Botany: An Introduction to Plant Biology - Mauseth, James D. 2017
Plant Structure
Flowers and Reproduction

Chapter Opener Image: These are the dry fruits (seed pods) of a small wild mustard. In mustard, each fruit has a thin septum and seeds are attached along the edges. Here, the walls of the fruits have fallen away, freeing the seeds, but several have not yet been blown away. With their disk-like shape, the seeds probably float on water, and rain will thus spread them away from the parent plant. Notice several tiny, brown seed-like structures: These started to develop as seeds but failed for some reason.
OUTLINE
✵ Concepts
✵ Asexual Reproduction
✵ Sexual Reproduction
- The Plant Life Cycle
- Flower Structure
- Gametophytes
- Fertilization
- Embryo and Seed Development
- Fruit Development
✵ Flower Structure and Cross-Pollination
- Cross-Pollination
- Stamen and Style Maturation Times
- Stigma and Pollen Incompatibility
- Monoecious and Dioecious Species
- Animal-Pollinated Flowers
- Wind-Pollinated Flowers
- Ovary Position
✵ Inflorescences and Pollination
✵ Fruit Types and Seed Dispersal
- True Fruits and Accessory Fruits
- Classification of Fruit Types
Box 9-1 Plants and People: Flowers, Fruits, Seeds, and Civilization
LEARNING OBJECTIVES
After reading this chapter, students will be able to:
✵ Discuss the advantages and disadvantages of asexual plant reproduction.
✵ Discuss the advantages and disadvantages of sexual plant reproduction.
✵ Explain one method of asexual reproduction.
✵ Identify the structures involved in the plant sexual reproduction cycle.
✵ List the four types of floral appendages.
✵ Describe microgametophytes and megagametophytes.
✵ Discuss endosperm development.
✵ Name the stages of embryo development.
✵ Compare cross-pollination, self-pollination, wind pollination, and animal pollination.
✵ Explain the advantages and disadvantages of inflorescences.
✵ Describe ways to classify fruits.
 Did You Know?
Did You Know?
✵ Flowers contain the organs and tissues angiosperms need for sexual reproduction; cones are equivalent structures in conifers.
✵ In addition to sexual reproduction, many plants also reproduce asexually, producing progeny that are clones of themselves.
✵ Seeds are always produced within a fruit or a cone; there are no exceptions (the tan-colored strawberry “seeds” are fruits that each contain one true seed, and the red sweet tissue is a “false fruit”).
✵ Pollen grains are complete plants: They have less than a dozen cells, have no leaves, no roots, and no stems, but are in fact complete plants.
![]() Concepts
Concepts
Reproduction can serve two very different functions: (1) producing offspring that have identical copies of the parental genes or (2) generating new individuals that are genetically different from the parents. Under certain environmental conditions, species that are genetically diverse survive better than genetically homogeneous species; under other conditions, just the opposite is true (in a genetically diverse species, individuals differ slightly from each other, as is true of humans). A plant that has been able to survive and grow to reproductive maturity is relatively well adapted to its location, so any progeny that are genetically identical to it are at least as well adapted as it is. Any progeny that are not genetically identical to the parent may or may not be well adapted to the conditions to which the parent is adapted.
If the environment is stable during several lifetimes, it is selectively advantageous for an organism to reproduce asexually by budding or sending out runners, thus producing new similarly adapted individuals. If the environment is not stable, however, such offspring may find themselves in conditions for which they are poorly adapted; if all are identical, all may die. Instability of the environment can result from many factors (e.g., landslides, avalanches, and road building in forests kill existing vegetation, opening up new, sunny sites that are good for quickly growing, sun-loving weedy plants). Irregular climatic events, such as unusually severe freezes, droughts, floods, or hurricanes, also disrupt plant communities. If all members of a species are equally susceptible to low temperatures, all may be killed by a rare freeze, but if the species is genetically diverse, some individuals may survive. Even though most die, the few survivors may be sufficient to repopulate the site (TABLE 9-1). A current concern is that we are adding so much carbon dioxide to the atmosphere that our climate is changing too rapidly for many plant and animal species to be able to adapt.
With asexual reproduction, progeny are never more fit than the parent, but during sexual reproduction, sex cells of one plant combine with those of one or several others, resulting in many new gene combinations. Sex cells are so small that many can be produced by a single plant, and many new combinations of genes can be “tested” rather inexpensively. For example, a single large tree can produce thousands of flowers and millions of pollen grains, each genetically unique, yet the tree uses only a few grams of carbohydrate, protein, and minerals (FIGURE 9-1). Similarly, thousands of egg cells can be produced using only a small amount of resources. The pollen from one plant can be blown or carried by pollinators to the flowers of hundreds or thousands of other plants, and one plant may receive pollen from numerous other individuals. The thousands of seeds produced by a single sexually reproducing plant represent thousands of natural genetic experiments. During seed and fruit maturation, those embryos with severely mismatched genes abort and use no further resources. The tree finally produces hundreds or thousands of fruits and seeds. The total reproductive effort may be a significant drain on the tree’s resources, but it produces numerous embryos, many of which are at least as genetically fit as the tree is and perhaps even more fit. In both stable environments and changing ones, sexual reproduction provides enough diversity of progeny that at least some are well adapted.
TABLE 9-1 Sexual and Asexual Reproduction


FIGURE 9-1 (A) These apple fruits have developed from flowers, and the seeds inside the fruits developed from egg cells inside the flowers. All egg cells produced by this tree have similar genes, but sperm cells were necessary to fertilize the eggs, and bees may have brought many types of sperm cells from many different apple trees. Although every seed here has the same maternal parent (the tree in the photo), it is possible that all the seeds have different paternal parents. The seeds are not genetically identical. (B) These anthers of a lily (Lilium) flower are each releasing thousands of pollen grains; because they were produced by meiosis with crossing over, each pollen grain is at least slightly different genetically from all the others produced by the same plant.
As a further example, think of sexual reproduction in humans: The children produced by a particular couple are variable, not identical to each other or to either parent. Some of the children may have a particularly advantageous combination of genes and be more healthy or intelligent or athletic or creative than either parent. Others may have combinations of genes that result in congenital problems such that the children survive only through medical help. Most children are more or less the same as the parents. The diversity is important.
Sexual reproduction also has negative aspects. Two individuals are required, and sex cells must move from one plant to another. In seed plants, pollen may be carried by wind, insects, and birds, but each results in the loss of many pollen grains or the need to produce nectar. Furthermore, potential sex partners may be widely scattered. For example, in a population of trees, those few individuals growing at the highest altitudes may have no neighbors, whereas those growing at lower altitudes have numerous close neighbors. The flowers of the highest individuals may receive no pollen and thus produce no seeds during some years. In contrast, plants that reproduce asexually can do so at any time, even when completely isolated. Some flowering plants are self-fertile and can undergo self-pollination, but they lose the benefit of receiving new genes from another plant.
Some plants reproduce both sexually and asexually FIGURE 9-2. Strawberries have flowers and sexual reproduction involving genetically diverse embryos and seeds, but they also spread rapidly and asexually by runners. Bamboos are perennial grasses that flower and set seed only occasionally (in some species, only once every 80 years), but their rhizomes grow vigorously and establish many new plants asexually. Kalanchoes produce large numbers of seeds each year, but they also produce such large numbers of plantlets along their leaf margins that they can be weeds in both nature and in greenhouses (Figure 9-2A).

FIGURE 9-2 (A) Kalanchoe plants are called maternity plants because they produce plantlets complete with stems, leaves, and roots along their leaf margins. Although hundreds of plantlets can be produced, all nuclear divisions are mitotic—duplication division—and thus, all of these plantlets are genetically identical to the parent; none is superior. (B) Chollas, species of Opuntia, have branches only weakly attached to the trunk. If an animal brushes against a branch, the spines stick to the animal and the branch is pulled from the plant. After the animal dislodges the branch, it roots and grows into a new plant. Long-distance distribution almost as extensive as that of seeds can occur. (C) All the leafless trees with white bark are quaking aspen (Populus tremuloides) in winter. Almost certainly, they are all parts of the same plant, each having grown from an adventitious shoot bud on a widely spreading root system. Some aspen clones fill entire valleys or hillsides. (D) All grapes used for wine are the same species (Vitis vinifera), but there are hundreds of varieties, each of which must be propagated vegetatively to ensure every plant produces grapes with exactly the same flavor. Here you can see an entire field planted just with cuttings. Every plant is a clone and each is genetically identical to the others. (E) The clonal nature of this dog cholla (Opuntia schottii) is easy to see: the oldest part of the plant was located at the center and then its branches began growing outward. The old, central parts have died, but the younger, newer branches on the edge continue to grow outward and the ring becomes larger.
Seeds, which are produced by sexual reproduction, often have a means of long-distance dispersal: Strawberries are eaten and the seeds later defecated; bamboo fruits and seeds are carried by winds. The consequence of this is that seeds may become widely scattered and germinate in numerous diverse sites, each site differing from the others in its microclimate, soil conditions, and exposure to predators and pathogens. Of course, a seed carrying an embryo with a combination of genes selectively advantageous for growth in a dry site may land in a wet site and not survive, but with the large numbers of seeds produced, it is statistically probable that some seeds will land in sites for which they are well suited.
In contrast, new plants that are produced asexually are usually not capable of long-distance dispersal; runners, rhizomes, and plantlets result in new plants that become established in the same microhabitat as their parent. After a single plant becomes established in a suitable site, by reproducing asexually it can quickly fill the area with replicas of itself, all of which are as fit as it is.
![]() Asexual Reproduction
Asexual Reproduction
Within angiosperms, numerous methods of asexual reproduction have evolved. One of the most common is fragmentation: A large spreading or vining plant grows to several meters in length, and individual parts become self-sufficient by establishing adventitious roots. If middle portions of the plant die, the ends become separated and act as individuals. Certain modifications improve the efficiency of fragmentation. In many cacti, branches are poorly attached to the trunk, and the plant breaks apart easily. The parts then form roots and become independent (Figure 9-2B). In some members of the saxifrage, grass, and pineapple families, plantlets are formed where flowers would be expected; these look like small bulbs and are called bulbils.
In willows and many thistles, adventitious shoot buds form on roots and then grow into plants. Adventitious buds may grow out even while the parent plant is still alive, and a small cluster of trees may in fact consist of just a single individual. A grove of aspens that covers several acres in Utah has been discovered to be a single plant (Figure 9-2C).
![]() Sexual Reproduction
Sexual Reproduction
Sexual reproduction in angiosperms involves flowers, which produce the necessary cells and structures. To understand flower structure, one must first understand the plant life cycle.
The Plant Life Cycle
The life cycle of mammals such as humans is simple: Diploid adults have sex organs that produce haploid sex cells called gametes, either sperms or eggs, by meiosis. Individuals that produce sperms are called males, of course, and individuals that produce eggs are females. One sperm and one egg are brought together, forming a new single diploid cell, the fertilized egg or zygote, which then grows to become a new individual that is diploid and resembles its parents.
In plants, the life cycle is more complex. The plants you are familiar with—trees, shrubs, and herbs—are all just one phase of the plant life cycle, called the sporophyte phase or sporophyte generation. A critical factor is that sporophytes are always diploid, like most adult animals, and they have organs (located in the flowers in angiosperms) with cells capable of undergoing meiosis. In animals, meiosis results in haploid gametes, but in plants, it results in haploid spores (FIGURE 9-3). The difference between gametes and spores is great: Gametes can fuse with other gametes in a process called syngamy or fertilization, thereby producing the diploid zygote. A gamete that does not undergo syngamy dies because it cannot live by itself and usually cannot grow into a new, haploid individual (unfertilized eggs of some insects such as bees are exceptional and develop into sterile workers).
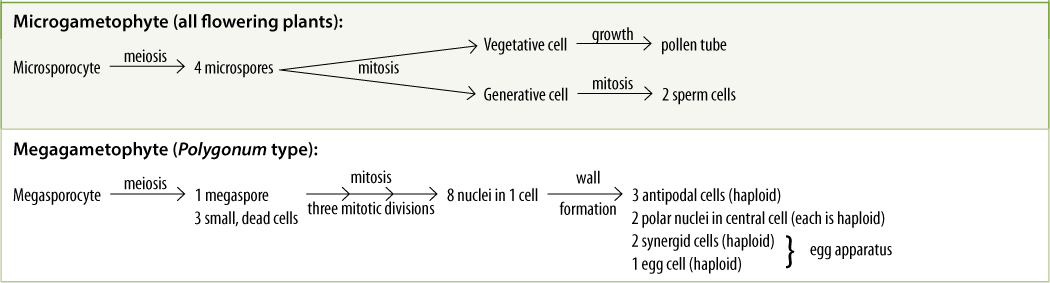
Plant spores are just the opposite: They cannot undergo syngamy, but each undergoes mitosis and grows into an entire new haploid plant called a gametophyte. It is called a gametophyte because it is the plant (-phyte) that produces gametes (gameto-). During sexual reproduction, when a sporophyte reproduces, it does not produce a new diploid plant like itself but rather a haploid plant (simple plants like hornworts, liverworts, and mosses are gametophytes). Furthermore, in all vascular plants, a haploid gametophyte does not even remotely resemble a diploid sporophyte. It is a tiny mass of cells with no roots, stems, leaves, or vascular tissues, but it is an entire plant (FIGURE 9-4). Gametes are formed by the haploid plants by mitosis, not meiosis. The gametes then undergo syngamy, forming a zygote that grows into a new, diploid sporophyte, and the life cycle is complete (FIGURE 9-5).
Mammalian gametes are of two types: small sperm cells (microgametes) that swim and large eggs (megagametes) that do not. This is also true of many plants and is known as oogamy. In oogamous plants, just as in oogamous mammals, sperms are produced by one type of individual and eggs by a different type of individual; hence, there are “male” or microgametophytes and “female” or megagametophytes (Figure 9-4). The two types of gametophytes have grown from two types of spores: microgametophytes from microspores and megagametophytes from megaspores. Having two types of spores is known as heterospory. Typically, just one kind of sporophyte occurs in a life cycle, and it produces both microspores and megaspores.
A life cycle like this, with two generations—sporophyte and gametophyte—is said to be an alternation of generations (Figure 9-5). Because gametophytes do not resemble sporophytes at all, this is an alternation of heteromorphic generations. This is a complex life cycle, with at least three distinct plants (one sporophyte and two gametophytes). The human life cycle, like that of all other animals, does not have anything equivalent to the haploid generation.
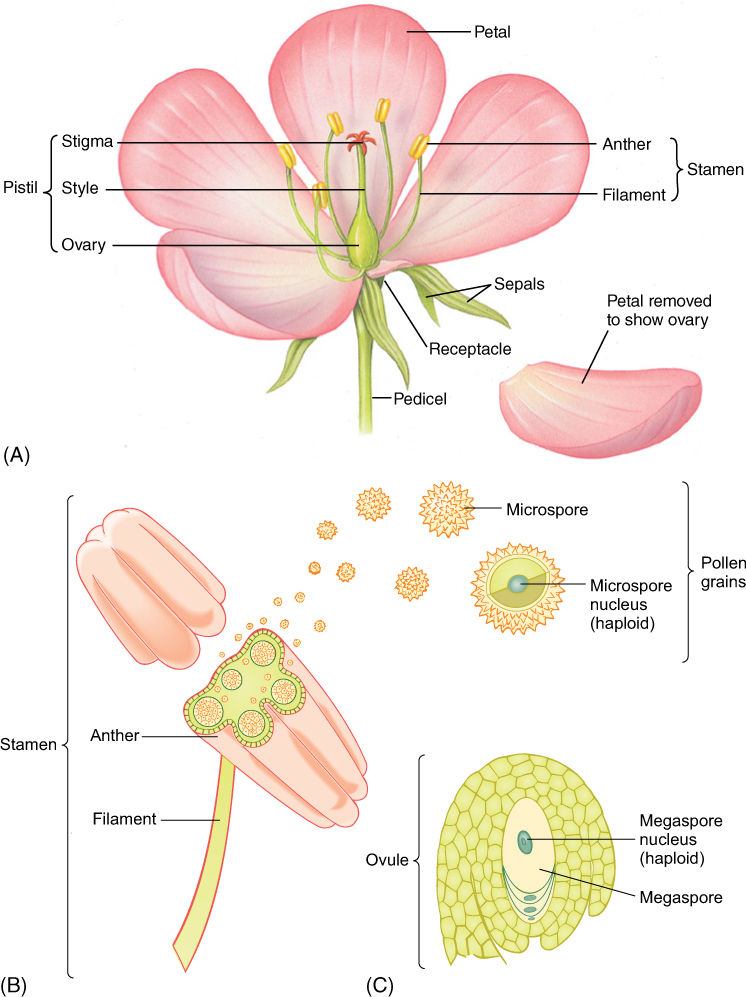
FIGURE 9-3 (A) Most flowers have four types of structures: sepals, petals, stamens, and a pistil. The pistil shown here is actually composed of five parts called carpels that have merged into one structure. (B) In the anthers of stamens, the central cells undergo meiosis and each produces four daughter cells called microspores or pollen grains. (C) In the flower’s central organs, the carpels are ovules, each containing only one cell that undergoes meiosis; often three of the daughter cells die and the one survivor becomes the megaspore.
Flower Structure
A flower is basically a stem with leaf-like structures, so almost everything discussed for vegetative shoots also applies to flowers. Flowers never become woody; secondary growth does not occur in flowers.
The flower stalk is a pedicel, and the very end of the axis, where the other flower parts are attached, is the receptacle (Figure 9-3). There are four types of floral appendages: sepals, petals, stamens, and carpels. Most flowers have all four types and are complete flowers. They typically have three, four, five, or more appendages of each type; for example, lilies have three sepals, three petals, three stamens, and three carpels. It is not uncommon for flowers of certain species to lack one or two of the four basic floral appendages, thus being called incomplete flowers. Flowers of Begonia (FIGURE 9-6) are incomplete, lacking petals but having either stamens or carpels; flowers of pigweed have all parts except petals.
Sepals
Sepals are the lowermost and outermost of the four floral appendages. They are modified leaves that surround and enclose the other flower parts as they mature. Sepals are typically the thickest, toughest, and waxiest of the flower parts. They protect the flower bud as it develops, keeping bacterial and fungal spores away, maintaining a high humidity inside the bud, and deterring insect feeding (FIGURES 9-7A and 9-7B). Sepals also protect the flower from nectar-robbing insects and birds. If flower buds develop in a protected position such as beneath a spiny cover or surrounded by regular leaves and branches, sepals are less important for protection and may be small or absent. It is not uncommon for sepals to be colorful (petaloid) and help attract pollinators. All the sepals together are referred to as the calyx.
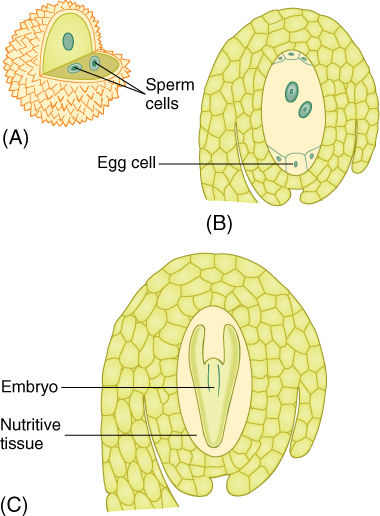
FIGURE 9-4 All seed plants produce two types of gametophytes. (A) Microspores (pollen grains) develop into microgametophytes (also called pollen grains). The microgametophyte body is so small that it has just three cells and fits inside the pollen cell wall. In angiosperms, pollen grains convert to microgametophyts before the anther opens and sheds the pollen. The microgametophyte produces two sperm cells located within its own protoplasm. (B) The megaspore develops into a megagametophyte. It is slightly larger than a microgametophyte and has seven cells, one of which has two nuclei. One of the cells is the egg. The megaspore and megagametophyte are never released from the flower of the parent sporophyte, so they appear to be just sporophyte tissues, not one plant growing inside another. (C) After one sperm cells fertilizes the egg cell, the new egg cell nucleus is diploid and the cell is a zygote. It develops, by mitosis, into a new sporophyte, shown here as the embryo in an immature seed. The entire ovule develops into a seed; the ovary develops into a fruit.
Petals
Above the sepals on the receptacle are petals, which together make up the corolla. Sepals and petals together constitute the flower’s perianth. Petals are also “leaf-like,” being broad, flat, and thin, but they differ from leaves in that they contain pigments other than chlorophyll, have fewer or no fibers, and tend to be thinner and more delicately constructed (FIGURES 9-7C and 9-7D). Petals are important not merely in attracting pollinators, but rather the correct pollinators. Each plant species has flowers of distinctive size, shape, color, and arrangement of petals, allowing pollinators to recognize specific species. Sexual reproduction results only if pollen is carried to other flowers of the same species; it cannot occur efficiently if pollen is carried to other plants indiscriminately. If a flower has a distinctive pattern and offers a good reward such as nectar or pollen (FIGURE 9-8), the pollinator is likely to search for and fly to other flowers with the same pattern, thereby enhancing cross-pollination; mutations are advantageous selectively if they cause flowers to have characters easily recognized by their pollinators (FIGURE 9-9). In addition to visible colors, many flowers have pigments that absorb ultraviolet light, creating patterns only insects can see. Without light, colors cannot be seen, and night-blooming species have white flowers lacking pigments. Their petals produce volatile fragrances, and insects and bats follow the aroma gradient to the flower.
Pollen is carried by wind in many species and by water in a few. Typically, petals do not develop in wind-pollinated species: They cannot attract wind so mutations that inhibit their differentiation prevent plants from wasting resources constructing nonfunctional structures (see Figure 9-27).
Stamens
Above the petals are stamens, known collectively as the androecium. Stamens are frequently referred to as the “male” part of the flower because they produce pollen, but technically, they are not male because the flower, being part of the sporophyte, produces spores, not gametes. Only gametes and gametophytes have sex.
Stamens have two parts, the filament (its stalk) and the anther, where pollen is actually produced. As part of the sporophyte, the anther is composed of diploid cells, and in each anther, four long columns of tissue become distinct as some cells enlarge and prepare for meiosis (FIGURE 9-10A). These microspore mother cells or microsporocytes continue to enlarge and then undergo meiosis, each producing four microspores (FIGURE 9-10B and TABLE 9-2). Neighboring anther cells, in a layer called the tapetum, act as nurse cells, contributing to microspore development and maturation. Microspores initially remain together in a tetrad, but later separate, expand to a characteristic shape, and form an especially resistant wall. They are then called pollen. The anthers open (dehisce) along a line of weakness and release the pollen.
The wall of a pollen grain is a cell wall; however, it is quite complex structurally. It has an inner layer called the intine, composed of cellulose, and an outer layer called the exine that consists of the polymer sporopollenin. It has one or several weak spots, germination pores, where the pollen opens after it has been carried to the stigma of another flower. Sporopollenin is remarkably waterproof and resistant to almost all chemicals; it protects the pollen grain and keeps it from drying out as it is being carried by wind or animals. The exine can have ridges, bumps, spines, and numerous other features so characteristic that each species has its own particular pattern (FIGURE 9-11). In many cases, it is possible to examine a single pollen grain and know exactly which species of plant produced it. Because sporopollenin is so resistant, pollen grains and their characteristic patterns fossilize well. By examining samples of old soil, botanists can determine exactly which plants grew in an area at a particular time in the ancient past.
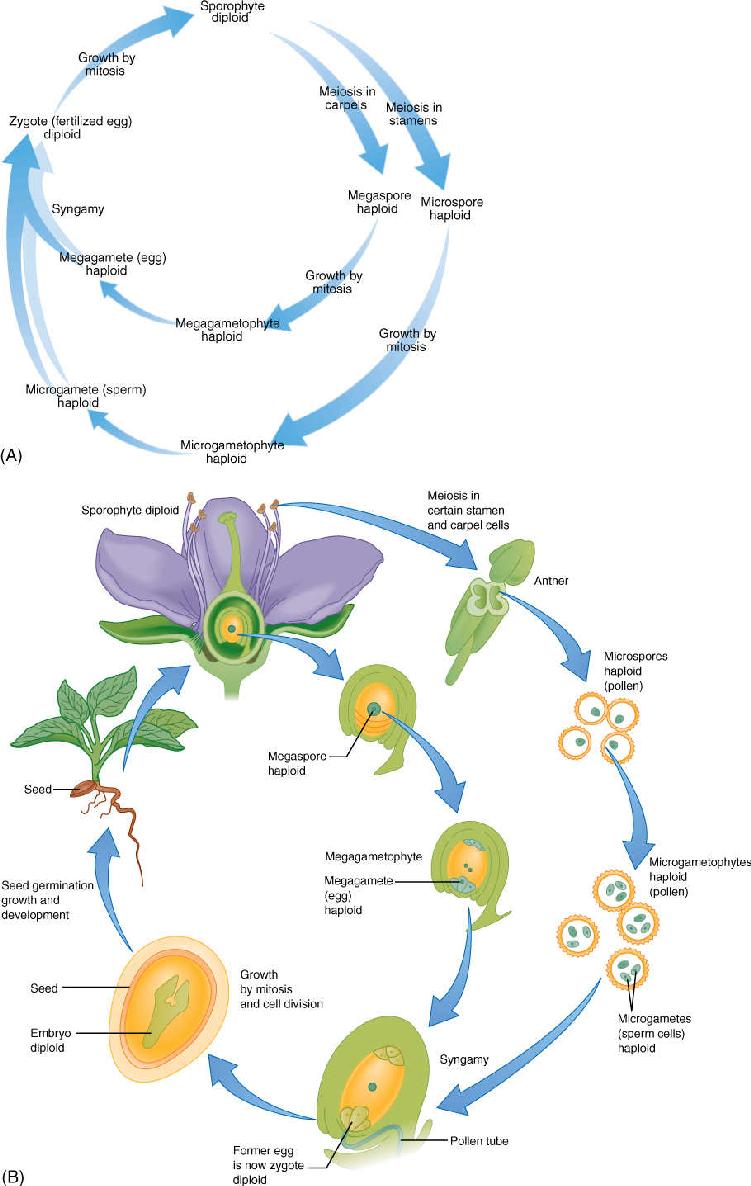
FIGURE 9-5 (A) Diagram of the life cycle of an angiosperm. (B) A flowering plant life cycle showing the actual structures involved.

FIGURE 9-6 In most species of flowering plants, the stamens and carpels are produced together in the same plant, but in a few species like this Begonia, stamens occur in flowers without carpels (A), whereas carpels occur in stamen-less flowers (B). In species in which stamen-less flowers occur on one plant and carpel-less flowers occur on a separate one, the two types of plants look so much alike that, in contrast to mammals, they usually cannot be identified as staminate (having stamens) or carpellate (having carpels) without looking at the flowers. In both types of Begonia flower shown here, the pink structures look like petals but are instead modified sepals; true petals are absent.

FIGURE 9-7 (A) The sepals of this rose form a tight covering over the rest of the flower as it develops, protecting the inner parts. When the microspores (pollen) and megaspores (in the ovules) are ready, the sepals bend outward and the flower opens. (B) Sunflowers and daisies are actually clusters of many small flowers that together have the appearance of a single flower. In this case, the entire cluster of flowers is protected by a set of bracts that look and function like sepals. It is not necessary for the individual flowers to have protective sepals. (C) Petals, like these of the mallow, are leaf like in being thin and flat, but in most species, they are less leaf like than sepals. (D) Leaves and petals of Arbutus can be compared here; these petals are highly modified, and the five petals of each flower have fused together, forming a tubular corolla.

FIGURE 9-8 Some flowers do not produce nectar; the pollinator eats part of the stamens and pollen instead. This is more expensive metabolically for the plant because pollen is rich in protein, whereas nectar is composed of carbohydrate. In these flowers of cannonball tree (Couropita), the lower stamens are modified for edibility; they are large and showy and produce little or no pollen. The pollen-producing stamens are small and inconspicuous, easily overlooked by pollinators.

FIGURE 9-9 Distinctive patterns like this are known as nectar guides; they direct the pollinator to the nectar and position the animal to pick up pollen.
Carpels
Carpels constitute the gynoecium, located at the highest level on the receptacle (FIGURE 9-12A). Carpels have three main parts: (1) a stigma that catches pollen grains, (2) a style that elevates the stigma to a useful position, and (3) an ovary where megaspores are produced. A flower can have zero (some imperfect flowers) to many carpels; usually they are fused together into a single compound structure, frequently called a pistil. Inside the ovary are placentae (singular, placenta), regions of tissue that bear small structures called ovules (FIGURE 9-12B). Ovules have a short stalk, called a funiculus, that carries water and nutrients from the placenta to the ovule by means of a small vascular bundle. The ovule has a central mass of parenchyma called a nucellus. Around the nucellus are two thin sheets of cells (integuments) that cover almost the entire nucellus surface, leaving only a small hole (micropyle) at the top (FIGURE 9-12C). As in anthers, some nucellus cells, usually only one in each ovule, enlarge in preparation for meiosis; these are megaspore mother cells or megasporocytes. After meiosis, usually three of the four megaspores degenerate, and only one survives, becoming very large by absorbing the protoplasm of the other three. Megaspores differ from microspores (pollen) because the ovule and the carpel do not dehisce and the megaspore remains enclosed inside the carpel.
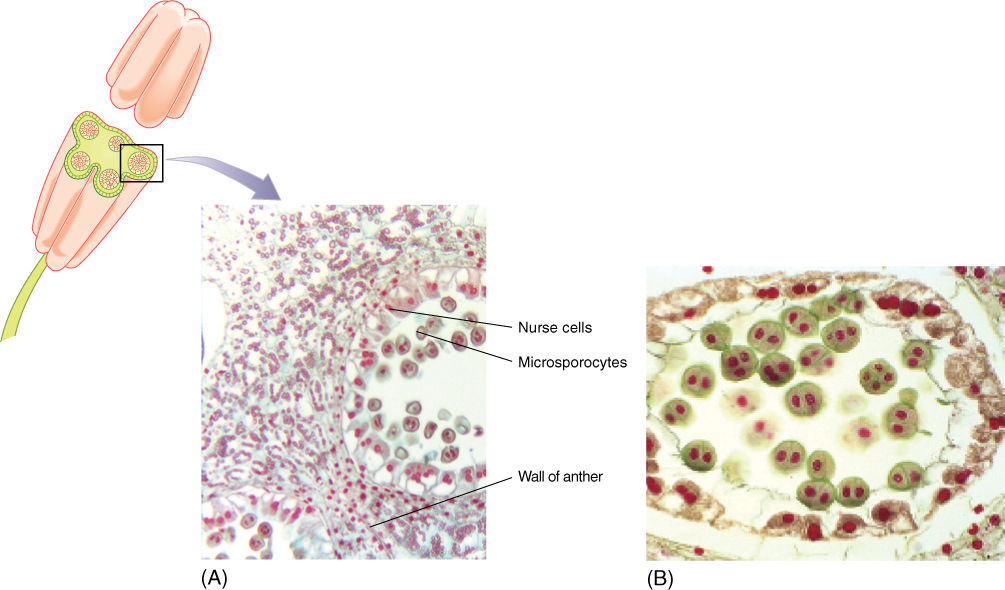
FIGURE 9-10 (A) Transverse section through an anther of a lily, showing microsporocytes in the meiotic division that will produce haploid microspores. The surrounding cells constitute the tapetum and aid in the maturation of the pollen grains (×200). (B) After meiosis, the four microspores remain together temporarily, but as they develop their specialized pollen cell wall, they usually separate from each other (×500).
TABLE 9-2 Stamen and Carpel Structure

An ovule develops into a seed after its egg is fertilized, and the surrounding ovary develops into a fruit. Each ovary might have either one or many placentae, each bearing one or many ovules; ovaries with just a single ovule develop into fruits with a single seed (such as avocado or peach). Manyseeded fruits (e.g., tomato, cantalope, and squash) have numerous placentae, each with many ovules. Orchid pistils always consist of three carpels, each of which might have tens of thousands of ovules, and some orchid fruits are estimated to contain up to one million tiny seeds.
Aside from bearing ovules, the rest of the carpel is at least somewhat leaflike. Figure 9-12A shows three carpels fused together as a pistil; the outermost layer is basically leaf lower epidermis and typically has stomata. The innermost layer is upper epidermis, and all middle layers are mesophyll and vascular bundles.
Gametophytes
Microgametophyte
Microspores develop into microgametophytes. In all angiosperms, each microgametophyte is very small and simple, consisting of at most three cells located within the original pollen cell wall (FIGURE 9-13). The microspore nucleus migrates to the side of the pollen grain and lies next to the wall. There it divides mitotically, producing a large vegetative cell and a small lens-shaped generative cell, which subsequently divides and forms two sperm cells. The entire microgametophyte consists of the vegetative cell and the two sperm cells (the microgametes). Although extremely simple, this is a full-fledged plant; see Chapters 20 and 21 for examples of gametophytes that are more complex than these of angiosperms and that are obviously complete plants.
In approximately 30% of angiosperm species, formation of sperm cells occurs even while pollen is still located within the anther. In the majority of angiosperm species, the pollen is released from the anther at about the time the generative cell has formed, and sperm cells are not produced until after the pollen has been carried to a stigma.
After a pollen grain lands on a stigma, it germinates by producing a pollen tube that penetrates into the loose, open tissues of the stigma (FIGURE 9-14). The pollen tube absorbs nutrients from the stigma and grows downward through the style toward the ovary. The microgametophyte (the pollen tube) is protected and nourished by the style tissue. As the pollen tube grows downward, it carries the sperm cells to the ovule. Almost all of the pollen cytoplasm is located at the tip of the pollen tube; the rest of the tube and the pollen grain are filled with a giant vacuole. Pollen grains are too small to store much starch, fat, or protein; if they could not absorb nutrients from the style, the pollen tubes could not grow long enough to reach the ovules.
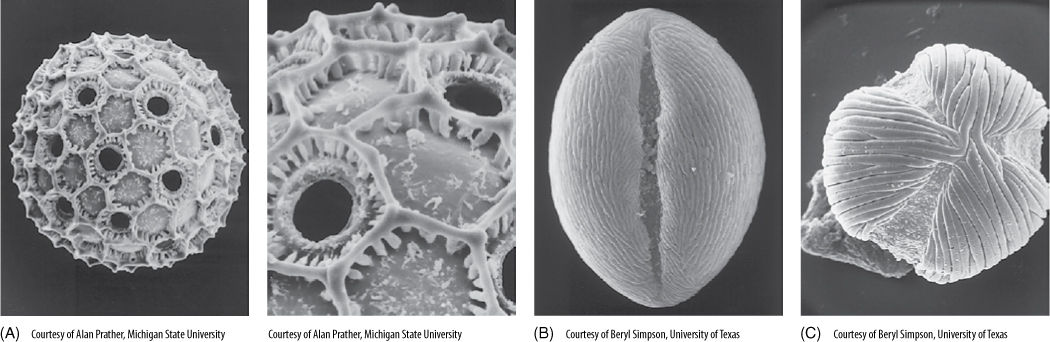
FIGURE 9-11 (A) Pollen grains of Cobaea, with the pollen wall forming hexagonal ridges. The numerous holes are germination pores where a pollen tube can emerge after the pollen lands on a stigma (×2,000). (B) Pollen of Lycium has a single long groove from which the pollen tube emerges (×4,000). (C) Pollen of Macrolobium has three germination grooves (×4,000).
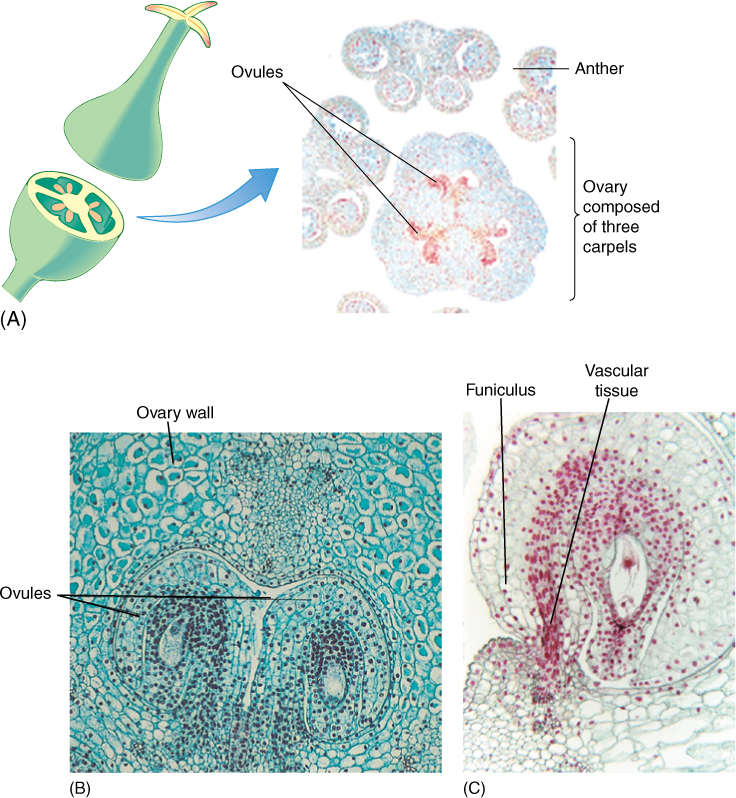
FIGURE 9-12 (A) In lily, the pistil consists of three carpels fused together. This ovary has been cut across to reveal the three ovary locules or chambers. In each chamber, two ovules are visible (×15). (B) Ovules are attached to the placenta by their funiculus, which contains both xylem and phloem. The central part of the ovule is the nucellus with (usually) just one megasporocyte, unlike anthers, which have numerous microsporocytes (×50). (C) Magnification of a single ovule. Surrounding the nucellus are the integuments. Growth of the ovule has been by mitosis, but now one cell, the megasporocyte—megaspore mother cell—is undergoing meiosis (×120).
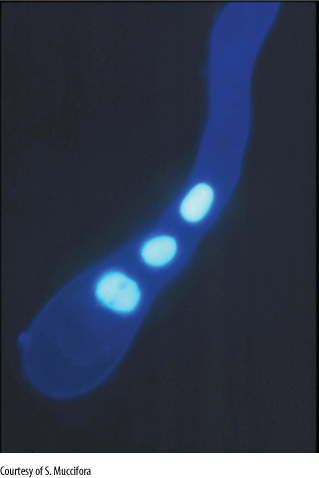
FIGURE 9-13 This is the tip of a growing pollen tube. It was treated to make the nuclei fluoresce (glow) so that they can be found easily despite being so small. The lowest nucleus is probably the tube nucleus, the upper two are probably sperm cell nuclei, each located within a sperm cell. The two sperm cells themselves are located within the protoplasm of the pollen tube (×5,000).
Megagametophyte
Within the ovule the surviving megaspore develops into a megagametophyte. In one type of development, the nucleus undergoes three mitotic divisions, producing two, four, and then eight haploid nuclei all in a single, undivided cell (FIGURE 9-15). The technical term for a multinucleate megagametophyte is embryo sac. The nuclei migrate through the cytoplasm, pulled by microtubules, until three nuclei lie at each end and two in the center. Walls then form around the nuclei, and the large, eight-nucleate megaspore becomes a megagametophyte with seven cells, one of which is binucleate. The seven cells are one large central cell with two polar nuclei, three small antipodal cells, and an egg apparatus consisting of two synergids and an egg (the megagamete) (TABLE 9-3). Like the microgametophyte, the megagametophyte is a distinct plant. As with the pollen, the megagametophyte obtains all of its nourishment from the parent sporophyte.
Plants and People
BOX 9-1 Flowers, Fruits, Seeds, and Civilization
Flowers and the fruits and seeds that result from them have always been important to us. One of the first hominids—animals on our line of evolution—was Australopithecus africanus; fossils of its teeth, 3 million years old, show that it was adapted to eating plants. Homo habilis lived only 1.8 million years ago and likewise had a diet strongly dependent on plants. Our species, Homo sapiens, appeared evolutionarily about 500,000 years ago and survived by hunting game and collecting fruits, seeds, and edible roots.
Approximately 11,000 years ago, a momentous change occurred. Small groups of humans began cultivating plants rather than just gathering them (TABLE B9-1). This happened in the Middle East, an area that is now Iran, Iraq, and Syria, also known as the Fertile Crescent. The plants were common, wild species such as wheat, barley, peas, and lentils. Farming required major changes in human society: People had to stay in one place to tend and protect crops rather than follow herds of game animals. Permanent villages were necessary, as were the rules and regulations needed when people live together in close proximity. Land, huts, and harvests are tempting targets for thieves, and thus, defense, both individual and collective, became a necessity—government had to be created.
Civilization advanced rapidly. Neolithic (New Stone Age) agricultural societies were widespread by 6000 BCE, and the oxdrawn plow was in use as early as 4500 BCE. New plants were cultivated: olives, date palms (FIGURE B9-1), and grapes for eating and for wine. Independently, the peoples of Southeast Asia domesticated rice and soybeans; in the New World the Incas, Mayans, and Aztecs cultivated potatoes, corn, tomatoes, beans, squashes, cocoa, pineapples, and peanuts.
TABLE B9-1 Locations Where Plants Were Domesticated
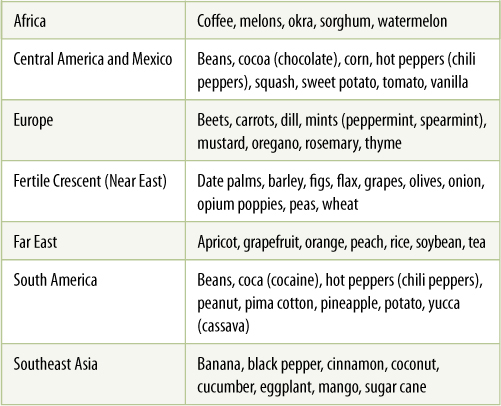

FIGURE B9-1 Images like this occur frequently in ancient Sumerian art. The person is either a priest or a king, and the object he faces is a very strange plant. In earlier depictions the plant is recognizable as a date palm, but through the years this important food tree became a symbol for life itself and artists began to draw it more stylized and with mystical symbols surrounding it. The person is holding a cone-like object in one hand and a pail in the other. Date palms grow as carpellate (“female”) plants and staminate (“male”) plants. To ensure a good crop, pollen can be collected in a pail and then applied to the carpellate plants with a brush to increase the rate of pollination and fruit production. The scene, carved before 2000 BCE, indicates that the very earliest farming societies understood this sophisticated aspect of botany.
The importance of fruits and seeds to the survival of both individual humans and societies is reflected in the prominent position they were given in early art. Ancient Egyptians carved likenesses of date palms, barley, and wheat as long ago as 3000 BCE, and on Crete in 1800 BCE, Minoan artists depicted, in addition to the plants already mentioned, figs, saffron crocus, pomegranate, and lupine. They also showed several plants that must have been grown purely as ornamentals: lilies, myrtle, narcissus, and roses.
As societies become wealthier, time and resources become available for leisure and pleasurable pursuits. Gardens dedicated to beautiful, fragrant flowers, not food crops or medicinal plants, arose early. Expeditions in search of exotic ornamental plants are described in the oldest epic poem in the world, the story of Gilgamesh of Sumer, and an ancient Egyptian monument, the Palermo Stone, records a plant-gathering expedition by King Snefru in 2900 BCE. Inventories list the extensive plant-collecting trips and gardens of Ramses III.
Growing beautiful flowers and enjoying exotic fruits have never lost their popularity; after the Spanish discovered America, the initial exploitation was for gold and silver, but there soon followed expeditions dedicated solely to gathering plants, especially ornamental ones. Two of the many famous plant explorers sent to the United States are David Douglas, who explored the Pacific Northwest for the Horticultural Society of London in the 1820s, and the father and son team, André and François-André Michaux, sent by the government of France in 1785.
Even today, gardening and growing flowers for pleasure are considered by many an essential part of our civilization. As technology increases, cultivating flowers, or at least having a potted plant on a window sill, seems to maintain contact with our past.
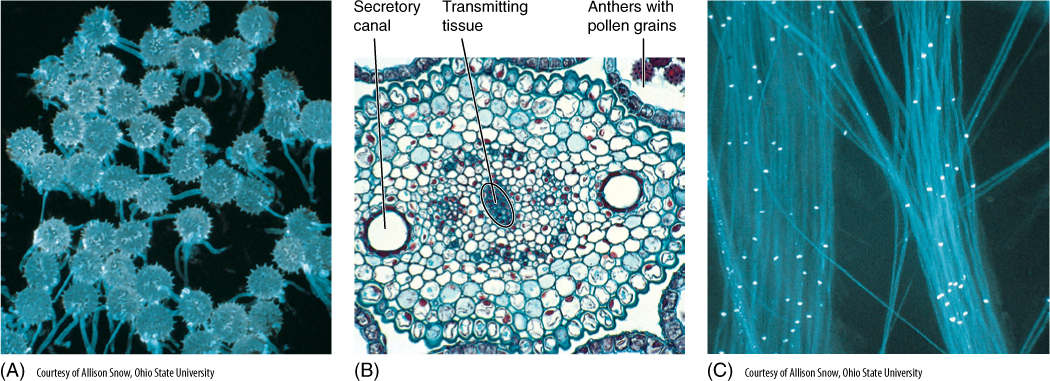
FIGURE 9-14 (A) Shortly after landing on a stigma of the correct species, a pollen grain may germinate by pushing out part of the wall as a pollen tube. This penetrates the loose tissues of the stigma and style. This was photographed with ultraviolet light, which causes the pollen wall and callose to fluoresce and shine (× 50). (B) Tissues of the style allow the pollen tubes to grow through them easily. Some styles are hollow and lined with a rich, nutritious transmitting tissue; other styles are solid but also have transmitting tissue. Sunflower (Helianthus) style (× 80). (C) These pollen tubes have penetrated deep into the style. The bright spots are callose plugs, which seal off the protoplasm at the tip from the empty parts of the pollen tube (× 200).
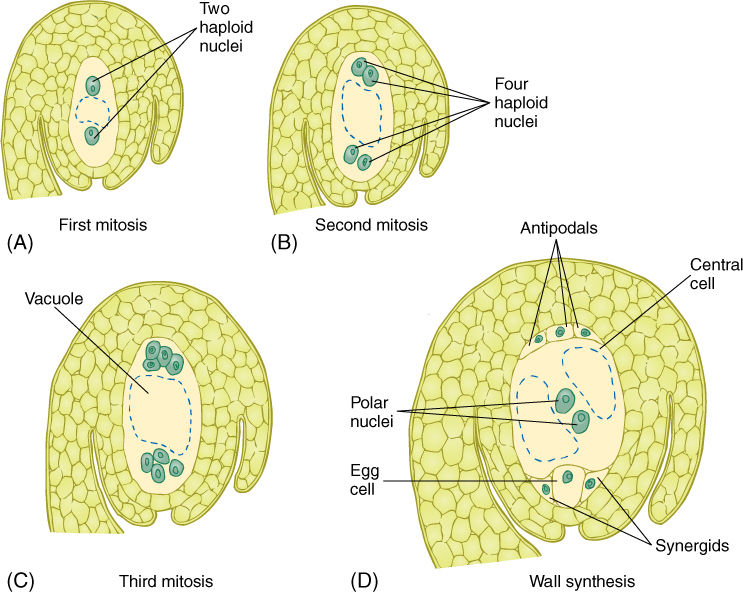
FIGURE 9-15 As a megaspore develops into a megagametophyte, three sets of mitosis occur (A) to (C) without any cytokinesis; thus, the megagametophyte is temporarily an eight-nucleate coenocyte (C). Then nuclei migrate; cytoplasm accumulates around each, and walls are established (D). Much of the egg is not covered by wall. The central cell is binucleate and is mostly vacuole.
TABLE 9-3 Gametophyte Development
Fertilization
Syngamy of sperm and egg involves both plasmogamy, fusion of the protoplasts of the gametes, and karyogamy, fusion of the nuclei. As a pollen tube grows downward through the style toward the ovule, it is guided to the ovule’s micropyle by some means. It penetrates the nucellus and reaches the egg apparatus, then enters one synergid. The pollen tube tip bursts and releases both sperm cells, one of which migrates through the synergid protoplasm toward the egg. As it does so, the sperm cell’s plasma membrane breaks down, and it loses most of its protoplasm. The sperm nucleus enters the egg, then is drawn to and fuses with the egg nucleus, establishing a diploid zygote nucleus.
Because the sperm sheds its protoplasm as it passes through the synergid, it contributes only its nucleus with the set of nuclear genes during karyogamy. The sperm does not carry mitochondria or plastids into the egg, and thus, organellar genes from the pollen parent are rarely inherited by the zygote. Instead, the mitochondrial and plastid genes of the embryo are inherited only from the ovule parent. Conifers are different: Their sperm cells retain their plastids while destroying their mitochondria. The zygote inherits mitochondria and a nucleus from the egg, a nucleus and plastids from the sperm cell.
In angiosperms only, the second sperm nucleus released from the pollen tube migrates from the synergid into the central cell. It undergoes karyogamy with both polar nuclei, establishing a large endosperm nucleus that is triploid, containing three full sets of genes. Because both sperm nuclei undergo fusions—one with the egg nucleus and the other with the polar nuclei—the process is called double fertilization. The endosperm nucleus is extraordinarily active and begins to divide very rapidly by mitosis, with cell cycles lasting only a few hours.
The endosperm nucleus initiates a dynamic cytoplasm and the central cell enlarges enormously, usually without cell division, into a huge cell with hundreds or thousands of nuclei (see FIGURE 9-16). Finally, nuclear division stops, and dense cytoplasm gathers around the nuclei. Walls are constructed, thus forming cells. An example of this is a coconut full of “milk.” The hollow center of the coconut is one single cell, and the milk is its protoplasm. The white coconut “meat” is the region where nuclei form cells. A green, immature coconut is full of milk but has almost no meat; as it ripens, the coenocytic milk is converted to a thick layer of cellular meat. All of this tissue, both coenocytic and cellular, is called endosperm, and it nourishes the development of the zygote. No other megagametophyte forms as much endosperm as a coconut does. More typical examples are grains such as wheat, rice, oats and corn, in which most of the grain is endosperm. Endosperm of these also pass through a milk stage, but by the time of harvest, the endosperm has become cellular, starch-filled, and dry enough to be hard (but if corn is picked for corn on the cob, it is collected just as endosperm is converting from coenocytic to cellular).
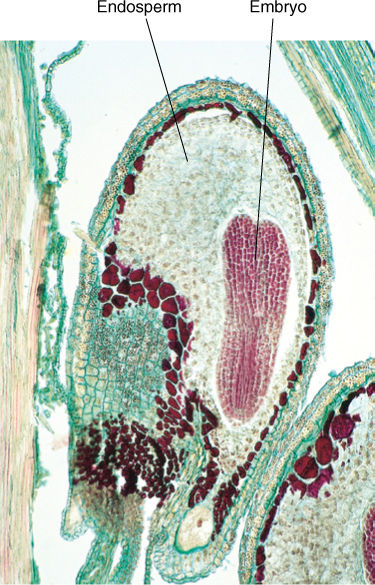
FIGURE 9-16 In many species, endosperm development is accompanied by some cytoplasmic division, and the endosperm is therefore a mass of multinucleate “cells,” each with variable amounts of protoplasm and nuclei and with irregular shapes. This is the only plant tissue in which nuclear and cytoplasmic divisions are so poorly coordinated, possibly because the endosperm is just a temporary tissue that is consumed by the embryo before or soon after seed germination (×180).
Embryo and Seed Development
As the endosperm nucleus proliferates, the zygote also begins to grow, but always by both nuclear and cellular divisions; a co-enocytic stage never occurs in the embryo. The zygote grows into a small cluster of cells, part of which later becomes the embryo proper, and the other part becomes a short stalk-like structure, the suspensor, which pushes the embryo deep into the endosperm (FIGURE 9-17). The suspensor is usually delicate and ephemeral in angiosperms; it is crushed by the later growth of the embryo and is not easily detectable in a mature seed.
Cells at one end of the suspensor continue to divide mitotically, developing into an embryo. The cells at first are arranged as a small sphere, the globular stage. The end of the embryo farther from the suspensor initiates two primordia that grow into two cotyledons in basal angiosperms and eudicots (FIGURES 9-18 and 9-19), such as beans and peanuts. While young, the cotyledon primordia give the embryo a heart shape; this is the heart stage. In monocots such as corn, only one cotyledon primordium grows out. “Dicot” is an abbreviation of “dicotyledon,” those plants whose embryos have two cotyledons. Monocots are monocotyledons, plants with only a single cotyledon on their embryos. Conifers like pine have numerous cotyledons.
Later, in the torpedo stage, the embryo is an elongate cylinder: A short axis is established, consisting of radicle (embryonic root), epicotyl (embryonic stem), and hypocotyl (the root/shoot junction). Finally, vascular tissue differentiates within the embryo. The epicotyl may bear a few small leaves, and the radicle often contains several primordia for lateral roots in its pericycle. Once mature, the embryo becomes quiescent and partially dehydrates, and the funiculus may break, leaving a small scar, the hilum. In green peas, the two halves of each pea are the two cotyledons, and the stalk attaching each pea to the pod (the fruit) is the funiculus.
In most basal angiosperms and eudicots, cotyledons store nutrients used during and after germination; during embryo development, the cotyledons become thick and filled with starch, oil, or protein, whereas the endosperm, which is supplying the nutrients, shrinks. When the seed is mature, the cotyledons are large, and the endosperm may be completely used up. We are eating mostly cotyledons when we eat beans, peas, peanuts, almonds, pecans, and other seeds that easily separate into two halves.
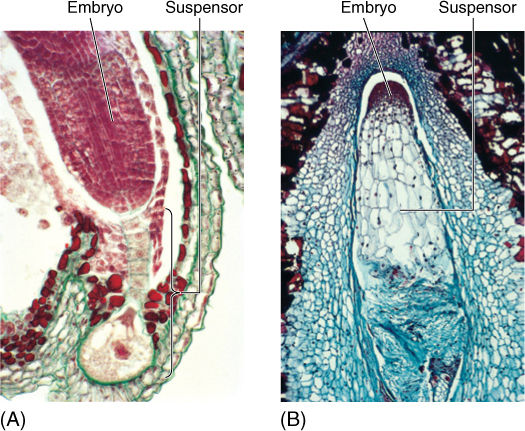
FIGURE 9-17 (A) and (B) In many species, endosperm development is accompanied by some cytoplasmic division, and the endosperm is therefore a mass of multinucleate “cells,” each with variable amounts of protoplasm and nuclei and with irregular shapes. This is the only plant tissue in which nuclear and cytoplasmic divisions are so poorly coordinated, possibly because the endosperm is just a temporary tissue that is consumed by the embryo before or soon after seed germination (× 180).
In monocots, the one cotyledon generally does not become thick and full; instead, the endosperm remains and is present in the mature seed. When eating cereals such as wheat, rice, oats, and corn, we are eating almost purely endosperm. During germination, the cotyledon acts as digestive/absorptive tissue, transferring endosperm nutrients to the embryo. Some eudicots are intermediate: Cotyledons store some starch and protein, but a considerable amount of endosperm remains in the seed, and both methods of nutrition are used during germination. A mature seed in which endosperm is rather abundant is an albuminous seed (Figure 9-19). If endosperm is sparse or absent at maturity, the seed is exalbuminous.
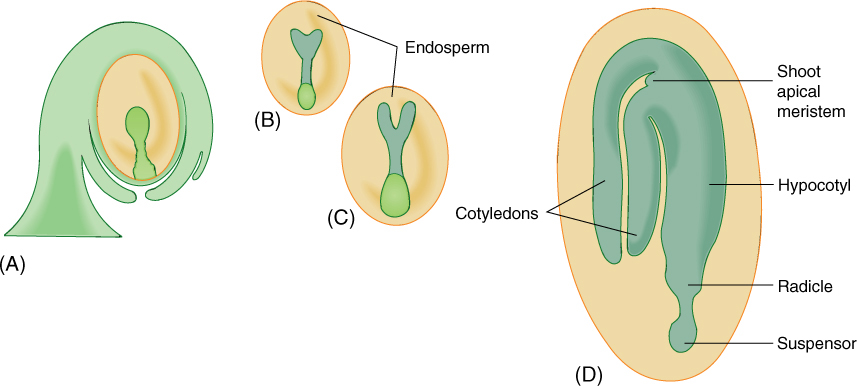
FIGURE 9-18 Embryo development. (A) Globular stage. (B) Heart stage. (C) Torpedo stage. Embryo root (radicle), cotyledons, and hypocotyl are present. At this stage, the first xylem and phloem may become distinguishable in the hypocotyl. (D) Mature embryo. A shoot apical meristem is shown; in some species, even some leaf primordia and a small stem are present.
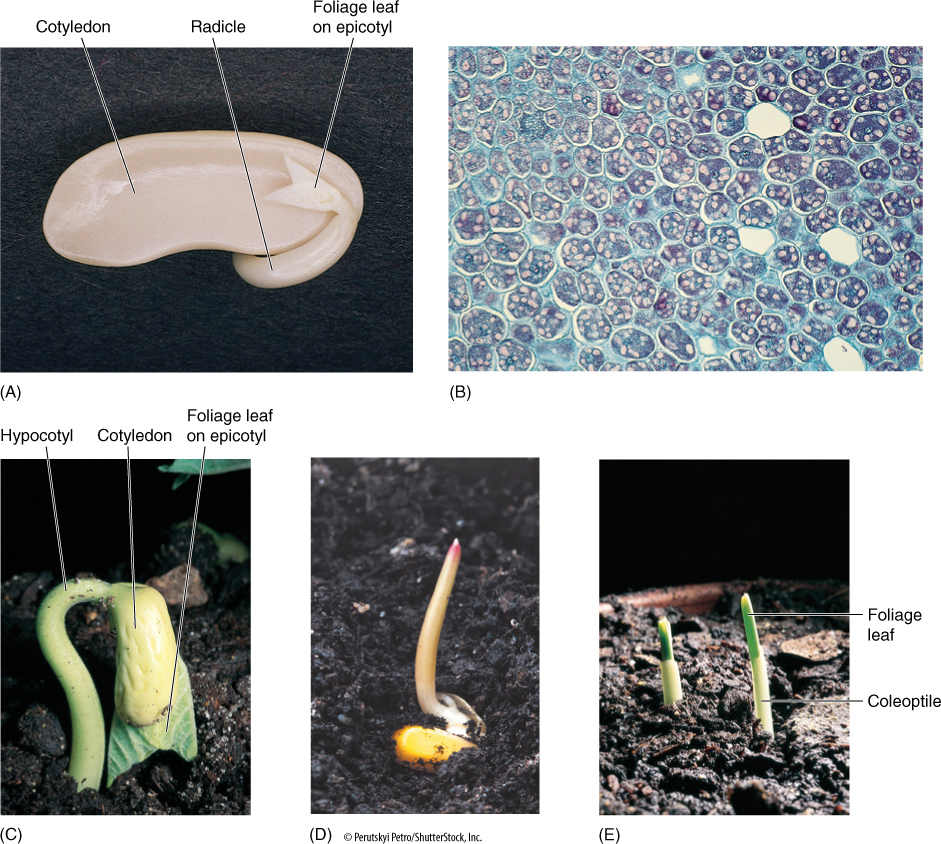
FIGURE 9-19 (A) This bean seed has begun germinating; the radicle has extended and will develop into a taproot. The two cotyledons (one was removed) were digesting their carbohydrates and proteins and transporting nutrients to the root and shoot apical meristems. Beans have a well-developed epicotyl with several partially expanded leaves; one is visible here. (B) These cells of bean cotyledon are full of starch (stained pink) and reserve protein (stained blue) (×100). (C) The small leaves present on the epicotyl are expanding during germination; because they were rather completely formed before the embryo became dormant, the leaves can now mature rapidly, and photosynthesis begins almost as soon as germination is complete. The hypocotyl is curved: This allows the leaves to be dragged gently up through the soil, protected by the cotyledons. (D) Most of a corn seed is the endosperm; the embryo is less than half the volume of the seed. During germination, the one cotyledon secretes digestive enzymes into the endosperm and absorbs the resulting monomers. (E) Corn seeds also produce many small leaves before becoming dormant, and they can begin photosynthesis immediately after germination. The leaves are protected from the soil by a tubular sheath called a coleoptile.
The amount of embryo growth and development that occurs before dormancy sets in is extremely variable. Orchids, bromeliads, and a few other species have small, dust-like seeds in which the embryo is only a small ball of cells with no cotyledons, radicle, or vascular tissue. In seeds of most angiosperm species, all of the parts are present, and the epicotyl contains two or three young leaves in addition to cotyledons; these leaves can begin photosynthesis immediately after germination (Figure 9-19). In corn, the embryo is even more advanced: It contains as many as six young leaves while in the seed. A fully mature corn plant often has only 10 or 12 leaves. Over half of the leaf production of the new sporophyte occurs while it is embedded in the parental gametophyte, which is itself embedded in the previous sporophyte.
The embryo and endosperm develop from the zygote and megagametophyte central cell, respectively, both located in the nucellus of the ovule. Soon after fertilization, or even before, synergids and antipodals break down in most species. The nucellus expands somewhat but later is crushed by the expansion of the embryo and endosperm and is usually not detectable in mature seeds.
The integuments that surround the nucellus expand and mature into the seed coat (also called the testa) as the rest of the ovule grows. In their last stages of maturation, they may become quite sclerenchymatous and tough (FIGURE 9-20). It is seed coat color that lets us distinguish between black, red, and pinto beans.
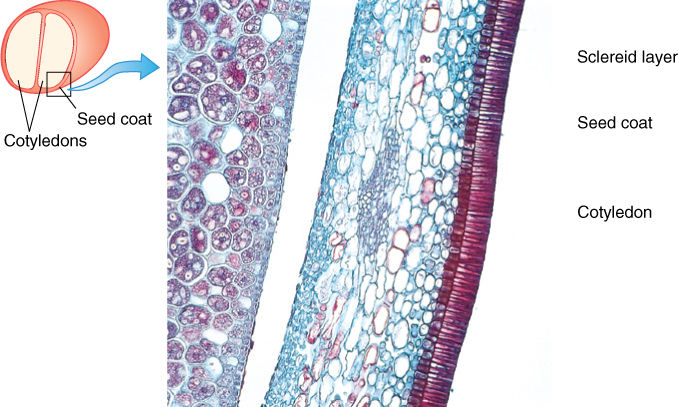
FIGURE 9-20 This seed coat of bean has an outer layer of sclereids and inner parenchymatous layers. The cotyledon is part of the embryo, and you might expect to see endosperm or nucellus between the cotyledon and seed coat; however, those have been crushed during the later stages of seed development (×200).
Fruit Development
As the ovule develops into a seed, the ovary matures into a fruit. Development varies with the nature of the carpels as well as the nature of the mature fruit. The stigma and style usually wither away, as do sepals, petals, and stamens, although they may persist at least temporarily (FIGURE 9-21). Often three layers become distinct during growth: The exocarp is the outer layer—the skin or peel; the middle layer is the mesocarp, or flesh; and the innermost layer, endocarp, may be tough like the stones or pit of a cherry or it may be thin (FIGURE 9-22). The relative thickness and fleshiness of these layers vary with fruit type, and often one or two layers are absent. The entire fruit wall, whether composed of one, two, or all three layers, is the pericarp.

FIGURE 9-21 In apples, the petals die and fall off after pollination. Stamens and sepals die and dry out, but they remain present, whereas carpels and receptacle develop into a fruit. In most species, sepals, petals, and stamens all abscise after pollination.

FIGURE 9-22 (A) A pecan nut is only the seed and innermost part (endocarp) of a fruit. The exocarp (green portion here) and mesocarp (brown) are tough, fibrous, and completely inedible. The fruit opens while on the tree, allowing the endocarp and seed to be shed. (B) Coconuts in stores are usually just the endocarp and seed; in nature, these are surrounded by a thick, fibrous, and buoyant mesocarp and a thin epicarp. A dark spot (an “eye”) on the endocarp is a soft spot where the seedling root and shoot will emerge during germination.
Before moving on to the next section, think about a peach tree in full bloom. The tree has thousands of flowers, each with stamens and carpels. When it has finished producing pollen and megaspores, it has technically finished its own reproduction and now has tens of thousands of offspring in the form of microgametophytes (pollen grains) and megagametophytes (inside each ovule in each carpel). Bees carry pollen from other peach trees to the stigmas of this one, and before long, it has thousands of pollen tubes growing inside its styles. You see one diploid peach tree, but inside it are thousands of haploid pollen tubes (which are also peach plants) and megagametophytes (also peach plants). All of the water, minerals, and photosynthates needed to keep all of these thousands of plants alive are being supplied by the leaves, roots, xylem, and phloem of the diploid peach tree. We cannot say that the haploid plants are parasitizing the tree because they are essential to its production of seeds. After fertilizations have occurred, hundreds of zygotes develop into embryos, each of which is the offspring of microgametophytes and megagametophytes and the “grand-offspring” of the peach tree itself. This tree has other “grand-offspring” as well, produced by its pollen that was carried to other peach trees. This tree is the ovule parent of its own seeds and the pollen parent of thousands of seeds in many other trees.
![]() Flower Structure and Cross-Pollination
Flower Structure and Cross-Pollination
The production and development of spores, gametes, zygotes, and seeds are complex, elaborate processes, but they are not the only functions of flowers. Flowers are also involved in the effective dispersal of pollen and seeds. Because numerous mechanisms carry out these processes, numerous types of flowers and fruits exist.
Cross-Pollination
Cross-pollination is the pollination of a carpel by pollen from a different individual; self-pollination is pollination of a carpel by pollen from the same flower or another flower on the same plant. In any plant population, there is genetic diversity. Random mutations will have produced some new versions of genes (each version is called an allele) that offer improved fitness and some that are deleterious. For example, people with blue eyes have the same genes for eye color as do people with brown eyes, it is just that they have different versions—alleles—of the eye color genes. With crosspollination, sperm cells and egg cells from different plants unite, resulting in new combinations of genes, at least a few of which may be better adapted than either parent. But self-pollination has about the same result as asexual reproduction because all genes (all alleles) come from the same parent. No possibility exists of bringing in new alleles that might provide more fitness than those inherited from the parent. However, if a plant is isolated by distance or lack of pollinators from potential cross-pollination partners, self-pollination allows it to set seed and propagate its genes rather than lose them when the plant dies.
Many mechanisms have evolved that decrease the probability of self-pollination and increase the chances of cross-pollination with its accompanying genetic diversity.
Stamen and Style Maturation Times
Self-fertilization in flowers that have both stamens and carpels is prevented if anthers and stigmas mature at different times (FIGURE 9-23). In many species, anthers release pollen while stigma tissues are immature and unreceptive; the style may not have elongated yet, and the stigmas may be near the base of the flower while the anthers are at the top, elevated by elongated filaments. Exposed pollen lives only briefly, being susceptible to desiccation in dry air and to damage to its DNA by ultraviolet light. When the stigma and style become mature, there may be no living pollen left in the flower; thus, all pollination is effected by younger flowers just opening their anthers.
This is not a very effective means of ensuring crosspollination. On plants with many flowers that do not open simultaneously, older flowers could be self-pollinated by freshly opened flowers of the same plant. Even if pollen does come from other plants, many of the near neighbors are probably closely related because many of the seeds of a plant fall and germinate near the plant, producing a cluster of plants that are at least as closely related as first cousins.
Stigma and Pollen Incompatibility
In many species, especially many important crop species, self-pollination is inhibited by compatibility barriers, chemical reactions between pollen and carpels that prevent pollen growth. In one incompatibility system, as a pollen tube grows, the stigma and style test proteins on the tube’s surface; if one of these proteins is produced by a gene that matches an incompatibility gene in the carpel-bearing plant, the stigma and style block any further growth of the pollen tube. In self-pollination, all pollen tube genes match those of the stigma and style, and blocking occurs: The plant cannot pollinate itself (FIGURE 9-24).

FIGURE 9-23 The stamens of this cactus flower are mature and shedding pollen, but the stigmas (green) are pressed together and are unreceptive. Later, the stamens wither and the stigmas spread open, ready to receive pollen from a different flower.
In another common system, the critical proteins are deposited on the outer surface of the developing pollen grain by the anther’s tapetum. Any match of proteins produced by the incompatibility genes blocks germination of the pollen grain. The interaction between a pollen tube and a style involves a haploid genome (pollen tube) and a diploid one (carpel), whereas the second system involves two diploid genomes (anther and carpel). The diploid/diploid system involves twice as many genes and therefore has a much greater probability of pollen rejection. With the diploid/diploid system, not only is self-pollination prevented, but inbreeding between close relatives is blocked as well.
Monoecious and Dioecious Species
Among incomplete flowers there is a significant difference between flowers that lack sepals or petals and those that lack stamens or carpels. The latter two organs are essential organs because they produce the critically important spores; if either organ is absent, sexual reproduction is dramatically affected. Flowers that lack either or both essential organs are not only incomplete but also imperfect flowers (see Figure 9-6). If a flower has both, it is a perfect flower even though it may lack either sepals or petals or both. Sepals and petals do not produce spores and are considered nonessential organs.
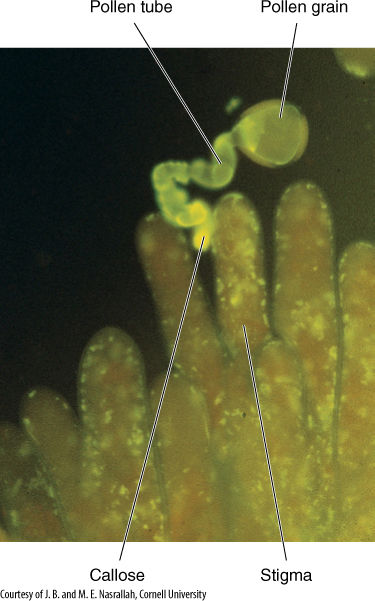
FIGURE 9-24 This pollen grain has started to grow on an incompatible stigma, but the stigma has blocked it, causing callose to form. The callose fluoresces in ultraviolet light, so it shines brightly in this micrograph (×200).
It is necessary to consider the entire plant and the entire species as well as individual flowers. Stamens produce pollen that results in sperm production, and carpels are involved in egg production; thus, a species must have both types of organs. Plants that have perfect flowers satisfy this requirement. But if some flowers of the species are imperfect, having no stamens, for instance, then other flowers must have stamens by being either perfect or by being imperfect because of a lack of carpels. A large number of combinations is possible: A species may have individuals that produce only staminate flowers and others that produce only carpellate flowers—this is dioecy, and the species (not the flower or the plant) is said to be dioecious (pronounced “dye EE cy” and “dye EE shus”). Examples of dioecious species are marijuana, dates, willows, and papaya. In dioecious species, the life cycle actually consists of four types of plants: (1) microgametophytes, (2) megagametophytes, (3) staminate sporophytes, and (4) carpellate sporophytes.
Monoecy is the condition of having staminate flowers located on the same plant as carpellate flowers; monoecious species include cattails and corn—ears are clusters of fertilized carpellate flowers, and tassels bear numerous staminate flowers (FIGURE 9-25). In some members of the cucumber family (including melons, squash, and pumpkins), the type of flower produced varies. Young plants and those growing in a poor environment produce staminate flowers, whereas older plants and those growing in a good environment produce perfect or carpellate flowers. If fertilization occurs, the carpels develop into large fruits, and only a healthy, robust plant can afford to do this. A young or poorly growing plant cannot supply enough carbohydrate and protein for fruit development, but it can supply enough to produce pollen.
Dioecy is an extreme adaptation that ensures cross-pollination; a plant that produces only one type of spore cannot pollinate itself. It is similar to the condition of separate genders in mammals—no individual can fertilize itself. Both conditions ensure that fertilization is by sex cells that are not identical genetically, thus increasing the genetic diversity of the offspring.
Animal-Pollinated Flowers
The evolution of animal-mediated pollination had a dramatic impact on the evolution of flowering plants. In wind-pollinated conifers, things such as brightly colored petals, fragrances, and nectar are a waste of material and energy. But once insects began visiting early angiosperms, mutations that resulted in pigments, fragrances, or sugar-rich secretions became adaptive. For pollen that is carried by wind, the probability that any particular grain will actually land on a stigma is very low, whereas if it is carried by an insect that flies from flower to flower, the probability of a pollen grain reaching a stigma is much improved.

FIGURE 9-25 (A) Ears of corn are really large groups (inflorescences) of carpellate flowers surrounded by protective leaflike bracts (the husks). The corn “silks” are long styles. (B) Corn tassels are inflorescences of staminate flowers. Each corn plant produces both ears and tassels: The species is monoecious.
When this insect-flower association began around 120 million years ago, neither insects nor flowers were particularly sophisticated. Insects probably visited all kinds of flowers, not recognizing the different species. As a result, pollen often landed on the stigma of the wrong species, where it was useless. Mutations in plants that increased a plant’s distinctiveness, its recognizability by an insect—flower color, size, shape, fragrances, and so on—became selectively advantageous. Rewards for the insect such as sugary nectar or protein-rich pollen and stamens were advantageous. Mutations in insects were adaptive if they increased the capacity of insects to recognize flowers that offered abundant nectar or pollen. Because it is expensive energetically for an insect to fly, the ability to recognize the most nutritious flowers from a distance while flying is advantageous. As a result, many lines of insects and flowers underwent coevolution, a flower becoming adapted for visitation by a particular insect and the insect for efficient exploitation of the flower. Coevolution has also occurred between flowers and birds and between flowers and bats.
The shape of the flower is particularly important, as a pollinator actually makes contact with it. Most flowers are radially symmetrical; that is, any longitudinal cut through the middle produces two halves that are mirror images of each other. These flowers (and stems and roots) are actinomorphic or regular (FIGURE 9-26). But all insects, birds, and bats are bilaterally symmetrical—only one longitudinal plane produces two halves that are mirror images. In many species, flowers and pollinators have coevolved in such a way that the flowers are now also bilaterally symmetrical—zygomorphic (Figure 9-26). When a pollinator approaches a zygomorphic flower, only one orientation is comfortable for it; any misalignment prevents the pollinator’s head or body from fitting the flower’s distinctive shape. As a result, as the pollinator feeds at the flower, pollen is placed on a predictable part of its body. When it visits the next flower, pollen is rubbed directly onto the stigma. Not only is pollen carried to the appropriate flower, but it is carried directly to the stigmas. In contrast, a pollinator can approach an actinomorphic flower from any direction, and pollen may be brushed onto any part of the body; when it visits the next flower, the pollen-carrying part may miss the stigmas and cross-pollination will not occur.

FIGURE 9-26 (A) Any median longitudinal section of this poppy results in two halves that are mirror images of each other; it is radially symmetrical, that is, actinomorphic. (B) These Penstemon flowers are bilaterally symmetrical, zygomorphic; only if cut from top to bottom are the two halves mirror images. A pollinator, also bilaterally symmetrical, fits well only if it approaches the flower properly. Anthers are at the top of each flower’s throat and will put pollen only onto a bee’s back.
Most plant/animal relationships are a battle: Animals try to eat plants, lay eggs in them, or do other harmful things. Plants defend themselves with poisons, spines, and sclereid barriers. Mutations that permit plants to make more effective deterrents benefit the plant but not its animal pests. But both plants and animals tend to benefit from the pollination relationship, and coevolution is possible. Mutations that make a plant more recognizable or more convenient to its pollinators help both the plant and the animal, just as certain mutations in pollinators are beneficial to both organisms. But neither side should give more than it has to. Some bees now are “nectar robbers,” being able to get nectar from flowers without having to carry any pollen. And a number of orchids, for example Chiloglottis, trick wasps into carrying pollen for free. These orchid flowers look, smell, and feel just like a female wasp, and male wasps try to mate with them. The wasps then fly off to the next orchid flower and the next, transferring pollen between each of them. The orchids get pollination without having to provide nectar, and the wasp gets nothing more than a good time.
Wind-Pollinated Flowers
In species that are wind pollinated, a totally distinct set of modifications is adaptive. Attracting pollinators is unnecessary; thus, mutations that prevent the formation of petals are selectively advantageous, and the energy saved can be used elsewhere in the plant. Sepals are also often reduced or absent, and the ovaries need no special protection; thus, the entire flower may be tiny. Zygomorphy provides no selective advantage. After pollen is released to the wind, the chance of any particular pollen grain landing on a compatible stigma is small, so huge numbers of grains must be produced. Large, feathery stigmas are adaptive by increasing the area that can catch pollen grains (FIGURE 9-27). In general, wind-pollinated individuals produce up to several thousand small flowers; although each flower is tiny, the entire plant has a large total stigmatic surface area.
Pollination is aided by the growth pattern of the plant population. Wind-pollinated species—like grasses, oaks, hickories, and all conifers—grow as dense populations in range lands or forests. Within 1 km2 may be found thousands of plants and, more importantly, millions of stigmas. Species that occur as widely scattered, rare individuals must rely on animal pollination.
Ovary Position
The ovary and ovules must be well protected from pollinators. Paradoxically, a flower must bring a hungry animal to within millimeters of protein-rich ovules in order to effect pollination. Adaptations that maximize the separation are long styles and stamen filaments. Burying the ovaries deep within the flower provides further protection. In some species, after all flower organ primordia are initiated at the receptacle apex (all parts are microscopically small when initiated), the primordia crowd together, and the bases of the stamens, petals, and sepals fuse, creating a thick layer of protective tissues around the ovaries, which appear to be located below the other organs (see Figures 9-21 and FIGURES 9-28A and 9-28B). Two terms describe this: We can either say that the ovary is an inferior ovary or that the other parts are epigynous. Inferior ovaries also can result if receptacle tissue grows upward around the ovary (FIGURE 9-28C). The more common arrangement, in which no fusion to the ovary occurs and the ovary is obviously above the other flower parts, is a superior ovary or hypogynous parts. Intermediate, partially buried ovaries are half-inferior with perigynous flower parts.

FIGURE 9-27 Many wind-pollinated trees have such tiny flowers people do not realize they are flowering plants at all. This is an entire set (an inflorescence) of flowers of poplar (Populus), each flower consisting of just numerous stamens.
![]() Inflorescences and Pollination
Inflorescences and Pollination
The positioning of flowers on an individual plant is important; few species have plants that produce only a single flower. Instead, many flowers are produced either within a single year or over a period of many years. A large mountain ash or cherry tree produces thousands of flowers every year for well over 100 years. Many important factors determine whether a flower is seen and visited by pollinators—for example, the positions of flowers relative to other flowers, leaves, and trunk; height from the ground; and distance from an open, uncluttered flight path for pollinators (FIGURE 9-29).
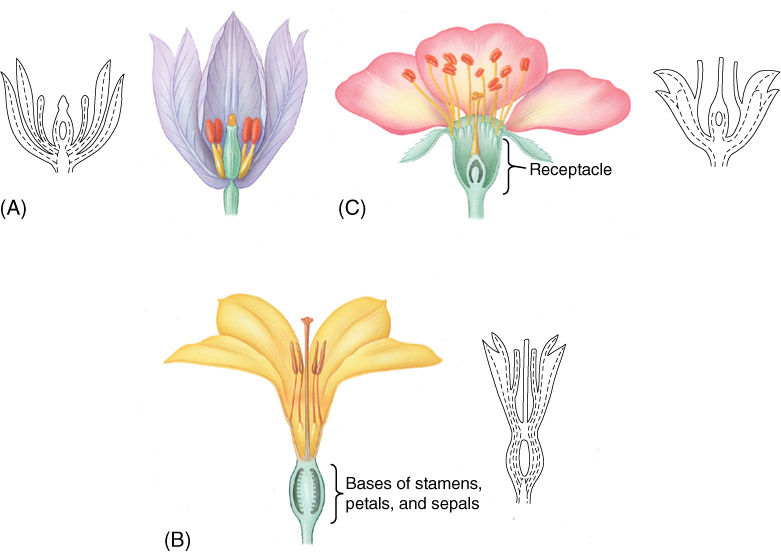
FIGURE 9-28 (A) In flowers with superior ovaries, carpels sit above the other organs, as would be expected because they are initiated last by the flower apical meristem. (B) This is an inferior ovary resulting from the fusion of the bases of sepals, petals, and stamens to the base of the carpel. Four appendages are involved whose vascular bundles (dashed lines) are still present and distinct. (C) In some species, after all of the appendage primordia are initiated, the receptacle tissue grows upward and surrounds the ovaries. This can be detected because the vascular bundles around the ovary are receptacle bundles, not appendage bundles.
Reproductive success is measured in terms of the number of healthy, viable seedlings that become established. One important factor for this is pollination. It might seem optimal to have large flowers that can be easily seen by pollinators; unfortunately, they can also be seen by herbivores, and because ovules and pollen are rich in protein, they are good food sources for pests. Production of the pedicel, receptacle, sepals, and petals can be thought of as packaging and advertising cost; the larger the flower is, the larger the cost. These costs can be made more acceptable (less disadvantageous selectively) by increasing the number of ovules (potential seeds) per flower. Large flowers tend to have numerous ovules and small flowers fewer. Small flowers exist because small plants cannot afford large flowers and their accompanying large seeds, which are very expensive. Also, a large flower with many ovules is a big risk—if it is found and eaten, a major reproductive investment has been lost; however, damage to a small flower with only a few ovules is less significant. Also, if many flowers are grouped together, an inflorescence, they give a collective visual signal to pollinators: One small flower may be overlooked, but not a hundred close together (FIGURE 9-30). Furthermore, in an inflorescence, the plant is able to control accurately the timing of the initiation, maturation, and opening of the flowers. Consequently, the plant can be in bloom and available to pollinators for several weeks even though each flower lasts only a day or two.

FIGURE 9-29 (A) Flowers of sausage tree (Kigelia) are pollinated by bats, which do not like to fly among the clutter of leaves because their sonar does not work well there. Long stalks allow flowers to hang in open air where bats have free access to them. (B) Angiosperms that grow under water in streams and lakes usually have flowers that project above the water’s surface, and are accessible to flying pollinators (Ranunculus aquatilis).

FIGURE 9-30 (A) Palm flowers are small and inconspicuous, but when grouped together, their inflorescence has even more impact than a single large flower. Thousands of tiny palm flowers are present here, each too small to be seen at this magnification. (B) The inflorescence of Combretum is even more striking than the individual flowers. Notice how the flowers mature and open sequentially.
Large flowers can produce more nectar than small flowers, but this makes them a more tempting target for nectar robbers, animals that take nectar without carrying pollen. Furthermore, a flower should not produce enough nectar to completely satisfy even a legitimate pollinator because no incentive would remain for the pollinators to go to another flower; it is better if the pollinators get only enough to make them interested in searching for more.
Many other factors affect the relative selective advantage or disadvantage of flower number and size as well as the number of ovules per flower. Considering the numerous types of plants, pollinators, and environments, the diversity of flower and inflorescence types is not surprising. In the simplest arrangement, flowers occur individually in leaf axils or as a transformation of the shoot apex. When grouped into inflorescences, two basic arrangements occur: (1) determinate inflorescences and (2) indeterminate inflorescences.
A determinate inflorescence has only a limited potential for growth because the inflorescence apex is converted to a flower, ending its possibilities for continued growth (FIGURES 9-31A and 9-31B). Typically, but not always, the terminal flower opens first, and then lower ones open successively. In the simplest type, below the terminal flower is a bract with an axillary flower, which also may have a bract and axillary flower, and so on.
In an indeterminate inflorescence, the lowest or outermost flowers open first, and even while these flowers are open, new flowers are still being initiated at the apex. A raceme has a major inflorescence axis, and the flowers are borne on pedicels that are all approximately the same length (FIGURE 9-31C). Catkins are similar to racemes, differing in that the flowers are imperfect, either staminate (Figure 9-27) or carpellate, and all flowers of a single inflorescence are the same, and thus, each species must have both staminate catkins and carpellate ones. Catkins almost always contain very small, wind-pollinated flowers. A spike is similar to a raceme except that the flowers are sessile, lacking a pedicel (FIGURE 9-31D). A spadix (plural, spadices) is a spike-like inflorescence with imperfect flowers, but both types occur in the same inflorescence, most often with staminate flowers located in the upper portion of the inflorescence and carpellate flowers in the lower portion, although they can intermingle. The main inflorescence axis is thick and fleshy with minute flowers embedded in it; the entire inflorescence is subtended or enclosed by a petal-like bract called a spathe. A panicle is a branched raceme with several flowers per branch.
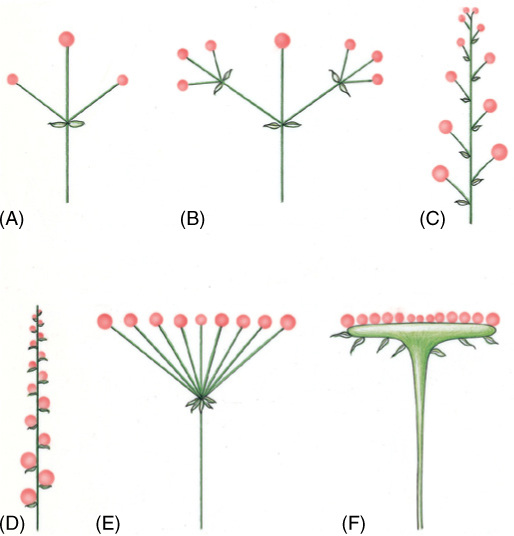
FIGURE 9-31 Inflorescence types: (A) Simple determinate inflorescence. (B) Compound determinate inflorescence. (C) Raceme. (D) Spike. (E) Umbel. (F) Head. Larger circles represent flowers that open earlier than those depicted as smaller circles.
Several types of indeterminate inflorescences have no dominant main axis. In umbels, the inflorescence stalk ends in a small rounded portion from which arise numerous flowers (FIGURE 9-31E). Their pedicels are long and arranged so that all flowers sit at the same height, forming a flat disk. A head is similar to an umbel except that the flowers are sessile and attached to a broad expansion of the inflorescence stalk (FIGURE 9-31F); numerous bracts may surround the inflorescence during development. Heads are almost synonymous with the aster family, sunflowers and dandelions being easily recognizable examples. In this group, the inflorescences are so compact and highly organized that they mimic single flowers; what appear to be the petals are really entire flowers, ray flowers, in which the petals are very large and fused together. The center of the inflorescence is composed of a different type of flower, disk flowers, in which the corollas are short and inconspicuous.
![]() Fruit Types and Seed Dispersal
Fruit Types and Seed Dispersal
Fruits are adaptations that result in the protection and distribution of seeds. Many different agents disperse fruits and the seeds they contain: Gravity, wind, water, and animals are the most common (TABLE 9-4). The principles involved in fruit function are somewhat opposed to each other: Fruits that are tough and full of fibers or sclereids, such as pecans, walnuts, brazil nuts, and coconuts, offer maximum protection but are heavy and expensive metabolically. Also, the protected seed must be able to break out to make germination possible; a more fragile fruit is better for that. If animals are to disperse the seeds, part of the fruit must be edible or otherwise attractive, whereas the seed and embryo must be protected from consumption. A division of labor often occurs—some parts being protective, others attractive, and still others allowing germination.
TABLE 9-4 Agents of Dispersal
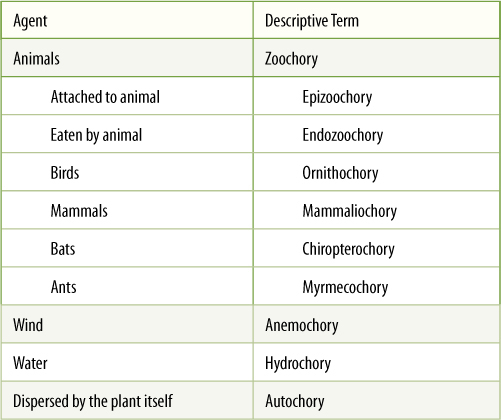
Seeds, fruits, and asexual propagules can be dispersed by many means. These are a few of the most common types.
True Fruits and Accessory Fruits
The term “pericarp” refers to the tissues of the fruit regardless of their origin. In most cases, this is the ovary wall, but in many species, especially those with inferior ovaries, the receptacle tissues or sepal, petal, and stamen tissues may also become involved in the fruit. The terms “pericarp” and “fruit” have been applied to both types of fruit; thus, now the term true fruit is used to refer to fruits containing only ovarian tissue, and accessory fruit (or false fruit) is used if any nonovarian tissue is present (FIGURE 9-32). Apples develop from inferior ovaries, and the bulk of the fruit is enlarged bases of sepals and petals; only the innermost part is true fruit derived from carpels (see Figure 9-21).

FIGURE 9-32 (A) The red, edible flesh of strawberry is really the receptacle, not carpel tissue. It is therefore an accessory fruit. The “seeds” are true fruits, each derived from one carpel of the flower. (B) Pineapples develop from the coalescence of all the many true fruits of one inflorescence, so they are a multiple fruit. In addition, many noncarpellary tissues become involved, and are therefore accessory fruits as well. Bracts are visible here.
Fusion of carpels also affects the nature of fruits. If the fruit develops from a single ovary or the fused ovaries of one flower, it is a simple fruit, the most common kind. If the separate carpels of one gynoecium fuse during development, an aggregate fruit results, such as raspberries. If during development all of the individual fruits of an inflorescence fuse into one fruit, it is a multiple fruit, as in figs, mulberries, and pineapple (Figure 9-32B). These are also largely accessory fruits because in addition to the ovary tissue, the inflorescence axis, bracts, and various flower parts contribute to the mature fruit.
Classification of Fruit Types
There are several ways of grouping or classifying fruits. In one method, emphasis is placed on whether the fruit is dry or fleshy. A dry fruit is one that is not typically eaten by the natural seed-distributing animals; fleshy fruits are those that are eaten during the natural seed distribution process.
A further classification of dry fruits emphasizes fruit opening: Dehiscent fruits break open and release the seeds, whereas indehiscent fruits do not (TABLE 9-5). For the most part, fleshy fruits are indehiscent. Animals chew or digest the fruit, opening it; if uneaten, the fruit rots and liberates the seeds. Although many dry fruits are dehiscent, others must rot, or the seed must break the fruit open itself.
Perhaps the simplest fruits are those of grasses (all cereals such as corn and wheat): Each fruit develops from one carpel containing a single ovule. During maturation, the seed fills the fruit and fuses with the fruit wall, which remains thin: “Seeds” of corn and wheat are actually both the fruit and its single seed, fused into a single structure. It has little protection and no attraction for animals. The fruit/seed falls and germinates close to the parent sporophyte. The fruits are indehiscent, and during germination, the embryo absorbs water, swells, and bursts the weak fruit. These fruits are caryopses.
TABLE 9-5 Fruit Types
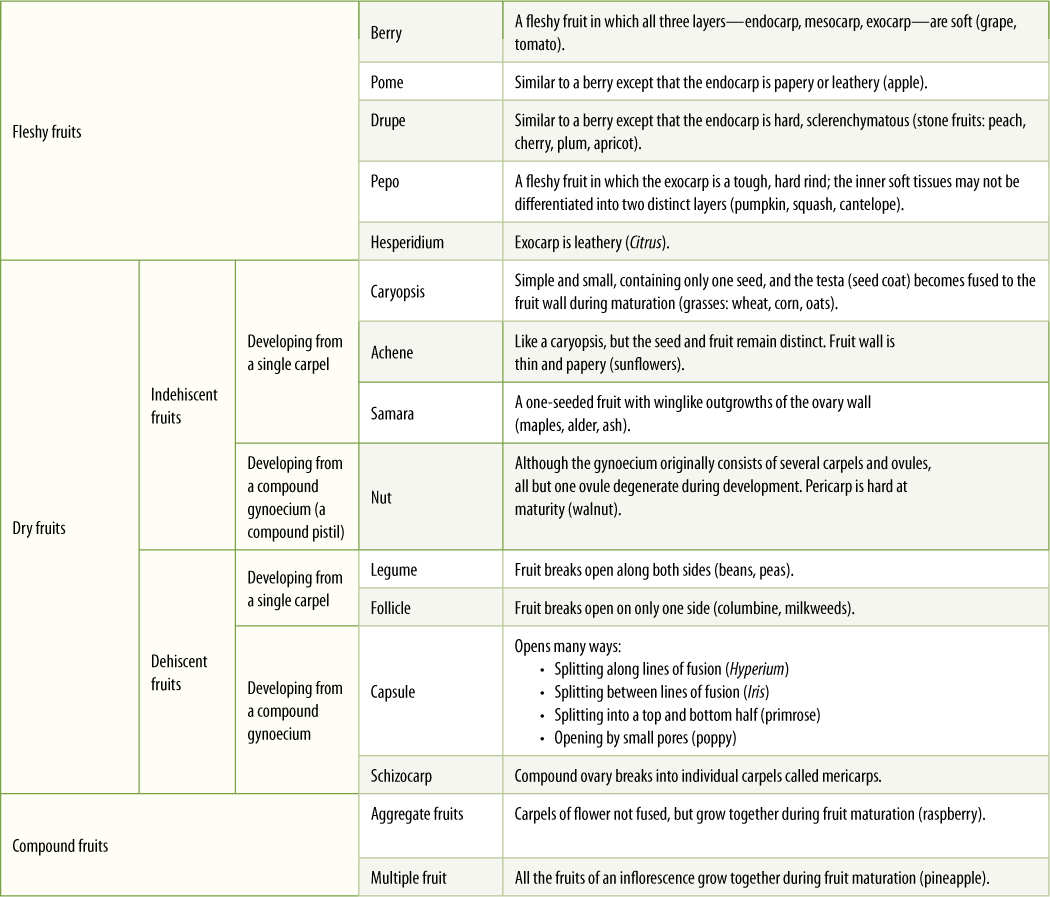
Beans and peas form from a single carpel that contains several seeds. The fruits, known as legumes, are dry and inedible (unless eaten while still young and green, such as snowpeas and green beans) like caryopses but do not fuse to the seeds. At maturity, the two halves of the fruit twist and break open (dehisce) along the two specialized lines of weakness (FIGURE 9-33). After liberation from the legume, the seeds are protected from small animals and fungi by only the seed coat.
Fruits or seeds carried by wind must be light; they often have wings (maples) or parachutes (dandelions) that keep them aloft as long as possible. Such fruits are dry and weigh much less than one gram. Fruits and seeds that are transported by water (ocean currents, streams, and floods) can be larger and heavier, but they must be buoyant and resist mildew and rot. An excellent example is coconuts that float from one island to another.
Animals carry fruit in a variety of ways. Dry fruits with hooks or stickers catch onto animal fur or feathers; sticky, tacky fruits glue themselves onto animals, and fleshy, sweet, colorful fruits are eaten. Edible fruits require particular specialization because the fruit must be consumed but the seeds must not be damaged; they must not be crushed by teeth or gizzard or be digested. Immature fruits deter frugivores by being hard, bitter, or sour, such as unripe apples or bananas. At maturity, the fruit is soft, sweet, flavorful, and typically strikingly different in color. The enclosed seeds often have hard seed coats or endocarps (peach and cherry) that allow them to pass through an animal unharmed (see Figure 9-22). Dinosaurs may have been especially poor seed dispersers: They had no hair for hooks to tangle in, and their digestive tracts were often too long, hot, and fermentative for seeds to survive.
Grapes and tomatoes are examples of simple fleshy fruits: They are berries because all parts of the fruit are soft and edible (Table 9-5). The seed coat provides some protection against being crushed during chewing, but perhaps more important are the size of the seeds and their slippery seed coat, which allows the seeds to slip out from between our molars. Pomes (apples and pears) differ from berries in two ways. First, they develop from inferior ovaries, and thus, the inner tissue is the true fruit and the outer tissue is accessory fruit. Second, the seeds are protected because the innermost fruit tissues, the cores, are bitter and tough. The most elaborate fleshy fruits are drupes (peaches and cherries), in which the innermost tissues, the endocarp, are extremely hard and totally inedible, full of sclerenchyma. The mesocarp becomes thick, fleshy, juicy, and sweet, and the exocarp forms the peel, whose color informs the frugivore of the fruit’s ripeness. The endocarp of a drupe is known as the pit and is often mistaken for the seed, but the true seed and its seed coat are located inside. Drupes provide maximum attraction to animals with minimum danger to the seed.
Because frugivores are a major danger to seeds, it is necessary to think about how edible fruits evolved: They must offer some advantage that outweighs the danger. Animals have predictable habits and migration patterns. Ants, birds, and mammals carefully select the sites where they feed, nest, and sleep; therefore, seeds are not distributed at random but are moved through the environment to specific sites. One example is mistletoe: Its seeds are sticky and adhere to a bird’s beak as it eats the fruit. The bird probably feeds and cleans itself in the same species of tree; thus, the seeds are rubbed off the beak directly onto a proper host tree. Few seeds are ever lost by dropping to the ground or being deposited in the wrong tree, as would happen with wind or water dispersal. Just as efficient is dispersal of fruits and seeds of marsh plants. The birds and animals that live in marshes typically spend little or no time in other habitats, so seeds carried by these animals are almost certainly distributed to favorable sites.

FIGURE 9-33 (A) At maturity, different layers in this dry fruit contract in opposite directions, causing the pod to twist and break open. (B) As these maple fruits drop, the wings cause them to rotate and float, thus remaining aloft and traveling much farther on the wind. (C) The spiral, hairy tails of these geranium fruits let them float on the wind over long distances.
A special benefit of seed distribution by animals is that the “deposited” seed may find itself in a small (or large) mound of “organic fertilizer” (FIGURE 9-34). Seeds adapted to pass through an animal’s digestive tract are resistant to digestive enzymes and can tolerate the high level of ammonium in fecal matter. Because many seeds of plants cannot tolerate these two factors, the adapted seeds find themselves in a microenvironment that is not only nutrient rich but also excludes some competitors.

FIGURE 9-34 This seedling is as happy as a clam at high tide: It has enough fertilizer to last for months. Many of its competitors have been digested; others are being suffocated.
 At the Next Level
At the Next Level
1. Pollination syndromes. Each type of pollination agent affects the evolution of the flowers it pollinates. Wind-pollinated flowers usually evolve to lack sepals and petals: Hummingbirds prefer flowers that are long, tubular, and red; bats fly at night and bat-pollinated flowers evolved to open after sunset, to have a strong fragrance, and even to reflect the bat’s sonar back to the bat such that the flower is found easily in the dark. Dozens of other pollination syndromes occur and are remarkable examples of co-evolution between plants and animals. Be sure to search for pseudo-copulation between wasps and orchids.
2. Frugivores and seed dispersal. Co-evolution has occurred between plants and animals such that seeds are dispersed to preferred sites rather than at random. For example, seeds of epiphytic plants and parasitic plants should be carried to new branches of suitable host trees, usually by having birds eat the fruits without damaging the seeds. As with pollination, many syndromes have evolved, some involving fruit as a reward for dispersal, others involving food bodies on the seeds themselves (such as elaiosomes, arils, and caruncles). Of special interest, several plant species produce edible fruits but we have not found any animals that eat them. One hypothesis is that the animals went extinct recently but the plants persist even though their seeds are no longer being dispersed effectively (many animals in the Americas became extinct just after human hunters reached these continents). Be sure to search on “extinct seed dispersers.”
3. Evolution of flowers. In most flowers, both stamens and carpels occur together within the same flower. Perfect flowers are common and evidence indicates that the earliest flowers were perfect. But in all other seed plants, pollen cones and seed cones are always separate: Pollen-producing structures never occur together with ovule-producing structures. Thus, we are still far from being certain of the steps involved in the evolution of early flowers, the early angiosperms, and their ancestors, but this is a fascinating area. There are conflicting theories and conflicting evidence. This is a challenging topic for anyone, especially for beginning botany students, but it is very rewarding.
4. Recommended books. A team of botanists and a visual artist created three wonderful books about plant reproduction. These are large coffee-table books with large illustrations, each image being a photograph or a scanning electron micrograph (an SEM image) of pollen or fruits or seeds. SEM images are naturally black and white, but the team colorized the images to enhance certain features so that the various parts are more readily understandable. Each is accompanied by just enough text to fully explain the biology without overwhelming the reader. I recommend them highly.
Pollen: The Hidden Sexuality of Flowers by Rob Kesseler and Madeline Harley (2006, Firefly Books).
Fruit: Edible, Inedible, Incredible by Wolfgang Stuppy and Rob Kesseler (2008, Firefly Books).
Seeds: Time Capsules of Life by Rob Kesseler and Wolfgang Stuppy (2012, Earth Aware Editions).
SUMMARY
1. Asexual reproduction results in new individuals genetically identical to the parent. The parent and all progeny are equally adapted to the habitat.
2. Sexual reproduction results in progeny that differ from each other genetically. There is a range of fitness, and some progeny may have a combination of maternal and paternal genes that causes them to be even better adapted than the parents.
3. All seed plants have a life cycle that consists of an alternation of heteromorphic generations. Microgametes and megagametes are produced by microgametophytes and megagametophytes that grow from microspores and megaspores. Most species have only one type of sporophyte, capable of producing both types of spores. Dioecious species have two types of sporophytes, one of which produces microspores and the other megaspores.
4. Flowers contain all or some of the following: nonessential organs—pedicel, receptacle, sepals, and petals; and essential organs—stamens and carpels.
5. In flowering plants, the microgametophyte consists initially of a vegetative cell and a generative cell, but the latter divides into two sperm cells. The megagametophyte consists of the egg cell, two synergid cells, a binucleate central cell, and three antipodal cells.
6. After fertilization, the zygote (fertilized egg) grows into an embryo; polar nuclei fuse with the second sperm nucleus, establishing the endosperm nucleus and endosperm, and the integuments develop into a seed coat. The entire structure is a seed. The ovary develops into a fruit.
7. Inflorescences are groups of many flowers. They may be determinate or indeterminate and may contain several types of specialized flowers.
8. Fruits protect seeds during development and may aid in their dispersal and maturation. Fruits are either dry, inedible, dehiscent, or indehiscent, or they are fleshy, edible, and indehiscent.
IMPORTANT TERMS
alternation of generations
anther
carpels
coevolution
compatibility barriers
complete flowers
cotyledons
cross-pollination
double fertilization
embryo sac
endosperm
epicotyl
fruit
gametes
gametophyte
heteromorphic generations
hypocotyl
imperfect flowers
incomplete flowers
inflorescence
karyogamy
ovary
ovules
pedicel
perfect flower
petals
plasmogamy
pollen
pollen tube
radicle
receptacle
seed coat
seeds
sepals
spores
sporophytes
stamens
stigma
style
syngamy
zygote
REVIEW QUESTIONS
1. Reproduction can serve two very different functions. What are they?
2. Under what conditions is it selectively advantageous for a plant to produce offspring identical to itself?
3. Imagine a plant that has been well adapted to a particular habitat, and it has been reproducing only asexually; thus, all of its offspring are identical to it. If the climate or other conditions change such that the plant can no longer survive in the new conditions, what will happen to all of its offspring? Why?
4. With asexual reproduction, are progeny ever more fit, more adapted than the parent?
5. Look at the apples in Figure 9-1a, all growing on a single tree. Do all of the seeds in all of the apples in the photograph have the same maternal parent? Do they all have the same paternal parent?
6. Sexual reproduction produces offspring that are not identical to each other or to either parent. Usually, some are more well adapted than the parents. Some are more poorly adapted, and most are about as adapted. What is one of the beneficial aspects of this diversity?
7. In stable populations, ones that are neither increasing nor decreasing in abundance (e.g., there are a million trees now and there will be a million trees a thousand years from now), about how many of a plant’s seeds survive and grow to adulthood, being able to replace it when it dies? If during the plant’s lifetime it produces 100,000 seeds, how many do not survive, do not grow, and cannot replace it when it dies. (Hint: Do not think of humans, as we are an increasing population, not a stable one. Almost all our children survive, but that is not true of any other species.)
8. Describe the life cycle of us humans. Are the tissues and organs in our bodies made up of diploid cells or haploid ones? Our reproductive organs make sperm cells in males and egg cells in females. Is this done by mitosis or meiosis? When our bodies make gametes, does every cell in our body become a sperm cell or an egg cell, or do just some of the cells in our reproductive organs do this? Can our haploid sperm and egg cells undergo mitosis and grow into new animals that look like us but are haploid instead? (Haploid eggs do grow in bees. They develop into males.) After a fertilized egg (a zygote) is formed, is it diploid or haploid? Can it immediately undergo meiosis to make four new sperm cells or egg cells, or can it only grow by mitosis into another person?
9. The plants we are familiar with are called sporophytes or the sporophyte generation. Do these plants have bodies made up of diploid cells or haploid ones? In which organs does meiosis occur (two are correct)—leaves, stems, stamens, roots, carpels, or petals? When plants undergo meiosis, do all cells of the plant undergo meiosis? Do all cells become haploid, or does meiosis occur in just a few cells in some flower parts?
10. In animals, meiosis produces gametes (sperm cells and egg cells), but that does not happen in plants. When some of the cells of a sporophyte undergo meiosis, what types of cells are produced? What do they grow into?
11. What is a gametophyte? How many different types of gametophytes are there in a plant life cycle? What do they look like (Hint: See Figure 9-5b)? Do the sporophytes and gametophytes of seed plants ever look like each other?
12. Draw and label a microgametophyte of a flowering plant. What type of gamete does it produce? Draw and label a megagametophyte of a flowering plant. What type of gamete does it produce?
13. Flowers typically have many parts, although some flowers can be missing some of the standard parts. What is the name of each of the following parts:
a. The stalk of the flower
b. The end of the stalk, where the other parts are attached
c. The parts that are usually green and protect the flower bud as it develops
d. The parts that attract pollinators
e. The parts that produce pollen
f. The parts that receive pollen and which contain ovules
14. If a flower has all of the parts c, d, e, and f in Question 13, they are said to be _______________ flowers. If they are missing any of these parts, they are said to be ________________________ flowers. Parts e and f are especially important because they produce the reproductive cells, the spores. If a flower has both e and f in Question 13, they are _____________________ flowers. If they are missing either e or f or both, they are _______________ flowers.
15. In flowers that are pollinated by wind or water, which of the parts in Question 13 is often missing?
16. What is the difference between a perfect and an imperfect flower? What is the difference between a complete and an incomplete flower?
17. A stamen usually has two parts. The stalk is called a ____________________ and an upper portion, the ______________________, which produces the pollen. Only some of the cells in the upper part undergo meiosis and become pollen grains (microspores). Those cells are called ____________________ mother cells or _________________________. Neighboring anther cells, in a layer called the ____________________, act as nurse cells.
18. Carpels usually have three parts: a _________________ that catches pollen grains, a _______________ that elevates the first part, and an _______________ where megaspores are produced. In this last part, there are placentae that bear small structures called _______________ each with a short stalk called a funiculus and a central mass of parenchyma called the nucellus. One cell in the nucellus will be the _______________ mother cell or ___________________.
19. The megaspore in most flowering plants grows into a megagametophyte that has seven cells and eight nuclei. Name and describe the seven cells.
20. After pollen lands on a stigma, it is far away from the ovule with the megagametophyte, which holds the egg (the megagamete). How are the two sperm cells transported from the stigma to the egg?
21. In angiosperms, as a sperm cell enters the egg, it loses both __________________ and _____________________ such that only the sperm cell nucleus contributes any DNA to the new zygote. In gymnosperms, however, the sperm cell loses only its _______________________ such that the zygote inherits both plastids and a nucleus from the sperm cell.
22. After the sperm cells enter a synergid, one fertilizes the egg in a two-step process. First there is fusion of the sperm cell’s protoplasm with that of the egg, a step called ______________________________. Then the sperm cell nucleus fuses with the egg cell nucleus, the second step, called _____________________________.
23. What happens to the second sperm nucleus, the one that does not fertilize the egg cell? What is the tissue that develops from this second “fertilization”? How is coconut related to this?
24. In most eudicot seeds, the parts of the embryo are very easy to see. Describe each of these parts:
a. Cotyledons
b. Radicle
c. Hypocotyl
d. Epicotyl
e. Extra credit portion: Answer the following question if you have read about the radicle with regard to a taproot system and a fibrous root system: How is the radicle involved in these different types of root systems?
25. What are albuminous and exalbuminous seeds? Consider corn, peas, and beans. Which of these seeds are which?
26. After pollination and fertilization, as the ovule develops into a seed, the ovary matures into a _________________. Many of these have three parts. The _________________ is the skin or peel. The ________________________ is the flesh, and the innermost layer, the ____________________, may be tough like the pit of a cherry.
27. What is the difference between self-pollination and cross-pollination? If pollen is transferred from the stamens of a flower to the stigma of the very same flower, is that cross-pollination or self-pollination? If it is transferred from the stamen of one flower to the stigma of another flower on the very same plant? If it is transferred from the stamen of one flower to the stigma of another flower on a different plant, but a plant that is a clone of the first one? Animals do not have pollen, of course, but why is it that most animals never have to worry about the equivalent problem of self-fertilization? Why cannot most animals fertilize themselves?
28. Describe a species that is dioecious. Name several examples (Hint: See Figure 9-6). In order for sexual reproduction to occur in a dioecious species, how many separate plants must be involved? Name them all.
29. Describe a monoecious species. Name several examples.
30. In several pairs of plant and animal species, plants have become modified such that only its animal partner can pollinate it, and the animals have become modified such that they are especially well adapted to pollinate just their plant partner. What is the name of this type of evolution that results in two organisms becoming particularly adapted to and dependent on each other?
31. Explain the following terms: inferior ovary, superior ovary, actinomorphic flower, zygomorphic flower. How is each of these modifications selectively advantageous?
32. What is an inflorescence? The inflorescences of a sausage tree (Figure 9-29) hang far down out of the tree. How is this of benefit to bats? Why can the inflorescence of Combretum (Figure 9-30B) attract more pollinators than can the individual flowers?
33. After pollination and then fertilization, what usually happens to each of the following: stigma, style, carpel, ovule, integuments, and zygote?
34. Fruits are often classified as dry or fleshy. What is the difference? Which of these two are dehiscent, which are indehiscent?
35. In ordinary English, we use the word fruit to mean something sweet and juicy; however, the following things are fruits: peanut shells, pea pods, bell peppers, and chili peppers. What is the characteristic that lets us know these really are fruits even though they are not sweet? In contrast, bananas are fruits that do not have this characteristic (they are sterile and new plants must be grown from buds that sprout near the base of the plant).
36. Some things that we call fruits are not true fruits but instead are accessory (false) fruits. What is the red part of a strawberry, and what are the true strawberry fruits (Hint: See Figure 9-32A)? In an apple, what is the fleshy part that we eat, and what is the core that we throw away?
Design Credits: Hummingbird: © Tongho58/Moment/Getty; Green plant cells: © ShutterStock, Inc./Nataliya Hora; Purple tulip: © ShutterStock, Inc./Marie C Fields; Dandelion: © ShutterStock, Inc./danielkreissl; Poppy: © ShutterStock, Inc./Saruri; Plant icon: © ShutterStock, Inc./Vector; Digging man icon: © ShutterStock, Inc./Z-art