Botany: An Introduction to Plant Biology - Mauseth, James D. 2017
Plant Physiology and Development
Energy Metabolism: Photosynthesis

Chapter Opener Image: The high-energy electrons moving through these power lines are dangerous and could easily kill a person. Photosynthesis also involves the movement of high-energy electrons, which, if not controlled properly, can and do damage chloroplasts and even the whole plant. There was no electricity flowing through these lines when they were being constructed, but a plant’s photosynthetic pigments become energized as soon as they are synthesized, and they are even dangerous in autumn while chloroplasts are being dismantled. Whenever light strikes chlorophyll, an essential but risky process begins.
OUTLINE
✵ Concepts
✵ Energy and Reducing Power
- Energy Carriers
- Reducing Power
- Other Electron Carriers
✵ Photosynthesis
- The Light-Dependent Reactions
- The Stroma Reactions
- Anabolic Metabolism
✵ Environmental and Internal Factors
- Light
- Leaf Structure
- Water
- C4 Metabolism
- Crassulacean Acid Metabolism
Box 10-1 Plants and People: Photosynthesis, Global Warming, and Global Climate Change
Box 10-2 Alternatives: Photosynthesis in Bacteria and Cyanobacteria
LEARNING OBJECTIVES
After reading this chapter, students will be able to:
✵ Recall the method of energy transportation from energized pigments into endergonic reactions.
✵ Explain why plants need reducing power.
✵ List four important qualities of carbon dioxide and water as photosynthesis compounds.
✵ Describe the (theoretically) ideal photosynthetic pigment.
✵ Compare chlorophyll a to this ideal.
✵ Summarize the function of accessory pigments.
✵ Describe the ways in which the light-dependent reactions and the stroma reactions function together.
✵ Explain the ways in which photosynthesis affects Earth’s climate.
 Did You Know?
Did You Know?
✵ The photosynthetic pigment chlorophyll absorbs mostly red and blue but is poor at absorbing green light. Green light bounces off and thus leaves appear to be green.
✵ Almost all energy used by animals, fungi, and everything else is the energy of sunlight captured by photosynthesis in plants, algae, and cyanobacteria.
✵ Light intensity must be just right; in habitats where sunlight is too intense (deserts, mountaintops), plants often have hairs or spines that shade the plant; if light is too dim (in a shady forest or deep in the ocean) not enough energy is available to allow life.
![]() Concepts
Concepts
Probably the most important concept concerning cells and all of life itself is that living organisms are highly ordered, highly structured systems. The universe as a whole is constantly becoming less orderly; its disorder (entropy) is increasing. Prokaryotes, protists, fungi, plants, and animals, however, represent phenomena in which particles become more orderly. A plant absorbs diffusely scattered molecules of carbon dioxide, water, and minerals and organizes them into organic molecules, cells, tissues, and organs. Each plant carries this out with such precision that each species of plant is easily distinguishable from others. After death, decay is the process by which an organism’s molecules become more disordered and scattered—their entropy increases.
Because living organisms are part of the natural world described by the laws of physics and chemistry, the decrease in the entropy of living organisms must obey physical laws. This is accomplished by putting energy into the living system, the source of energy being sunlight. To be accurate, we must consider the sun and life together: Atomic reactions that generate sunlight cause greater disorder in the sun than sunlight causes order in living organisms. The entire system (sun + life) becomes more disordered. Because there is no means of putting energy into an organism’s body after death, an increase in entropy cannot be prevented.
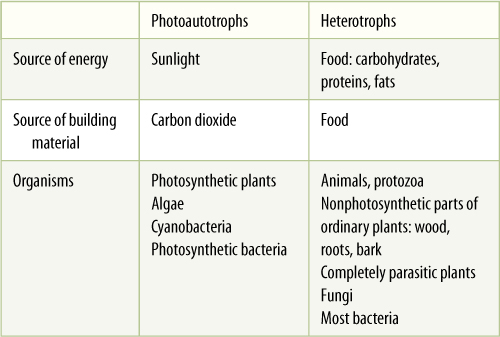
Sunlight maintains and increases the orderliness of life by two methods: (1) directly, in the process of photosynthesis, which produces complex organic compounds, and (2) indirectly, in the respiration of those organic compounds, either by the organism itself or by another organism that eats it. These two methods of supplying energy and maintaining orderliness—photosynthesis and respiration—are the basis for a major, fundamental distinction in the types of organisms. Photoautotrophs are organisms that gather energy directly from light and use it to assimilate small inorganic molecules into their own tissues. Photoautotrophs include all green plants, all cyanobacteria, and the few bacteria capable of photosynthesis. Heterotrophs are organisms that cannot do this but instead take in organic molecules and respire them, obtaining the energy available in them. Heterotrophs include all animals, all completely parasitic plants (FIGURE 10-1), all fungi, and nonphotosynthetic prokaryotes. Gathering energy by taking in organic material has the advantage that part of the material can be used as construction material instead of fuel. At least some of the amino acids, fatty acids, and sugars in food can be built into the organism’s own polymers and the rest respired for energy. Photoautotrophs must build all of their own molecules using just carbon dioxide, water, and various nitrates, sulfates, and other minerals.
Tremendously important consequences follow from the fact that photoautotrophs and heterotrophs differ in their sources of energy and building material (TABLE 10-1). Sunlight and carbon dioxide do not need to be stalked, hunted, and captured; thus, sensory organs, muscles, and central nervous systems like those of animals are unnecessary. Conversely, the ocean is full of microscopic bits of food, and animals such as sponges and corals can gather it the way plants gather carbon dioxide. The mode of nutrition has had overriding influence on the bodies and metabolisms of plants and animals.
Tissues and organs are also either photoautotrophic or heterotrophic. Chlorophyllous leaves and stems are photoautotrophic, whereas roots, wood, and flowers are heterotrophic and survive on carbohydrates imported through phloem. During winter, if all leaves have abscised, the entire plant may be composed of heterotrophic tissues, and it maintains its metabolism by respiring stored starch.

FIGURE 10-1 (A) All organisms must obtain the energy needed to drive their metabolism. Most plants obtain energy through photosynthesis using the green pigment chlorophyll. All animals and fungi obtain their energy by either eating plants or by eating animals that ate plants. Some plants, called holoparasites, like this orange dodder (Cuscuta), do not carry out photosynthesis. Instead, they send specialized roots called haustoria [visible in (B) as short pegs] into the bodies of ordinary green, photosynthetic plants and extract sugars, water, and minerals from them and convert the sugars into the organic compounds they need. (B) Close-up showing haustoria penetrating the leaf of a host.
TABLE 10-1 Differences Between Photoautotrophs and Heterotrophs
Tissues often change their type of metabolism; young seedlings are white and heterotrophic while germinating underground; they survive on nutrients stored in cotyledons or endosperm. Seedlings become photoautotrophic only after they emerge into sunlight. Immature fruits may be green and photosynthetic; however, in the last stages of maturation, chloroplasts are converted to chromoplasts, and metabolism then depends on imported or stored nutrients (chromoplasts are plastids that contain large amounts of pigments other than chlorophyll). Young leaf primordia are green, but they grow more rapidly than their own photosynthesis would permit; they have a mixed metabolism of photosynthesis and carbohydrate import.
Photosynthesis is a complex process by which carbon dioxide is converted to carbohydrate. This involves endergonic reactions driven by adenosine triphosphate (ATP) and requiring new bonding orbitals filled by electrons carried to the reaction by reduced nicotinamide adenine dinucleotide phosphate (NADPH). Before this can happen, ATP and NADPH themselves must be formed in highly endergonic reactions driven by light energy. In order to understand this, you must first understand the nature of light and pigments along with the concept of reducing power.
![]() Energy and Reducing Power
Energy and Reducing Power
Energy Carriers
Energy enters the biological world through photosynthesis, a process that converts light energy to chemical energy. The sun’s light is captured by certain plant pigments that use the energy in chemical reactions. Unfortunately, the energized pigments can enter into only two chemical reactions, although plants have thousands of different reactions in their metabolism. Several theoretical ways exist of transporting energy from energized pigments into endergonic reactions:
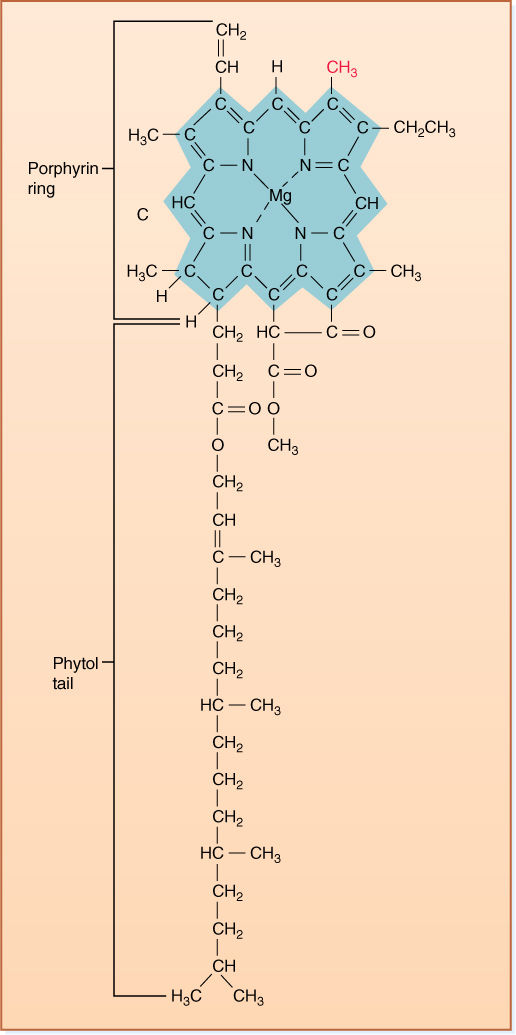
FIGURE 10-2 The tail of chlorophyll a contains only hydrogen and methyl functional groups, so it is hydrophobic and dissolves into the chloroplast’s membrane lipids, immobilizing it. The porphyrin ring system of alternating double and single bonds acts as an antenna that captures light energy. The magnesium atom carries the electrons involved in photosynthesis. If the colored methyl group were an aldehyde group (—CHO), the pigment would be chlorophyll b.
1. Allow the pigments to enter into every reaction necessary. A problem is that the energized pigments are large molecules; therefore, they are not very mobile and never move across membranes (FIGURE 10-2). Furthermore, they are too energetic; they can react with almost anything and would be difficult to control.
TABLE 10-2 Methods of Synthesizing ATP
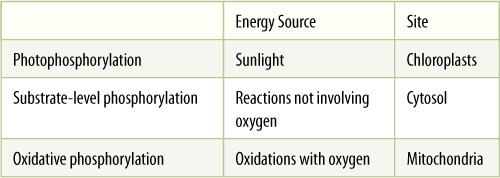
2. Allow the energized pigments to make one or several smaller, less energetic intermediates that can be moved and controlled easily. Such a method has evolved: Photosynthetic reactions produce ATP, an extraordinary molecule. Its high-energy phosphate bonds carry enough energy to force almost any reaction to proceed, and it can enter into almost every reaction for which energy is needed. In those that it does not enter, other energy carriers, often relatives of ATP, are involved; the most frequent is guanosine triphosphate, which also carries high-energy phosphate bonds.
Although ATP is an essential molecule, it constitutes only a tiny fraction of the plant body. Each molecule is recycled and reused repeatedly, thousands of times per second. ATP is converted to ADP and phosphate by metabolic reactions, but the phosphate can be reattached with a high-energy bond by the reactions of either photosynthesis or respiration. Each molecule is an energy carrier, shuttling between reactions that release energy and those that consume it.
There are three methods by which adenosine diphosphate (ADP) can be phosphorylated to ATP (TABLE 10-2). The first, photophosphorylation, involves light energy in photosynthesis; animals, fungi, and nonchlorophyllous plant tissues cannot perform photophosphorylation because they lack the necessary pigments and organelles. Instead, they respire some of the high-energy compounds they have consumed as food or imported by phloem. Compounds with high-energy phosphate groups are produced, and these compounds force their phosphate onto ADP, making ATP. This is substrate-level phosphorylation. In the last stages of respiration, ADP is phosphorylated to ATP by oxidative phosphorylation. Each process occurs in a distinct site within the cell, and each captures energy from distinct types of exergonic reactions. Photophosphorylation occurs only in chloroplasts in light, but substrate-level and oxidative phosphorylation occur in all parts of the plant at all times.
Reducing Power
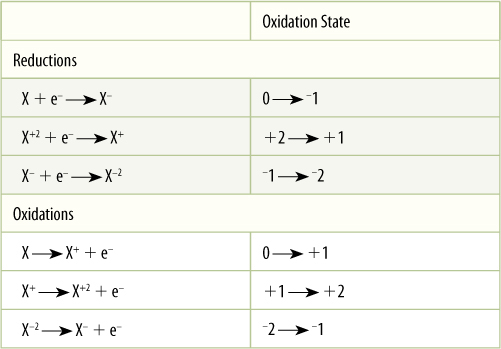
Earth’s atmosphere is about 21% oxygen; therefore, many compounds are found in their oxidized form: carbon as carbon dioxide (CO2), sulfur as sulfate (SO2—4), nitrogen as nitrate (NO3—), and so on. “Oxidized” means that an atom does not carry as many electrons as it could. In carbon dioxide, each oxygen can be considered to have pulled two electrons almost completely away from the carbon, and the carbon is said to be at a +4 oxidation state. This is speaking figuratively; electrons spend more time near the oxygen than they do near the carbon, sulfur, or nitrogen, but they are not torn completely away; these bonds are covalent, not ionic (TABLE 10-3).
When electrons are added to an atom, it becomes reduced. Think of a reduction reaction as one that reduces the positive charge on an atom and an oxidation reaction as one that increases the positive charge (TABLE 10-4). A preliminary rule of thumb is that oxidized compounds often (but not always) contain a great deal of oxygen, whereas reduced compounds contain hydrogen. Oxygen has a strong tendency to pull electrons away from an atom and raise that atom’s partial positive charge, but hydrogen becomes more stable by giving up electrons, reducing its partner’s partial positive charge. Electrons can be transferred only between atoms or molecules, so each reaction in Table 10-4 is only a “half reaction.” Every oxidation occurs simultaneously with a reduction. The full reaction is known as an “oxidation-reduction reaction,” or “redox reaction.”
TABLE 10-3 Calculation of Oxidation States

TABLE 10-4 Reductions and Oxidations
Whereas compounds in the environment are predominantly in the oxidized state because of our oxygen-rich atmosphere, most compounds in organisms are in the reduced state. Carbon is often in the form of carbohydrate, where its oxidation state is +0; nitrogen is frequently present as an amino group, NH3 (N3- H1+ H1+ H1+); and sulfur is present as SH2 (S2- H1+ H1+). Thus, in addition to needing energy, organisms also need reducing power, the ability to force electrons onto compounds. Reducing power is especially important to plants because they take in carbon dioxide and water—the most highly oxidized forms of carbon and hydrogen—and then convert them to carbohydrates, fats, and other compounds that are very reduced. Heterotrophs have less of a problem: When they consume plants, they get compounds that have already been reduced. They do need some reducing power, however, when they synthesize compounds that are very reduced, such as fatty acids.
Just as with energy, an optimum solution for moving and handling reducing power—electrons—is to use small molecules that are semistable and mobile. The two molecules used most often are nicotinamide adenine dinucleotide (NAD+; FIGURE 10-3) and the closely related nicotinamide adenine dinucleotide phosphate (NADP+). Both can pick up a pair of electrons and a proton, thereby becoming reduced to NADH and NADPH. When they reduce a compound by transferring their electrons to it, the proton is released and NAD+ or NADP+ is regenerated. Rather than having a large number of carrier molecules, the cell recycles each molecule, using it thousands of times a second as it shuttles between electron-producing and electron-consuming reactions.
Because NAD+ and NADP+ take electrons away from other molecules, they are oxidizing agents—they oxidize the material they react with. It is also possible to say that the material has reduced the NAD+ or NADP+. During the process, NADH and NADPH, two strong reducing agents, are formed. They have a powerful tendency to place electrons onto other molecules, reducing those molecules and becoming oxidized themselves (Figure 10-3B). The tendency to accept or donate electrons varies greatly and is known as a molecule’s redox potential (TABLE 10-5). Cells contain a variety of electron carriers that differ in their tendency to accept or donate electrons.
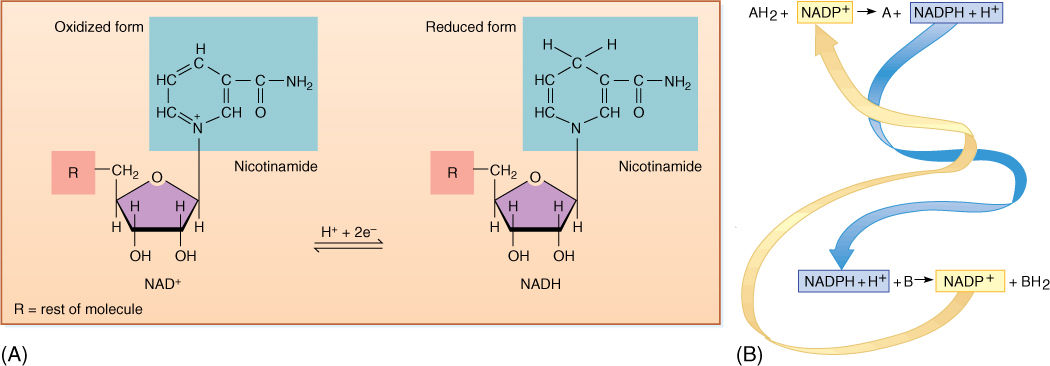
FIGURE 10-3 (A) Here, only the portion of NAD+ involved in carrying electrons is shown. The oxidized state (NAD+ and NADP+) carries a partial positive charge on the nitrogen and three double bonds in the ring. When it is reduced by two electrons, the positive charge disappears from the nitrogen, numerous bonding orbitals within the ring are changed, and the top carbon picks up a proton. No bonding orbitals are formed between NAD+ and the electron donor; therefore, NADH is free to diffuse away after picking up electrons. Similarly, as it donates electrons to some substrate, the ring bonding orbitals revert to the NAD+ form, none of which binds it to the substrate, so the NAD+ diffuses away. The same is true for NADP+. (B) In the upper reaction, two electrons are passed from a hypothetical molecule AH2 to NADP+: AH2 has been oxidized to A, and NADP+ has been reduced to NADPH + H+. The NADPH is free to diffuse to another site, where it passes the two electrons onto another hypothetical molecule, B, reducing it to BH2 and becoming oxidized back to NADP+.
TABLE 10-5 Redox Potentials of Electron Carriers
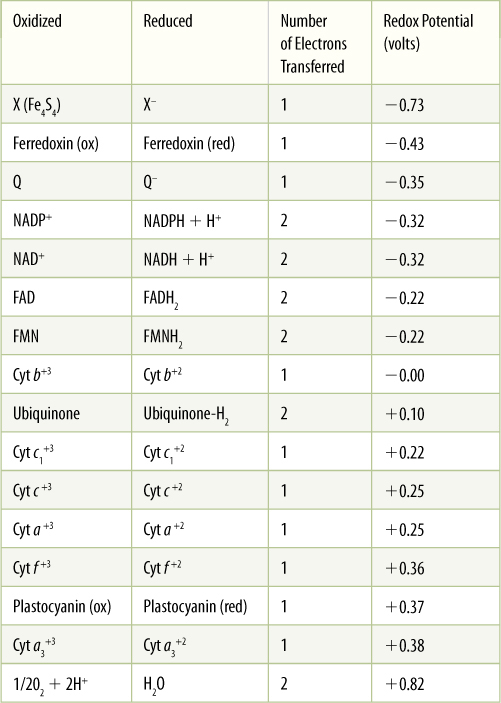
Compounds with a large negative redox potential (at the top of the list) tend to donate electrons and exist in the oxidized state: X, ferredoxin, NADP+, FAD. Compounds with a large positive redox potential (at the bottom of the list) tend to accept electrons and exist in the reduced form: Cyt a3+2, H2O.
Other Electron Carriers
Cytochromes
Cytochromes are small proteins that contain a cofactor, heme, which holds an iron atom (FIGURE 10-4); the iron carries electrons and cycles between the +2 and +3 oxidation states. Cytochromes are intrinsic membrane proteins; they are an integral part of the chloroplast’s thylakoid membranes and cannot be removed without destroying the membrane. Consequently, they carry electrons only between sites that are extremely close together within a membrane rather than diffusing throughout the stroma as NADPH does.
Plastoquinones
Plastoquinones, like cytochromes, transport electrons over short distances within a membrane (Figure 10-4B). After they pick up two electrons, they also bind two protons. Their long hydrocarbon tail causes them to be hydrophobic, so they dissolve easily into the lipid component of chloroplast membranes.
Plastocyanin
Like cytochromes, plastocyanin is a small protein that carries electrons on a metal atom—in this case copper. When oxidized, the copper ion is in the +2 oxidation state, but as it picks up the electron, it is reduced one level to the +1 oxidation state. Plastocyanin is loosely associated with chloroplast membranes; it can move a short distance along the surface, but it does not travel far.
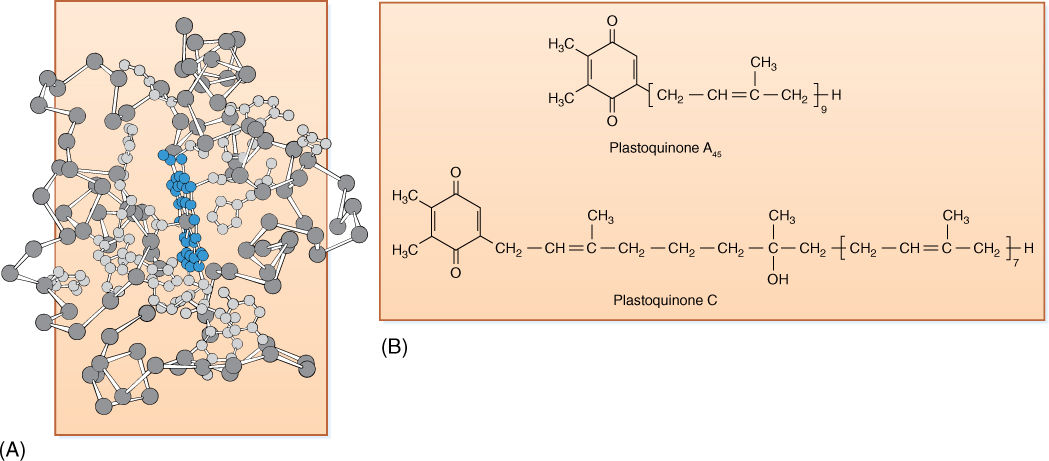
FIGURE 10-4 (A) In cytochromes, iron is not bonded directly to any amino acid but is held by heme, a box-like porphyrin ring similar to the portion of chlorophyll that holds the magnesium ion. (B) The distinguishing feature of the quinone class of electron carriers is that each has two ketone groups (the double-bonded oxygen) whose carbon atoms are part of a ring structure.
![]() Photosynthesis
Photosynthesis
As the name implies, photosynthesis is a process that uses light energy to synthesize something. The term is so general that it could be applied to many different types of reactions, but whenever a botanist uses the term “photosynthesis,” the reaction being discussed is the combination of carbon dioxide with water to form carbohydrate (FIGURE 10-5). Think about why these particular compounds are part of photosynthesis. First, both carbon dioxide and water are abundant and cheap, occurring almost everywhere in large quantities. The exception is the lack of water in severe deserts like the Sahara, where very little life exists simply because water is scarce. It is important to have a metabolism based on abundant compounds. It is necessary also for raw materials to be cheap; that is, the plant must be able to obtain them without expending much energy. Water and carbon dioxide are excellent because they diffuse into the plants automatically from soil, air, or water.
Another important quality of carbon dioxide and water is that they are very stable and contain little chemical energy, so it is possible to deposit a large amount of energy into them. The carbohydrates they form are a good means of storing energy because all reactions leading to carbohydrate breakdown have high energy-of-activation barriers. Despite being energy rich, carbohydrates are stable and chemically unreactive.
Finally, both the reactants and the products of photosynthesis are nontoxic; it is safe to absorb large quantities of carbon dioxide and water and to store high concentrations of carbohydrates. Many substances that are critical for life are extraordinarily toxic if they become even slightly concentrated; chlorine, sodium, ammonium, and many vitamins are just a few examples.
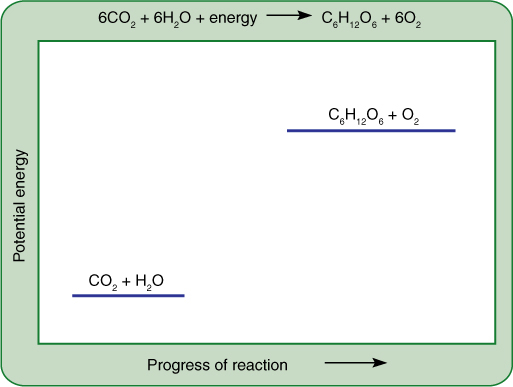
FIGURE 10-5 Although this chemical equation succinctly summarizes photosynthesis, it reveals virtually nothing of the reaction mechanism or the many carriers and enzymes that participate. We cannot draw a reaction diagram because photosynthesis does not occur by the direct interaction of six molecules of carbon dioxide with six of water; however, the relative potential energies can be shown, indicating that this is an endergonic process.
During photosynthesis, the carbon of carbon dioxide is reduced and energy is supplied to it, converting it to carbohydrate (Figure 10-5). The carbon atom in carbon dioxide is at the +4 oxidation state, whereas carbon atoms in carbohydrate are, in general, at +0. Four electrons must be found and placed into new bonding orbitals around the carbon atom to reduce it. This is not easy because carbon is more stable in the oxidized state than in the reduced state: Carbohydrates such as wood and sugar can burn, releasing energy, but carbon dioxide does not. Therefore, a source of electrons and a source of energy are necessary for photosynthesis: The electron source is water, and the energy source is light. Water and light, however, do not act on carbon dioxide directly; instead, they create the intermediates ATP and NADPH by a process called the light-dependent reactions (also called the thylakoid reactions). In a separate set of reactions, the stroma reactions (formerly known as dark reactions), ATP and NADPH interact with carbon dioxide and actually produce carbohydrate (FIGURE 10-6).
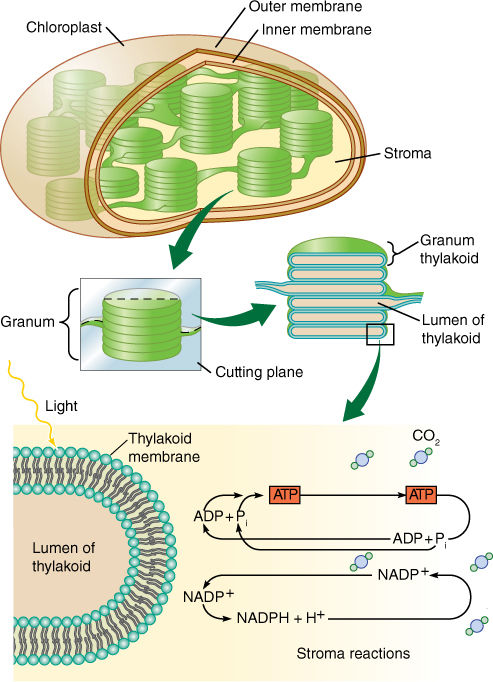
FIGURE 10-6 Light-dependent reactions of photosynthesis occur by means of membrane-bound carriers, but the actual formation of carbohydrate occurs in the chloroplast liquid (stroma). ATP-ADP and NADP+-NADPH diffuse between the two regions. No region of the chloroplast is far from a membrane, so the distances traveled are only a few hundred times the diameter of a molecule.
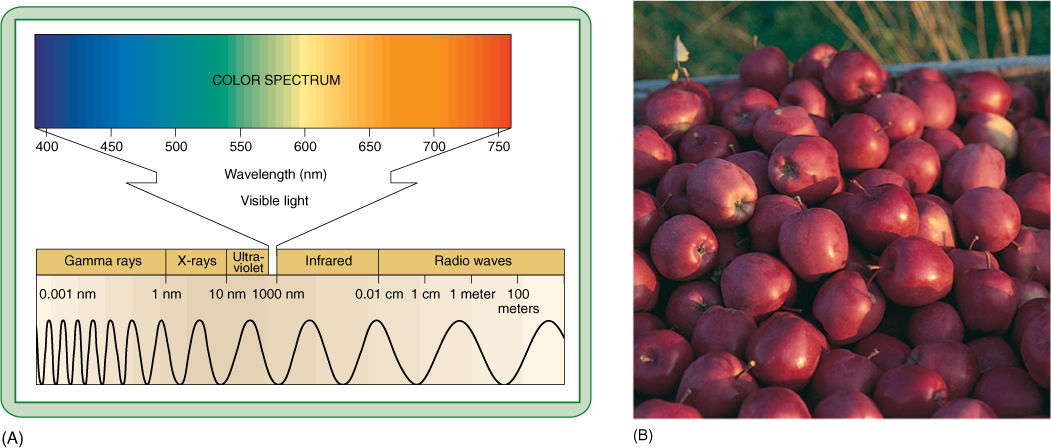
FIGURE 10-7 (A) On the left end of this electromagnetic spectrum are gamma rays. If treated as waves, they have extremely short wavelengths, less than 1 nm. If treated as particles, each quantum is highly energetic. Moving to the right, wavelengths become longer and quanta less energetic. The region to which our eyes respond—the visible region—is enlarged. (B) These apples have an abundance of pigments that absorb all colors except red. Earlier, while still developing, they had the pigment chlorophyll, which absorbs red and blue strongly, but reflects green.
The Light-Dependent Reactions
The Nature of Light
Light is one small segment of the electromagnetic radiation spectrum, which encompasses gamma rays, X-rays, ultraviolet light, infrared light, microwaves, and radio waves, in addition to visible light (FIGURE 10-7). Radiation can be thought of and treated physically either as a set of particles called quanta (singular, quantum), also called photons, or as a set of waves. The various types of radiation differ from each other only in their wavelengths and the amounts of energy each individual quantum contains. Short wavelengths (cosmic rays, gamma rays, and ultraviolet light) have relatively large amounts of energy in each quantum, whereas long wavelengths (infrared, microwaves, radar, and radio waves) have relatively little. Because humans use visible light for vision (that is why it is visible), we are more sensitive to and familiar with this region of the spectrum. We cannot see other wavelengths but we can feel infrared radiation as heat, and our bodies react to ultraviolet light by becoming tanned or burned. We distinguish differences in quantum energy (wavelength) of visible light as differences in color. Most of us see all wavelengths from red (760 nm) through orange, yellow, green, blue, indigo, to violet (390 nm). Certain insects see some near ultraviolet but in general, all animals see in the range from 350 to 760 nm, which is also the radiation that plants use for photosynthesis.
The Nature of Pigments
Most materials absorb certain wavelengths more than other wavelengths. If a substance absorbs all wavelengths except red, then red light either bounces off or passes through it, and the substance appears red to us. Any material that absorbs certain wavelengths specifically and therefore has distinctive color is a pigment, but we more often think of pigments as substances that absorb light as part of their biological function. Some pigments, such as melanin (the pigment of our skin), absorb light and thereby protect other light-sensitive substances. Other pigments, such as those in flowers, fruits, or the skins of animals, are important for the light they do not absorb, which gives the pigments their color, allowing them to be useful to the organisms in attracting mates, pollinators, or frugivores or in hiding from predators (Figure 10-7B).
Photosynthetic pigments transfer absorbed light energy to electrons that then enter chemical reactions. Because only absorbed energy can be used, a theoretically ideal photosynthetic pigment would be black: It would absorb and use all light, not letting any escape. The pigment should at least absorb high-energy radiation (ultraviolet light and gamma rays) instead of the fairly weak visible light. Rather surprisingly, the critical pigment, chlorophyll a, is not like this at all (see Figure 10-2). It absorbs only some red and some blue light, letting most of the rest pass through, especially high-energy radiation. In general, plants only capture about 5% of the energy available in the electromagnetic radiation that strikes them. Why does chlorophyll a lack what seem like ideal characteristics?
Plants and People
BOX 10-1 Photosynthesis, Global Warming, and Global Climate Change
Our atmosphere is critically important to life on Earth; ironically, its composition is the product of that very life. The free oxygen (O2) we breathe is produced solely by oxygenic photosynthesis; there is no other source. Oxygenic photosynthesis originated 2.8 billion years ago: We know this because for millions of years the newly produced oxygen reacted with iron, forming a worldwide stratum of rust in ancient rocks. After all iron had been oxidized, free oxygen began accumulating in the atmosphere, and its concentration has been increasing ever since. Simultaneously, photosynthesis pulled carbon dioxide out of the atmosphere, converting it first to 3-phosphoglyceraldehyde and then to all of the other organic compounds that exist. Every single organic molecule started out as carbon dioxide snared by RuBP carboxylase. Most organic molecules are digested and respired by aerobic organisms, a process that returns carbon dioxide back to the atmosphere rather quickly. But millions of tons of trees have died and fallen into stagnant swamps where a lack of oxygen prevented decay: The carbon in their wood—all of the cellulose, hemicellulose, and lignin—was converted to coal and did not return to the atmosphere. Petroleum also is probably derived from photosynthetically fixed carbon dioxide. The point is that respiration does not release all carbon back to the atmosphere; therefore, photosynthesis is gradually causing carbon dioxide concentration in the air to decrease.
Three groups of organisms have had especially important impacts on atmospheric carbon dioxide: coccoliths, mollusks, and humans. Coccoliths are microscopic algae that build shells of calcium carbonate, as do mollusks. When they die, their shells and the carbon they contain sink to the bottom of the ocean and decompose only slowly. All limestone and vast carbonate deposits on the ocean floor represent millions of tons of carbon dioxide removed from the atmosphere by clams, barnacles, and unimaginable numbers of algae.
We humans were no different from any other aerobic organism until we made a fateful discovery: how to use fire. Since then, we not only oxidize food in our mitochondria, but we also oxidize wood, coal, oil, and gas, putting carbon dioxide back into the atmosphere and raising its concentration measurably.
Why does the concentration of atmospheric carbon dioxide matter? Think of carbon dioxide as a pigment; its absorption spectrum is low for visible light but high for infrared wavelengths. Visible light from the sun passes easily through the atmosphere: It is not absorbed by nitrogen, oxygen, or carbon dioxide. As it strikes Earth’s surface, some is reflected immediately back out into space, and a small amount is absorbed by biological pigments such as chlorophyll in leaves or rhodopsin in eyes, where it powers photosynthesis or vision; however, most visible light has no effect other than to warm rocks, soil, and water, causing them to radiate the extra energy away as long-wavelength infrared light. Many of these infrared quanta pass directly back through the atmosphere without hitting a carbon dioxide molecule because the concentration of carbon dioxide is so low (0.03% of air), but many quanta are absorbed by atmospheric carbon dioxide molecules, causing them to become warmer. This energy is trapped in the Earth/atmosphere system and warms our world. This is called the greenhouse effect because the glass in greenhouses works the same way, as does the glass in a parked car. Carbon dioxide is a greenhouse gas.
An important balance exists between the atmospheric concentration of carbon dioxide and life: With less carbon dioxide, more heat would be lost and Earth would be frozen, like Mars. With more, more heat would be trapped and our world would be as hot as Venus, at 800°C, with lakes of molten lead. During the industrial age, we have been adding carbon dioxide to the atmosphere by burning oil, gas, and coal, and we have destroyed forest trees that can remove the carbon dioxide by photosynthesis. The concentration of carbon dioxide is increasing in the atmosphere, and the average temperature is also increasing. This is global warming, and it could cause mean temperatures to be 2°C or 3°C (3°F or 4°F) warmer in the next century.
Global warming is having numerous consequences. First, surface water of the oceans is becoming warmer; therefore, more water evaporates into the air. Much of our weather in North America comes as winds blow eastward across the North Pacific. The water is cold and the air picks up only enough moisture to keep the Pacific Northwest wet; by the time it moves to the Central Plains states, it has so little moisture left that only grasses, not forests, thrive. But as surface waters of the Pacific become slightly warmer, vastly more moisture will evaporate into the wind and be carried to the Mississippi drainage basin. This increased rainfall could cause much better farming conditions in the Central Plains, and catastrophic flooding in most river valleys where cities are located. El Niño years show the gigantic flooding that results from slight warming in just one area of an ocean.
Global warming is also causing rapid melting of snow and glaciers in mountains and of ice caps in the Arctic and Antarctic. It is difficult to comprehend, but Antarctica is a large continent covered by ice 1 to 2 miles thick. As the world’s ice fields melt, that water is added to oceans, increasing their volume and causing sea level to rise. Sea levels are rising even more because as ocean water warms, it expands. Coastal cities will be flooded, and so will coastal wetlands where ducks, geese, and hundreds of other species live.
The additional freshwater flowing from melting ice caps and flooding rivers has other impacts. Freshwater is lighter, more buoyant than seawater. It spreads outward from a river’s mouth as a cap, only gradually mixing with seawater as waves agitate the two. With more flooding, this cap becomes more extensive. Being fresh, marine algae cannot live in this layer, so they are forced into lower, darker waters where they cannot photosynthesize so well. The mixing of freshwater with seawater dilutes the nutrients in seawater, also slowing the growth of algae, and algae are of course the basis of the entire food web in the oceans: All ocean life is ultimately dependent on algae.
Another consequence of global warming is that not all areas are being affected equally. Wind patterns are being changed such that certain areas are becoming warmer, others cooler, some wetter, and others drier. This is global climate change; if circulation of ocean currents like the Gulf Stream is altered, climates in vast areas of the world will change dramatically. At what point will we decide that weather patterns have truly changed and that a particular city should be abandoned because of frequent flooding? Some agricultural areas will become too dry to farm, causing at least personal misery as crops fail, perhaps causing massive starvation. New areas will become optimal for farming, but it will take a great deal of confidence to be certain that the new weather patterns are stable enough for people to risk starting again in an unknown area. Millions of lives will be (are being) disrupted, some suddenly and catastrophically in floods and others slowly and inexorably as conditions decline. In addition to the problems global climate change causes us, we are also forcing it onto all of the other organisms with which we share the planet and which bear no responsibility for the damage we are doing.

FIGURE B10-1A After this tree is killed by the chain saw, it will stop photosynthesizing and will stop removing carbon dioxide from the atmosphere. Even if converted to lumber and used as part of a house, it will eventually decay or rot or be burned, being oxidized back to carbon dioxide and increasing the amount of CO2 in the air.
The Kyoto Protocol is a treaty designed to reduce production of greenhouse gases. Signed by 166 countries, it went into effect in 1994, but the United States—the largest producer of carbon dioxide—did not sign the treaty. Some people contend that if we do not act decisively, greenhouse gases will build up rapidly. Others point out that China and India—with combined populations of almost 2 billion people—are modernizing so rapidly that soon they will be producing more carbon dioxide than the United States is. They conclude that greenhouse gases will be produced as long as coal and oil are available and people need energy.
One human interaction with the atmosphere seems to be going well. The atmosphere protects us from harmful radiation from space by absorbing high-energy cosmic rays, X-rays, and ultraviolet light; very little of these wavelengths reach ground level. If not for water in the atmosphere, the first two would kill us outright. Ultraviolet light can induce mutations in our DNA and cause skin cancer and blindness, but atmospheric oxygen (produced by oxygenic photosynthesis) weakly blocks some ultraviolet light; fortunately, a by-product of its absorbing high-energy quanta is the conversion of oxygen to ozone (3O2 → 2O3), which is even more effective at blocking ultraviolet light. Man-made chlorofluorocarbons from air conditioners and cans of hairspray and deodorant, among other products, had been escaping into the atmosphere and destroying ozone, thus increasing the amount of ultraviolet radiation reaching us. Effort has been made to eliminate production of chlorofluorocarbons worldwide, and its levels in the atmosphere are dropping, thus allowing ozone to accumulate again and continue protecting us. This one success story shows that we have the power to protect the environment, the environment that protects us.

FIGURE B10-1B Tropical rain forests are not the only forests being damaged. Clear-cutting like this is common in the western United States and Canada.
First, chlorophyll, like all other biological pigments, does not use high-energy quanta because they have too much energy. Each is so powerful that it would knock electrons completely away from the pigment, disrupting bonding orbitals and causing the molecule to break apart. Notice in Figure 10-2 that all bonds in the chlorophyll ring system are double bonds that alternate with single bonds (conjugated double bonds); this bond system is excellent for absorbing quanta, but if even a single electron is knocked out, the entire structure becomes useless. It is selectively disadvantageous for a plant to produce a photosynthetic pigment that is destroyed by the light it absorbs; the molecule would break down, and all of the ATP that had been expended earlier in its construction would be wasted. Fortunately for our own molecules, the atmosphere’s ozone layer protects us by absorbing high-energy radiation.
Long-wavelength radiations, such as infrared and microwaves, have so little energy per quantum that they cannot appreciably boost an electron’s energy. Instead, they make the pigment molecule warmer, as in a microwave oven, but this is not especially useful for chemical synthesis. Visible light contains just the right amount of energy per quantum. When one of these quanta is absorbed by the pigment, an electron is activated—raised to an orbital of a higher energy level. We say that the electron and the molecule go from the ground state to an excited state (FIGURE 10-8). Under the right conditions, this high-energy, excited electron can be used in chemical reactions. If it is not used, it returns to its original, stable ground orbital by emitting a new quantum of light, one with less energy and a longer wavelength than the one that it absorbed. The release of light by a pigment is called fluorescence (Figure 10-8C).
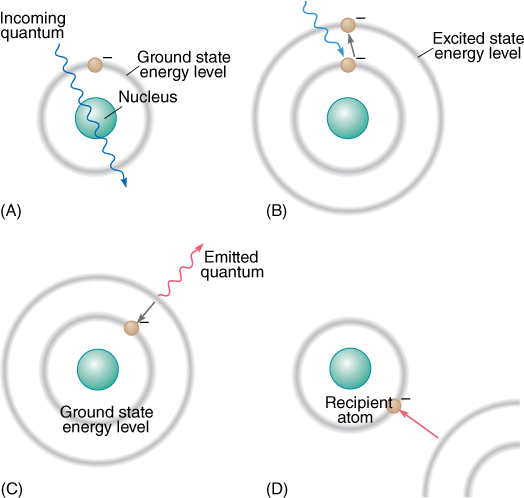
FIGURE 10-8 (A) If a quantum has the wrong wavelength for a pigment, it passes through the pigment’s bonding orbitals without being absorbed. Chlorophyll looks green because most green light passes through it. (B) If the quantum has the correct wavelength, the correct amount of energy, it is absorbed, and the electron must move to a new orbital whose energy level corresponds to the electron’s new energy load. (C) The excited state is unstable; it may stabilize itself by having the electron emit enough energy (fluoresce) to drop back to its original ground state energy level. (D) The electron can also be stabilized by moving to a more stable orbital on an entirely different atom. This is the critical process in photosynthesis; without this step, life would not exist.
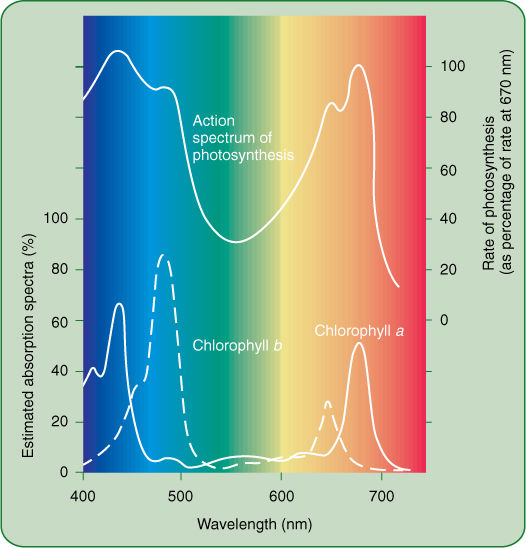
FIGURE 10-9 The absorption spectra of chlorophyll a and chlorophyll b and the action spectrum of photosynthesis. On the bottom axis is the wavelength of light, with short (blue) wavelengths to the left and long (red) ones to the right. The vertical axis of the absorption spectra is the amount of light absorbed by the pigment (scale on left); for the action spectrum, it is the amount of photosynthesis carried out (scale on right). Chlorophylls absorb little of the very short wavelength light at 400 nm, and little photosynthesis occurs; however, light at slightly longer wavelengths, about 425 nm, is absorbed well by chlorophyll a, and photosynthesis proceeds. Quanta with intermediate wavelengths pass right through the pigment, and photosynthesis is low, but in the 650 to 680 nm range (red), considerable absorption occurs. Because the absorption spectra of chlorophyll a and b differ, more wavelengths are harvested. If the two matched perfectly, chlorophyll b would be useless.
Two of the most useful pieces of information about a photochemical process are its action spectrum and the absorption spectrum of its pigment (FIGURE 10-9). An absorption spectrum is a graph that shows which wavelengths are most strongly absorbed by a pigment, whereas an action spectrum shows which wavelengths are most effective at powering a photochemical process. To initiate a photochemical process, light must first be absorbed; therefore, the action spectrum of a process must match the absorption spectrum of the pigments responsible. The absorption spectrum of chlorophyll a shows that it absorbs red light (especially 660 nm) and blue light (440 nm) very well and other wavelengths only slightly. It would be better if it could absorb a greater number of wavelengths, but it simply does not. Chlorophyll a is the essential photosynthetic pigment in all plants, algae, and cyanobacteria and has existed unaltered by evolution for about 3 billion years. To put this in perspective, the entire Milky Way Galaxy takes 250 million years to make one rotation about its center, so in 12 full rotations of our galaxy, no alteration in the structure of chlorophyll a has been selectively advantageous.
Accessory pigments are molecules that strongly absorb wavelengths not absorbed by chlorophyll a. The absorbed energy is passed on to chlorophyll a. In effect, the accessory pigments overcome the narrow absorption of chlorophyll a and broaden the action spectrum of photosynthesis. We know that accessory pigments are involved because the action spectrum of photosynthesis does not perfectly match the absorption spectrum of chlorophyll a. The most common accessory pigments in land plants are chlorophyll b and the carotenoids (FIGURE 10-10); algae have other types. Chlorophyll a and chlorophyll b are large, flat molecules with almost identical porphyrin ring structures. Their phytol tails are hydrophobic and dissolve into the lipid portion of the thylakoid membrane. When packed tightly in a membrane by their phytol tails, their porphyrin rings lie parallel to each other, which causes the electron orbitals of one molecule to interact with those of the two adjacent molecules. Hundreds of chlorophylls act somewhat like one molecule, and the energy absorbed by one can be rapidly transferred to another in a different part of the complex. This transfer, called resonance, allows chlorophyll b to absorb wavelengths that chlorophyll a would miss and then to transfer the energy to chlorophyll a for use in chemical reactions. Carotenoids are poor at this type of resonance and transfer only approximately 10% of their energy; they seem to be more important in absorbing excessive light and thus protecting chlorophylls.
Rather than occurring at random, the pigments are held in place by proteins called light-harvesting complex proteins that are also embedded in the thylakoid membrane. For example, one well-studied protein is known to bind 14 molecules of chlorophyll a and b and as well as four carotenoids. The entire set of 300 chlorophylls, carotenoids, and their proteins are called an antenna complex. When light strikes any pigment of an antenna complex, the energy is transferred to a reaction center, a structure that contains a pair of special molecules of chlorophyll a whose properties differ from those of all other molecules of chlorophyll a in the complex (FIGURE 10-11). The reaction center’s pair of chlorophyll a molecules are the only ones that can actually donate an excited electron onto an electron acceptor molecule.
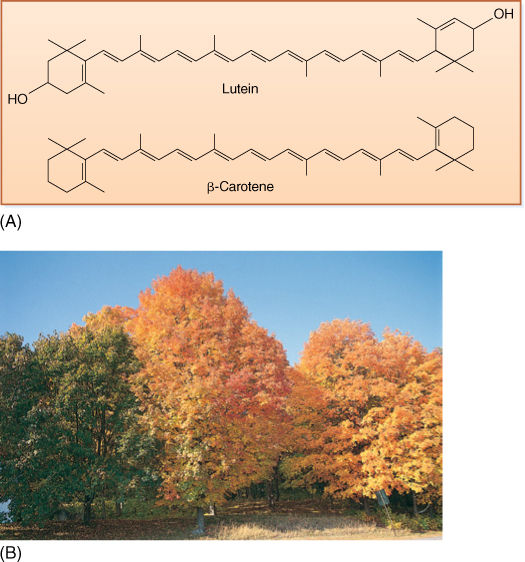
FIGURE 10-10 (A) There are two types of carotenoids: Carotenes lack oxygen, but xanthophylls (such as lutein) have it. Both are accessory pigments that protect the chlorophyll from excess sunlight. (B) Carotenoids are always present in leaves, but usually the abundant chlorophyll masks their presence. In autumn, chlorophyll breaks down, and we can see the carotenoids.
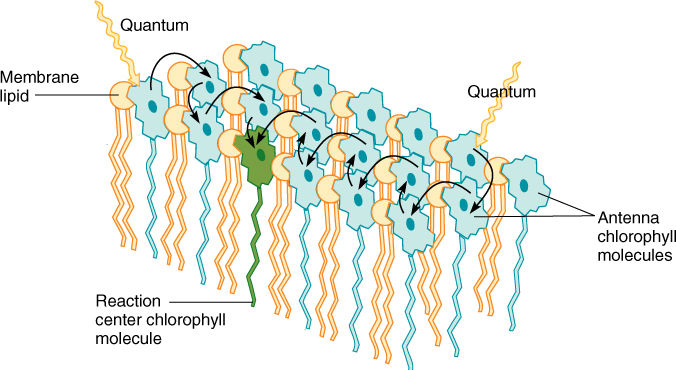
FIGURE 10-11 Only special chlorophyll a molecules—reaction centers—undergo the initial photochemical reaction of photosynthesis, but they are surrounded by other chlorophyll a molecules as well as accessory pigments; regardless of which pigment absorbs light, the energy is transferred to the reaction center.
It seems strange for chlorophyll molecules to pass energy from one to another, but it is necessary because chlorophyll is so poor at absorbing light. Even in bright sunlight, any particular chlorophyll molecule absorbs only a few quanta each second. This is much too slow to keep the later steps of photosynthesis operating properly. By having an antenna complex, the several hundred pigment molecules all transfer their energy into one reaction center, which thus acts as if it is absorbing thousands of quanta each second. Also, this organization allows all the electron carriers and other crucial molecules to be clustered around just the reaction center chlorophyll a dimer rather than being scattered among all the photosynthetic pigments.
When light energy reaches a reaction center in chlorophyll a, the electron is so unstable that it either reacts with almost anything or fluoresces, wasting its energy. Chlorophyll’s electron must react only with the proper molecule, which should be close enough for the reaction to take place instantly, before fluorescence can occur. All pigments and carriers that work together are packed into a granule called a photosynthetic unit, and the thylakoid membranes are filled with millions of these granular arrays (FIGURE 10-12). Each photosynthetic unit contains an antenna complex and reaction center, but in some photosynthetic units, chlorophyll b is plentiful, and in others, it is less abundant. Those with little chlorophyll b have been named photosystem I; those in which chlorophyll b is present at levels almost equal to a are photosystem II. The photosystem I units are involved in the following reactions.
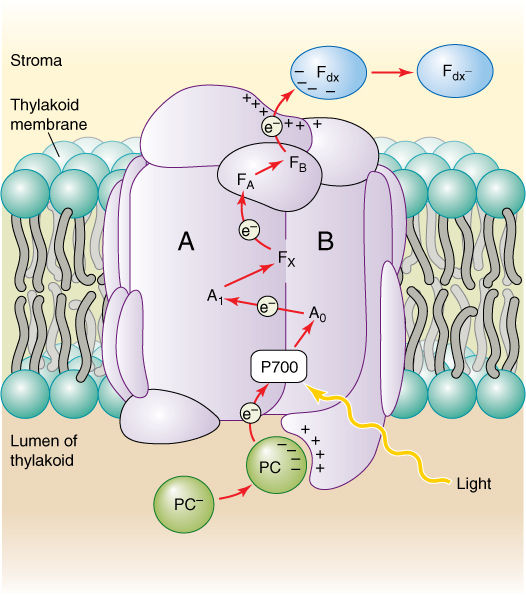
FIGURE 10-12 Reaction center of photosystem I. A and B are two major proteins that bind chlorophyll; many other components are clustered around these proteins. PC = plastocyanin; Fdx = ferredoxin; A0, A1, FX, FA, and FB are electron carriers.
Photosystem I
The pair of chlorophylls of the photosystem I reaction center is given the special name P700 because they absorb red light of 700 nm most efficiently. The energy excites an electron of P700, which is then absorbed by a membrane-bound electron acceptor known as “FX.” This is a transfer of an electron; no bonding orbital is formed. The exact chemical nature of FX is not known, but it contains iron and sulfur and is sometimes designated Fe4S4. When X absorbs an electron from P700, it becomes a powerful reducing agent, with a redox potential of -0.73 volts (Table 10-5). The transferred electron is still extremely unstable, and the reduced FX immediately passes it onto ferredoxin, which is also located in the thylakoid membrane (FIGURE 10-13). Ferredoxin is a small protein (10,500 to 11,000 daltons; a dalton is the weight of one hydrogen atom) with an active site consisting of two iron atoms bound to two sulfur atoms. Reduced ferredoxin is also a strong reducing agent, with a redox potential of -0.43 volts. Electrons are passed from ferredoxin to an enzyme, ferredoxin-NADP+ reductase, which then reduces NADP+, converting it to NADPH, as its name indicates. Ferredoxin carries only one electron, but two are needed simultaneously to reduce NADP+. Ferredoxin-NADP reductase carries two electrons, but it can be reduced one electron at a time; then it transfers those two electrons together to NADP+. Although NADPH is also a strong reducing agent, it is stable enough to move away from the membrane safely without the risk of reducing things indiscriminately, as the previous electron carriers might.
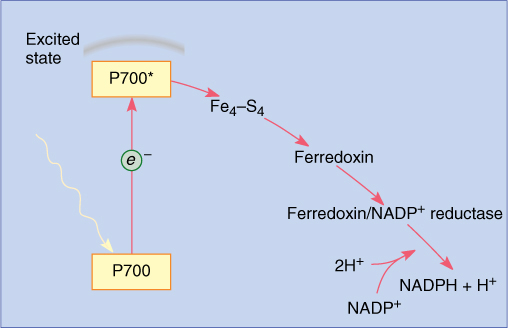
FIGURE 10-13 In photosystem I, energy is absorbed by a pair of P700 chlorophyll a molecules, raising two electrons to an excited energy level; from here, they pass onto Fe4—S4 (“FX”), then onto ferredoxin, and finally onto ferredoxin-NADP reductase. After two electrons have reduced ferredoxin-NADP reductase, they are transferred simultaneously to NADP+, reducing it to NADPH + H+.
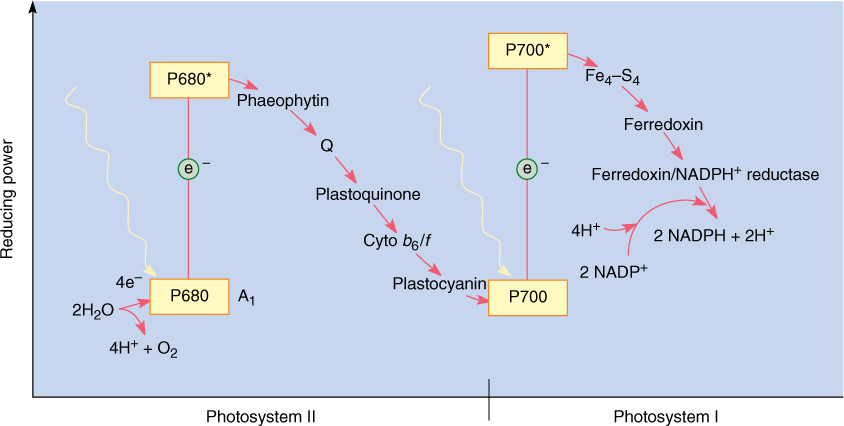
FIGURE 10-14 The two photosystems work together to transfer electrons from water to NADPH. The scale on the left indicates reducing power, the redox potential (see Table 10-5). The higher a molecule is in the chart, the greater its capacity to force electrons onto another molecule. Because of its shape, this diagram is called a Z scheme.
Photosystem II
Photosystem I efficiently produces NADPH, but the reaction center P700 chlorophyll a loses electrons during the process. In this oxidized state, bonding orbitals could easily rearrange, causing the molecule to break down and be destroyed. There must be a mechanism that adds electrons back to the P700, reducing it so that it can work repeatedly.
The mechanism that reduces P700 is photosystem II (FIGURE 10-14). Photosystem II can be best described by working backward from photosystem I: A molecule of plastocyanin, which contains copper, donates an electron to the chlorophyll a of the photosystem I reaction center. The plastocyanin is now oxidized, lacking an electron; it must reacquire one because it also is too expensive a molecule to donate just one electron and then never work again. It receives its new electron from a complex of cytochrome molecules, called the cytochrome b6/f complex, which in turn gets an electron from a molecule of plastoquinone. This receives electrons from another carrier, Q, a molecule of quinone, which in turn receives electrons from phaeophytin. Phaeophytin is actually a chlorophyll a molecule that does not contain a magnesium atom. Phaeophytin becomes oxidized as it donates an electron to Q, so it must obtain another electron, which it does when a chlorophyll a molecule absorbs light and is activated. This is a different chlorophyll a from the one in photosystem I; it is the reaction center of photosystem II and has the name P680.
We may seem to be going in circles: taking an electron from one chlorophyll a, P680, to pass it onto another chlorophyll a, P700, which then sends it to NADP+, but the physical differences between the two molecules of chlorophyll a are crucial. The one in photosystem II gets new electrons from water, not plastocyanin. The important thing is that water is cheap enough to just throw away after the electrons are removed. Water breaks down into protons (H+), which the plant uses, and oxygen (O2), which it discards. Whereas all electron carriers are large, expensive molecules that the plant must construct itself, water is simply brought in. The electrons are stripped off, the protons are used, and the oxygen is discarded through stomata. Ten quanta are needed to break down each water molecule.
Photosystems I and II together are an efficient system. Electrons are passed from water to P680 in photosystem II, their energy is boosted by light, and then they move through an electron transport chain—the various electron carriers—to P700 in photosystem I. Their energy is boosted by light again, and they pass through a short second electron transport chain to NADP+, reducing it to NADPH. This last step requires that protons be added to NADP+; these protons are present in the water surrounding the membrane (water is always a mixture of H2O, H+, and OH-). It would be simpler if photosystem I could receive electrons directly from water, but that does not happen. Besides, the electron transport chain between P680 and P700 is necessary for the production of ATP.
The Synthesis of ATP
The light-dependent reactions produce the reducing agent NADPH that actually places electrons onto the carbon of carbon dioxide in the stroma reactions, but the stroma reactions are highly endergonic and must be driven by being coupled to the exergonic splitting of ATP. The necessary ATP is also generated by the light reactions, but the process is indirect. It is photophosphorylation because light is involved, but a more specific name is often used: chemiosmotic phosphorylation. To understand it, we must take a closer look at the structure of chloroplasts. The inner membrane of chloroplasts folds inward, forming flattened sacs called thylakoids (FIGURE 10-15). In certain regions, these swell slightly and form rounded vesicles. All thylakoids in one region form vesicles at the same spot, so they occur in sets called grana (singular: granum). Thylakoids that lie between grana are frets. The liquid surrounding the thylakoid system is the stroma, but notice especially that there is another compartment, the thylakoid lumen.
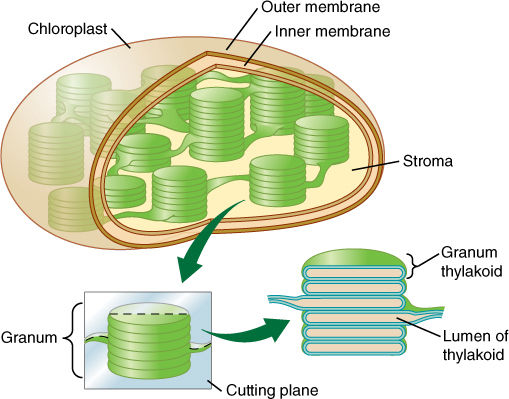
FIGURE 10-15 Grana are stacks of small thylakoid vesicles compressed together; frets are regions of thylakoid that connect one granum to another. The lumen of the thylakoid region is continuous with that of the fret region. The liquid surrounding all of the thylakoids is stroma.
The thylakoid lumen is a critically important compartment because some of the enzymes and electron carriers of the photosystems are embedded in the membrane layer facing the lumen, whereas other enzymes are in the membrane layer facing the stroma (FIGURE 10-16). Reactions that break down water and produce oxygen and protons are located on the lumen side of the thylakoid membrane, in the granum areas. This membrane is not permeable to protons; therefore, as light reactions run, the thylakoid interior accumulates protons, and their concentration increases. The molecules of ferredoxin-NADP reductase that generate NADPH are located on the other side of the membrane, facing the stroma. The protons they attach to NADP+ are those present as a result of the natural breakdown of water: H2O → H+ + OH-. As protons are absorbed, their concentration in the stroma decreases. Furthermore, during electron transport between P680 and P700, the electron carrier plastoquinone moves a proton from the stroma to the thylakoid lumen every time it carries an electron between phaeophytin and the cytochrome b6/f complex. This also contributes to the increased concentration of protons in the thylakoid lumen and to the decreased concentration of protons in the stroma.
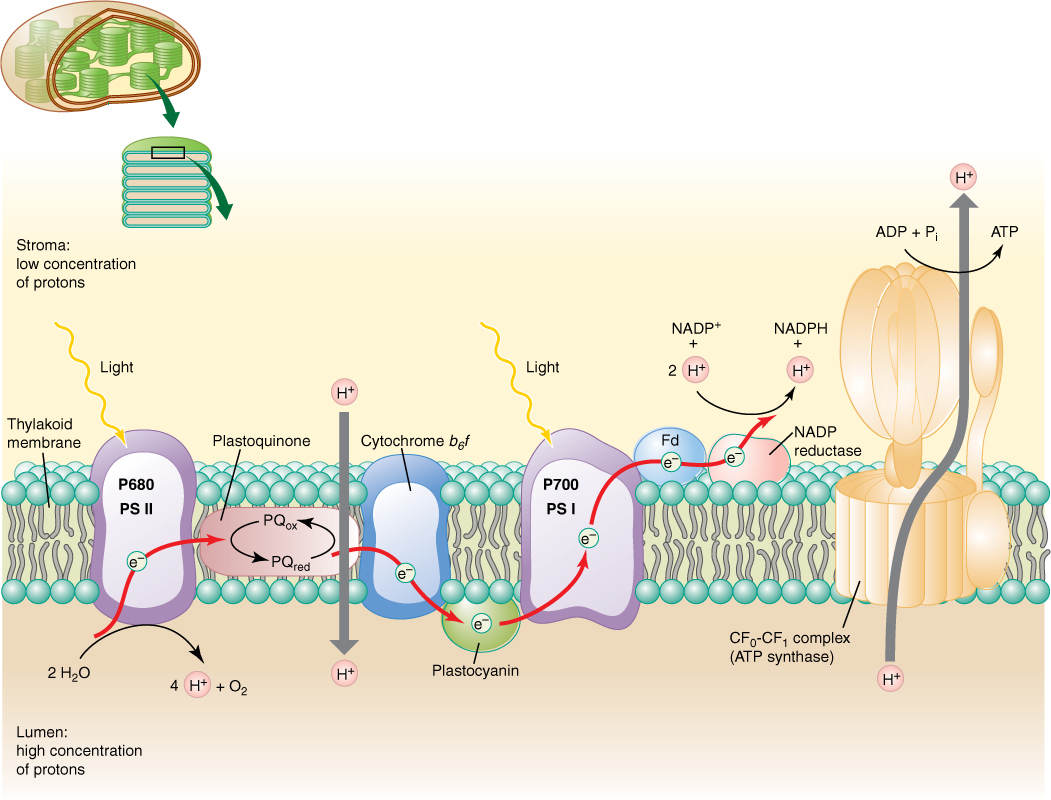
FIGURE 10-16 The water-splitting, proton-producing reactions of photosystem II take place on the lumen side of the thylakoid membrane. Plastoquinone is like NADP+ in that when it picks up electrons it also picks up a proton. This occurs on the stroma side of the membrane, but the reduced plastoquinone must diffuse to the other side of the membrane to pass electrons on to the cytochrome b6/f complex. The proton then dissociates and is deposited in the lumen, adding to the growing pool of protons. When NADPH is formed, it picks up protons from the stroma. This and the plastoquinone pumping result in a deficiency of protons in the stroma. Protons return to the stroma by passing through ATP synthetases; their passage is exergonic and powers the phosphorylation of ADP to ATP.
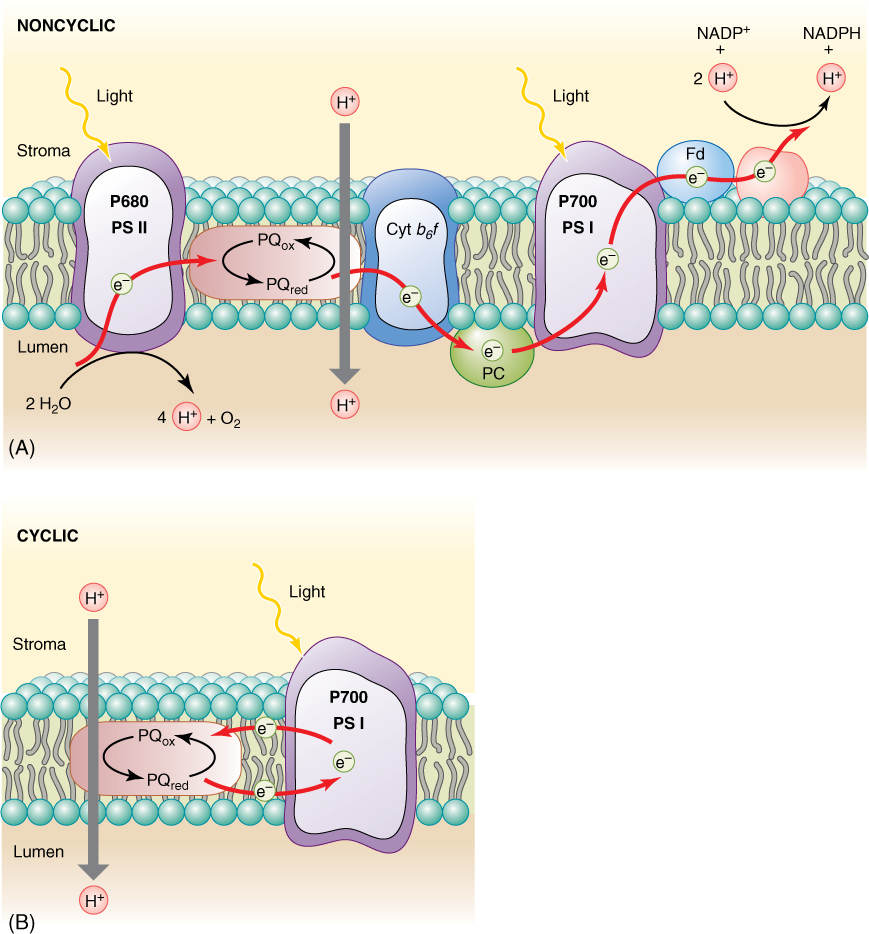
FIGURE 10-17 (A) In noncyclic electron transport, electrons flow through the Z scheme from water to NADPH. (B) Cyclic electron transport is much simpler: Electrons flow from P700 to plastoquinone, which carries a proton to the lumen and returns the electron to P700.
The strong difference between the concentrations of protons inside the thylakoid lumen and exterior to it in the stroma quickly becomes so powerful that protons begin to flow out of the lumen through special channels in the membrane. These channels are complex sets of enzymes that can synthesize ATP from ADP and phosphate; the entire complex is called ATP synthetase (Figure 10-16). The ATP synthetase of chloroplasts is known specifically as the CF0-CF1 complex. CF0 is the portion of the enzyme spanning the membrane where the actual proton channel is located. CF1 is the portion of the enzyme that phosphorylates ADP to ATP. The power required to force phosphate onto ADP and establish the high-energy bonding orbitals of ATP comes from the flow of protons through the ATP synthetase channels. In a car battery, electron flow through wires powers the starter; in chloroplasts, proton flow through ATP synthetase channels powers phosphorylation of ADP to ATP.
When electrons flow smoothly from water to NADPH, the process is called noncyclic electron transport (see Figure 10-14 and FIGURE 10-17). The chemiosmotic potential that builds up does not produce quite enough ATP for the stroma reactions: There is too little ATP relative to the amount of NADPH produced. This problem is overcome by an alternate route for electrons. After they reach ferredoxin in photosystem I, they can be transferred to the plastoquinones of photosystem II instead of being used to make NADPH. The plastoquinones carry the electrons along just as though they had gotten them from Q and use their energy to pump more protons into the thylakoid lumen. This is cyclic electron transport, and with it, chloroplasts make extra ATP without making extra NADPH, thus producing ATP and NADPH in the proper ratios for the stroma reactions. Cyclic electron transport is a simple light-powered proton pump, and similar types occur in bacteria. This may have been the original power system that evolved first, billions of years ago.
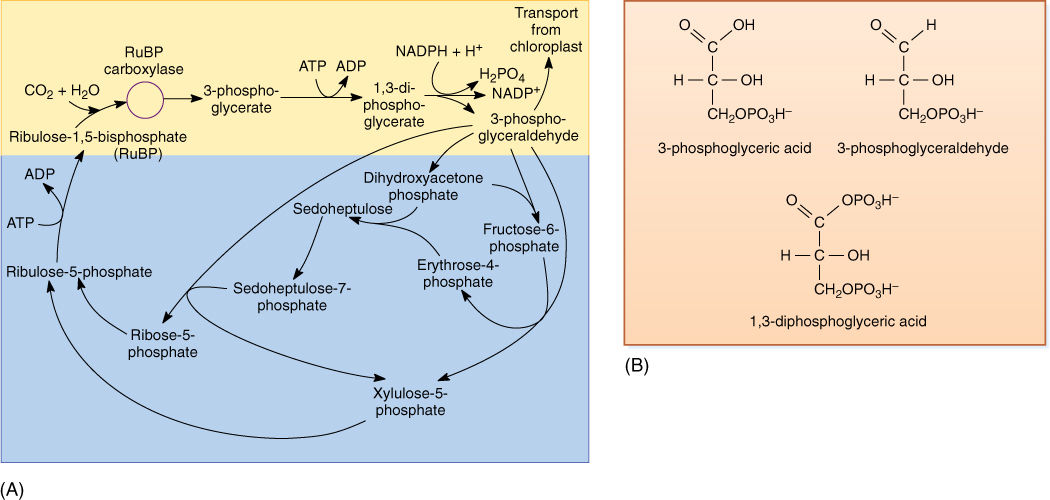
FIGURE 10-18 (A) In the yellow area are the first steps of the stroma reactions, also known as the C3 cycle; the product is two molecules of 3-phosphoglyceraldehyde. Some of this is transported out of the chloroplast, and the rest undergoes reactions (blue area) that form a new molecule of the acceptor, RuBP. (B) At various times, acids such as phosphoglycerate and malate are written as phosphoglyceric acid and malic acid; the “-ic acid” ending refers to the whole acid. The “-ate” ending refers to the acid’s anion, the negatively charged portion left after the proton dissociates. In protoplasm, most of the acids occur as free anions, not intact neutral acids still holding their protons.
The Stroma Reactions
Conversion of carbon dioxide to carbohydrate occurs in the stroma reactions, also called the Calvin/Benson cycle, or the C3 cycle (FIGURE 10-18). These reactions take place in the stroma, mediated by enzymes that are not bound to thylakoid membranes (see Figure 10-6). In the first step, an acceptor molecule (ribulose-1,5-bisphosphate; RuBP) reacts with a molecule of carbon dioxide. Because RuBP contains five carbons and one more is added from carbon dioxide, you might expect a product that contains six carbons. However, the new molecule breaks apart immediately, while still on the enzyme; stable bonding orbitals cannot be formed between all six carbon atoms while so many oxygen atoms are present and pulling electrons to themselves. Instead, orbitals rearrange and two identical molecules are formed that each contains three carbons: 3-phosphoglycerate, hence the name C3 cycle. The abbreviation PGA is often used for 3-phosphoglycerate.
The enzyme that carries out this reaction has many names; the most common is RuBP carboxylase (RUBISCO). This is one of the largest and most complex enzymes known—a giant complex of two kinds of protein subunits. There are eight copies of a small protein, each with a molecular weight of 14,000 to 15,000 daltons, and eight copies of a large protein, each with a molecular weight of 53,000 to 55,000 daltons. The entire enzyme has a molecular weight of about 480,000 daltons. Not only is the tertiary structure of each protein subunit important, but their quaternary structure as a complex is critical: When all eight subunits are properly assembled, the large subunits form the functional active site. RuBP carboxylase can constitute up to 30% of the protein in a leaf, making it the most abundant protein on Earth. Without it, there would be almost no life at all; all photosynthesis that produces oxygen is mediated by this enzyme. A few photosynthetic bacteria use an enzyme composed of just large subunits, lacking any small ones. RuBP carboxylase is crucial to the production of food; without it, heterotrophs would starve.
Like chlorophyll a, RuBP carboxylase is by no means ideal. Its active site recognizes and binds to carbon dioxide only poorly, and it has low substrate specificity, frequently putting oxygen rather than carbon dioxide onto RuBP. Yet this enzyme is highly conserved evolutionarily. The amino acid sequences of RuBP carboxylase from all plants are virtually identical. Apparently, all mutations that cause any change in structure, however slight, disturb the active sites and are selectively disadvantageous.
It is important to realize that the first step of the stroma reactions is carboxylation only. Electrons and energy are added in the next two steps: ATP donates a high-energy phosphate group to the 3-phosphoglycerate, converting it to 1,3-diphosphoglycerate, which then is reduced by NADPH to 3-phosphoglyceraldehyde (PGAL); a phosphate comes off in this step also. The carbon is now both reduced and energized.
The rest of the stroma reactions are complex, but the important point is that as they operate some 3-phosphoglyceraldehyde can be taken out of the chloroplast and used by the cell to build sugars, fats, amino acids, nucleic acids—basically anything the plant needs. The rest of the PGAL remains in the chloroplast and undergoes several more stroma reactions, which convert it to RuBP, the original acceptor molecule. The principle involved is important: To incorporate carbon dioxide, the plant needs the acceptor RuBP, and the two react on a one-to-one basis. To assimilate large amounts of carbon dioxide, the plant either needs large amounts of RuBP or needs to use a few RuBP molecules repeatedly. Plants use the second strategy. As 3-phosphoglyceraldehyde is formed, some of it is reconverted to RuBP by the rest of the stroma reactions, and some is exported to the cytoplasm. The chloroplast does not need to import quantities of RuBP from the rest of the cell; it just recycles the small amount that it has. Imagine a chloroplast that has 1,000,000 carbon atoms inside it as the various intermediates of the stroma reactions; after three carbon dioxides have been assimilated, there are 1,000,003 carbon atoms. When one molecule of 3-phosphoglyceraldehyde is exported, the carbon pool returns to 1,000,000, and a steady state is maintained. In very young leaves with growing chloroplasts, little or no 3-phosphoglyceraldehyde is exported; it is retained, and the pools of C3 metabolites increase in numbers of molecules.
Anabolic Metabolism
3-phosphoglyceraldehyde is an amazingly versatile molecule: Using it plus water, nitrates, sulfates, and minerals, plants construct everything inside themselves. The entire fabric of the organism can be synthesized. This is also the basis of all animal metabolism because animals either eat plants or eat other animals that eat plants.
Most biological molecules are larger than 3-phosphoglyceraldehyde so it must be rearranged and altered in the cytoplasm to build up larger, more complex molecules. This constructive metabolism is called anabolism, and it consists of anabolic reactions.
Anabolic pathways are numerous, but two are especially important with regard to energy metabolism: the synthetic pathways of polysaccharides and fats, which are storage forms of energy and carbon. The NADPH and ATP produced by photosynthesis are excellent sources of energy, but they cannot be stored for even a short time. They are so reactive and unstable that they would break down. A plant cannot stockpile them to survive times when photosynthesis is impossible nor can they be transported over long distances; therefore, even if leaves had an abundant supply, roots would starve.
Several types of storage compounds have evolved that solve these problems.
1. Short-term storage: ATP and NADPH can be used within the cell and last only briefly.
2. Intermediate-term storage: The simple sugar glucose and the disaccharide sucrose are stable enough to be moved from cell to cell, either in the vascular tissue of a plant or in a blood stream. They are also sufficiently stable to last for weeks or months. A problem with storing large quantities of monosaccharide or disaccharide is that they cause cells to absorb water by osmosis.
3. Long-term storage: Starch is a large, high-molecularweight polymer of glucose, too large to be transported. It is even more stable than glucose, lasts for years, and does not cause the cell to absorb water. Lipids are an even more concentrated storage form of energy that can be synthesized rapidly and stored in large quantities.
The Synthesis of Polysaccharides
The anabolic synthesis of glucose is gluconeogenesis (FIGURE 10-19). In reactions similar to those of C3 metabolism, part of the PGAL exported to the cytoplasm is converted to dihydroxyacetone phosphate; one molecule of this condenses with one molecule of unconverted 3-phosphoglyceraldehyde to form the sugar fructose-1,6-bisphosphate. This loses a phosphate to become fructose-6-phosphate, and part of this is rearranged, converting it to glucose-6-phosphate. Both fructose-6-phosphate and glucose-6-phosphate are versatile, useful molecules that enter many metabolic pathways. In plants, the glucose-6-phosphate is polymerized into polysaccharides: amylose, amylopectin, or cellulose.
Both amylose and amylopectin are components of starch. Amylose is an unbranched polymer of glucose, containing between 500 and 20,000 glucose residues. Amylopectin is also a polymer of glucose, but it branches and can be extraordinarily large, with as many as one million glucose residues in each molecule. The enzymes that add glucose to amylose and amylopectin can only work at the ends of the molecules; because amylose has only one end, it is synthesized slowly, whereas amylopectin might have thousands of enzymes simultaneously adding glucose to the thousands of its branches. Similarly, amylopectin can be digested back to glucose monomers much more rapidly than amylose.
The speed of starch degradation is important to us animals as well as to plants. Most of our food is rich in starch, and its digestion begins even before we swallow because our saliva contains salivary amylase. We digest the amylopectin portion of starch rapidly, which makes glucose available to our bloodstream quickly (such foods have a high glycemic index). In contrast, the amylose portion of starches is digested more slowly.
Plants store starch in both chloroplasts and amyloplasts. Other than a few rare exceptions, animals never synthesize starch, we store glucose as glycogen instead. During daylight hours, as photosynthesis is occurring, most of the 3-phosphoglyceraldehyde is retained within the chloroplast and gluconeogenesis converts it to glucose. It is polymerized to starch within the chloroplast, and by afternoon, chloroplasts are usually swollen because of the large starch grains they contain. At night, the starch is depolymerized back to glucose, which is transported from the chloroplasts to the cytosol. Much of it is then converted to sucrose, loaded into phloem, and transported to various parts of the plant that need carbohydrate. By morning, most chloroplasts have little or no starch left in them. In organs involved in long-term storage of starch, such as potato tubers and starchy seeds like wheat and rice, sugar moves from the phloem into parenchyma cells, then enters amyloplasts where is repolymerized to starch again.

FIGURE 10-19 (A) Gluconeogenesis is an anabolic pathway in which large molecules are built up from small ones. This process may occur in chloroplasts, amyloplasts, or cytosol. (B) Most of the sugar (sucrose) we eat and drink comes from sugar cane, which is a type of grass with large stems more than 2 m tall. This harvester cuts the shoots off at ground level, slices the stems into pieces called “joints,” and loads them into the cart behind the tractor. Pieces of leaves are so light they blow away. (C) These “joints” of sugar cane must be taken to a refinery and have their sap squeezed out and refined into raw sugar within 18 hours. Immediately after the stems are cut in the field, enzymes begin to convert the disaccharide sucrose into the monosaccharides glucose and fructose, both of which are less valuable than sucrose. (D) Sucrose is obtained by boiling cane sap to concentrate the sucrose until it becomes so concentrated it crystallizes. The crystals are centrifuged out and stored in giant piles as “raw sugar,” shown here; the remaining liquid is molasses. Raw sugar is converted to refined (white) sugar by dissolving it again and reboiling it to recrystallize it, after which it is filtered. The raw sugar in this warehouse will last indefinitely if it is kept dry.
![]() Environmental and Internal Factors
Environmental and Internal Factors
A plant’s photosynthesis is affected by its environment in many ways.
Light
From a plant’s viewpoint, light has three important properties: (1) quality, (2) quantity, and (3) duration.
Quality of sunlight refers to the colors or wavelengths it contains. Sunlight is pure white because it contains the entire visible spectrum. During sunset and sunrise, sunlight passes tangentially through the atmosphere, and a large percentage of the blue light is deflected upward; consequently, light at ground level is enriched in red, which is easily visible. This period of red-enriched light lasts only a few minutes and probably has little effect on photosynthesis. At noon, sunlight passes nearly vertically through the atmosphere, more blue light is transmitted, and even though the blueness of the sky suggests that all reds, greens, and yellows have been blocked, in fact, enough of all of these wavelengths penetrate to Earth’s surface to allow efficient photosynthesis. This is true of plants in deserts, grasslands, and the top layer—the canopy—of a forest; however, herbs and shrubs that grow near soil level in a forest are understory plants, and the light that they receive has already passed through the leaves of the canopy (FIGURE 10-20). As light penetrates those leaves, red and blue are absorbed by chlorophyll, so the dim light received by understory plants is especially depleted in these critical wavelengths. It is selectively advantageous for them to have extra amounts of accessory pigments so that they can gather the wavelengths available and pass the energy on to chlorophyll a. Similarly, algae that grow near the surface of lakes or oceans receive complete light, but water absorbs red and violet. Algae at deep regions receive mostly green and blue light and must have special accessory pigments capable of absorbing these wavelengths efficiently (FIGURE 10-21).
Quantity of light, which refers to light intensity or brightness, is affected by several factors. More light is available for photosynthesis on a clear than on a cloudy day; understory plants receive dim light; lower branches and branches on the shaded side of a plant receive less light (Figure 10-20B). Plants growing in the shadow of mountains or in deep canyons receive much less light than plants that grow on slopes that face the sun. Plants growing near the equator receive intense light because the sun is always more or less directly overhead at noon, whereas plants near the poles receive very little light. Even during the summer the sun is low at noon, and light is scattered by the atmosphere.
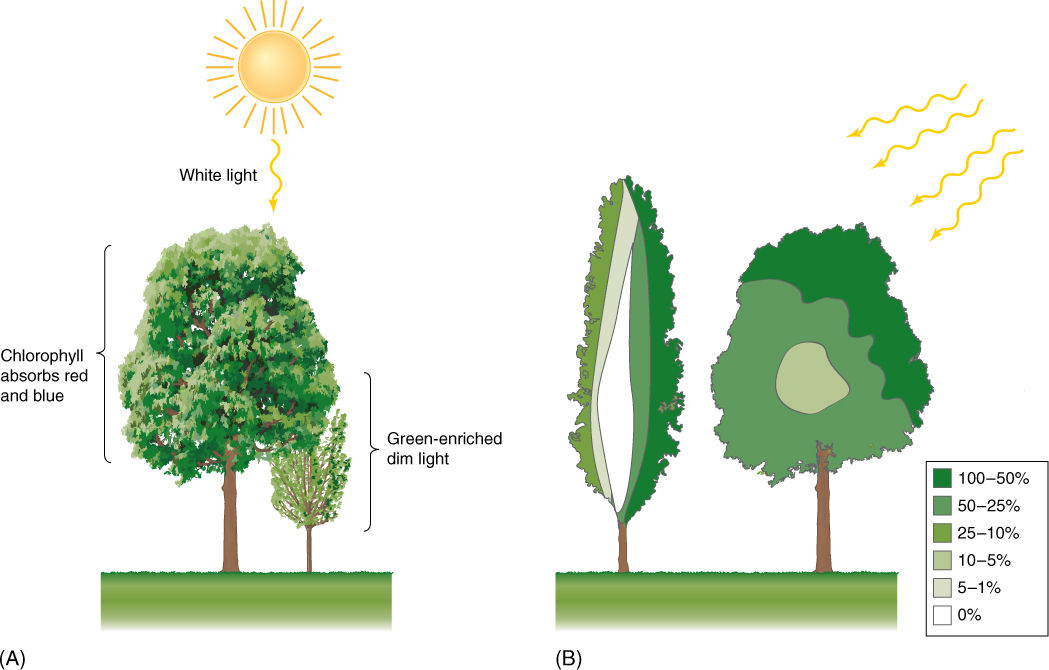
FIGURE 10-20 (A) Plants growing in the shade of other plants receive not only dim light, but light depleted of red and blue. (B) Not all leaves of a tree receive equal amounts of light; this self-shading is more severe in latitudes farther away from the equator. Colors indicate the percentage of full sunlight that reaches the various parts of a tree.

FIGURE 10-21 Many kelps (brown algae) grow in deep ocean water and receive mostly green and blue light. Their accessory pigments absorb these wavelengths; therefore, the kelps appear yellow-brown to us.
Think about how intensity of sunlight varies during the day and affects photosynthesis. Examine the solid line labeled “300 ppm CO2” in FIGURE 10-22. This was derived from many experiments in which the rate of photosynthesis was measured for plants grown under different intensities of light, but all with 300 ppm carbon dioxide in the air. Near the left side at point (a), those plants that received dim light absorbed little carbon dioxide, whereas those grown in brighter light absorbed more carbon dioxide. Under these normal levels of carbon dioxide, light is the limiting factor. Photosynthesis is slow on dull, overcast days but faster on brighter days. At point (b) in the graph, plants that received more light did not photosynthesize faster than those that received slightly less. Where the curve turns flat, there was enough light to saturate the process. In these conditions, the limitation was lack of carbon dioxide; in the experiments in which more carbon dioxide was available (blue line, 1000 ppm CO2), photosynthesis went faster. Thus, at point (b), carbon dioxide was the rate-limiting factor.
Under conditions of adequate water, light, and fertilizers, the rate of photosynthesis in whole plants increases with increasing carbon dioxide concentrations, being 30% faster with concentrations between 600 to 750 ppm. It should be noted that the experiment in Figure 10-22 was performed in the 1970s, but by 2015, atmospheric carbon dioxide already reached 400 ppm.
At point (c), light was so intense that it damaged the plant by overheating it and bleaching the pigments.
At point (a), if the lack of light prevents photosynthesis from proceeding faster, there must be adequate amounts of carbon dioxide. As soon as ATP and NADPH are produced, they move to the stroma and are used by the waiting enzymes and carbon dioxide; then ADP and NADP+ diffuse back to the thylakoid membranes and wait for another quantum. Conversely, at point (b), where light is bright and the low concentration of carbon dioxide is rate limiting, there is so much light that as quickly as ADP and NADP+ come to the thylakoids, they are reprocessed immediately into ATP and NADPH. These then move to the stroma, where they must wait for a carbon dioxide molecule.
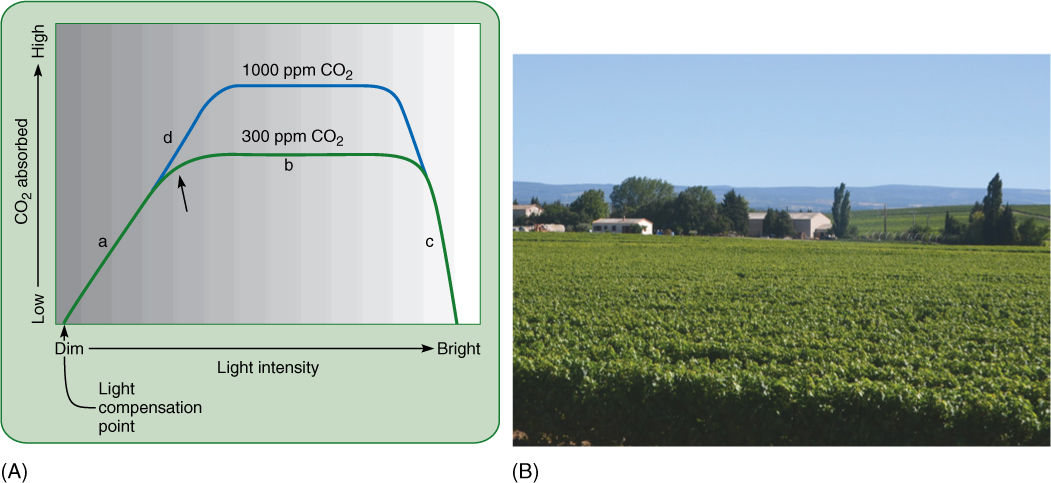
FIGURE 10-22 (A) Light and photosynthesis; details are given in text. The unit ppm is parts per million; for 300 ppm, in 1 million liters of air, 300 liters are carbon dioxide. (B) These grape vines have abundant sunlight, water, and fertilizer. They could probably photosynthesize more rapidly only if more carbon dioxide were present.

FIGURE 10-23 (A) While young, the leaves of dusty miller are completely obscured by trichomes, protecting the leaf from strong sunlight and insects. (B) These cacti (Epithelantha) live in environments where sunlight is extremely intense; their spines are so abundant and closely spaced that they shade the stem and prevent chlorophyll from being damaged. (C) Plants of Dudleya brittonii are protected from excess sunlight and water loss by a thick layer of white wax; this particular wax strongly reflects ultraviolet light.
To the left of the light compensation point, it appears that there was no photosynthesis, even though some dim light was provided. The problem actually lies with the technology of measuring photosynthesis. Either the amount of carbon dioxide absorbed or the amount of oxygen released must be measured, but both of these gases are involved in respiration as well as photosynthesis. To the left of the light compensation point, photosynthesis was absorbing carbon dioxide more slowly than respiration was releasing it; thus, it appears as though no photosynthesis occurred. The same problem arises when we try to measure respiration: Photosynthesis distorts the measurement, but then we can turn off the lights and stop photosynthesis. The light compensation point is the level of light at which photosynthesis matches respiration. Plants that are grown for a long time in conditions below the light compensation point respire faster than they photosynthesize; they gradually consume their reserve carbohydrates and fats and starve to death. For plants grown in light brighter than the light compensation point, photosynthesis exceeds respiration, and the extra sugar can be used for growth and reproduction.
When considering the effects of the amount of light, we need to consider the shape and organization of a plant. Photosynthesis in most ordinary leaves is saturated even on overcast days: One-quarter to one-half of ordinary sunshine is all most leaves can use. In contrast, many lower leaves on trees and bushes are shaded by upper leaves, and they may be exposed only to small flecks of sunlight that move irregularly over the leaf’s surface. Even on sunny days, many of the lower leaves are not receiving enough light to photosynthesize at their maximum capacity. Consequently, whereas an upper leaf maybe saturated in partial sunlight, most plants are not: As light intensity increases, more of their lower leaves photosynthesize faster so total photosynthesis for the entire plant increases.
In the brightest environments, the air is so clear that sunlight is frequently too intense during summer months. Protective adaptations are necessary, and in many species, mechanisms have evolved that provide shade. A common method is the production of a thick layer of dead trichomes, plant hairs (FIGURE 10-23). A heavy coating of wax can also reflect light, and cutin is especially good at absorbing the more harmful short wavelengths. Part of the value of carotenoids and other accessory pigments is that they shade the chlorophyll, absorbing some of the most damaging wavelengths.
The intensity of light, the actual number of quanta that strike a given area per unit time (e.g., that strike 1 cm2/s), is greatest at noon in midsummer when the sun is most directly overhead and is less whenever the sun is lower—morning, evening, and winter. Light may be too intense at midday but optimal when the sun is lower; some species (iris, eucalyptus) have adapted to this by means of vertical leaves. The lamina face is exposed fully only in mornings and afternoons, but at noon, only the leaf edge is exposed. In other plants, leaves orient vertically automatically when stressed—they wilt and hang down.
Understory plants of forests are adapted to low light. If a roadway is cut into a forest and plants adjacent to the cut are suddenly exposed to full sunlight, the shock may cause them to wilt and die. The same phenomenon occurs when trees are blown down during storms, floods, or avalanches and the surrounding plants are exposed.
The duration of sunlight refers to the number of hours per day that sunlight is available. At the equator, days are 12 hours long throughout the year. Farther north or south, days become longer in summer; maximum length occurs near the poles, where the day is 24 hours long in midsummer, and only night occurs in midwinter. In middle latitudes, winter days are short, and sunlight is weak because the sun is so low in the sky. Under these conditions, even evergreen plants are unable to undergo very much photosynthesis; however, because temperatures are low, the plants are growing little and have a low rate of respiration. Even deciduous trees and biennials can survive by means of stored nutrients.
During summer, days are longer and light is brighter because the sun is higher in the sky. The amount of energy obtained by photosynthesis easily exceeds that consumed by respiration and growth. In many plants, longer days cause greater amounts of photosynthesis, but in others, chloroplasts become so full of starch that photosynthesis stops, even though light is present. At night, starch is converted to sugar, which is then transported out of chloroplasts and can be used for growth or stored in amyloplasts in tubers, corms, or other such organs. By morning, leaf chloroplasts can resume photosynthesis.

FIGURE 10-24 (A) These living stone plants (Lithops) of African deserts conserve water in several ways. They have only two leaves at a time; when two new ones form, the old two die. Not enough water is available for four leaves. The leaves are fleshy and pressed together, such that they form a cylinder with minimal surface area through which water can be lost. (B) This is a plant of Haworthia cooperi. Its short stem is located several centimeters underground, and it produces many cylindrical leaves that are just long enough to reach the soil surface. The tips of the leaves, visible here, have transparent epidermis, and the mesophyll is also transparent because it has no intercellular spaces. Consequently, light enters through the window-like tip and then passes deep into the rest of the underground leaf, where cells with chloroplasts are located: photosynthesis actually occurs underground. Because most of each leaf is subterranean, they stay cooler in summer, they are less visible to animals, and the air in the soil is richer in carbon dioxide than is air above ground. (C) This transverse section of a leaf of Aloe vera is so transparent you can read through it. It is excellent for transmitting light to parts of the leaf that are underground.
Leaf Structure
Leaf structure of most temperate and tropical plants is quite standard: palisade parenchyma above and spongy mesophyll below. This structure is excellent for absorbing carbon dioxide but inefficient for conserving water. If plants of hot, dry habitats had this leaf architecture, they would have to keep their stomata closed so much of the time that they would starve. Instead, their leaf cells are frequently packed together without intercellular spaces. Water loss is reduced because the small internal surface area retards water evaporation, but with so little surface area, it is difficult to dissolve carbon dioxide from the air into the cytoplasm. This slows photosynthesis, but apparently this tradeoff, slow growth versus water conservation, is selectively advantageous.
Another method of minimizing water loss while maintaining photosynthesis is to reduce external surface by means of cylindrical leaves. Water movement from interior air spaces to exterior air is minimized because so few stomata are present (FIGURE 10-24). Photosynthesis is reduced because absorption of carbon dioxide is slowed.
Water
The amount of water available greatly affects photosynthesis. Most plants keep their stomata open during the day, permitting entry of carbon dioxide, but water is inevitably lost. At night, carbon dioxide cannot be used, and stomata are closed, retaining water within the plant. If the soil becomes dry and water is not readily available, a plant keeps its stomata closed even during the day, and carbon dioxide cannot enter (FIGURE 10-25). The small amount of carbon dioxide produced by the respiration of the leaves can be reused photosynthetically, but this is a minor amount. In addition to numerous structural modifications that conserve water, metabolic adaptations also exist; two of the most important are C4 metabolism and Crassulacean acid metabolism (CAM).
FIGURE 10-25 (A) Light intensity exceeded the light compensation point for these trees just after 6 AM, and photosynthesis increased rapidly; however, by early afternoon, photosynthesis dropped even though light and temperature were adequate. The problem was a lack of water (water stress), and probably stomata had begun to close around noon. (B) On a cloudy day, water stress never became a problem and stomata remained open. Even though there was less total light than in (A), there was more photosynthesis for the day.
C4 Metabolism
An important factor for plants is the amount of water lost for each molecule of carbon dioxide absorbed. This ratio is called the plant’s water use efficiency. Ideally, this ratio is low. Carbon dioxide diffuses into a leaf faster if its concentration in air is higher or if its concentration inside leaf protoplasm is lower. Plants can do nothing about the carbon dioxide level in the external air, and many also have poor control over the protoplasmic concentration. RuBP carboxylase has a low affinity for carbon dioxide; as carbon dioxide concentration drops, enzyme-substrate binding slows. Even while carbon dioxide is still rather abundant in protoplasm, the enzyme is only rarely picking it up. With this relatively high concentration, carbon dioxide diffusion into the leaf is slow, whereas water loss may be high. Under normal conditions, each molecule of RuBP carboxylase fixes only one to twelve carbon dioxide molecules per second. This is extremely slow: Most enzymes catalyze thousands of reactions per second.
RuBP carboxylase occasionally binds to oxygen instead of carbon dioxide, acting as an oxygenase and producing one molecule of 3-phosphoglycerate and one of phosphoglycolate. This latter molecule is transported from the chloroplast to peroxisomes and mitochondria, where some of it is converted to the useful amino acids glycine and serine, but much of the phosphoglycolate is broken down to two molecules of carbon dioxide (FIGURE 10-26). The breakdown is photorespiration, an energy-wasting process. The energy and reducing power used to produce the two reduced carbons of phosphoglycolate are completely lost. Photorespiration is extremely exergonic but is not used to provide power to endergonic reactions. Because phosphoglycolate is toxic, photorespiration protects the plant whenever RuBP carboxylase picks up oxygen; however, it is an expensive defense because up to 30% of all ATP and NADPH produced by the chloroplast can be immediately lost by photorespiration.
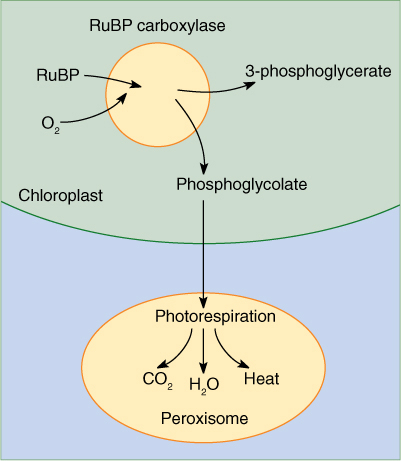
FIGURE 10-26 If RuBP carboxylase puts oxygen onto RuBP, one of the products is phosphoglycolate, which is transported to a peroxisome and broken down during photorespiration. All of the energy that was present in the phosphoglycolate is wasted. Mitochondria are also involved.
Apparently RuBP carboxylase cannot be significantly improved. At the time when RuBP carboxylase originated by evolution, there was no free oxygen at all in the atmosphere, and carbon dioxide concentrations were very high. RuBP carboxylase was well-adapted to that environment. The first type of photosynthesis to evolve began to remove carbon dioxide from the air but it did not produce any oxygen (see BOX 10-2), so conditions were still appropriate for RuBP carboxylase. Photosynthesis that produces oxygen originated in cyanobacteria about 2.8 billion years ago, and ever since, Earth’s atmosphere has slowly changed such that RuBP carboxylase does not function well: Oxygen has increased and carbon dioxide has decreased. Massive volcanic eruptions occasionally increase the concentration of carbon dioxide in the air, but molecule by molecule, RuBP carboxylase pulls it out, altering its environment to its own detriment. RuBP carboxylase has existed for billions of years, and virtually no structural mutations have survived; they must not have produced superior versions of the enzyme.
An alternative is to improve the working conditions of RuBP carboxylase. RuBP carboxylase should be compartmentalized in a site where carbon dioxide concentration is high and oxygen concentration is low. This has evolved in some plant groups and is known as C4 metabolism or C4 photosynthesis. Basically, C4 metabolism is a mechanism by which carbon dioxide is absorbed, transported through, and concentrated in a leaf, whereas oxygen is kept away from RuBP carboxylase.
C4 metabolism occurs in leaves with Kranz anatomy (FIGURE 10-27). In such leaves, mesophyll is not distributed as palisade and spongy parenchyma; rather, each vascular bundle has a prominent chlorophyllous sheath of cells, and around the sheath are mesophyll cells. Mesophyll cells contain the enzyme PEP (phosphoenolpyruvate) carboxylase, which has a very high affinity for carbon dioxide. Unlike RuBP carboxylase, as the carbon dioxide concentration drops lower and lower, PEP carboxylase continues binding to it rapidly and firmly. Carbon dioxide concentrations inside the leaf are kept very low, and carbon dioxide diffuses inward rapidly whenever stomata are open. The ratio of water lost to carbon dioxide absorbed is favorably low. Also, PEP carboxylase has a high specificity for carbon dioxide; it never picks up oxygen. PEP carboxylase is an ideal enzyme except that it does not perform the critical reaction that results in 3-phosphoglycerate. Despite its shortcomings, RuBP carboxylase is still the only enzyme that carries out the necessary reaction.
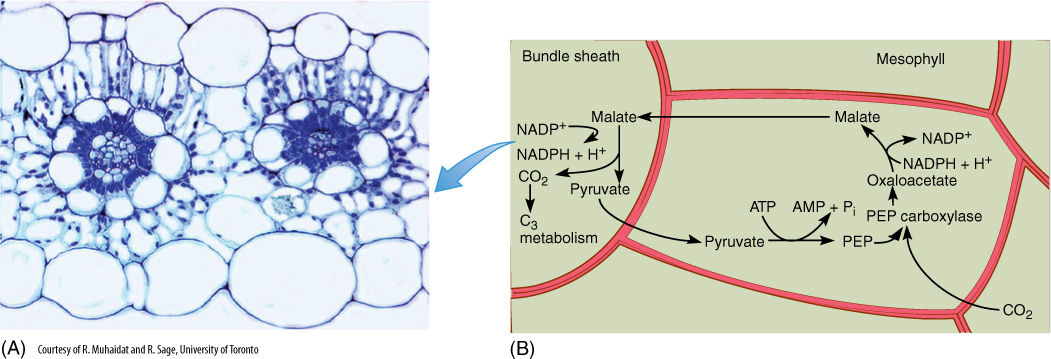
FIGURE 10-27 (A) In leaves with Kranz anatomy, the bundle sheath around all veins is prominent and rich in chloroplasts. The bundle sheath chloroplasts are located as closely as possible to the vascular tissues. The mesophyll cells are arranged in a sheath around the bundle sheath (×250). (B) Knowing the reactions of C4 metabolism is not sufficient for understanding it; only by realizing that some reactions occur in separate compartments does it become logical.
PEP carboxylase adds carbon dioxide to PEP, producing oxaloacetate, which has four carbons, hence the name C4 metabolism. Oxaloacetate is reduced to malate by a molecule of NADPH, and further reactions may occur, depending on the species. Malate from throughout the mesophyll moves into the bundle sheath and breaks down into pyruvate by releasing carbon dioxide (Figure 10-27B). This reaction is powerful enough to drive the formation of a new molecule of NADPH, so, the process results in the transport by malate of both carbon dioxide and reducing power. Because all of the malate from a large volume of mesophyll decarboxylates in the small volume of the bundle sheath, carbon dioxide concentration in the sheath is very high. Typically, the concentration of carbon dioxide in bundle sheath cells is 10 times higher than it is in air. Also, because NADPH is synthesized by this unusual method in bundle sheath cells, the bundle sheath chloroplasts primarily carry out cyclic electron transport, pumping protons and making ATP. Without noncyclic electron transport, there is no breakdown of water or production of oxygen. RuBP carboxylase, located exclusively in the bundle sheath chloroplasts, is in ideal conditions—a high carbon dioxide concentration with low or no oxygen—and carries out efficient C3 stroma reactions with low production of phosphoglycolate. C4 plants have little photorespiration (TABLE 10-6).
Like any other carrier, malate must be shuttled back to its recharge site, the mesophyll. In the bundle sheath it is converted to pyruvate by the release of carbon dioxide; pyruvate moves back to the mesophyll and receives a phosphate group from ATP, which converts it to PEP.
The selective advantage of C4 metabolism depends on the environment. Approximately 50 million years ago, the concentration of carbon dioxide in the air dropped to a critical level and global temperatures cooled. Twenty million years later, Earth entered the Pleistocene Ice Age and ever since has been dry and low on carbon dioxide. Conditions such as these favor the increased water use efficiency of C4 metabolism, and it began to evolve independently many times (perhaps as many as 62 times). At present it occurs in 19 separate plant families. Photorespiration increases with temperature, so it is more of a problem in hot climates. Under warm, dry conditions, C4 metabolism has a strong selective advantage over C3 metabolism: Much less water is lost during carbon dioxide absorption. Also, abundant light is available to generate the extra ATP needed to convert pyruvate to PEP. The ATP used to make PEP means that C4 metabolism is not free; under cool conditions, photorespiration may be slow enough that it loses less energy than C4 metabolism. Also, many cool habitats are also moist, so water conservation by stomatal closure is not as critical. The critical temperature above which C4 metabolism is more advantageous selectively than C3 metabolism varies among species but averages around 25°C: In general, the optimum temperature range for C3 photosynthesis is 20—25°C (68—77°F), and for C4 photosynthesis it is 25—30°C (77—86°F).
TABLE 10-6 Types of Carbon Dioxide Processing
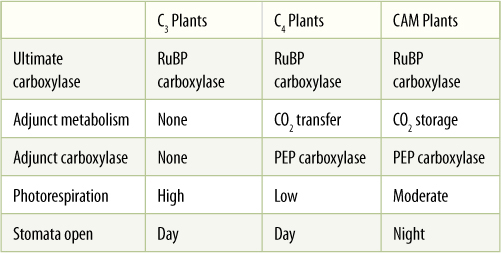
TABLE 10-7 Plant Families Having C4 Species
C4 metabolism and Kranz anatomy have evolved several times; most C4 species are monocots of hot climates such as corn, sugarcane, sorghum, and several other grasses, but a considerable number of eudicots, also from warm, dry regions, are also C4 species (TABLE 10-7).
Crassulacean Acid Metabolism
Crassulacean acid metabolism (CAM) is a second metabolic adaptation that improves conservation of water while permitting photosynthesis. It is so named because it was first discovered in those members of the family Crassulaceae that have succulent leaves (FIGURE 10-28 and TABLE 10-8). The metabolism is almost identical to that in C4 plants: PEP is carboxylated, forming oxaloacetate, which is then reduced to malate or other acids. These acids are not transported but simply accumulate, in effect storing carbon dioxide. This occurs at night. These plants differ from C3 and C4 plants in that their stomata are closed during the hottest periods and open only at night when it is cool. Coolness reduces transpiration, and usually air is calmer at night; therefore, water molecules near stomata are not blown away immediately and may diffuse back into the plant. Opening stomata at night is effective for conserving water, but the lack of light energy creates a problem for photosynthesis: A plant cannot store ATP and NADPH during the day for use at night; they are not stable enough and each cell has too little. Hence, carbon dioxide is stored on acids until daytime, when stomata close and the malate or other acids break down, releasing carbon dioxide for C3 metabolism. The released carbon dioxide cannot escape because stomata close at dawn.

FIGURE 10-28 (A) Sempervivum arachnoideum, a member of the Crassulaceae. Like many desert succulents, it has CAM metabolism as well as other adaptations that conserve water. (B) This barrel cactus (Ferocactus) is a CAM plant. (C) Photosynthetic cells of CAM plants have large vacuoles, which permits the accumulation of C4 acids. These are cortex cells, arranged in palisades, similar to leaf palisade mesophyll (×250).
Crassulacean acid metabolism is not particularly efficient; the total amount of carbon dioxide is so small that it may be entirely used in C3 metabolism after just a few hours of sunlight. Furthermore, RuBP carboxylase is protected from photorespiration only in the morning when internal carbon dioxide levels are high. Later in the day, photorespiration may be high, but the carbon dioxide released by the breakdown of phosphoglycolate is trapped and refixed.
Alternatives
BOX 10-2 Photosynthesis in Bacteria and Cyanobacteria
A process as complex as photosynthesis had to evolve by numerous steps, and many early types of photosynthesis are present in bacteria and cyanobacteria. By examining them, we can understand plant photosynthesis more fully.
Cyanobacterial photosynthesis is the most similar to that of plants. This is not surprising because chloroplasts arose as endosymbiotic cyanobacteria living inside early eukaryotic cells. Cyanobacterial light reactions are almost identical to those in chloroplasts, having chlorophyll a but lacking chlorophyll b. Their accessory pigments are phycobilins, open-chain tetrapyrrole rings (FIGURE B10-2A), which act like carotenoids by absorbing wavelengths that chlorophyll cannot and then transferring the energy to chlorophyll and activating an electron. One class of phycobilins, phycocyanin, absorbs most strongly at approximately 620 to 640 nm and is blue; the other class, phycoerythrin, is red because it absorbs maximally at 550 nm. Both occur bound to proteins, and these aggregate into small nodules (phycobilisomes) visible by electron microscopy. Most other aspects of photosynthesis in cyanobacteria are identical to those of plants; electrons are taken from water, passed through photosystem (PS) II to PS I, and are then used to reduce NADP+ to NADPH. Oxygen is liberated as a waste product, as in plants; therefore, both have oxygenic photosynthesis. Cyanobacteria do not have chloroplasts, but they do have extensive sheets of infolded plasma membrane that contain pigments and electron carriers; the folded membrane forms accumulation spaces for protons and the generation of a chemiosmotic gradient.
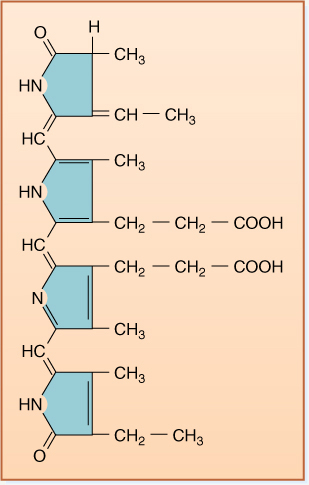
FIGURE B10-2A A typical phycobilin, containing four pyrrole groups—a tetrapyrrole. The two ends of the pyrrole are not attached to each other as they are in chlorophyll.
Purple bacteria and green bacteria do not contain chlorophyll, either a or b, but instead have bacteriochlorophylls (FIGURE B10-2B). Like chlorophyll, these are closed tetrapyrroles with a long tail, but they have certain side groups that the chlorophylls lack. Carotenoid accessory pigments are present, as in chloroplasts. PS I operates, but PS II is not present to put electrons back onto bacteriochlorophyll. Instead, the original electron comes back through a series of carriers, so electron flow is cyclic, similar to that in plant photosynthesis (FIGURE B10-2C; compare with Figure 10-17). One carrier is quinone; thus, protons are pumped across the photosynthetic membranes, creating a strong chemiosmotic gradient that causes ATP to be generated, just as in eukaryotic chloroplasts. Because there is no PS II, oxygen is not formed. This is anoxygenic photosynthesis. The photosynthetic apparatus of purple bacteria consists of extensive arrays of membranes connected to the plasma membrane. In green bacteria, photosynthetic membranes are cylindrical vesicles that occur in clusters surrounded by another “membrane.” The enclosing membrane does not look like a typical bilayered membrane in electron micrographs and is probably different physiologically as well.
Notice that bacterial photosynthesis is basically just light-powered proton pumping; it is extremely simple and not very effective. Bacteriochlorophyll does not capture enough light energy to activate an electron strongly enough for it to move onto NADP+; instead, electrons must cycle repeatedly, gradually building up enough of a chemiosmotic gradient to finally produce NADPH. In contrast, chlorophyll captures enough energy to reduce NADP+ in just two steps. Also, plants and cyanobacteria obtain all of the electrons they need from water, which is always present wherever organisms are alive. Purple and green bacteria recycle electrons repeatedly as they pump protons, but the electrons used to reduce NADP+ must come from other substances, such as sulfur and hydrogen, which are not always reliably present.
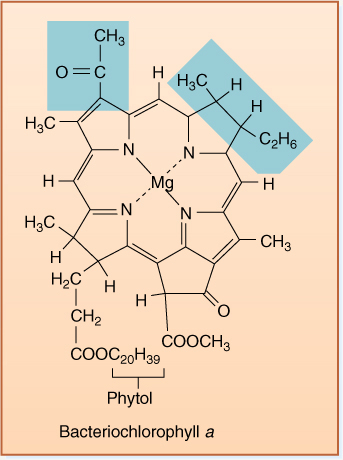
FIGURE B10-2B Bacteriochlorophyll a is similar to chlorophyll a, differing only at the sites indicated by color. Is this similarity analogous, the product of two independent lines of evolution, or homologous, the genes for one pigment having evolved from those for the other? These two molecules are not sufficient evidence to judge, but we are almost certain that all photosynthetic systems are closely related evolutionarily.
Although plant photosynthesis is complex, think about it in terms of being composed of several parts that originated as simple pumps, cyclic electron flows, and chemiosmotic synthetic pathways. Each could originate as a simple metabolic function that then evolved to become more sophisticated. Gradually, one part was added to another, then another, and finally, chlorophyll-based oxygenic photosynthesis resulted.
FIGURE B10-2C Electrons flow cyclically from activated bacteriochlorophyll a, pumping protons with quinone carriers. This establishes a chemiosmotic gradient that generates ATP. Bph = bacteriopheophytin.
TABLE 10-8 Plant Families Having CAM Species
Crassulacean acid metabolism is selectively advantageous in a hot, very dry climate where survival rather than rapid growth is most important. In these habitats, unaided C3 metabolism is so wasteful of water that C3 plants cannot survive. In the most arid regions, C4 plants barely get by, growing in slightly less stressful microhabitats such as near temporary streams and ponds, or growing and flowering quickly during the cool, moist months of winter and spring and then dying in the summer and surviving only as seeds. Under such conditions, CAM plants have a selective advantage; they may not grow luxuriantly, but they do grow and reproduce more successfully than C3 and C4 plants.
Under milder, moister conditions, Crassulacean acid metabolism is not selectively advantageous. Water conservation is less of a benefit, and the limited capacity to absorb and store carbon dioxide is a distinct disadvantage. C3 and C4 plants photosynthesize all day, whereas CAM plants may stop before noon.
Crassulacean acid metabolism has evolved several times and is also present in the cactus family, many orchids, bromeliads, lilies, and euphorbias. All have other metabolic and structural adaptations for water conservation, such as succulent bodies filledwith water-storing parenchyma and covered by a tough epidermis with a thick cuticle and wax layer.
 At the Next Level
At the Next Level
1. Photosynthesis and carbon sequestration. Box 10-1 is an introduction to the interplay between greenhouse gases, photosynthesis, and global warming. An Internet search on this will lead you to many sites, some written by crackpots, so be careful to use your judgment as to whether a site is reliable. One important topic for you to focus on is carbon sequestration: removing carbon dioxide from the atmosphere and locking it away as wood, calcium carbonate seashells, or as carbon dioxide pumped into empty oil wells. Another important topic is whether photosynthesis will automatically speed up as levels of carbon dioxide increase in the air—after all, the current level of carbon dioxide is one of the lowest that has ever occurred—and it could be that plants are now growing more slowly than they ever did in the past. But we are not certain and research is in progress.
2. Photosynthesis and biofuels. Many people suggest that we should cultivate plants such as corn and sugar cane and then ferment the corn and sugar to alcohol to use as a substitute for petroleum. Fuels produced by photosynthesis are biofuels. The problem is that the production of these crops requires gasoline and diesel for tractors to plow, plant, till, harvest, haul, and refine the corn and sugar. If we put in more energy then we get out, we are not only wasting our time, we are making the situation worse. Researching and thinking about biofuels is an important topic by which you can take your thinking to the next level.
3. Chloroplast structure and reactions centers. Our understanding of the biochemistry and biophysics of photosynthesis is expanding rapidly. Advanced textbooks in plant physiology can give you an orderly explanation of photosynthesis, and with the information you have in this chapter, you should have a good foundation. A textbook or review article must be new: Progress is so rapid that a 10-year-old book on photosynthesis is out of date. Light harvesting complexes are especially important.
4. Global warming. Convincing proof exists that global warming is being caused by us, by our burning fossil fuels; it is called anthropogenic global warming. An important step for you to think about is whether global warming is good or bad. We are in an interglacial period of an ice age: Earth has occasionally been colder than it is right now, but more often it has been much warmer and with much more common dioxide in the atmosphere. What will be the effect of warmer temperatures? Some species will go extinct, but new species may come into existence and others will flourish. Remember: The scientific method can provide us with facts about this, but you must use your own ethical standards to decide if it is good or bad.
SUMMARY
1. All physical systems have a tendency to become disordered, to increase in entropy. Living organisms have a high degree of order and regularity, maintained by the input of energy, the ultimate source of which is the sun.
2. All photosynthetic plants are autotrophs, but some parasitic plants and many plant tissues are heterotrophic.
3. When electrons are passed from one atom to another, the recipient becomes reduced, and the donor becomes oxidized. Most of a plant’s raw materials are highly oxidized and must be reduced as they are assimilated.
4. Photosynthetic pigments respond to light quanta that have just enough energy to boost electrons one or two energy levels.
5. Accessory pigments have absorption spectra different from that of chlorophyll a, and they therefore absorb different wavelengths and transfer energy to chlorophyll a.
6. Photosystems I and II work together, transferring electrons from water to NADPH. The electron’s energy is boosted twice, once at P680 and again at P700.
7. The light-dependent reactions result in a chemiosmotic gradient. Driven by concentration differences, protons flow from the thylakoid lumen to the stroma through ATP synthetase channels, powering the phosphorylation of ADP to ATP.
8. Cyclic electron transport permits production of extra ATP without synthesis of NADPH or production of free oxygen. Noncyclic electron transport results in production of both ATP and NADPH, but the amount of ATP is not sufficient for the stroma reactions.
9. All photosynthetic plants use the C3 stroma reactions mediated by RuBP carboxylase: A molecule of RuBP is carboxylated, energized, and reduced, resulting in two molecules of 3-phosphoglyceraldehyde.
10. C4 metabolism is an adjunct to C3 metabolism, not a replacement. PEP carboxylase, which acts as the initial carboxylating enzyme, has a great affinity for carbon dioxide.
11. Crassulacean acid metabolism is similar to C4 metabolism, except that it accumulates and stores carbon dioxide at night while stomata are open and releases it during the day while stomata are closed.
IMPORTANT TERMS
absorption spectrum
action spectrum
anabolic reactions
anoxygenic photosynthesis
antenna complex
ATP synthetase
bacteriochlorophylls
C3 (Calvin/Benson) cycle
C4 metabolism (C4 photosynthesis)
chemiosmotic phosphorylation
chlorophyll a
Crassulacean acid metabolism (CAM)
cyclic electron transport
cytochromes
electron transport chain
entropy
global warming
gluconeogenesis
granum
greenhouse effect
heterotrophs
light compensation point
light-dependent reactions
noncyclic electron transport
oxidation state
oxidative phosphorylation
oxidized compounds
oxygenic photosynthesis
ozone
3-phosphoglyceraldehyde
photoautotrophs
photons
photophosphorylation
photorespiration
photosystem I
photosystem II
pigment
plastocyanin
plastoquinones
quality of sunlight
quantum
reaction center
reduced compounds
reducing power
ribulose-1,5-bisphosphate; RuBP
RuBP carboxylase (RUBISCO)
stroma
stroma reactions
substrate-level phosphorylation
thylakoid lumen
Z scheme
REVIEW QUESTIONS
1. What is the meaning of the word entropy? Does the entropy of a plant increase or decrease while it is alive? After it is dead?
2. Name several examples of photoautotrophs and several of heterotrophs. How do photoautotrophs obtain energy? Can a plant be heterotrophic while a seedling and photoautotrophic when older?
3. ATP is an important chemical involved in many of a plant and animal’s metabolic reactions. Yet any plant has only a small amount of it. Can you explain this? When ATP enters a reaction and forces it to proceed, what is the ATP converted into? What then happens to that molecule?
4. Name the three methods of phosphorylation.
5. What is a reduction reaction? Why does a reduction reaction always occur simultaneously with an oxidation reaction?
6. In organic molecules, we calculate the oxidation state of carbon by assuming that each oxygen has an oxidation state of _____________________. Each hydrogen has an oxidation state of _____________________. Calculate the oxidation state of carbon in each of the following: CO2, CH2O, and malic acid.
7. Two of the following are oxidizing agents and two are reducing agents. Which are which: NAD+, NADP+, NADH, and NADPH?
8. In photosynthesis, what is the ultimate source of electrons? What are the benefits of this molecule in terms of its toxicity and the cost of the plant to obtain it?
9. Describe the absorption spectrum of chlorophyll. Why does it match the action spectrum of photosynthesis?
10. Chlorophyll does not use high-energy quanta. Why not? What would happen to the chlorophyll if it did? It also does not use long wavelength radiation either. Why not?
11. The most common accessory pigments in land plants are chlorophyll ____________________ and the ____________________. Algae that live in deep water have other accessory pigments because only _______________-________________ light penetrates deeply into water.
12. Name the electron carriers that transport electrons from photosystem II to photosystem I. Which ones contain metal atoms, and which do not?
13. When photosystem I produces NADPH, its reaction center P700 chlorophyll a loses electrons. What would happen if photosystem II did not supply new electrons to P700?
14. When electrons are removed from water, protons are liberated. Does this occur in the stroma or inside the thylakoid lumen? Can protons move directly across the membrane? Describe the chemiosmotic mechanism of ATP synthesis in chloroplasts.
15. Is ADP converted to ATP directly by the reaction center chlorophylls? Do the enzymes that synthesize ATP obtain the necessary energy by interacting directly with the reaction center chlorophylls?
16. What chemical is the acceptor of carbon dioxide in the C3 cycle? What enzyme catalyzes the reaction, and what is the product?
17. RuBP carboxylase is by no means an ideal enzyme. Describe some of the problems with its active site and its substrate specificity. If we compare the amino acid sequences of this enzyme from many different species, they are almost identical. What is the significance of this uniformity?
18. Which chemicals are useful for energy storage on a short-term basis? Which are for intermediate term and which are for long term?
19. What is the “quality” of light? How does it differ for plants in deserts, grasslands, and the canopy of a forest versus for plants in the understory? How does it differ for algae that grow near the surface of a lake or ocean versus those that inhabit deep water far below the surface?
20. How is the quantity of light affected by a plant’s location relative to the equator or the poles? On one side of a mountain or the other? On one side of a valley or the other?
21. Imagine a leaf in bright light but an atmosphere with no carbon dioxide. Would RuBP carboxylase be functioning? Would the NADP be in the reduced or oxidized form?
22. Name some of the brightest environments. Describe some protective adaptations that plants may use to shade themselves.
23. An important factor for plants is the amount of water lost for each molecule of carbon dioxide absorbed. How could the plant be harmed if it loses a lot of water for each carbon dioxide molecule, that is, if the ratio is high? Would this be more important for a plant in a rainy habitat or one in a desert?
24. In a C4 plant, where is PEP carboxylase located? Where is RuBP carboxylase located?
25. In a CAM plant, are stomata open during the day or the night? How does this affect the amount of water the plant loses when its stomata are open?
26. As a CAM plant makes and stores acids during the night, how does this affect the plant’s acidity (its pH)? Think about the acidity of your own blood. Do you think it is allowed to vary by any large amount?
27. In habitats where water conservation is not especially necessary, is CAM metabolism more or less advantageous than C3 or C4 metabolism? Why?
28. What is global warming? What is the main gas that causes it? What would happen if the Earth’s atmosphere had a lower concentration of CO2 than it has now? What would happen if it had more?
29. What is the Kyoto Protocol? How many countries have signed it? Name one country that has not signed it. What are two substances burned in the United States (and all other countries) that produce CO2? Which two countries have large populations and may soon surpass the United States in production of greenhouse gases?
Design Credits: Hummingbird: © Tongho58/Moment/Getty; Green plant cells: © ShutterStock, Inc./Nataliya Hora; Purple tulip: © ShutterStock, Inc./Marie C Fields; Dandelion: © ShutterStock, Inc./danielkreissl; Poppy: © ShutterStock, Inc./Saruri; Plant icon: © ShutterStock, Inc./Vector; Digging man icon: © ShutterStock, Inc./Z-art