease that is severe enough to warrant treatment. Invasive infections may require treatment with an additional antibiotic such as a quinolone or an aminoglycoside.
The preventive measures for salmonella infections described above are equally applicable to the prevention of Campylobacter infections, but there are no requirements for the screening of food handlers because contamination of food by this route is very uncommon.
Cholera
Cholera is an acute infection of the gastrointestinal tract caused by the comma-shaped Gram-negative bacterium V. cholerae (Fig. 22.10). The disease has a long history characterized by epidemics and pandemics. The last cases of cholera acquired in the UK were in the nineteenth century following the introduction of the bacterium by sailors arriving from Europe, and in 1849 Snow published his historic essay On the Mode of Communication of Cholera.
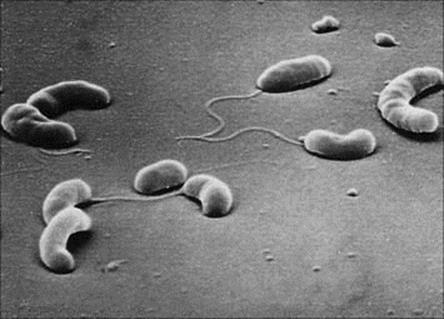
Figure 22.10 Scanning electron micrograph of Vibrio cholerae showing comma-shaped rods with a single polar flagellum (× 13 000).
(Courtesy of D.K. Banerjee.)
Cholera flourishes in communities with inadequate clean drinking water and sewage disposal
The disease remains endemic in SE Asia and parts of Africa and South America. Unlike salmonellae and Campylobacter, V. cholerae is a free-living inhabitant of fresh water, but causes infection only in humans. Asymptomatic human carriers are believed to be a major reservoir. The disease is spread via contaminated food; shellfish grown in fresh and estuarine waters have also been implicated. Direct person-to-person spread is thought to be uncommon. Therefore, cholera continues to flourish in communities where there is absent or unreliable provision of clean drinking water and sewage disposal. Cases still occur in resource-rich countries (e.g. the Gulf Coast of Louisiana and Texas in the USA), but high standards of hygiene mean that secondary spread should not occur.
V. cholerae serotypes are based on somatic (O) antigens
Serotype O1 is the most important and is further divided into two biotypes: classical and El Tor (Fig. 22.11). The El Tor biotype, named after the quarantine camp where it was first isolated from pilgrims returning from Mecca, differs from classical V. cholerae in several ways. In particular, it causes only mild diarrhea and has a higher ratio of carriers to cases than classic cholera; carriage is also more prolonged, and the organisms survive better in the environment. The El Tor biotype, which was responsible for the seventh pandemic, has now spread throughout the world and has largely displaced the classic biotype.
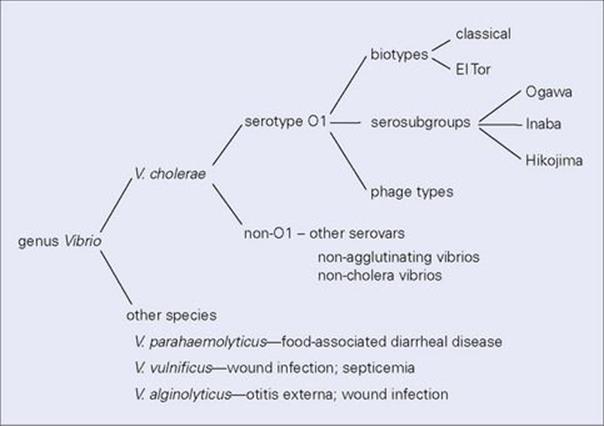
Figure 22.11 Vibrio cholerae serotype O1, the cause of cholera, can be subdivided into different biotypes with different epidemiologic features, and into sero-subgroups and phage types for the purposes of investigating outbreaks of infection. Although V. cholerae is the most important pathogen of the genus, other species can also cause infections of both the gastrointestinal tract and other sites.
In 1992, a new non-O1 strain (O139) arose in south India. It spread rapidly, infected O1-immune individuals, caused epidemics, and was the eighth pandemic strain of cholera. V. cholerae O139 appeared to have originated from the El Tor O1 biotype when the latter acquired a new O (capsular) antigen by horizontal gene transfer from a non-O1 strain. This provided the recipient strain with a selective advantage in a region where a large part of the population was immune to O1 strains.
Other species of Vibrio cause a variety of infections in humans (Fig. 22.11). V. parahaemolyticus is another cause of diarrheal disease, but this is usually much less severe than cholera (see below).
The symptoms of cholera are caused by an enterotoxin
The symptoms of cholera are entirely due to the production of an enterotoxin in the gastrointestinal tract (see Ch. 17). However, the organism requires additional virulence factors to enable it to survive the host defences and adhere to the intestinal mucosa. These are illustrated in Figure 22.12(see also Ch. 13).
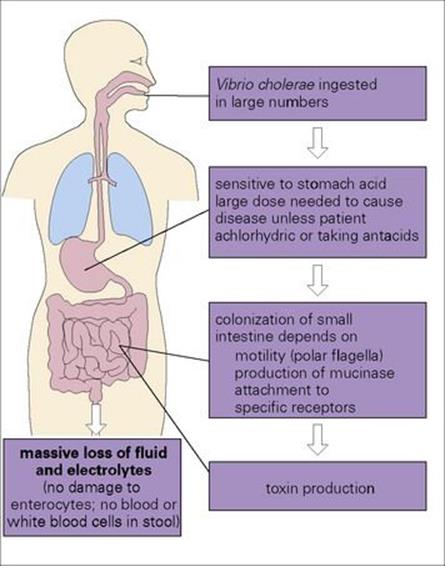
Figure 22.12 The production of an enterotoxin is central to the pathogenesis of cholera, but the organisms must possess other virulence factors to allow them to reach the small intestine and to adhere to the mucosal cells.
The clinical features of cholera are summarized in Table 22.3. The severe watery non-bloody diarrhea is known as rice water stool because of its appearance (Fig. 22.13) and can result in the loss of 1 L of fluid every hour. It is this fluid loss and the consequent electrolyte imbalance that results in marked dehydration, metabolic acidosis (loss of bicarbonate), hypokalaemia (potassium loss) and hypovolaemic shock resulting in cardiac failure. Untreated, the mortality from cholera is 40–60%; rapidly instituted fluid and electrolyte replacement reduces the mortality to < 1%.

Figure 22.13 Rice water stool in cholera.
(Courtesy of A.M. Geddes.)
Culture is necessary to diagnose sporadic or imported cases of cholera and carriers
In countries where cholera is prevalent, diagnosis is based on clinical grounds, and laboratory confirmation is rarely sought. It is worth remembering that ETEC infection can resemble cholera in both its severity and the management of infected individuals, as fluid and electrolyte replacement are of paramount importance. The methods are given in the Appendix.
Prompt rehydration with fluids and electrolytes is central to the treatment of cholera
Oral or intravenous rehydration may be used. Antibiotics are not necessary, but tetracycline may be given, as some evidence indicates that this reduces the time of excretion of V. cholerae thereby reducing the risk of transmission. There have, however, been reports of tetracycline-resistant V. cholerae in some areas.
As with other diarrheal disease, a clean drinking water supply and adequate sewage disposal are fundamental to the prevention of cholera. As there is no animal reservoir, it should in theory be possible to eliminate the disease. However, carriage in humans, albeit for only a few weeks, occurs in 1–20% of previously infected patients, making eradication difficult to achieve.
Cholera vaccines are not recommended for most travellers
A killed whole-cell vaccine is available and is given parenterally, but is effective in only about 50% of those vaccinated, with protection lasting for only 3–6 months. It is no longer recommended by the World Health Organization (WHO) for travellers to cholera-endemic areas, although it may be required in certain countries. Oral vaccines (not available in the USA) appear to provide somewhat better protection.
Shigellosis
Symptoms of Shigella infection range from mild to severe depending upon the infecting species
Shigellosis is also known as bacillary dysentery (in contrast to amoebic dysentery; see below) because in its more severe form it is characterized by an invasive infection of the mucosa of the large intestine, causing inflammation and resulting in the presence of pus and blood in the diarrheal stool. However, symptoms range from mild to severe depending upon the species of Shigella involved and on the underlying state of health of the host. There are four species:
• Shigella sonnei causes most infections at the mild end of the spectrum.
• Shigella flexneri and S. boydii usually produce more severe disease.
• Shigella dysenteriae is the most serious.
Shigellosis is primarily a paediatric disease. When associated with severe malnutrition it may precipitate complications such as the protein deficiency syndrome ‘kwashiorkor’. Like V. cholerae, shigellae are human pathogens without an animal reservoir, but unlike the vibrios, they are not found in the environment, being spread from person to person by the faecal–oral route and less frequently by contaminated food and water. Shigellae appear to be able to initiate infection from a small infective dose (10–100 organisms) and therefore spread is easy in situations where sanitation or personal hygiene may be poor (e.g. refugee camps, nurseries, daycare centres and other residential institutions).
Shigella diarrhea is usually watery at first, but later contains mucus and blood
Shigellae attach to, and invade, the mucosal epithelium of the distal ileum and colon, causing inflammation and ulceration (Fig. 22.14). However, they rarely invade through the gut wall to the bloodstream. S. dysenteriae produce a (Shiga) toxin similar to that associated with enterohaemorrhagic E. coli (EHEC; see above), which can cause damage to the intestinal epithelium and glomerular endothelial cells, the latter leading to kidney failure (haemolytic-uraemic syndrome, HUS; see above).
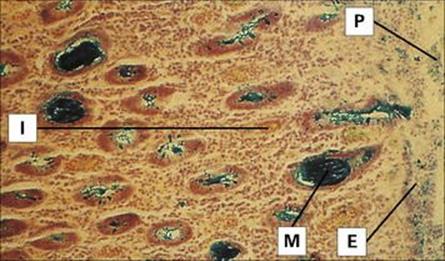
Figure 22.14 Shigellosis. Histology of the colon showing disrupted epithelium covered by pseudomembrane and interstitial infiltration. Mucin glands have discharged their contents and the goblet cells are empty. E, epithelium; I, interstitial infiltration; M, mucin in glands; P, pseudomembrane (colloidal iron stain).
(Courtesy of R.H. Gilman.)
The main features of shigella infection are summarized in Table 22.3. Diarrhea is usually watery initially, but later contains mucus and blood. Lower abdominal cramps can be severe. The disease is usually self-limiting, but dehydration can occur, especially in the young and elderly. Complications can be associated with malnutrition (see above).
Antibiotics should only be given for severe shigella diarrhea
Rehydration may be indicated. Antibiotics, especially those that also decrease intestinal motility, should not be given except in severe cases. Plasmid-mediated resistance is common, and antibiotic susceptibility tests should be performed on shigella isolates if treatment is required.
Education in personal hygiene and proper sewage disposal are important. Infected individuals may continue to excrete shigellae for a few weeks, but longer-term carriage is unusual; therefore, with adequate public health measures and no animal reservoir, the disease is potentially eradicable.
Other bacterial causes of diarrheal disease
The pathogens described in the previous sections are the major bacterial causes of diarrheal disease. Salmonella and Campylobacter infections and some types of E. coli infections are most often food-associated, whereas cholera is more often water-borne and shigellosis is usually spread by direct faecal–oral contact. Other bacterial pathogens that cause food-associated infection or food poisoning are described below.
V. parahaemolyticus and Yersinia enterocolitica are food-borne Gram-negative causes of diarrhea
V. parahaemolyticus is a halophilic (salt-loving) vibrio that contaminates seafood and fish. If these foods are consumed uncooked, diarrheal disease can result. The mechanism of pathogenesis is still unclear. Most strains associated with infection are haemolytic due to production of a heat-stable cytotoxin and have been shown to invade intestinal cells (in contrast to V. cholerae, which is non-invasive and cholera toxin, which is not cytotoxic).
The clinical features of infection are summarized in Table 22.3. The methods used for the laboratory diagnosis of V. parahaemolyticus infection are given in the Appendix (e.g. special media for cultivation). Prevention of infection depends upon cooking fish and seafood properly.
Yersinia enterocolitica is a member of the Enterobacteriaceae and is a cause of food-associated infection especially among infants and particularly in colder parts of the world. The reason for this geographic distribution is unknown, but it has been speculated that it is because the organism prefers to grow at temperatures of 22–25 °C. Y. enterocolitica is found in a variety of animal hosts including rodents, rabbits, pigs, sheep, cattle, horses and domestic pets. Transmission to humans from household dogs has been reported. The organism survives and multiplies, albeit more slowly, at refrigeration temperatures (48 °C) and has been implicated in outbreaks of infection associated with contaminated milk as well as other foods.
The mechanism of pathogenesis is unknown, but the clinical features of the disease result from invasion of the terminal ileum, necrosis in Peyer’s patches and an associated inflammation of the mesenteric lymph nodes (Fig. 22.15). The presentation, with enterocolitis and often mesenteric adenitis, can easily be confused with acute appendicitis, particularly in children. The clinical features are summarized in Table 22.3. The laboratory diagnosis is outlined in the Appendix. As with V. parahaemolyticus, an indication of a suspicion of yersinia infection is useful so that the laboratory staff can process the specimen appropriately.
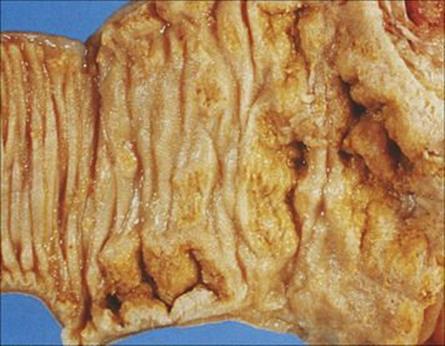
Figure 22.15 Yersinia enterocolitica infection of the ileum, showing superficial necrosis of the mucosa and ulceration.
(Courtesy of J. Newman.)
Clostridium perfringens and Bacillus cereus are spore-forming Gram-positive causes of diarrhea
The Gram-negative organisms described in the previous sections invade the intestinal mucosa or produce enterotoxins, which cause diarrhea. None of these organisms produces spores. Two Gram-positive species are important causes of diarrheal disease, particularly in association with spore-contaminated food. These are Clostridium perfringens and Bacillus cereus.
Cl. perfringens is associated with diarrheal diseases in different circumstances, and the pathogenesis is summarized in Figure 22.16:
• Enterotoxin-producing strains are a common cause of food-associated infection.
• Much more rarely, β-toxin-producing strains produce an acute necrotizing disease of the small intestine, accompanied by abdominal pain and diarrhea. This form occurs after the consumption of contaminated meat by people who are unaccustomed to a high-protein diet and do not have sufficient intestinal trypsin to destroy the toxin. It is traditionally associated with the orgiastic pig feasts enjoyed by the natives of New Guinea, but also occurred in people released from prisoner-of-war camps.
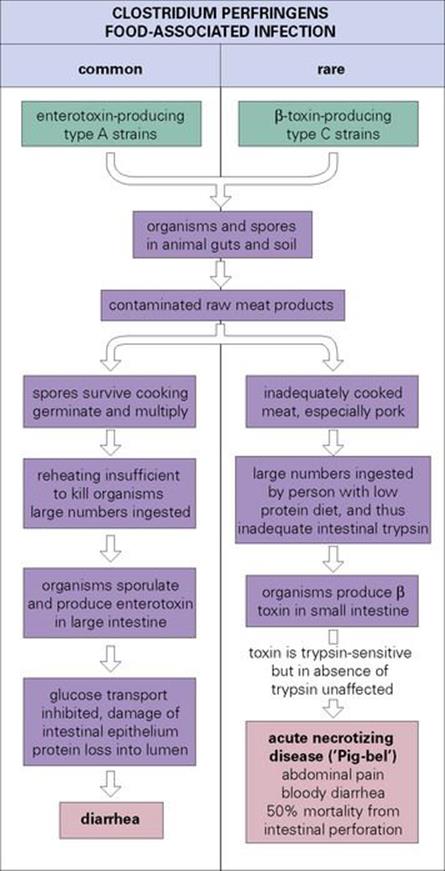
Figure 22.16 Clostridium perfringens is linked with two forms of food-associated infection. The common, enterotoxin-mediated infection (left) is usually acquired by eating meat or poultry that has been cooked enough to kill vegetative cells, but not spores. As the food cools, the spores germinate. If reheating before consumption is inadequate (as it often is in mass catering outlets), large numbers of organisms are ingested. The rare form associated with β-toxin-producing strains (right) causes a severe necrotizing disease.
The clinical features of the common type of infection are shown in Table 22.3. The laboratory investigation of suspected Cl. perfringens infection is outlined in the Appendix. The organism is an anaerobe and grows readily on routine laboratory media. Enterotoxin production can be demonstrated by a latex agglutination method.
Antibacterial treatment of Cl. perfringens diarrhea is rarely required. Prevention depends on thorough reheating of food before serving, or preferably avoiding cooking food too long before consumption.
Cl. perfringens is also an important cause of wound and soft tissue infections, as described in Chapter 26.
Bacillus cereus spores and vegetative cells contaminate many foods, and food-associated infection takes one of two forms:
• diarrhea resulting from the production of enterotoxin in the gut
• vomiting due to the ingestion of enterotoxin in food.
Two different toxins are involved, as illustrated in Figure 22.17. The clinical features of the infections are summarized in Table 22.3. Laboratory confirmation of the diagnosis requires specific media as described in the Appendix. The emetic type of disease may be difficult to assign to B. cereus unless the incriminated food is cultured.
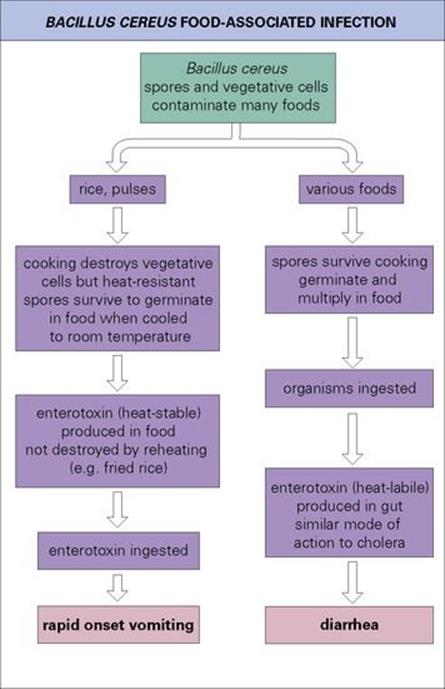
Figure 22.17 Bacillus cereus can cause two different forms of food-associated infection. Both involve toxins.
As with Cl. perfringens, prevention of B. cereus food-associated infection depends upon proper cooking and rapid consumption of food. Specific antibacterial treatment is not indicated.
Antibiotic-associated diarrhea – Clostridium difficile
Clostridium difficile infection is the most commonly diagnosed bacterial cause of hospital-acquired infectious diarrhea in resource-rich countries.
Treatment with broad-spectrum antibiotics can be complicated by antibiotic associated Cl. difficile diarrhea
All the infections described so far arise from the ingestion of organisms or their toxins. However, diarrhea can also arise from disruption of the normal gut flora. Even in the early days of antibiotic use, it was recognized that these agents affected the normal flora of the body as well as attacking the pathogens. For example, orally administered tetracycline disrupts the normal gut flora, and patients sometimes become recolonized not with the usual facultative Gram-negative anaerobes but with Staphylococcus aureus, causing enterocolitis, or with yeasts such as Candida. Soon after clindamycin was introduced for therapeutic use, it was found to be associated with severe diarrhea in which the colonic mucosa became covered with a characteristic fibrinous pseudomembrane (pseudomembranous colitis; Fig. 22.18). However, clindamycin is not the cause of the condition; it merely inhibits the normal gut flora and allows Cl. difficile to multiply. This organism is commonly found in the gut of children and to a lesser extent in adults, but can also be acquired from other patients in hospital by cross-infection. Cl. difficile is a spore former and survives in the environment as it is resistant to heat and acid, for example. The spores contaminate the environment and become vegetative bacteria that can be transmitted between patients on the wards. In common with other clostridia, Cl. difficile produces exotoxins, two of which have been characterized: one is a cytotoxin and the other an enterotoxin that cause haemostasis and tissue necrosis in the colon, resulting in diarrhea.
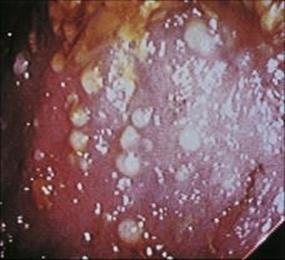
Figure 22.18 Antibiotic-associated colitis due to Clostridium difficile. Sigmoidoscopic view showing multiple pseudomembranous lesions.
(Courtesy of J. Cunningham.)
Toxin A and toxin B are encoded within a short chromosomal segment carried by pathogenic strains of Cl. difficile, referred to as the pathogenicity locus, as is a regulatory gene tcdC. There is also a binary toxin encoded by two chromosomal genes separate from the chromosomal pathogenicity locus. One gene mediates cell surface binding and intracellular translocation and the other causes cell death.
An emergent epidemic Cl. difficile variant strain called Cl. difficile 027 has been shown to produce more toxin A and toxin B than most hospital strains. A study reported that the binary toxin genes were associated with partial deletions in the tcdC gene that down-regulates the toxin A and B genes, and that severe Cl. difficile-associated diarrhea was significantly associated with them. Finally, Cl. difficile 027 was associated with much higher levels of toxins A and B than in other strains. This strain detected in the USA, Canada, the UK and other parts of Europe is not only highly transmissible but causes more severe disease in individuals in both hospitals and the community. It has been associated with higher case fatality rates, with some infected individuals requiring a colectomy and intensive care unit support, and has also been shown to be more resistant to the fluoroquinolone antibiotics than other strains.
Although initially associated with clindamycin, Cl. difficile diarrhea has since been shown to follow therapy with many other broad-spectrum antibiotics; hence the term antibiotic-associated diarrhea or colitis. The infection is often severe and may require treatment with the anti-anaerobic agent metronidazole, or with oral vancomycin. However, the emergence of vancomycin-resistant enterococci, probably originating in the gut flora, has led to the recommendation that oral vancomycin be avoided wherever possible (see Ch. 33).
Viral diarrhea
Over 3 million infants die of gastroenteritis each year, and viruses are the most common cause
Non-bacterial gastroenteritis and diarrhea are usually caused by viruses. Infection is seen in all parts of the world, especially in infants and young children (Fig. 22.19). Its impact is staggering, as in parts of Asia, Africa and Latin America more than 3 million infants die of gastroenteritis each year, and children may have a total of 60 days of diarrhea in each year. It has a major effect on nutritional status and growth. In the USA, about 200 000 children under 5 years of age are hospitalized each year because of infectious gastroenteritis.
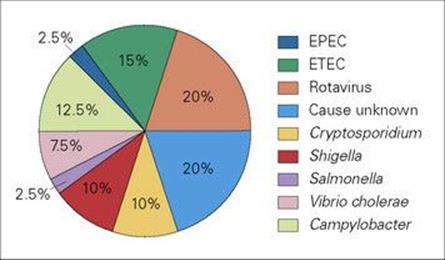
Figure 22.19 Diarrheal disease is a major cause of illness and death in children in resource-poor countries. This illustration shows the proportion of infections caused by different pathogens. Note that in as many as 20% of infections, a cause is not identified, but many of these are likely to be viral. EPEC, enteropathogenic E. coli; ETEC, enterotoxigenic E. coli.
(Data from the WHO.)
Although viruses appear to be the most common causes of gastroenteritis in infants and young children, viral gastroenteritis is not distinguishable clinically from other types of gastroenteritis. The viruses are specific to humans, and infection follows the general rules for faecal–oral transmission. Oral transmission of non-bacterial gastroenteritis was first demonstrated experimentally in 1945, but it was not until 1972 that viral particles were identified in faeces by electron microscopy. It has been difficult or impossible to cultivate most of these viruses in cell culture.
Rotaviruses
These are morphologically characteristic viruses (Fig. 22.20) named after the Latin word rota meaning a wheel, with a genome consisting of 11 separate segments of double-stranded RNA. Different rotaviruses infect the young of many mammals, including children, kittens, puppies, calves, foals and piglets, but it is thought that viruses from one host species occasionally cross-infect another. There are at least two human serotypes.
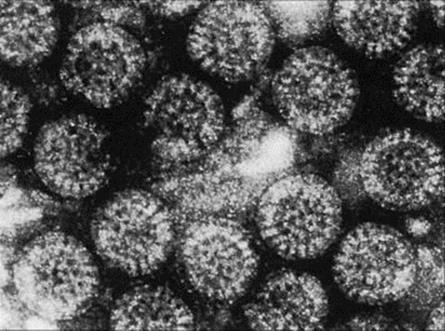
Figure 22.20 Rotavirus. The virus particles (65 nm in diameter) have a well-defined outer margin and capsules radiating from an inner core to give the particle a wheel-like (hence ‘rota’) appearance.
(Courtesy of J.E. Banatvala.)
Replicating rotavirus causes diarrhea by damaging transport mechanisms in the gut
The incubation period is 1–2 days. After virus replication in intestinal epithelial cells there is an acute onset of vomiting, which is sometimes projectile, and diarrhea which lasts from 4 to 7 days. The replicating virus damages transport mechanisms in the gut, and loss of water, salt and glucose causes diarrhea (Fig. 22.21). Infected cells in the intestine are destroyed, resulting in villous atrophy. The villi, long finger-like projections, become flattened, resulting in the loss of both the surface area for absorption and the digestive enzymes, and raised osmotic pressure in the gut lumen causes diarrhea. There is no inflammation or loss of blood. Exceedingly large numbers of virus particles, 1010–1011/g, appear in the faeces. For unknown reasons, respiratory symptoms such as cough and coryza are quite common. The disease is more severe in infants in resource-poor countries.
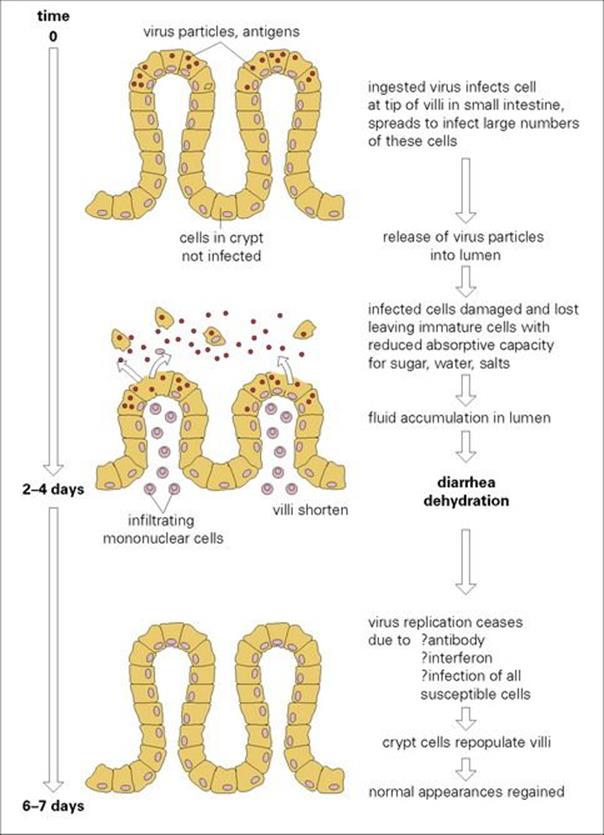
Figure 22.21 The pathogenesis of rotavirus diarrhea. This may differ with other viral infections of the gastrointestinal tract.
Infection is most common in children under 2 years of age, and has a seasonal pattern, being most frequent in the cooler months of the year in temperate climates. IgA antibodies in colostrum give protection during the first 6 months of life. Outbreaks are sometimes seen in nurseries. Older children are less susceptible to infection, nearly all of them having developed antibodies, but occasional infections occur in adults.
Rotaviruses are well-adapted intestinal infectious agents. As few as 10 ingested particles can cause infection, and by generating diarrhea laden with enormous quantities of infectious particles, together with their stability in the environment, these organisms have ensured their continued transmission and survival.
Rotavirus infection is confirmed by viral RNA or antigen detection
Laboratory diagnosis is generally not available in resource-poor countries, but during the acute stages, the diagnosis is made by detecting viral RNA or antigen using PCR or ELISA methods, respectively (see Ch. 32). The characteristic 65-nm particles can be seen in faecal samples by electron microscopy. They show cubic symmetry and an outer capsid coat arranged like the spokes of a wheel (Fig. 22.20).
Fluid and salt replacement can be life-saving in rotavirus diarrhea
Dehydration occurs readily in infants, and fluid and salt replacement orally or intravenously can be life-saving. There are no antiviral agents available, but a variety of live attenuated oral vaccines have undergone successful trials. In 2006, the US Food and Drug Administration (FDA) announced the approval of a live, oral vaccine for use in preventing rotavirus gastroenteritis in infants.
Other viruses
Other viruses causing diarrhea in humans include caliciviruses, astroviruses, adenoviruses and coronaviruses
Caliciviruses are 27 nm in diameter, single-stranded RNA viruses. They include the noroviruses, previously known as the small round structured viruses (SRSV) or Norwalk-like viruses (NLV) that cause ‘winter vomiting disease’ and sapoviruses. They have not yet been cultivated in vitro and cause gastroenteritis when fed to adult volunteers. One of the first identified norovirus outbreaks was in a school in Norwalk, Ohio, in 1969. Infection is common in older children and adults. These viruses are highly infectious, spread rapidly and nosocomial infection is common. The incubation period is 12–72 h. In up to 50% of cases there may be chills, headache, myalgia or fever as well as nausea, abdominal pain, vomiting and diarrhea. Recovery may occur within 24–48 h but may take longer. Noroviruses bind to cell surface carbohydrates of the ABH histo-blood group antigens and some strains have different binding affinities for different patterns of these antigens. In addition, these antigens are expressed to varying degrees in different individuals, resulting in some people being resistant to infection with specific norovirus strains. Laboratory diagnosis, important in outbreaks and for epidemiologic studies, is usually by PCR, electron microscopy or ELISA. Viruses in this group are often implicated in diarrhea associated with food- or water-borne routes occurring after eating sewage-contaminated shellfish such as cockles or mussels. In particular, noroviruses are a major cause of gastroenteritis in healthcare settings and many outbreaks have been reported in crowded environments such as cruise ships. Noroviruses show a high level of variability, resulting in both limited cross-protection between strains and reduced immunity in the population. In addition, due to this diversity, diagnostic assays have to be modified in order to optimize detection, and vaccine design either has to involve a cross-protective component or the development of a multivalent vaccine.
Astroviruses are 28-nm single-stranded RNA viruses of which five serotypes are known and have characteristic five- or six-pointed star patterns. Most infections occur in childhood and are mild. Adenoviruses are unenveloped, 70–80-nm double-stranded DNA viruses of which types 40 and 41 are associated with gastroenteritis. Types 40 and 41 can only be grown in specialized cell culture lines. They are second to rotaviruses as a cause of acute diarrhea in young children in temperate climates. The role of coronaviruses and human bocavirus infections in causing gastroenteritis is uncertain.
Although outbreaks of gastroenteritis often have a viral aetiology it may be difficult to be sure about the exact role of a given virus when it is identified in faeces, as there are a number of viruses that replicate in the gastrointestinal tract, enteroviruses, for example, which are not associated with acute diarrheal illness.
Food poisoning
In this chapter, the term ‘food poisoning’ is restricted to the diseases caused by toxins elaborated by contaminating bacteria in food before it is consumed (see above). The emetic toxin of B. cereus fits this definition, as do the diseases associated with the consumption of Staph. aureusenterotoxin and Cl. botulinum toxin.
Staphylococcus aureus
Eight different enterotoxins are produced by different strains of Staph. aureus
Twenty two serologically distinct enterotoxins have been reported to be produced by strains of Staph. aureus, the best understood of which are enterotoxins A–E (Table 22.4). All are heat stable and resistant to destruction by enzymes in the stomach and small intestine. Their mechanism of action is incompletely understood; however, similar to the TSST-1 toxin of toxic shock syndrome (see Ch. 26), they generally behave as superantigens (see Ch. 16), binding to major histocompatibility complex (MHC) class II molecules, which results in T-cell stimulation. Their effect on the central nervous system results in severe vomiting within 3–6 h of consumption. Diarrhea is not a feature, and recovery within 24 h is usual.
Table 22.4 Staphylococcal enterotoxins
|
Enterotoxin |
||
|
A |
Most commonly associated with food poisoning |
|
|
B |
Associated with staphylococcal enterocolitis (rare) |
|
|
C |
Rare |
|
|
D |
Second most common |
|
|
Alone or in combination with A |
||
|
E |
Rare |
|
|
TSST-1 |
Toxic shock syndrome toxin, not food-associated |
|
Staphylococcus aureus produces at least eight immunologically distinct enterotoxins, the most important of which are listed here. Strains may produce one or more of the toxins simultaneously. Enterotoxin A is by far the most common in food-associated disease.
Up to 50% of Staph. aureus strains produce enterotoxin, and food (especially processed meats) is contaminated by human carriers. The bacteria grow at room temperature and release toxin. Subsequent heating may kill the organisms, but the toxin is stable. Often there are no viable organisms detectable in the food consumed, but enterotoxin can be detected by a latex agglutination test.
Botulism
Exotoxins produced by Cl. botulinum cause botulism
Botulism is a rare but serious disease caused by the exotoxin of Cl. botulinum. The organism is widespread in the environment, and spores can be isolated readily from soil samples and from various animals including fish. Seven serologically distinct toxins have been identified, but only four, A, B, E, and less frequently F, are associated with human disease. While not destroyed by digestive enzymes, the toxins are inactivated after 30 min at 80 °C. The toxins are ingested in food (often canned or reheated) or produced in the gut after ingestion of the organism; they are absorbed from the gut into the bloodstream and then reach their site of action, the peripheral nerve synapses. The action of the toxin is to block neurotransmission (see Ch. 17).
Infant botulism is the most common form of botulism
There are three forms of botulism:
1. food-borne botulism
2. infant botulism
3. wound botulism.
In food-borne botulism, toxin is elaborated by organisms in food, which is then ingested. In infant and wound botulism, the organisms are, respectively, ingested or implanted in a wound, and multiply and elaborate toxin in vivo. Infant botulism has been associated with feeding babies honey contaminated with Cl. botulinum spores.
The clinical disease is the same in all three forms and is characterized by flaccid paralysis leading to progressive muscle weakness and respiratory arrest. Intensive supportive treatment is urgently required, and complete recovery may take many months. Improvements in supportive care have reduced the mortality from around 70% to approximately 10%, but the disease, although rare, remains life-threatening. In addition, since botulinum toxin is one of the most potent biological toxins known, there is concern regarding its potential use as an agent of biowarfare.
Laboratory diagnosis of botulism involves injecting faecal and food samples into mice
Laboratory diagnosis depends largely upon demonstrating the presence of toxin or culturing the bacteria. However, a bioassay may need to be used if serum is available, whereby the serum would be injected into mice that have been protected with botulinum antitoxin or left unprotected. Culture of faeces or wound exudate for Cl. botulinum as well as toxin detection by polymerase chain reaction (PCR)-based assays for toxin sequences and ELISA (see Ch. 32) tests for functional toxin activity.
Polyvalent antitoxin is recommended as an adjunct to intensive supportive therapy for botulism
Since botulinum toxins are antigenic, they can be inactivated and used to produce antitoxin in animals. When botulism is suspected, antitoxin should be promptly administered along with supportive care, which may include mechanical ventilation (due to difficulty in breathing) and intravenous and nasogastric nutritional support (due to difficulty in swallowing). Antibiotics are generally used only for treatment of secondary infections.
It is not practicable to prevent food becoming contaminated with botulinum spores, so prevention of disease depends upon preventing the germination of spores in food by:
• maintaining food at an acid pH
• storing food at < 4°C
• destroying toxin in food by heating for 30 min at 80 °C.
Helicobacter pylori and gastric ulcer disease
Helicobacter pylori is associated with most duodenal and gastric ulcers
It is now well established that the Gram-negative spiral bacterium H. pylori is associated with over 90% of duodenal ulcers and 70–80% of gastric ulcers (Fig. 22.22). H. pylori does not appear to play a role in gastroesophageal reflux disease (GERD) or non-ulcer dyspepsia, most commonly presenting with persistent or recurrent pain in the upper abdomen in the absence of structural evidence of disease. Diagnosis may be made on the basis of histologic examination of biopsy specimens, although the non-invasive urea breath test (H. pylori produces large amounts of urease) is the most rapid means of detecting the organism’s presence. Faecal Helicobacter pylori antigen testing is another non-invasive test. H. pylori can be cultured in the laboratory, but is difficult to grow.
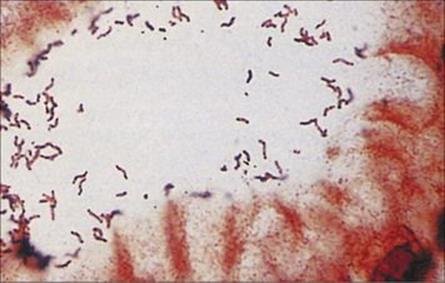
Figure 22.22 Helicobacter pylori gastritis. Silver stain showing numerous spiral-shaped organisms adhering to the mucosal surface.
(Courtesy of A.M. Geddes.)
The mechanism of pathogenicity is still being elucidated but involves a number of virulence factors, including a cytotoxin, acid-inhibiting protein, adhesins, urease (which aids survival in the acidic environment) and other factors which disrupt the gastric mucosa. Eradication of H. pylori to promote the remission and healing of ulcers requires combination therapy such as a proton pump inhibitor and two antibiotics such as clarithromycin and amoxicillin (see Ch. 33). However, studies suggesting that H. pylori may actually provide protection from some oesophageal and gastric cancers have led to active discussion regarding whether the organism should be eliminated from asymptomatic patients. It has been postulated that H. pylori interferes with the secretion of gastric acid that is associated with GERD, which in turn may precede oesophageal cancer. It is particularly confusing as it has also been associated with the development of stomach ulcers and cancer.
The interrelationship between H. pylori and gastric disease is thus complex and remains to be further clarified.
Parasites and the gastrointestinal tract
Many species of protozoan and worm parasites live in the gastrointestinal tract, infecting some 3.5 billion people worldwide. Only a few are a frequent cause of serious pathology (Fig. 22.23) and these will form the focus of this part of the chapter.
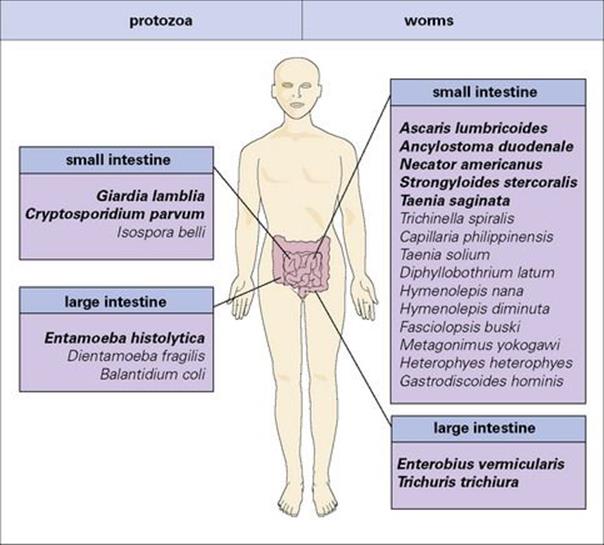
Figure 22.23 Gastrointestinal parasites of humans. The majority of these infections are found in resource-poor countries, but all species also occur in the resource-rich world and some have come to prominence because of their association with AIDS. The most important parasite species are highlighted in bold type.
Transmission of intestinal parasites is maintained by the release of life cycle stages in faeces
The different life cycle stages include cysts, eggs and larvae. In most cases new infections depend either directly or indirectly on contact with faecally derived material, infection rates therefore reflecting standards of hygiene and levels of sanitation. In general, the stages of protozoan parasites passed in faeces are either already infective or become infective within a short time. These parasites are therefore usually acquired by swallowing infective stages in faecally contaminated food or water. Worm parasites, with two major exceptions (pinworm and the dwarf tapeworm), produce eggs or larvae that require a period of development outside the host before they become infective. Transmission routes are more complex here:
• Some species are acquired through food or water contaminated with infective eggs or larvae, or are picked up directly via contaminated fingers.
• Some have larvae that can actively penetrate through the skin, migrating eventually to the intestine.
• Others are acquired by eating animals or animal products containing infective stages.
The symptoms of intestinal infection range from very mild, through acute or chronic diarrheal conditions associated with parasite-related inflammation, to life-threatening diseases caused by spread of the parasites into other organs of the body. Most infections fall into the first of these categories.
Protozoan infections
Three species are of particular importance:
• Entamoeba histolytica
• Giardia intestinalis
• Cryptosporidium parvum.
All three can give rise to diarrheal illnesses, but the organisms have distinctive features that allow a differential diagnosis to be made quite easily (Fig. 22.24). Other intestinal protozoa of concern, particularly in immunosuppressed patients, include Cyclospora cayetanensis, Isospora belli and the microsporidia.
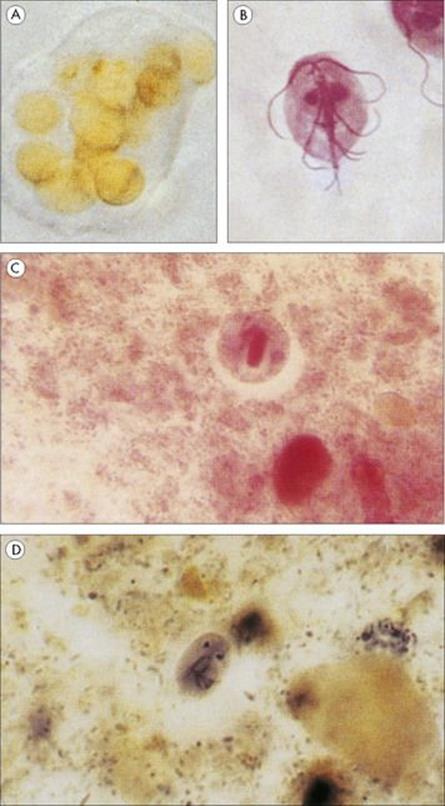
Figure 22.24 Protozoan infections of the gastrointestinal tract. (A) Entamoeba histolytica. Trophozoite found in the acute stage of the disease, which often contains ingested red blood cells. (B) Giardia intestinalis trophozoite associated with acute infection in humans. (Courtesy of D.K. Banerjee.) (C) Cyst of E. histolytica, with only one of the four nuclei visible. The broad chromatid bar is a semicrystalline aggregation of ribosomes. (H&E stain). (D) Oval cyst of G. intestinalis showing two of the four nuclei. (Iron haematoxylin stain.)
(Courtesy of R. Muller and J.R. Baker.)
Entamoeba histolytica
Entamoeba histolytica infection is particularly common in subtropical and tropical countries
For many years, it was considered that infections with E. histolytica could be asymptomatic or pathogenic, with dysentery a key symptom when the amoebae invaded the mucosa. Two species are involved: E. histolytica being invasive and E. dispar being non-invasive. E. histolytica occurs worldwide, but is most often found in subtropical and tropical countries, where the prevalence may exceed 50%. The trophozoite stages of the amoebae live in the large intestine on the mucosal surface. Reproduction of these stages is by simple binary fission, and there is periodic formation of resistant encysted forms, which pass out of the body. These cysts can survive in the external environment and act as the infective stages. Infection occurs when food or drink is contaminated either by infected food handlers or as a result of inadequate sanitation. Transmission can also take place as a result of anal sexual activity. The cysts pass intact through the stomach when swallowed and excyst in the small intestine, each giving rise to four progeny. These adhere to the epithelial cells and damage them by phagocytosis and cytolysis. They can invade the mucosa and feed on host tissues including red blood cells, giving rise to amoebic colitis.
E. histolytica infection may cause mild diarrhea or severe dysentery
Infections with E. dispar are asymptomatic. Invasion of the mucosa by E. histolytica may produce small localized superficial ulcers or involve the entire colonic mucosa with the formation of deep confluent ulcers (Fig. 22.25). The former causes a mild diarrhea, whereas more severe invasion leads to ‘amoebic dysentery’, which is characterized by mucus and blood in the stools. Dysenteries of amoebic and bacillary origin can be distinguished by a number of features (Table 22.5).
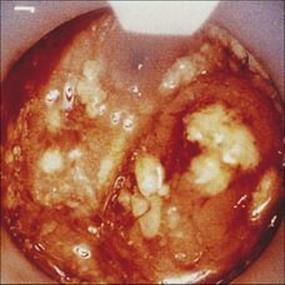
Figure 22.25 Amoebic colitis. Sigmoidoscopic view showing deep ulcers and overlying purulent exudate.
(Courtesy of R.H. Gilman.)
Table 22.5 Features of bacillary and amoebic dysentery
|
Bacillary |
Amoebic |
|
|
Organism |
Shigella |
Entamoeba |
|
Polymorphs and macrophages in stool |
Many |
Few |
|
Eosinophils and Charcot–Leyden crystals in stool |
Few or absent |
Often present |
|
Organisms in stool |
Many |
Few |
|
Blood and mucus in stool |
Yes |
Yes |
Complications include perforation of the intestine, leading to peritonitis, and extraintestinal invasion. Trophozoites can spread via the blood to the liver, with the formation of an abscess, and may secondarily extend to the lung and other organs. Rarely, abscesses spread directly and involve the overlying skin.
E. histolytica infection can be diagnosed in asymptomatic patients from the presence of characteristic four-nucleate cysts in the stool
These cysts may be infrequent in light infections, and repeated stool examination is necessary. Care must be taken to differentiate E. histolytica from other non-pathogenic species that might be present (Fig. 22.26). Trophozoites can be found in cases of dysentery (when the stools are loose and wet), but they are fragile and deteriorate rapidly; specimens should therefore be preserved before examination. ELISA tests are available, as is a triage panel assay that can distinguish between E. histolytica/E. dispar Cryptosporidium parvum and Giardia intestinalis. Differentiation of E. histolytica from E. dispar requires immunological tests or PCR.
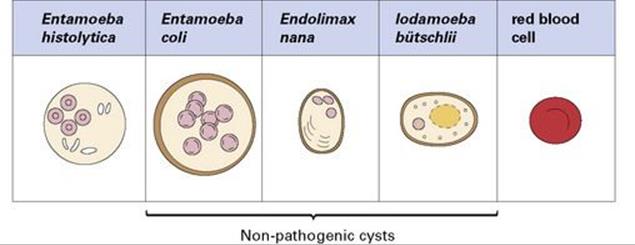
Figure 22.26 Characteristics of cysts (size and number of nuclei) are used to differentiate pathogenic from non-pathogenic protozoa. A red blood cell is shown for comparison.
Acute E. histolytica infection can be treated with metronidazole or tinidazole
Recovery from infection is usual, and there is some immunity to reinfection. Metronidazole or tinidazole kill amoebic trophozoites in both intestinal and extraintestinal sites of infection and result in rapid clinical improvement, but relapse of the infection may occur unless a second antiamoebic agent is given to eradicate amoebae from the gut lumen. Examples are diloxanide furoate or paromomycin. Prevention of amoebiasis in the community requires the same approaches to hygiene and sanitation as those adopted for bacterial infections of the intestine.
Giardia intestinalis
Giardia was the first intestinal microorganism to be observed under a microscope. It was discovered by Anton van Leeuwenhoek in 1681, using the microscope he had invented to examine specimens of his own stool. It has a global distribution and is a frequent cause of traveller’s diarrhea.Giardia is the most commonly diagnosed intestinal parasite in the USA, having been detected in both drinking and recreational water. There is confusion over nomenclature, and the species infecting humans is also commonly referred to as G. lamblia, and sometimes as G. duodenalis(human).
Like Entamoeba, Giardia has only two life cycle stages
The two life cycle stages are the flagellate (four pairs of flagella) binucleate trophozoite and the resistant four-nucleate cyst. The trophozoites live in the upper portion of the small intestine, adhering closely to the brush border of the epithelial cells by specialized attachment regions (Fig. 22.27). They divide by binary fission and can occur in such numbers that they cover large areas of the mucosal surface. Cyst formation occurs at regular intervals, each cyst being formed as one trophozoite rounds up and produces a resistant wall. Cysts pass out in the stools and can survive for several weeks under optimum conditions. Infection occurs when the cysts are swallowed, usually as a result of drinking contaminated water. The minimum infective dose is very small: 10–25 cysts.
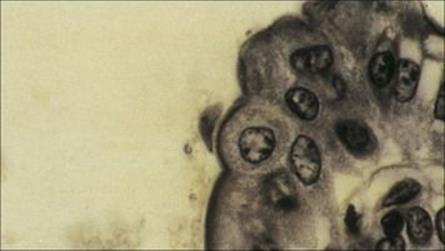
Figure 22.27 Trophozoite of Giardia intestinalis attached to the mucosal surface of the small intestine. (Iron haematoxylin stain.)
(Courtesy of R. Muller and J.R. Baker.)
Epidemics of giardiasis have occurred when public drinking supplies have become contaminated, but smaller outbreaks have been traced to drinking from rivers and streams that have been contaminated by wild animals. Apart from water-borne transmission, Giardia can be passed from person to person, especially within families, with food-borne transmission being rare. Giardia may also be transmitted sexually among homosexual men. The genus Giardia is widely distributed in mammals, and there is suggestive evidence for cross-infection between certain animal hosts (e.g. beaver) and humans. Much of this is circumstantial, but case reports provide more direct evidence.
Mild Giardia infections are asymptomatic, more severe infections cause diarrhea
The diarrhea may be:
• self-limiting, with 7–10 days being the usual course
• chronic, and develop into a serious condition, particularly in patients with deficient or compromised immunologic defences.
It is thought to arise from inflammatory responses triggered by the damaged epithelial cells and from interference with normal absorptive processes. Characteristically, the stools are loose, foul-smelling and often fatty.
Diagnosis of Giardia infection is based on identifying cysts or trophozoites in the stool
Repeated examination is necessary in light infections, when concentration techniques improve the chances of finding cysts. Duodenal intubation or the use of recoverable swallowed capsules and threads may aid in obtaining trophozoites directly from the intestine. Alternatives to microscopic methods are increasingly available, including faecal antigen ELISA tests with good specificity, immunochromatographic tests in cassette form, and PCR in some centres.
Giardia infection can be treated with a variety of drugs
Metronidazole and tinidazole are commonly used. Nitazoxanide or albendazole are alternatives and mepacrine hydrochloride is sometimes used. Community measures for prevention include the usual concerns with hygiene and sanitation, and improved treatment of drinking water supplies (largely filtration and chlorination) where these are suspected as a source. Care in drinking from potentially contaminated natural waters is also indicated.
Cryptosporidium hominis and Cryptosporidium parvum
The protozoan genus Cryptosporidium is widely distributed in many animals
Awareness of Cryptosporidium as an important cause of diarrhea in humans was established during the early years of the AIDS epidemic, although similar parasites were known to be widely distributed in many animals. There are two major species, C. hominis causing human infection, andC. parvum primarily infecting animals (including cattle), though cross-infection to humans does occur. The parasite has a complex life cycle, going through both asexual and sexual phases of development in the same host. Transmission requires ingestion of a minimum of 10 or so of the resistant oocyst stage (4–5 mm in diameter) in faecally contaminated material (Fig. 22.28). In the small intestine, the cyst releases infective sporozoites, which invade the epithelial cells, remaining closely associated with the apical plasma membrane. Here they form schizonts, which divide to release merozoites, and these then re-invade further epithelial cells. Eventually, a sexual phase occurs and oocysts are released. Transmission probably occurs most often via drinking water contaminated by oocysts, either from other humans or from animals. In 1993, Cryptosporidiumcaused a massive outbreak of watery diarrhea affecting 403 000 people in Milwaukee, USA. It was transmitted through the public water supply and probably originated from cattle.
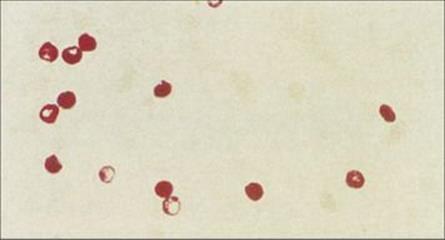
Figure 22.28 Cryptosporidium oocysts in faecal specimen.
(Courtesy of S. Tzipori.)
Cryptosporidial diarrhea ranges from moderate to severe
Symptoms of infection with Cryptosporidium range from moderate diarrhea to more severe profuse diarrhea that is self-limiting in immunocompetent individuals (lasting 15–40 days), but can become chronic in immunocompromised patients. Cryptosporidiosis is a common infection in people with AIDS. In individuals with CD4+ T-cell counts < 100/mm3 diarrhea is prolonged, may become irreversible, and can be life-threatening.
Routine faecal examinations are inadequate for diagnosing cryptosporidial diarrhea
Concentration techniques and special staining (e.g. modified Ziehl–Neelsen stain) are necessary to recover and identify the oocysts. Direct immunofluorescence and antigen detection ELISA tests are also used. PCR is available in reference centres and is becoming more widely available.
In general, only immunocompromised patients need antiparasitic treatment for cryptosporidial diarrhea
Highly active antiretroviral therapy (HAART) in individuals with AIDS infected with Cryptosporidium has been reported to improve the diarrhea symptoms. This may be due to the protease inhibitors used in combination therapy interfering directly with the cryptosporidial proteases involved in the protozoal life cycle. In addition, HAART results in lowering of the HIV load and immune reconstitution. Paromomycin reduces oocyst output but does not clear infection. Nitazoxanide is effective in HIV-negative patients but is only partially active in those co-infected with HIV. Public health measures are similar to those outlined for controlling giardiasis, although Cryptosporidium is more resistant to chlorination. Some water treatment facilities deploy an additional ozonation step to inactivate cryptosporidia.
Cyclospora, Isospora and the Microsporidia
Cyclospora, like Isospora belli and Cryptosporidium, is a coccidian parasite, whose life cycle stages take place in epithelial cells of the mucosa. Cyclospora and Isospora have only been found in humans, unlike other coccidia that are zoonotic.
Cyclospora cayetanensis, identified in 1994, is one of the causes of diarrhea in travellers, but it can also be acquired from contaminated imported food; for example, Guatemalan raspberries were thought to be the cause of five diarrheal outbreaks in the USA in the years 1995 to 2000. Diarrhea can be prolonged and is severe in immunosuppressed individuals. Trimethoprim-sulphamethoxazole (co-trimoxazole) treatment is effective. Ciprofloxacin is partially effective.
AIDS patients infected with Isospora belli may show particularly severe symptoms, persistent diarrhea causing weight loss and even death. Treatment is with co-trimoxazole.
Infections with microsporidia, an unusual group, have also become recognized as a cause of diarrhea in AIDS and other immunosuppressed patients. Enterocytozoon bieneusi is the commonest cause, although Encephalitozoon intestinalis also occurs. Transmission appears to be direct. Albendazole treatment is effective against Encephalitozoon intestinalis but has disappointing activity against Enterocytozoon bieneusi. Where possible, immune reconstitution is the mainstay of treatment.
‘Minor’ intestinal protozoa
The human intestine may harbour a large number of protozoa, many of which appear to be quite harmless. Some have a questionable role in disease: these include Blastocystis hominis, Dientamoeba fragilis and Sarcocystis hominis.
Worm infections
The most important intestinal worms clinically are the nematodes known as ‘soil-transmitted helminths’
Soil-transmitted helminths fall into two distinct groups:
• Ascaris lumbricoides (large roundworm) and Trichuris trichiura (whipworm), in which infection occurs by swallowing the infective eggs
• Ancylostoma duodenale and Necator americanus (hookworms) and Strongyloides stercoralis, which infect by active skin penetration by infective larvae which then undertake a systemic migration through the lungs to the intestine.
With the exception of Trichuris all the soil-transmitted nematodes inhabit the small bowel.
The pinworm or threadworm Enterobius vermicularis is probably the commonest intestinal nematode in resource-rich countries and is the least pathogenic. The females of this species, which live in the large bowel, release infective eggs onto the perianal skin. This causes itching, and transmission usually occurs directly from contaminated fingers, but the eggs are also light enough to be carried in dust.
The soil-transmitted helminths are commonest in the warmer resource-poor countries. About one-quarter of the world’s population carry these worms, children being the most heavily infected section of the population. Transmission is favoured where there is inadequate disposal of faeces, contamination of water supplies, use of faeces (night-soil) as fertilizer, or low standards of hygiene (see below). Vast numbers of eggs are released in the lifetime of each female (tens of thousands by Trichuris and Ancylostoma and hundreds of thousands by Ascaris).
Life cycle and transmission
Female Ascaris and Trichuris lay thick-shelled eggs in the intestine, which are expelled with faeces and hatch after being swallowed by another host
The thick-shelled eggs of Ascaris and Trichuris are shown in Figure 22.29. The eggs require incubation for several days at optimum conditions (warm temperature, high humidity) for the infective larvae to develop. Once this occurs, the eggs remain infective for many weeks or months, depending upon the local microclimate. After being swallowed, the eggs hatch in the intestine, releasing the larvae. Those of Ascaris penetrate the gut wall and are carried in the blood through the liver to the lungs, climbing up the bronchi and trachea before being swallowed and once again reaching the intestine. The adult worms live freely in the gut lumen, feeding on intestinal contents. In contrast, Trichuris larvae remain within the large bowel, penetrating into the epithelial cell layer, where they remain as they mature.
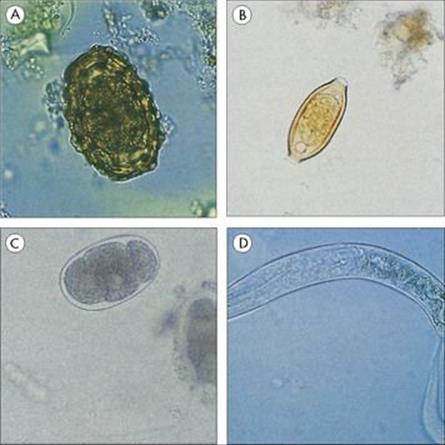
Figure 22.29 Eggs and larvae of intestinal nematodes passed in faeces. (A) Egg of Ascaris (fertile). (B) Egg of Trichuris. (C) Egg of hookworm. The embryo continues to divide in the faecal sample and may be at the 16- or 32-cell stage by the time the sample is examined. (D) Larva ofStrongyloides stercoralis.
(Courtesy of J.H. Cross.)
Adult female hookworms lay thin-shelled eggs that hatch in the faeces shortly after leaving the host
A hookworm egg is shown in Figure 22.29. The larvae of these hookworms (A. duodenale and N. americanus) feed on bacteria until infective, and then migrate away from the faecal mass. Infection takes place when larvae come into contact with unprotected skin (or additionally, in the case of Ancylostoma, are swallowed). They penetrate the skin, migrate via the blood to the lungs, climb the trachea and are swallowed. Adult worms attach by their enlarged mouths to the intestinal mucosa, ingest a plug of tissue, rupture capillaries and suck blood.
The adult female Strongyloides lays eggs that hatch in the intestine
The life cycle of Strongyloides is similar to that of hookworms, but shows some important differences. The adult worm exists as a parthenogenetic female that lays eggs into the mucosa. These eggs hatch in the intestine and the released larvae usually pass out in the faeces (Fig. 22.29). Development outside the host can follow the hookworm pattern, with the direct production of skin-penetrating larvae, or may be diverted into the production of a complete free-living generation, which then produces infective larvae. Under certain conditions, and particularly when the host is immunocompromised, Strongyloides larvae can re-invade before they are voided in the faeces. This process of autoinfection can give rise to the severe clinical condition known as disseminated strongyloidiasis, also known as hyperinfection, which is often complicated by Gram-negative bacterial septicaemia. All soil-transmitted helminths are relatively long-lived (several months to years), but authenticated cases show that Strongyloides infections can persist for more than 30 years, presumably through continuous internal autoinfection.
Clinical features
In most individuals, worm infections produce chronic mild intestinal discomfort rather than severe diarrhea or other conditions. Infections may lead to hypersensitivity responses and can also reduce responses to vaccination. Each parasite has a number of characteristic pathologic conditions linked with it.
Large numbers of adult Ascaris worms can cause intestinal obstruction
The migration of Ascaris larvae through the lungs can cause severe respiratory distress (pneumonitis), and this stage is often associated with pronounced eosinophilia. Intestinal stages of infection can cause abdominal pain, nausea and digestive disturbances. In children with a suboptimal nutritional intake, these disturbances can contribute to clinical malnutrition. Large numbers of adult worms can cause a physical blockage in the intestine, and this may also occur as worms die following chemotherapy. Intestinal worms tend to migrate out of the intestine, often up the bile duct, causing cholangitis. Perforation of the intestinal wall can also occur. Worms have occasionally been reported in unusual locations, including the orbit of the eye and the (male) urethra. Ascaris is highly allergenic, and infections often give rise to symptoms of hypersensitivity which may persist for many years after the infection has been cleared.
Moderate to severe Trichuris infection can cause chronic diarrhea
As with all intestinal worms, children are the members of the community most heavily infected with Trichuris. Although previously regarded as of little clinical significance, research has shown that moderate to heavy infections in children can cause a chronic diarrhea (Fig. 22.30), reflected in impaired nutrition and retarded growth. Occasionally, heavy infections lead to prolapse of the rectum.
Figure 22.30 Trichuriasis in a healthy, infected, child. Proctoscopic view showing numerous adult Trichuris trichiura attached to the intestinal mucosa.
(Courtesy of R.H. Gilman.)
Hookworm disease can result in iron-deficiency anaemia
Invasion of hookworm larvae through the skin and lungs can cause a dermatitis and pneumonitis, respectively. The blood-feeding activities of the intestinal worms can lead to an iron-deficiency anaemia if the diet is inadequate. Heavy infections cause a marked debility and growth retardation.
Strongyloidiasis can be fatal in immunosuppressed people
Heavy intestinal infection with strongyloidiasis causes persistent and profuse diarrhea with dehydration and electrolyte imbalance. Profound mucosal changes can also lead to a malabsorption syndrome, which is sometimes confused with tropical sprue. People with diseases that suppress immune function, such as AIDS and cancer, or who are being treated with immunosuppressive drugs are susceptible to the development of disseminated strongyloidiasis. Invasion of the body by many thousands of autoinfective larvae can be fatal. Gram-negative bacterial septicaemia or meningitis can ensue.
The most common sign of pinworm (threadworm) infection is anal pruritus. Occasionally, this is accompanied by mild diarrhea. Migrating worms sometimes invade the appendix and have been linked with appendicitis. Invasion of the vagina has been reported in female children.
Laboratory diagnosis
All five of the soil-transmitted species can be diagnosed by finding eggs or larvae in the fresh stool, and direct smears or concentration techniques can be used. Acute infections with Ascaris, hookworms and Strongyloides are often accompanied by a marked blood eosinophilia. Although this is not diagnostic, it is a strong indicator of worm infection. Enzyme immunoassays can be used to detect Strongyloides antibody and has a 90% sensitivity of detection. However, there is cross-reactivity with IgG made against other nematode infections and one cannot determine whether the infection occurred recently or in the past.
The eggs of Ascaris, Trichuris and hookworms are characteristic
These eggs are shown in Figure 22.29 and are easily recognizable. Identification of the species of hookworm requires culture of the stool to allow the eggs to hatch and the larvae to mature into the infective third stage. The presence of adult Ascaris can sometimes be confirmed directly by radiography (Fig. 22.31).
Figure 22.31 Filling defect in the small intestine due to the presence of Ascaris, seen on a radiograph after a barium meal.
(Courtesy of W. Peters.)
The presence of larvae in fresh stools is diagnostic of Strongyloides infection.
Pinworm infection is diagnosed by finding eggs on perianal skin
Although adult pinworms sometimes appear in the stools, the eggs are seldom seen because they are laid directly onto the perianal skin (Fig. 22.32). They can be found by wiping this area with a piece of clear adhesive tape (the ‘Scotch tape’ test) and examining the tape under the microscope.
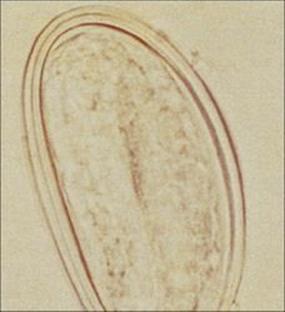
Figure 22.32 Egg of Enterobius on perianal skin.
(Courtesy of J.H. Cross.)
Treatment and prevention
Enterobius is treated with mebendazole or piperazine; Ascaris with mebendazole or piperazine; hookworm with mebendazole or albendazole; Trichuris with mebendazole or albendazole. Strongyloides requires treatment with ivermectin; thiabendazole is also effective, but is less well tolerated by the patient. At the community level, prevention can be achieved through improved hygiene and sanitation, making sure that faecal material is disposed of properly.
Other intestinal worms
Many other worm species can infect the intestine, but most are uncommon in resource-rich countries
Of the human tapeworms:
• The beef tapeworm Taenia saginata, transmitted through infected beef, is the most widely distributed. However, infection is usually asymptomatic, apart from the nausea felt on passing the large segments. Diagnosis involves finding these segments or the characteristic eggs in the stool (Fig. 22.33).
• Diphyllobothrium latum, the fish tapeworm, is widely distributed geographically, but infection is restricted to individuals eating raw or undercooked fish carrying the infective larvae. The eggs of this species have a terminal ‘lid’ and are the diagnostic stage in the stool (Fig. 22.34).
• Hymenolepis nana, the dwarf tapeworm, occurs primarily in children, infection occurring directly by swallowing eggs (Fig. 22.34). This worm has the ability to undergo autoinfection within the host’s intestine, so that a large number of worms can build up rapidly, leading to diarrhea and some abdominal discomfort.
All these tapeworms can be removed by praziquantel or niclosamide.
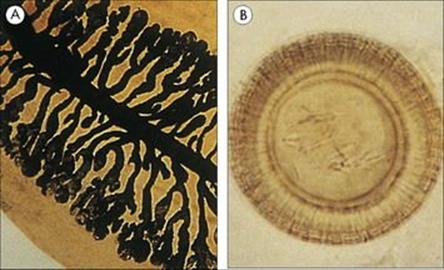
Figure 22.33 Taenia saginata. (A) Gravid proglottid stained with India ink to show numerous side branches. (B) Egg containing six-hooked (hexacanth) larva.
(Courtesy of R. Muller and J.R. Baker.)
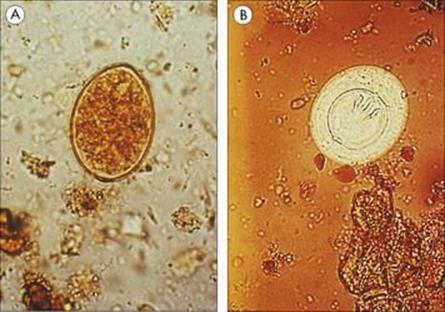
Figure 22.34 Eggs of (A) Diphyllobothrium latum and (B) Hymenolepis nana.
(Courtesy of R. Muller and J.R. Baker.)
Intestinal symptoms (predominantly diarrhea and abdominal pain) are also associated with infections by the nematode Trichinella spiralis, which is better known clinically for the pathology caused by the blood-borne muscle phase (see Chs 26 and 28). Infection with the two species of schistosome associated with mesenteric blood vessels (Schistosoma japonicum and S. mansoni) can also cause symptoms of intestinal disease. As the eggs pass through the intestinal wall, they cause marked inflammatory responses, granulomatous lesions form, and diarrhea may occur in the early acute phase. Heavy chronic S. mansoni infection is associated with inflammatory polyps in the colon, while severe involvement of the small bowel is more common with S. japonicum.
Systemic infection initiated in the gastrointestinal tract
We opened this chapter by noting that infections acquired by the ingestion of pathogens could remain localized in the gastrointestinal tract or could disseminate to other organs and body systems. Important examples of disseminated infection are the enteric fevers and viral hepatitis types A and E. Listeriosis also appears to be acquired via the gastrointestinal tract. For the sake of clarity and convenience, other types of viral hepatitis will also be discussed in this chapter.
Enteric fevers: typhoid and paratyphoid
The term ‘enteric fever’ was introduced in the last century in an attempt to clarify the distinction between typhus (see Ch. 27) and typhoid. For many years these two diseases had been confused, as the common root of their names suggests (typhus, a fever with delirium; typhoid, resembling typhus), but even before the causative agents were isolated (typhoid caused by S. typhi and typhus caused by Rickettsia spp.), it was pointed out that it was ‘just as impossible to confuse the intestinal lesions of typhoid with the pathologic findings of typhus as it was to confuse the eruptions of measles with the pustules of smallpox’. In fact, enteric fevers can be caused by S. typhi and three additional salmonella species, but the name ‘typhoid’ has stuck.
S. typhi, and paratyphi types S. paratyphi A, S. schottmuelleri (previously named S. paratyphi B), and S. hirschfeldii (previously named S. paratyphi C) cause enteric fevers
These species of Salmonella are restricted to humans and do not have a reservoir in animals. Therefore, spread of the infection is from person to person, usually through contaminated food or water. After infection, people can carry the organism for months or years, providing a continuing source from which others may become infected. Typhoid Mary, a cook in New York City in the early 1900s, is one such example. She was a long-term carrier who succeeded in initiating at least 10 outbreaks of the disease (see Ch. 16, Box 16.1).
The salmonellae multiply within, and are transported around, the body in macrophages
After ingestion, the salmonellae that survive the antibacterial defences of the stomach and small intestine penetrate the gut mucosa through the Peyer’s patches, probably in the jejunum or distal ileum (Fig. 22.35). Once through the mucosal barrier, the bacteria reach the intestinal lymph nodes, where they survive and multiply within macrophages. They are transported in the macrophages to the mesenteric lymph nodes and thence to the thoracic duct and are eventually discharged into the bloodstream. Circulating in the blood, the organisms can seed many organs, most importantly in areas where cells of the reticuloendothelial system are concentrated (i.e. the spleen, bone marrow, liver and Peyer’s patches). In the liver, they multiply in Kupffer cells. From the reticuloendothelial system, the bacteria re-invade the blood to reach other organs (e.g. kidney). The gallbladder is infected either from the blood or from the liver via the biliary tract, the bacterium being particularly resistant to bile. As a result, S. typhi enters the intestine for a second time in much larger numbers than on the primary encounter and causes a strong inflammatory response in Peyer’s patches, leading to ulceration, with the danger of intestinal perforation.
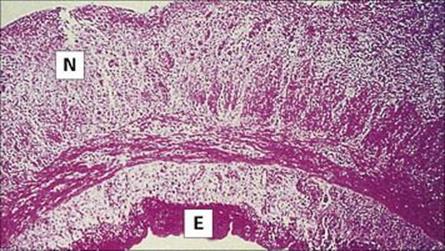
Figure 22.35 Typhoid. Section of ileum showing a typhoid ulcer with a transmural inflammatory reaction, focal areas of necrosis (N) and a fibrinous exudate (E) on the serosal surface (H&E stain).
(Courtesy of M.S.R. Hutt.)
Rose spots on the upper abdomen are characteristic, but absent in up to half of patients with enteric fever
After an incubation period of 10–14 days (range 7–21 days), the disease has an insidious onset with non-specific symptoms of fever and malaise accompanied by aches and respiratory symptoms, and may resemble a flu-like illness (see Ch. 15). Diarrhea may be present, but constipation is just as likely. At this stage, the patient often presents with a fever of unknown origin (FUO; see Ch. 29). In the absence of treatment, the fever increases and the patient becomes acutely ill. Rose spots – erythematous maculopapular lesions that blanch on pressure (Fig. 22.36) – are characteristic on the upper abdomen, but may be absent in up to half of patients. They are transient and disappear within hours to days. Without treatment, an uncomplicated infection lasts 4–6 weeks.
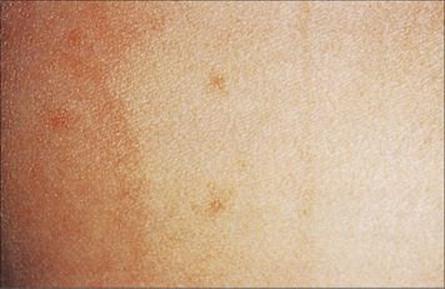
Figure 22.36 Rose spots on the skin in typhoid fever.
(Courtesy of W.E. Farrar.)
Before antibiotics, 12–16% of patients with enteric fever died, usually of complications
The complications can be classified into:
• those secondary to the local gastrointestinal lesions (e.g. haemorrhage and perforation; Fig. 22.37)
• those associated with toxaemia (e.g. myocarditis, hepatic and bone marrow damage)
• those secondary to a prolonged serious illness
• those resulting from multiplication of the organisms in other sites, causing meningitis, osteomyelitis or endocarditis.
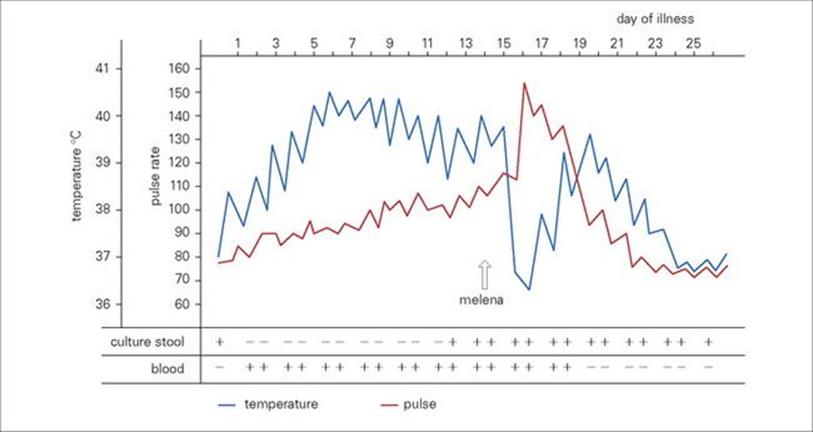
Figure 22.37 The clinical course of typhoid fever. Chart of temperature, pulse rate and bacteriologic findings in a patient whose illness was complicated by massive haemorrhage.
(Courtesy of H.L. DuPont.)
Before antibiotics became available, 12–16% of patients died, usually of complications occurring in the third or fourth week of the disease. Relapse after an initial recovery was also common.
One to three per cent of patients with enteric fever become chronic carriers
Patients usually continue to excrete S. typhi in the faeces for several weeks after recovery, and 1–3% become chronic carriers, which is defined as S. typhi excretion in faeces or urine for 1 year after infection. Chronic carriage is more common in women, in older patients and in those with underlying disease of the gallbladder (e.g. stones) or urinary bladder (e.g. schistosomiasis).
Diagnosis of enteric fever depends upon isolating S. typhi or paratyphi types using selective media
Diagnosis cannot be made on clinical grounds alone, although the presence of rose spots in a febrile patient is highly suggestive. Samples of blood, faeces and urine should be cultured on selective media. An antibody response to infection can be detected by an agglutination test (Widal test), but non-specific cross-reaction with other enterobacteria may also cause an increase in H and O antibody levels. Interpretation of the results is complicated and depends on knowing the normal antibody titres in the population and whether the patient has been vaccinated. A demonstration of a rising titre between acute- and convalescent-phase sera is more useful than examination of a single sample. At best, the results confirm the microbiologic diagnosis, at worst they are misleading.
Antibiotic treatment should be started as soon as enteric fever is diagnosed
Ciprofloxacin or ceftriaxone followed by cefixime have been effectively used in antimicrobial chemotherapy, which should continue for at least 1 week after the patient’s temperature has returned to normal. Some antibiotics appear active in vitro, but do not achieve a clinical cure, presumably because they do not reach the bacteria in their intracellular location. Isolates of S. typhi resistant to a variety of antimicrobial agents have been reported.
Prevention of enteric fever involves public health measures, treating carriers and vaccination
Breaking the chain of spread of infection from person to person depends upon good personal hygiene, adequate sewage disposal and a clean water supply. These conditions exist in the resource-rich world, where outbreaks of enteric fever are rare but still occur.
Typhoid carriers are a public health concern and should be excluded from employment involving food handling. Every effort should be made to eradicate carriage by antibiotic treatment, and if this is unsuccessful, removal of the gallbladder (the most common site of carriage) should be considered.
A single-dose injectable vaccine (Typhim Vi) which contains capsular polysaccharide antigen and an oral, live-attenuated, vaccine (strain Ty21a) are available and recommended for travellers to resource-poor countries. However, with both vaccines protection is complete in only 50–80% of recipients.
Listeriosis
Listeria infection is associated with pregnancy and reduced immunity
Listeria monocytogenes is a Gram-positive coccobacillus that is widespread among animals and in the environment. It is a food-borne pathogen, associated particularly with uncooked foods such as pâté, contaminated milk, soft cheeses and coleslaw. Studies of cases involving unpasteurized milk suggest that fewer than one thousand organisms may cause disease, and the ability of the organism to multiply, albeit slowly, at refrigeration temperatures allows an infective dose to accumulate in goods stored in this way. Even then, the population at risk appears primarily to be:
• pregnant women, with the possibility of infection of the baby in the uterus or during birth
• immunocompromised individuals including those with cancer, AIDS, on immunosuppressive drugs
• elderly individuals.
The disease usually presents as meningitis (see Ch. 24).
Viral hepatitis
An alphabetical litany of viruses directly target the liver, from hepatitis A to E
Hepatitis means inflammation and damage to the liver, and has differing aetiologies including non-infectious multisystemic conditions and drug toxicity as well as infectious agents. The latter include viruses and less commonly bacteria (e.g. Leptospira spp.), and other microorganisms. There is a broad spectrum of clinical illness ranging from asymptomatic, symptomatic with malaise, anorexia, nausea, abdominal pain and jaundice, to acute life-threatening liver failure, which is rare. Jaundice is a clinical term for the yellow tinge to the skin, sclera and mucous membranes. This is a result of liver cell damage which means that the liver cannot transport bilirubin into the bile, causing increased bilirubin levels in the body fluids. More than half of the liver must be damaged or destroyed before liver function fails. Regeneration of liver cells is rapid, but fibrous repair, especially when infection persists, can lead to permanent damage called cirrhosis. Cirrhosis results in a small, shrunken liver with poor function.
At least six different viruses are referred to as hepatitis viruses (Table 22.6), and generally they cannot be distinguished clinically. However, hepatitis A and E viruses are transmitted by the faecal–oral route and do not result in a carrier state; both infections resolve. In contrast, hepatitis B, D (delta), and C are transmitted by similar routes involving blood-contaminated equipment, although sexual transmission of hepatitis B is much more common than in hepatitis C, and all can lead to chronic carriage. Some agents have been reported that were thought to be involved in the spectrum of what is referred to as non-A–E hepatitis. However, there is no evidence that the GB, hepatitis G and TT viruses infect the liver directly, the liver being affected as a bystander. Other viruses also cause hepatitis as part of a disease syndrome and are dealt with in other chapters. Dramatic elevations of serum aminotransferase concentration (alanine aminotransferase, ALT; aspartate aminotransferase, AST) are characteristic of acute viral hepatitis. Specific laboratory tests to make the serological diagnosis of hepatitis A, B, D, C and E virus infections are available, as are PCR tests to detect and quantify the hepatitis B and C virus load in those with chronic infections. With the exception of hepatitis A and B there are no licensed vaccines, and specific antiviral treatments with and without immunomodulators are available for hepatitis B and C.
Table 22.6 The main viruses causing hepatitis in humans
Hepatitis A
This disease is caused by hepatitis A virus (HAV), a single-stranded unenveloped RNA virus that has its own genus Hepatovirus in the Picornaviridae family. There is only one serotype, and the virus is endemic worldwide.
HAV is transmitted by the faecal–oral route
Virus is excreted in large amounts in faeces (108 infectious doses/g) and spreads from person to person by close contact (poor hand hygiene), by intimate contact (anal intercourse) or by contamination of food or water. The incubation period is 3–5 weeks, with a mean of 4 weeks; virus is present in faeces 1–2 weeks before symptoms appear and during the first week (sometimes also the second and third week) of the illness. Person-to-person transmission can lead to outbreaks in places such as schools and camps, and viral contamination of water or food is a common source of infection (Fig. 22.38). In resource-poor countries up to 90% of children have been infected by 5 years of age, whereas in resource-rich countries up to 20% of young adults have been infected. The latter figure used to be higher but is mostly a result of improved sanitation and less overcrowding.
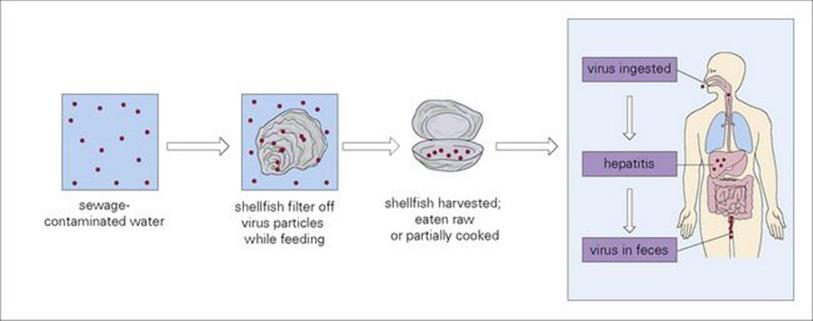
Figure 22.38 Contamination of shellfish by hepatitis A virus (HAV) can lead to human infection.
Clinically, hepatitis A is milder in young children than in older children and adults
After infection, the virus enters the blood from the gastrointestinal tract, where it may replicate. It then infects liver cells, passing into the biliary tract to reach the intestine and appears in faeces (Fig. 22.39). Relatively small amounts of virus enter the blood at this stage. Events during the rather lengthy incubation period are poorly understood, but liver cells are damaged, and this is thought to be due to direct viral action. Common clinical manifestations are fever, anorexia, nausea and vomiting; jaundice is more common in adults. The illness generally has a more sudden onset than hepatitis B. The best laboratory method for diagnosis is to detect HAV-specific IgM antibody in serum.
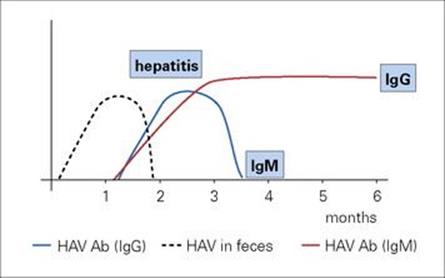
Figure 22.39 The clinical and virologic course of HAV. Ab, antibody.
Pooled human normal immunoglobulin (HNIG) contains antibody to HAV and will prevent or attenuate infection if given as pre- or post-exposure prophylaxis. There is no antiviral therapy, but an effective formaldehyde-inactivated vaccine should be offered to a number of groups at particular risk of infection. These include travellers to HAV-endemic countries, sewage workers, child daycare centre staff, institutional care workers, male homosexuals, and individuals with chronic liver disease. The vaccine is used alone or together with HNIG in certain situations in the post-exposure setting providing it can be given to contacts within 14 days of the onset of jaundice in the infected individual.
Hepatitis E
Hepatitis E virus (HEV) spreads by the faecal–oral route
This disease, also known as enteric non-A-non-B hepatitis, is caused by a small single-stranded RNA virus, and shares similarities with the caliciviruses. It has been classified in the genus Hepevirus in the family Hepeviridae, with four genotypes and one serotype.
Genotypes 1 and 2 have been involved in epidemics in resource-poor countries and infect humans only, and genotypes 3 and 4 infect humans and other animals in both resource-rich and -poor settings and are zoonoses. The virus is excreted in faeces and spreads by the faecal–oral route. It is the major cause of sporadic (up to 60%) as well as epidemic hepatitis in Asia, in the latter due to water-borne routes of transmission. In addition, there are sporadic cases in resource-rich countries.
HEV has been identified in a variety of animals, especially pigs, rabbits, chickens and sika deer and they constitute a probable reservoir for infection. Hepatitis E is likely to be another example of a zoonotic infection and experimental data showing the potential for transmission between pigs and humans have been reported. In Japan, eating undercooked pig liver and uncooked meats has been identified as a source of infection. The incubation period is 6–8 weeks. The disease is generally mild, but is severe in pregnant women, with a high mortality, up to 20% during the third trimester, due to fulminant hepatitis. The virus is eliminated from the body on recovery and there are no carriers. The diagnosis is made using serological tests to detect HEV-specific IgM. The 3-D crystal structure of the HEV capsid protein has been determined, which will lead to potential vaccines and antiviral agents. Two recombinant vaccines have undergone successful clinical trials.
Hepatitis B
This disease is caused by hepatitis B virus (HBV) infections, a hepadna (hepatitis DNA) virus (Box 22.3) containing a partially double-stranded circular DNA genome and three important antigens: HB surface antigen, HB core antigen and HBe antigen (Fig. 22.40; Table 22.7). HBe antigen is a soluble component secreted by the virus core, is expressed on the hepatocyte surface, and is targeted by the host immune system. Infection with a given strain of HBV confers resistance to all strains, but antigenic variation occurs. The four serological subtypes (adw, adr, ayw and ayr) have been superseded by the genetic classification in which the eight genotypes A to H have been determined. These can influence the clinical outcome of infection and response to antiviral treatment, and are useful in epidemiologic studies.
![]()
Box 22.3 Lessons in Microbiology
Hepatitis A
In August 1988, the Florida Department of Health and Rehabilitation Services traced 61 people who had suffered serologically confirmed infection with HAV. These individuals resided in five different states, but 59 of them had eaten raw oysters from the same growing areas in Bay County coastal waters. The oysters had been gathered illegally from outside the approved harvesting areas and were contaminated with HAV. The mean incubation period of the disease was 29 days (range 16–48 days). Probable sources of faecal contamination near the oyster beds included boats with inappropriate sewage disposal systems and discharge from a local sewage treatment plant that contained a high concentration of faecal coliforms.
Hepatitis B
One of the largest outbreaks of hepatitis B virus infections in Europe occurred in London in 1998. A patient went to an alternative medicine clinic and was treated with a technique called autohaemotherapy. This involved mixing a small sample of the patient’s blood with saline, then injecting the blood and saline mixture into her buttocks or acupuncture points. She subsequently developed acute hepatitis B and the public health doctors were contacted and an investigation started having identified the practices in the clinic that could have resulted in her becoming jaundiced.
A lookback exercise involving 352 patients who had attended the clinic between January 1997 and February 1998 and four staff was carried out. Evidence of exposure to hepatitis B was found in samples from 57 (16%) of this group. Hepatitis B surface antigen was detected in blood samples collected from a total of 33 patients and staff, 23 of whom had acute hepatitis B. Molecular analysis revealed that 30 (91%) samples had identical nucleotide sequences and were part of a large community outbreak of hepatitis B. Five patients were chronic hepatitis B carriers, one of whom was the likely source of infection, with the vehicle being the contaminated saline in a vial that was used to mix the blood on a number of occasions for the other patients involved in the outbreak.
This demonstrated once again that only single-use vials must be used in healthcare settings, together with the benefits in those countries that offer universal immunization against hepatitis B to their populations.
Hepadnaviruses
Hepadnaviruses are also found in woodchucks, ground squirrels and Pekin ducks. In each case, the infection persists in the body, with HBsAg-like particles in the blood and chronic hepatitis and liver cancer as sequelae. These viruses often infect non-hepatic cells. In northeast USA, for instance, 30% of woodchucks carry their own type of hepadnavirus and most develop liver cancer by later life. The virus replicates not only in liver cells, but also in lymphoid cells in the spleen, peripheral blood and thymus and in pancreatic acinar cells and bile duct epithelium.
![]()

Figure 22.40 During acute infection, and in some carriers there are 106–107 infectious (Dane) particles/mL of serum (A), and as many as 1012 HB surface antigen (HBsAg) particles/mL (B). (C) Electron micrograph showing Dane particles and HBsAg particles.
(Courtesy of J.D. Almeida.)
Table 22.7 Characteristics of hepatitis B virus (HBV) antigens (Ag) and antibodies (Ab)
|
HBsAg |
Envelope (surface) antigen of HBV particle also occurs as free particles (spheres and filaments) in blood; indicates infectivity of blood |
|
HBsAb |
Antibody to HBsAg; provides immunity; appears late (not in carriers) |
|
HBcAg |
Antigen in core of HBV |
|
HBcAb |
Antibody to HBcAg; appears early |
|
HBeAg |
Antigen derived from core; indicates high transmissibility |
|
HBeAb |
Antibody to the soluble component of core |
HB surface antigen can be found in blood and other body fluids
HBV can be transmitted by various routes, including:
• sexual intercourse
• vertically from mother to child (intrauterine, peri- and postnatal infection; see Ch. 23)
• via blood and blood products, blood-contaminated needles and equipment which may be used by injecting drug users
• in association with tattooing, body-piercing and acupuncture, again due to reusing needles which may be contaminated by blood.
Transmission has been reported in healthcare settings such as renal units and has been associated with blood contaminated haemodialysis equipment. This has been reduced dramatically since the introduction of regular HB surface antigen (HBsAg) monitoring of patients and disposable dialysis cartridges. In addition, incidents have been reported involving HBV transmission from hepatitis B carrier healthcare workers (HCWs) to their patients while carrying out exposure-prone procedures, such as cardiothoracic surgery, due to intraoperative needlestick injuries resulting in blood-to-blood contact. Hepatitis B immunization and HBsAg screening of HCWs reduces the incidence of these transmission events. Blood and organ donors are also screened for HBsAg and HB core antibody in many counties worldwide, reducing the potential for transmission to recipients.
The number of HBV carriers worldwide is estimated to be over 350 million, and they play a major role in transmission. The HBV carrier prevalence is estimated to be up to 0.5% in north, west and central Europe, North America and Australia, up to 0.7% in east Europe, the Mediterranean littoral, Central and South America, Russia and southwest Asia, and up to 20% in southeast Asia, sub-Saharan Africa and China. In countries where infant and childhood infection is common (possibly because there is a high carrier rate in mothers), overall carrier rates are higher.
HBV is not directly cytopathic for liver cells, and the pathology is largely immune mediated
After entering the body, the virus reaches the blood, then the liver, where the result is inflammation and necrosis. Much of the pathology is immune mediated, as infected liver cells are attacked by virus-specific cytotoxic T cells. The incubation period ranges from 6 weeks to 6 months, the median being 2.5 months.
As liver damage increases, clinical signs of hepatitis appear (Fig. 22.41); the disease is generally more severe than hepatitis A. The immune response slowly becomes effective, virus replication is curtailed, and eventually, although sometimes not for many months, the blood becomes non-infectious.
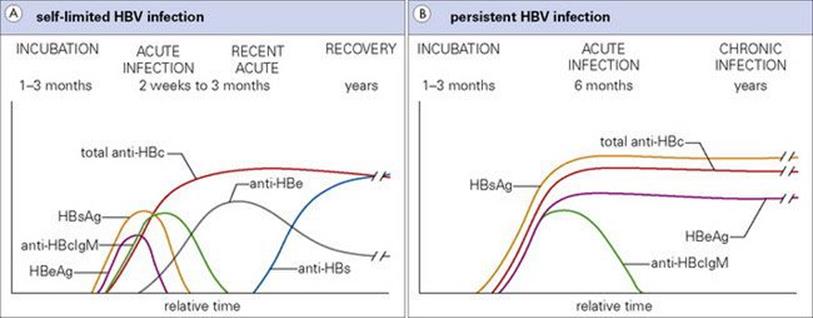
Figure 22.41 (A) Clinical and virologic course of hepatitis B, with recovery. (B) Clinical and virologic course in a carrier of hepatitis B.
(Redrawn from: Farrar, W.E., Wood M.J., Innes, J.A. et al. (1992) Infectious Diseases, 2nd edn. London: Mosby International.)
Certain groups of people are more likely to become carriers of hepatitis B
People with a more vigorous immune response to the infection clear the virus more rapidly, but tend to suffer a more severe illness. However, about 10% of infected adults fail to eliminate the virus from the body, and become carriers. The blood remains infectious, often for life, and although continuing liver damage can cause chronic hepatitis, the damage is often so mild that the carrier remains in good health. Certain groups of people are more or less likely to become carriers, as follows:
• Immunodeficient patients develop a milder disease due to the effect of reducing the host response to the infection, but are more likely to become carriers.
• There is a marked age-related effect. For example, in a study carried out in Taiwan, 90–95% of perinatally infected infants became carriers compared with 23% of those infected at 1–3 years of age and only 3% of those infected as university students.
• Gender is another factor, with males being more likely to become carriers.
Complications of hepatitis B are cirrhosis and hepatocellular carcinoma
Complications of hepatitis B include:
• Cirrhosis, as a result of chronic active hepatitis. This is an irreversible form of liver injury which may lead to primary hepatocellular carcinoma.
• Hepatocellular carcinoma is one of the 10 most common cancers worldwide. Hepatitis B carriers are 200 times more likely to develop liver cancer than non-carriers. This is not seen until 20–30 years after the infection. The cancer cells contain multiple integrated copies of HBV DNA and this could be the carcinogenic factor (see Ch. 17).
Serological tests are used in the diagnosis of HBV infection
HBsAg appears in the serum during the incubation period in the form of infectious Dane particles (see Fig. 22.41). The characteristic serological picture in an acute HBV infection includes the detection of HBsAg, HB core IgM and HBe antigen. The HBsAg concentration generally falls and finally disappears during recovery and convalescence. As HBsAg disappears, the HB core IgM level wanes over the next 3 months, HB core total antibody (IgM and IgG) is detected but is almost all IgG by this stage, and HB surface antibody becomes detectable. Therefore, evidence of past infection will give the following serological profile (Table 22.8): HBsAg negative, HB core total antibody positive and HB surface antibody positive. HBV carriage is defined by detecting HBsAg in blood for a period of 6 months after the acute infection. When HBe antigen is detected, there are large amounts of virus in the blood and the carrier is considered to be of high infectivity, and when it disappears HBe antibody may become detectable. HBe antibody-positive carriers are considered to be of low infectivity. However, HBV DNA load is becoming a more useful marker of infectivity as mutations have been detected in the region encoding the e antigen which result in absence of e antigen production yet infectious virus is still assembled. They are known as precore mutant viruses. Therefore, these patients will be HBe antigen negative and HBe antibody positive but could be highly infectious.
Table 22.8 Interpretation of hepatitis B virus serological results
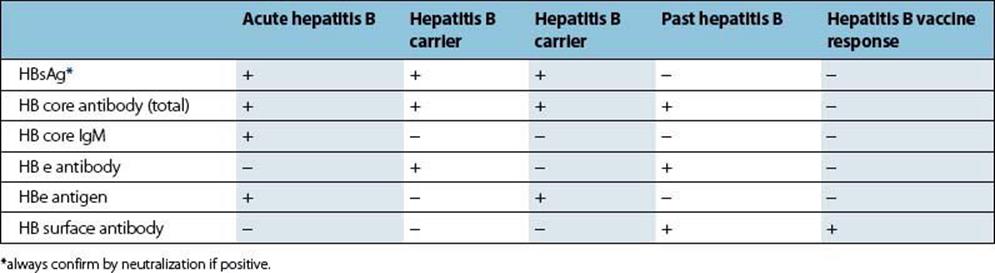
Antiviral therapy is available
Two classes of drugs used to treat hepatitis B virus infections are pegylated interferon and nucleotide/nucleoside analogues. Management of HBV carriers has been revolutionized with the advent of oral antiviral therapy, in particular lamivudine (3TC), adefovir, entecavir, emtricitabine and tenofovir. Previously, therapy with interferon α2b, an immunomodulator, was used, but only 30% of selected patients achieved sustained responses. In addition, interferon (IFN) treatment has significant side effects. However, the better pharmacokinetics of pegylated interferon α2a has improved the results with respect to sustained response after treatment has been discontinued, especially in the e antigen-positive carriers. Better responses to IFN are seen in females under 50 years old, infected in adulthood with HBV genotype A or B, with a lower HBV DNA load and alanine aminotransferase more than twice the upper limit of normal. Moreover, with the range of antivirals available, courses of treatment are available that depend on a number of factors related to the virus as well as the stage of liver disease. For example, entecavir or tenofovir can be used if lamivudine resistance develops, which it does in 70% of those treated after 5 years.
Hepatitis B infection can be prevented by immunization
The original vaccine was produced in 1981 and consisted of purified HBsAg, prepared from the plasma of carriers, which was chemically treated to kill any contaminating viruses. The current vaccine is genetically engineered HBsAg produced in yeast or mammalian cells. Three injections of vaccine over a 6-month period give good protection in over 90% of healthy adults. Immunization is recommended, especially for those who may be exposed to blood or blood products, such as receiving multiple transfusions or dialysis patients, all healthcare workers, sexual contacts of individuals with acute or chronic hepatitis B, and injecting drug users. One problem is that up to 10% of healthy individuals fail to produce the protective HB surface antibody, even when re-immunized. This could be due to genetically determined defects in the immune repertoire or because of the induction of immune suppressor cells.
After accidental exposure to infection, hepatitis B immunoglobulin (HBIG) can be used to provide immediate passive protection. This is prepared from the serum of individuals with high titres of HB surface antibody. It may also be used together with hepatitis B vaccine to prevent transmission to children of HBV-carrier mothers.
Hepatitis C
Hepatitis C virus was the most common cause of transfusion-associated non-A-non-B viral hepatitis
Hepatitis C virus (HCV) was discovered in 1989 as the cause of 90–95% of cases of transfusion-associated non-A-non-B hepatitis. It is a single-stranded RNA virus related to the flaviviruses and pestiviruses. The discovery of HCV was a tour de force in molecular virology. The viral RNA was extracted from blood, a complementary DNA (cDNA) clone was made, and viral antigen produced. Serum from individuals with non-A-non-B hepatitis was then tested for the presence of antibody to the viral antigen. The introduction of first-generation HCV antibody screening tests between 1990 and 1992, and subsequent improvement in sensitivity and specificity of these assays and genome detection methods, has resulted in a massive reduction in transfusion-associated HCV infection. It is estimated that more than 170 million people worldwide are infected with HCV.
HCV transmission routes share similarities with hepatitis B
HCV is present in blood, and transmission routes include blood and blood products, blood-contaminated needles and equipment which may be used by injecting drug users, and in association with tattooing, body-piercing and acupuncture, again due to reusing potentially blood-contaminated needles from other clients. Transmission has been reported in healthcare settings such as renal units because of contaminated dialysis equipment and other fomites, including gloves. Although the introduction of regular HCV monitoring of patients and disposable dialysis cartridges has helped in infection control, transmission has also occurred by other routes, probably often involving contaminated gloves worn by HCWs, which are not changed between patients. In addition, there have been incidents involving HCV transmission from HCV carrier HCWs carrying out exposure-prone procedures on their patients, such as intraoperative needlestick injuries resulting in blood-to-blood contact during cardiothoracic surgery. Unlike hepatitis B, HCV transmission is uncommon vertically, from mother to infant, and by sexual intercourse. There may be other methods of spread, as the route of transmission is unknown in up to 40% of infected individuals.
The HCV envelope binds to the hepatocyte cell surface membrane allowing viral entry, probably by endocytosis. Some of the HCV proteins interfere with the host response and other evasive measures include the high degree of genetic diversity due to the high error rate of RNA replication.
Six HCV genotypes and multiple subtypes have been identified. Genotype determination is predictive of antiviral therapy response, genotype 1 being associated with poor response. Viral and host factors affect the disease progression rate, with high HCV load in blood, genotype, and the degree of viral heterogeneity referred to as the quasispecies, being associated with more rapid progression. Viral clearance is associated with both the development and persistence of strong HCV-specific cytotoxic T-cell and helper T-cell responses. Being infected with one genotype does not protect against the others; therefore multiple infections are possible, thus making the production of a cross-protective vaccine more difficult.
About 75–85% of HCV-infected individuals develop chronic HCV
The incubation period is 2–4 months, with a mean of 7 weeks. Subclinical infection is the rule, with mild disease occurring in about 10% of individuals. Virus is often detectable in the blood after recovery from the acute illness, and carriers are a source of infection. Up to 2% of apparently healthy individuals in the USA have HCV antibody, and as a result, between 2.7 and 3.9 million people have an active infection. About 75–85% of HCV-infected individuals will develop chronic HCV and 10–15% will progress to cirrhosis within the first 20 years, with a resultant 1–4% risk per year of liver cancer in those with established cirrhosis. It is also a leading indication for liver transplantation. The rate of chronic HCV infection depends on the infected individual’s age, gender, ethnicity and immune response.
Diagnostic tests for HCV infection involve serological assays to detect HCV antibody, qualitative and quantitative HCV RNA detection methods and genotype analysis. HCV RNA is present in approximately 70% of individuals with HCV antibody.
Treatment with pegylated IFNα and ribavirin is the standard of care
The aim of treatment is a sustained virological response (SVR) which means that HCV RNA cannot be detected 6 months after completing a course of treatment. Antiviral therapy with ribavirin has made a huge impact on the management of HCV infection, used in combination with pegylated interferon alpha. Originally, IFNα monotherapy resulted in up to 40% initial response rates, but under 20% were sustained responses. Treatment with pegylated IFNα, in which polyethylene glycol is attached to interferon and extends the half-life and duration of activity, and ribavirin has resulted in an SVR in 45% of patients with genotype 1 or 4 infections (48 weeks’ treatment) and 80% of those with genotype 2 or 3 (24 weeks’ treatment). Combining pegylated interferon and ribavirin with viral protease inhibitors such as telaprevir or boceprevir improves the SVR rates in the more difficult to treat HCV infections. No vaccine is available.
Hepatitis D
Hepatitis D virus can only multiply in a cell infected with HBV
This is caused by hepatitis D virus (HDV or delta virus), which has a very small, circular, single-stranded RNA genome and is a defective virus, so-named because it can successfully multiply in a cell only when the cell is infected with HBV at the same time. When HDV buds from the surface of a liver cell it acquires an envelope consisting of HBsAg (Fig. 22.42). The HBs envelope makes the 35–37-nm virus particle infectious by attaching it to hepatic cells.
Figure 22.42 Structure of hepatitis D virus in serum. Ag, antigen.
Spread of HDV is similar to that of HBV and HBC
Infected blood contains very large amounts of virus (up to 1010 infectious doses/mL in experimentally infected chimpanzees), and spread is similar to that of the other parenterally transmitted hepatitis viruses.
HDV infection may occur at the same time as an HBV infection, and the resulting disease is often more severe than with HBV alone. Alternatively, HDV superinfection of an HBV carrier may occur, which may accelerate the course of the chronic hepatitis-B-related liver disease. Infection is uncommon in the UK and USA, but common in the Mediterranean littoral, parts of South America and Africa. Worldwide, HDV infection may occur in approximately 5% of HBV carriers.
The diagnosis is made by serological tests for HD antigen (‘delta’ antigen) or HD IgM and IgG. HBsAg will also be present.
There is no HDV-specific vaccine, but successful hepatitis B immunization prevents infection with hepatitis D.
Viral hepatitis, the rest of the alphabet
After the discovery of HCV, a small percentage of hepatitis infections known to be transmitted by blood transfusion have yet to be attributed to a virus infection, although hepatitis G virus referred to as GB virus C, transfusion transmitted virus or Torque Teno virus (TTV) and SENV 25–28 have been detected in individuals with post-transfusion hepatitis. There are even more human hepatitis viruses waiting to be discovered.
Parasitic infections affecting the liver
Few protozoa affect the liver. Some worms live there as adults and others migrate through the liver to reach other locations.
Inflammatory responses to the eggs of Schistosoma mansoni result in severe liver damage
Liver pathology in parasitic infections is most severe in S. mansoni infection. Although the worms spend only a relatively short time in the liver before moving to the mesenteric vessels, eggs released by the females can be swept by the bloodstream into the hepatic circulation and be filtered out in the sinusoids. The inflammatory response to these trapped eggs is the primary cause of the complex changes that result in hepatomegaly, fibrosis and the formation of varices (Fig. 22.43).
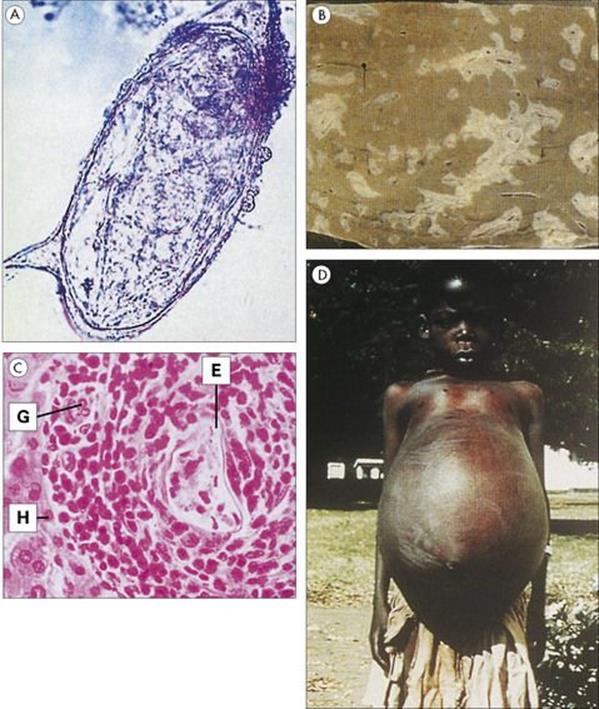
Figure 22.43 The portal fibrosis of Schistosoma mansoni is the end result of huge numbers of granulomas formed around worm eggs deposited in the liver. In the related Schistosoma haematobium infection, a similar process occurs in the wall of the bladder. (A) Egg of S. mansoni (× 400). (Courtesy of R. Muller.) (B) Pipe-stem fibrosis in the liver as a result of coalescent calcified granulomas. (Courtesy of R. Muller.) (C) Cellular reaction around an egg in the liver. E, egg containing miracidium; G, giant cell; H, hepatic cell. (Courtesy of R. Muller.) (D) Clinical schistosomiasis with massive hepatosplenomegaly and ascites due to portal obstruction.
(Courtesy of G. Webbe.)
Whereas schistosomiasis is widespread in tropical and subtropical regions, other parasitic infections affecting the liver are much more restricted in their distribution (e.g. clonorchiasis, fascioliasis, hydatid disease).
In Asia, infections with the human liver fluke Clonorchis sinensis are acquired by eating fish infected with the metacercarial stage. Juvenile flukes released in the intestine move up the bile duct and attach to the duct epithelium, feeding on the cells and blood and tissue fluids. In heavy infections, there is a pronounced inflammatory response, and proliferation and hyperplasia of the biliary epithelium, cholangitis, jaundice and liver enlargement are possible consequences. Chronic infection with Clonorchis sinensis or Opisthorchis viverrini is a recognized cause of intrahepatic cholangiocarcinoma.
A number of animal liver flukes can also establish themselves in humans. These include species of Opisthorchis (in Asia and Eastern Europe) and the common liver fluke Fasciola hepatica. In general, the symptoms associated with these infections are similar to those described for C. sinensis.
The larval stages of the dog tapeworm Echinococcus granulosus can develop in humans when the eggs are swallowed. Larvae from the eggs move from the intestine into the body and frequently develop into large hydatid cysts (cystic echinococcosis) in the liver. These can be diagnosed on scans as large cavities. Apart from pressure damage to surrounding tissues, rupture of the cysts leads to secondary cysts and may cause anaphylaxis. Cysts can be removed surgically or treated with benzimidazole drugs. E. multilocularis, acquired from eggs passed by wild carnivores, develops in the liver not as cysts but as a ramifying mass resembling a carcinoma (alveolar echinococcosis). E. multilocularis is treated by radical excision plus benzimidazole therapy. Inoperable cysts require life-long drug therapy. Liver transplantation is sometimes used.
Other parasitic infections associated with liver pathology are malaria, leishmaniasis, ascariasis and extraintestinal amoebiasis, which causes liver abscesses.
Liver abscesses
Despite the name, an amoebic liver abscess does not consist of pus
E. histolytica can move from the gastrointestinal tract and cause disease in other sites, including the liver (see above). However, the term ‘amoebic liver abscess’ is not strictly accurate because the lesion formed in the liver consists of necrotic liver tissue rather than pus. True liver abscesses – walled-off lesions containing organisms and dead or dying polymorphs (pus) – are frequently polymicrobial, containing a mixed flora of aerobic and anaerobic bacteria (Fig. 22.44). Lesions caused by E. granulosus in hydatid disease can become secondarily infected with bacteria. The source of infection may be local to the lesion or another body site, but is usually undiagnosed. Broad-spectrum antimicrobial therapy is required to cover both aerobes and anaerobes.
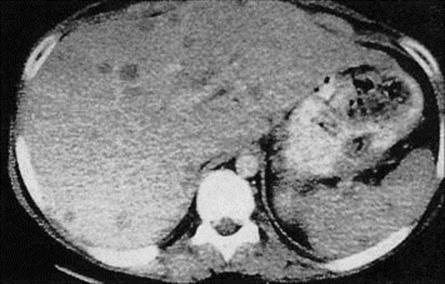
Figure 22.44 Multiple pyogenic liver abscesses due to Pseudomonas aeruginosa.
(Courtesy of N. Holland.)
Biliary tract infections
Infection is a common complication of biliary tract disease
Although infection is not often the primary cause of disease in the biliary tract, it is a common complication. Many patients with gallstones obstructing the biliary system develop infective complications caused by organisms from the normal gastrointestinal flora such as enterobacteria and anaerobes. Local infection can result in cholangitis and subsequent liver abscesses or invade the bloodstream to cause septicaemia and generalized infection. Removing the underlying obstruction in the biliary tree is a prerequisite to successful therapy. Antibacterial therapy is usually broad spectrum, covering both aerobes and anaerobes.
Peritonitis and intra-abdominal sepsis
The peritoneal cavity is normally sterile, but is in constant danger of becoming contaminated by bacteria discharged through perforations in the gut wall arising from trauma (accidental or surgical) or infection. The outcome of peritoneal contamination depends upon the volume of the inoculum (1 mL of gut contents contains many millions of microorganisms), and the ability of the local defences to wall off and destroy the microorganisms.
Peritonitis is generally classified as primary (without apparent source of infection) or secondary (e.g. due to perforated appendicitis, ulcer, colon)
Peritonitis usually begins as an acute inflammation in the abdomen which may progress to the formation of localized intra-abdominal abscesses. In general, the aetiologic agents responsible for primary and secondary peritonitis and intraperitoneal abscesses are different. Spontaneous bacterial peritonitis (SBP) is most commonly associated with cirrhosis of the liver. SBP is typically due to Gram-negative enteric bacteria, most commonly E. coli. Secondary peritonitis and intra-abdominal abscesses more often involve a mixture of organisms, especially the Gram-negative anaerobe Bacteroides fragilis. Mycobacterium tuberculosis and Actinomyces can also cause intraperitoneal infection (Fig. 22.45). In the absence of appropriate antibiotic therapy, infections are frequently fatal, and even with appropriate treatment the mortality remains at 1–5%. Empiric antibiotic therapy for SBP commonly involves third-generation cephalosporins (see Ch. 33) with re-evaluation when culture results are available. Initial antimicrobial treatment of secondary peritonitis must especially target the Gram-negative anaerobe B. fragilis (e.g. metronidazole) and Gram-negative aerobic pathogens (e.g. third-generation cephalosporins, etc.) as well as taking steps to eliminate the source of contamination. Mycobacterial infection requires specific antituberculosis therapy (see Ch. 33), while actinomycosis responds well to prolonged treatment with penicillin.
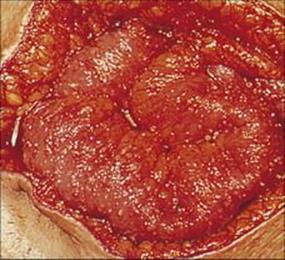
Figure 22.45 Tuberculous peritonitis. Oedematous bowel with multiple lesions on the peritoneal surface.
(Courtesy of M. Goldman.)
![]()
 Key Facts
Key Facts
• Diarrheal disease is a major cause of morbidity and mortality in the resource-poor world. A wide range of diverse microbes cause infections of the gastrointestinal tract. Diarrhea, the most common symptom, ranges from mild and self-limiting to severe with consequent dehydration and death.
• Gastrointestinal pathogens are transmitted by the faecal–oral route. They may invade the gut, causing systemic disease (e.g. typhoid), or multiply and produce locally acting toxins and damage only the gastrointestinal tract (e.g. cholera). The number of organisms ingested and their virulence attributes are critical factors in determining whether infection becomes established.
• Microbiologic diagnosis is usually impossible without laboratory investigations, but the patient’s history, including food and travel history, provides useful pointers.
• The major bacterial causes of diarrhea are E. coli, salmonellae, Campylobacter, V. cholerae and shigellae. Other less common causes include Cl. perfringens, B. cereus, V. parahaemolyticus and Y. enterocolitica. Food poisoning (i.e. the ingestion of bacterial toxins in food) is caused byStaph. aureus and Cl. Botulinum.
• E. coli is the major bacterial cause of diarrhea in resource-poor countries and of traveller’s diarrhea. Distinct groups within the species (ETEC, EHEC, EPEC and EIEC) have different pathogenic mechanisms – some are invasive, others toxigenic.
• Salmonellae and Campylobacter are common in resource-rich countries, have large animal reservoirs and spread via the food chain. Both cause disease by multiplication in the gut and the production of locally acting toxins.
• V. cholerae and shigellae have no animal reservoirs, and the diseases are potentially eradicable. Transmission is prevented by good hygiene, clean drinking water and hygienic disposal of faeces. The pathogenesis of cholera depends upon production of cholera enterotoxin, which acts on the gastrointestinal mucosal cells. In contrast, Shigella invades the mucosa, causing ulceration and bloody diarrhea, symptoms similar to those of amoebic dysentery.
• H. pylori is associated with gastritis and duodenal ulcers. Removal of the bacterium by combination treatment with antibiotics and proton pump inhibitors reduces symptoms and encourages healing.
• Disruption of the normal bacterial flora of the gut (usually due to antibiotic treatment) allows organisms normally absent or present in small numbers (e.g. Cl. difficile) to multiply and cause antibiotic-associated diarrhea.
• Viral gastroenteritis causes appalling morbidity and mortality, especially in young children in the resource-poor world. The chief culprits are the rotaviruses, which are specific to humans, spread by the faecal–oral route and restrict their multiplication to the gastrointestinal epithelial cells, which they destroy. Very small numbers can initiate infection and multiply in the gut to produce enormous numbers for excretion and transmission to new hosts.
• Ingestion of food or water contaminated with S. typhi or paratyphi types can result in the systemic infection enteric (typhoid) fever. These pathogens invade the gut mucosa and are ingested by, and survive in, macrophages. They are transported via the lymphatics to the bloodstream from whence they seed many organs and give the characteristic multisystem disease. Positive diagnosis depends upon culture of the organism. Specific antibiotic therapy is required and specific prevention is achievable through immunization.
• Hepatitis is usually caused by viruses (e.g. hepatitis A–E). Hepatitis A and E are transmitted by the faecal–oral route and the rest by contaminated blood or the sexual route. Infection with HBV and HCV often leads to chronic hepatitis and can result in liver cancer.
• Many protozoa and worms live in the intestine, but relatively few cause severe diarrhea. Important protozoa are E. histolytica, G. intestinalis and Cryptosporidium, which are acquired by ingestion of infective cysts in faecally contaminated food or water. Important worms are Ascaris,Trichuris, Strongyloides and hookworm. They have more complex routes of transmission, with the eggs or larvae requiring a development period outside the human host.
• Parasitic infections involving the liver include infections by Schistosoma mansoni in the tropics and subtropics, and Clonorchis sinensis, the human liver fluke, in Asia. Other parasitic infections with important liver pathology include malaria, leishmaniasis, extraintestinal amoebiasis, hydatid disease and ascariasis.
• Infection of the biliary tree is usually secondary to obstruction. The normal intestinal flora causes mixed infections, which may extend to produce liver abscesses and septicaemia.
• Peritonitis and intra-abdominal sepsis follow contamination of the normally sterile abdominal cavity with intestinal microbes. The presentation is acute, and infection can be fatal. Antibiotic therapy against both aerobic and anaerobic bacteria is essential.
![]()