Botany: An Introduction to Plant Biology - Mauseth, James D. 2017
Plant Physiology and Development
Development and Morphogenesis

Chapter Opener Image: Every leaf on this wall is facing the same direction, each is turned such that it receives maximum sunlight. This is no coincidence; instead, individual leaf blades perceive the direction of the optimum amount of light, then transmit that information to cells in their own petiole causing some to swell, some to shrink such that the blade is rotated and receives optimum illumination. Environmental information has been perceived and transmitted and stimulated a response. Similarly, various parts of a single plant, such as shoots, can generate and send a signal to other parts, such as roots, informing them that more water is needed (usually indicating it is springtime) or less (as leaves abscise in autumn).
OUTLINE
✵ Environmental Complexity
- Light
- Gravity
- Touch
- Temperature
- Water
- Pests and Damage
✵ Responding to Environmental Stimuli
- Tropic Responses
- Nastic Responses
- Morphogenic Responses
- Taxis
✵ Communication Within the Plant
- Perception and Transduction
- Chemical Messengers
- Signal Amplification Cascades
✵ Activation and Inhibition of Shoots by Auxin
- Cell Elongation
- Apical Dominance
- Differentiation of Vascular Tissues
✵ Interactions of Hormones in Shoots
✵ Hormones as Signals of Environmental Factors
- Leaf Abscission
- Tropisms
✵ Flowering
- Ripeness to Flower
- Photoperiodic Induction to Flower
- Endogenous Rhythms and Flowering
- ABC Model of Flower Organization
Box 14-1 Alternatives: Simple Bodies and Simple Development in Algae
Box 14-2 Botany and Beyond: Names of Genes
Box 14-3 Plants and People: Environmental Stimuli and Global Climate Change
LEARNING OBJECTIVES
After reading this chapter, students will be able to:
✵ Discuss cell communication and coordination in plant growth.
✵ Recall the three principles of plant response to the environment.
✵ Summarize the effects of light, gravity, touch, temperature, water, and damage on plant development.
✵ List the four classes of plant responses to environmental stimuli.
✵ Restate how presentation time and threshold influence plant growth.
✵ Recall the names and functions of the most well-studied hormones.
✵ List three potential cell responses to auxin.
✵ Give two examples of how hormones communicate environmental change.
✵ Summarize how differing photoperiods affect plant growth and flowering.
✵ Describe how endogenous rhythms impact plant flowering.
✵ Explain the ABC model of flower organization.
 Did You Know?
Did You Know?
✵ All organisms begin life as a spherical fertilized egg, with differences in genes, gene expression, growth, and morphogenesis producing all the different types of individuals.
✵ During growth and development, various parts of a plant send chemical messages to other parts, such that development is coordinated.
✵ Organisms obtain information from their environment: If grown in continuous light and unchanging temperatures, plants do not develop properly.
✵ Plants grown in microgravity on the International Space Station may bloom and produce seeds, but they often have odd shapes due to lack of gravity.
![]() Concepts
Concepts
All plants begin as a zygote—a single cell—that undergoes enlargement and cell division as it grows and develops into an individual plant. It undergoes morphogenesis (generation of the shape of the plant and its various organs) and differentiation (an increase in complexity as some cells become different from each other). Often the zygote divides into a basal cell and an apical cell: This establishes polarity, the formation of the root/shoot axis. Also, the zygote’s spherical symmetry changes to the embryo’s radial symmetry. Radial symmetry dominates roots and stems, and typically, only leaves, sepals, and other flower parts have bilateral symmetry.
Cell differentiation occurs with the first cell division. The basal cell develops a size, shape, and metabolism that differ from those that the apical cell develops. Later, surface cells differentiate into epidermis cells, including guard cells and trichomes; inner cells differentiate into cortex, phloem, xylem, and pith. Such differentiation requires that each cell be capable of identifying its position relative to other cells and also of informing other cells of the types of developmental and morphogenic changes it is undergoing. For example, cells that will differentiate into epidermis cells must first detect that they are on the plant’s surface. Also, surface cells in roots must develop into epidermis tissue that consists only of pavement cells and root hairs, whereas surface cells of stems of the same plant develop into an epidermis with pavement cells, trichomes, and guard cells; those of petals might develop into pavement cells only, but filled with pigments. To develop properly, a pre-epidermis cell must not only detect that it is a surface cell, but also which organ it is a part of.
Consider also the differentiation of guard cells. They must occur in pairs (due to being sister cells in some species but derived from separate mother cells in other species), and they must coordinate where the stomatal pore forms; it would do no good if each were stimulated to develop as a guard cell but then each tried to form a pore on one of the sides that does not face the other guard cell. Furthermore, pairs of guard cells do not occur at random on stems and leaves, nor are they clustered together: There is a pattern establishment mechanism that informs each cell of its location relative to other epidermis cells. It appears that after one cell begins to differentiate as a guard mother cell, it produces a substance that moves into surrounding cells and inhibits them from also differentiating as guard mother cells. As the inhibitor spreads, it becomes more dilute until it no longer prevents distant cells from acting as guard mother cells. We now know that the inhibitor is a peptide (a very short protein) produced by a gene called EPIDERMAL PATTERNING FACTOR-LIKE (EPFL). The peptide is secreted from guard cell mother cells and diffuses outward into the adjacent young epidermis cells (FIGURE 14-1). There, it interacts with another protein produced by a gene called ERECTA, and the combination of peptide and protein prevents neighboring cells from becoming new guard cells. The peptide cannot diffuse very far, so young epidermis cells that are located far enough away from differentiating guard cells are not inhibited; consequently stomata form with a well-defined spacing between them. Rarely, things go wrong and inhibition is too great (stomata are then too far apart). Botanists have produced a mutation in Arabidopsis thaliana called too many mouths (tmm) in which the pattern establishment mechanism does not work correctly and stomata are formed side-by-side with few or no pavement cells between guard cells. There are additional control mechanisms as well. For example, if subsurface cells are differentiating into fibers, they inhibit adjacent surface cells from undergoing guard cell morphogenesis.
Inner cells of young regions of shoots also must detect their position and differentiate accordingly. Typically, the first cells below the apical meristem to become visibly differentiated are sieve tube elements of protophloem. They are located part way between the epidermis and the center of the stem, and each develops several micrometers away from other newly forming sieve tube elements: Like guard mother cells, each region that begins to differentiate as phloem must inhibit adjacent regions from also becoming phloem, thus establishing a pattern of one ring of bundles in most seed plants, numerous bundles in monocots. Vascular bundles always contain a strand of xylem located just interior to the phloem; thus, the first sieve tube elements may be capable of stimulating another pattern: xylem formation. This pattern, however, does not affect all cells equally; only cells located between the phloem and pith are stimulated to undergo xylogenesis, whereas cells located between the phloem and epidermis are unresponsive.
Whereas guard cell morphogenesis requires coordinated differentiation of two cells, morphogenesis of vessels requires coordination of thousands of cells. As each new vessel element differentiates from a young subapical parenchyma cell, it must recruit the next cell above itself to also develop into a vessel element, thus extending the vessel upward. And each element must make perforations aligned with the perforation of the element below it (which is slightly older and more mature) and the one above it (which is slightly younger). Each newly recruited cell must then recruit the next cell to be part of the vessel and so on. Differentiation of sieve tubes requires not only the progressive recruitment of sieve tube members upward in the shoot’s newly developing vascular bundle, but also the coordinated differentiation of a companion cell for each sieve tube member.
In most cases, we assume that cells inhibit or promote activities in nearby cells by secreting chemicals, but it may be that the enlargement of some cells—or the lack of enlargement—causes physical stresses in tissues. Compression or stretching may also be important in establishing patterns of cells and tissues.
Development and morphogenesis also occur at the level of organs and the whole plant (FIGURE 14-2). While still small, a seed plant embryo establishes a shoot apical meristem and a root apical meristem. The shoot apical meristem generates new cells for both itself and for the elongating stem. It also establishes leaf primordia in characteristic phyllotactic patterns, and leaf primordia in turn establish axillary buds. Although each axillary bud develops its own apical meristem, a chemical messenger (a hormone) from the stem’s apical meristem inhibits most of the stem’s axillary buds, keeping them quiescent and thus controlling the pattern of branching and the shape of the plant. Vascular bundles differentiate upward toward the shoot apex and branch at the proper place such that leaf and bud traces interconnect the stem’s vascular system with those of leaves, flowers, and branches.
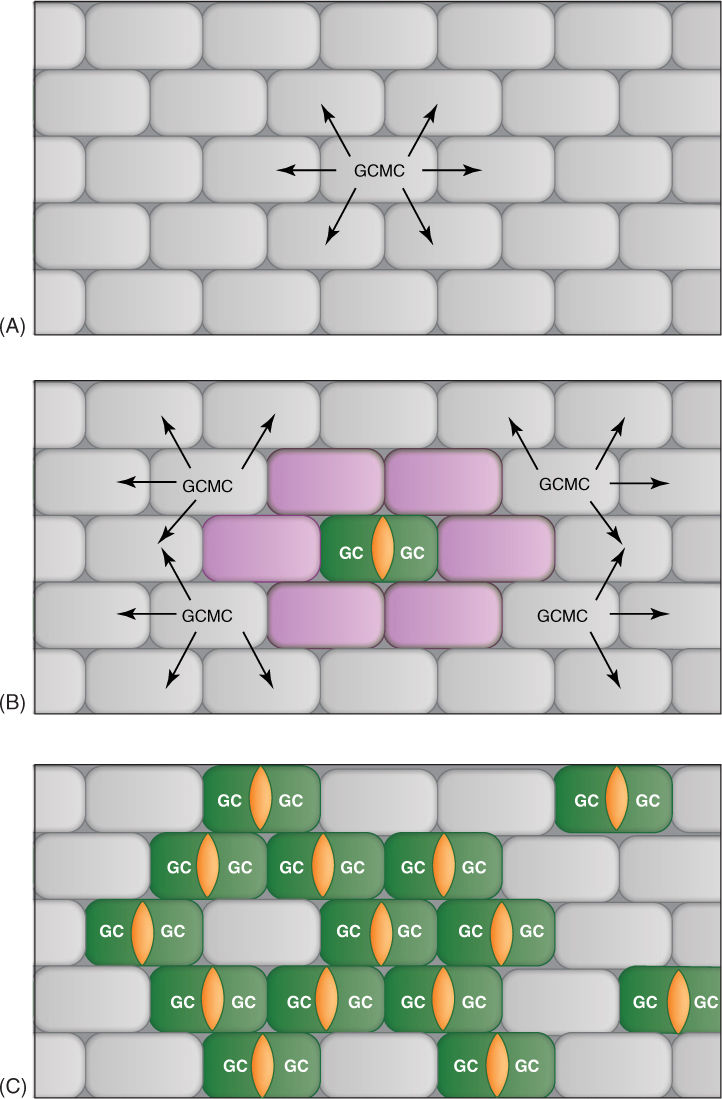
FIGURE 14-1 (A) A cell has become a guard cell mother cell (GCMC) and is exporting the inhibiting peptide EPFL. The peptide enters adjacent cells and reacts with a protein produced by another gene, ERECTA. The combination of peptide and protein [shown in purple in (B)] inhibits these cells from becoming guard cell mother cells, but cells located father away received no peptide and thus are not inhibited. Four are shown as guard cell mother cells that are now inhibiting cells adjacent to themselves. (C) Rarely, inhibition fails and the proper pattern does not develop; here, too many cells develop into guard cells. This mutant gene form has been named too many mouths.
GC = Guard Cell
GCMC = Guard Cell Mother Cell
Orange = developing stomatal pore

FIGURE 14-2 A seed has just germinated, and the microscopic shoot apical meristem is probably just starting to make its first leaf primordia and the first cells of the nodes and internodes. At this point, it is impossible to tell what species this is as many eudicot seedlings look just like this. But as new cells are formed, each differentiates in a particular way, guided by its genes and hormones such that the leaves have characteristic features, as do the nodes, internodes, axillary buds, and later on the flowers and fruits. Ultimately, the actions of all the genes guide the development of a plant body that is easy to recognize as being a particular species. This is the result of differential gene expression, as explained in this chapter.
Plant bodies tend to be much simpler than those of animals, but even so, plant growth and development are not trivial. A great deal of information is required to establish polarity and symmetry, then patterns of cells, tissues, and organs. And the differentiation and morphogenesis that converts each pattern into actual cells, tissues, and organs require even more information. Virtually all information needed for plant development is stored as genes located in nuclei, plastids, and mitochondria. As an embryo develops and later as a seed germinates, it develops into a complete plant relying entirely on its own self-contained information and the intercommunication between its parts.
But plants live in habitats with environmental factors such as light, gravity, water, temperature, seasons, and other organisms. All plants have evolved to have the capacity to detect at least some environmental factors and to alter their development and morphogenesis in adaptive ways. This is an important concept: Plants respond to certain environmental cues in ways that make the plants more likely to survive and reproduce successfully in the habitats they find themselves in. For example, all living stem and leaf epidermis cells synthesize some cuticle: If they are alive, they make cutin. But many plants also can detect environmental moisture and respond adaptively by producing a thicker, more effective cuticle if conditions are dry and a thinner, less expensive cuticle if it is humid. Similarly, seeds germinate and grow if given water and warm temperatures, but if buried under several centimeters of soil when they germinate, they detect gravity and respond adaptively: Shoots grow upward, eventually reaching light, whereas roots grow downward into deep soil that may be more moist. Gravity is not necessary for growth, but it acts as a source of information about where the seedling is located with respect to light and moist soil.
In these and many other cases, the environment provides information that allows plants to alter aspects of development and morphogenesis that would occur anyway. In other cases, environmental cues are needed for the plant to alter its development radically and to complete its life cycle. Many plants detect season (spring, summer, and autumn) by measuring the length of nights, and this stimulates the adaptive response of producing flowers at the right time of year or restricting growth and initiating preparations for winter. Without this environmental information, the plants would continue to grow vegetatively without ever forming flowers or without producing cold-resistant dormant buds, and this would be disastrous for the plant. Note that here too all information needed to actually produce flowers or cold resistance is present in the plant’s genes—it is just that the plant itself cannot activate those genes until triggered by environmental signals.
It is important to distinguish between adaptive responses and mere consequences of environmental stress. When fire kills a plant, the plant’s death is a result of overly adverse conditions; it is not adaptive to the plant. Similarly, a late frost in spring may cause developing leaves to become malformed. The resulting shape is not adaptive; it is merely that frost disturbed leaf morphogenesis destructively.
This chapter describes several environmental cues that plants detect, the chemical signals used by plant parts to communicate with each other, and several widespread types of development and morphogenesis. The following chapter discusses genes and the mechanisms by which their information is accessed and used.
![]() Environmental Complexity
Environmental Complexity
If all plants were extremely small and lived in completely uniform, nonvarying environments, most would probably be simple and would experience little selective pressure for the evolution of complex shapes, tissues, organs, and metabolism. The most uniform, constant conditions occur in oceans and large lakes, where water buffers rapid changes in temperature, acidity, oxygen concentration, and other factors. Under such stable conditions, small organisms such as algae, protozoans, and sponges are simple. But most organisms exist in a heterogeneous environment: Gravity comes from only one direction. The sun is either to the side or overhead but never below. Temperatures are lower on the shady side of a plant. Moisture depends on depth below or height above the soil surface. This mosaic of conditions changes over minutes, days, seasons, or longer periods of time. It is selectively advantageous for plants to sense these differences and changes and to respond to them.
Most plants are so large that their bodies exist in several different microenvironments. Consider a small tree: Its roots are in soil, which is usually moister, cooler, and darker than air; the highest branches are in open air, exposed to full sunlight and the full force of wind, storms, rain, and snow. The trunk base and lower branches are in an intermediate environment—less stable than soil and less variable and severe than open air. A vertical tree trunk is oriented to best resist gravitational attraction (its own weight), whereas horizontal branches are highly stressed unilaterally by gravity.
In springtime, the shoot can become warm enough for active metabolism even though the soil remains cold or frozen. The plant parts must communicate with each other, or shoot buds would become active and expand before roots were capable of transporting water to new leaves. In autumn, increasing night length and declining air temperatures signal impending winter and the need for dormancy; roots are informed about changing seasons by chemical signals from the shoot.
The need for intercommunication and coordination also exists within a limited region of the body. For example, leaf parts act in a coordinated fashion during development such that the petiole has enough xylem and phloem to facilitate the transport needs of the blade. Too little conductive tissue causes the blade to suffer water stress or an inability to export sugars; too much conductive tissue is a waste of energy and material. Even on the intracellular level, organelles must communicate with each other because their metabolisms are interrelated.
All levels of communication have in common a basic mechanism. Information about the environment or the metabolic status of the organ must be perceived. The plant must sense environmental cues such as changes in temperature, moisture, or day length, or the nucleus must receive chemical signals if conditions in the surrounding cytoplasm change. Next, information must be transduced, or changed to a form that can be either acted upon or transported. Finally, there must be a response: The plant must enter dormancy, produce flowers, change the type of leaf production, and so on (FIGURE 14-3). If any of these steps is missing, the plant cannot respond to the environment. These principles apply to all organisms, and of course, higher animals have the most elaborate mechanisms. Their sophisticated sense organs for sight, taste, touch, hearing, and smell perceive external conditions. These sense organs transduce the perceived information to a transportable form such as nerve impulses or hormones that are secreted into the blood stream. Animal responses are also typically highly elaborate, involving precisely controlled movement and increased activity of organs.
Sensory systems and response mechanisms of plants tend to be simpler, and their signal transport is usually slow, involving movement through cortex parenchyma or phloem. This does not mean that animal systems are superior to those of plants. “Superior” has no meaning. The proper question is this: “Which is more adaptive, which is more advantageous selectively?” In terms of evolution and natural selection, animal systems are successful adaptations for organisms that must detect food or predators and must move in order to capture that food or avoid being captured. Any animal with a plant-type mechanism of perception, transduction, and response would soon starve or be eaten.
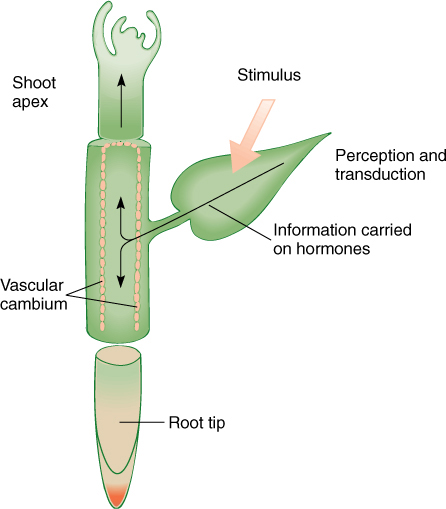
FIGURE 14-3 The signal for dormancy and preparation for winter is short days (long nights) in autumn, perceived by leaves. The chemical messenger transported from the leaves causes the shoot tip to produce bud scales instead of leaves; the vascular cambium fills with many small vacuoles and stops mitosis and cytokinesis. Young xylem mother cells differentiate to a predetermined stopping point and then become dormant; roots slow their growth greatly but usually do not stop completely. Roots and the cambial region cannot perceive the approach of winter themselves but depend on leaves as sites of perception. Although the shoot apices could perceive it themselves, the entire plant is integrated as a whole by being cued by the leaves.
On the other hand, rapid response mechanisms would not be adaptive in plants. They are extremely expensive to build and maintain and are unnecessary because most environmental conditions important to plants change only slowly. Eyes, nerves, muscles, or adrenal glands are not needed by plants for absorption of carbon dioxide, water, minerals, or light or for perception of autumn and preparation for winter dormancy. For sexual reproduction, animal-pollinated plants do need sophisticated perception and response mechanisms, but basically, they simply “rent” those of their pollinators, paying with nectar or other rewards. Finally, the very sophistication of animal sensory/response systems makes animals more vulnerable, more easily injured, and more dependent on avoiding dangers such as fires, floods, freezes, and predators. Plants are typically much more resilient than animals, being able to survive burning because of thick bark or resprouting from rhizomes and bulbs; many withstand flood by being tough or flexible. Predators can consume most of a plant’s leaves, wood, or roots without actually killing the plant.
Light
Besides energy for photosynthesis, light also provides two important types of information about the environment: (1) the direction or, more precisely, the gradient of light. This allows a plant to grow or to orient its leaves toward a region of bright light, which increases the light available for photosynthesis. (2) The duration of light (length of the day) provides information about the time of year. Air temperature is unsuitable because cool autumn temperatures may be followed so quickly by severe cold that plants do not have enough time to become dormant. But day length is an infallible indicator of season.
Gravity
It is selectively advantageous for many plants to orient themselves or their parts with respect to the direction of gravity. In some cases, gravity itself is important because it causes weight stress. A vertical stem supports more weight than does a similar stem growing at random. Whenever a plant is bent or tilted because of flooding or the slipping of a hillside, the plant must change its growth back to an upright direction; if it continued to grow at an angle, it would need to produce many more fibers to support its weight. In some situations, the direction of gravity is instead a guide to other important factors. Roots that grow downward are more likely to encounter water and minerals. Shoots that grow upward grow above other plants and encounter better conditions for photosynthesis, pollination, and seed distribution. Normally, shoots do this by growing toward the brightest light, the open sky, but shoots of seeds that germinate deep in the soil must determine which way is up while in the dark. Direction of gravity is their only reliable guide. Most bilaterally symmetrical flowers must be aligned with the body symmetry and flight pattern of their pollinator. Such flowers must be bilaterally symmetrical vertically and project horizontally because insects and birds do not fly upside down or sideways. The flower must orient itself along the same environmental gradient that the pollinator uses—the gravitational gradient of up versus down (FIGURE 14-4A).

FIGURE 14-4 (A) These Penstemon flowers have detected and responded to gravity in two ways. First, they all project horizontally, not at random and not toward the sun. Second, they are bilaterally symmetrical (their right side is a mirror image of their left side), and the plane of symmetry is vertical. Thus, flower symmetry matches the symmetry of bees, and flower projection allows bees to land easily. (B) Weight is a source of information to the plant about the amount of collenchyma or sclerenchyma needed to counteract the gravitational attraction on the plant. Initially, the flower stalk was strong enough only to support the weight of a flower; by sensing and responding to gravity, it is now strong enough to support the weight of an apple.
Although gravity does not change with time, the force it exerts on a particular organ does change as the weight supported by the organ changes. The pedicel of an apple flower supports almost no weight, but the same pedicel must later support the weight of a fully grown fruit (FIGURE 14-4B). The extra fibers are not produced until needed. Similarly, a young branch must be strong enough to support a small amount of weight, whereas a larger branch must support more.
Touch
Although plants do not move around like animals, their parts frequently grow against objects and respond to this contact. Certain types of contact are detrimental, for example, when a root grows against a stone or a branch rubs against another branch. In these cases, a thick bark is adaptive as a protective layer, and it is produced only where needed, only in the area being touched. Other types of contact are beneficial: After a tendril touches an object, it grows around the object and uses it as a support. When a fly touches sensitive trigger hairs on a Venus’ flytrap, the trap closes, catching and holding the insect during digestion. In some cases, the contact is between two growing primordia and is a normal developmental feature. Many flowers that have fused petals or carpels start with separate primordia that grow together and fuse, acting as a single unit during development (FIGURE 14-5). In each case, the physical action of touching is similar, but each organ responds in a distinct way that is adaptive for the plant; the response of each would be inappropriate if it occurred in the others.
Temperature
Temperature fluctuates in a predictable pattern on both a daily and a yearly basis. Changing temperatures can induce many specific types of plant development. Although most plants appear to be quiescent and virtually lifeless in winter, a considerable amount of critically important metabolism is occurring. This metabolism usually does not proceed at temperatures above 1°C to 7°C.
Cold temperatures are required for the normal flowering of biennial and many perennial plants. Species of perennial trees that are adapted to habitats with cold winters, such as apples, typically require near-freezing temperatures to break the dormancy of their flower buds, which they had formed in the previous summer. If the trees are grown in areas with warm winters, they form flowers that never open. In contrast, biennial plants spend their first year in a vegetative phase and cannot be induced to form flowers. Their first cold winter vernalizes them, it causes them to switch to a state in which they can sense and respond to a stimulus that induces flower formation (vernalization; FIGURE 14-6). In the year after the vernalizing winter, the plants resume vegetative growth, but when exposed to the proper stimulus (usually the short nights of early summer), they respond by producing flower buds. In their first year (prevernalization), summer has no effect; in their second year (postvernalization), it induces flowering. If never vernalized, they never produce flowers.
Low temperatures are required to induce deep dormancy in temperate trees and shrubs. Short days of autumn induce plants to initiate preparations for winter and enter a mild state of dormancy, but the most resistant stages are not entered until the plant actually experiences a week or two of cool temperatures. In contrast, cool temperatures are required to break the dormancy of many seeds, allowing them to germinate when their habitat becomes warm and moist.
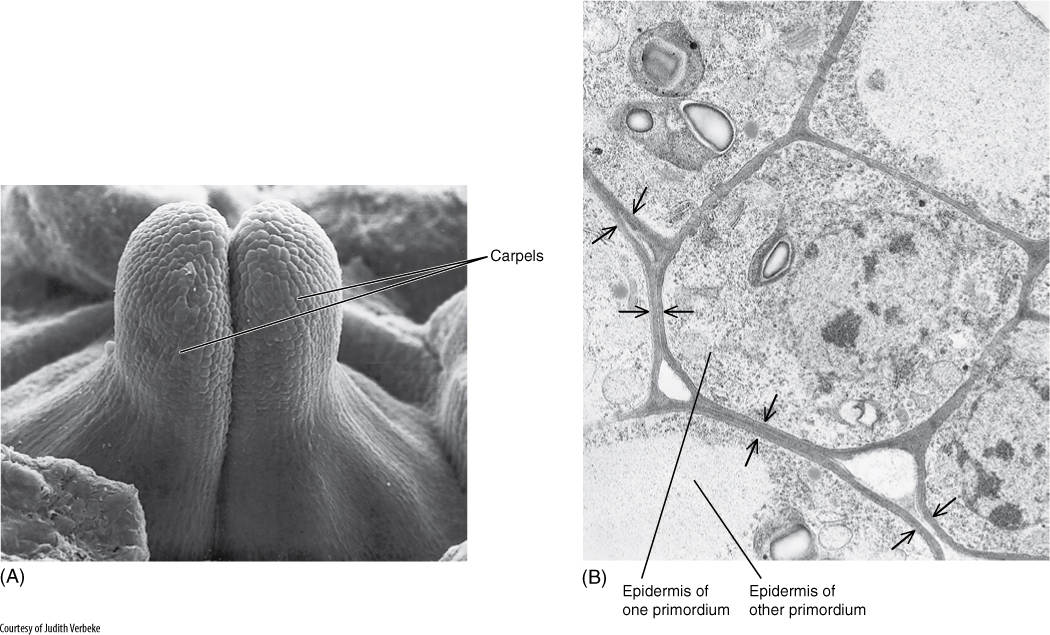
FIGURE 14-5 (A) The carpels of Catharanthus roseus are initiated separately and consist of protoderm and ground meristem, but they soon crowd into each other and fuse into one syncarpelous gynoecium (×70). (B) At the points of contact (arrows), the protoderm changes into mesophyll rather than epidermis (×6,000).

FIGURE 14-6 (A) In its first year a biennial plant has only a very short stem, with all its leaves attached close together, and is unable to flower. (B) In its second year a biennial plant produces a large group of flowers, almost always on a tall stalk. After the fruits and seeds mature, the entire plant dies. This is mullein, Verbascum thapsus.
Water
Although water is an absolute prerequisite for life, its presence probably does not act like a signal in the way other factors do. If enough water is available, plants grow; if not, plants wilt and perhaps even die. Although roots often appear to grow toward water, they actually grow in all directions, and those which, by chance, grow toward water grow more rapidly because they are in a favorable environment. Roots that grow away from water grow slowly, but only because they enter an environment too dry to permit growth. Roots do not turn and grow toward water in the way they turn and grow toward gravity.
Water scarcity triggers specific adaptive responses. One of the first is production of the hormone abscisic acid, which causes guard cells to lose potassium and close stomatal pores. This occurs in most plants even while cells have enough water to carry on basic metabolism. If water stress continues or becomes more severe, new responses are triggered that may inhibit production of new leaves, increase the cuticle on existing leaves, or even initiate abscission of leaves.
Pests and Damage
Plants must detect when they are being attacked by pests or being damaged physically. Pests such as bacteria, fungi, and insects attack plants chemically with toxins and digestive enzymes. Plants also suffer physical damage when larger animals trample them or chew into leaves and twigs. Plants have at least two mechanisms that perceive such damage. First, purely physical damage breaks a plant’s own cell wall molecules, producing fragments of cellulose and hemicellulose. Although cell walls might be physically broken in many ways, most fragments will have certain physical and chemical patterns, such as bits of cellulose molecules protruding from the edges with portions of hemicellulose attached. These are called damage-associated molecular patterns (frequently just called DAMPs), and when the fragments diffuse inward to a cell’s plasma membrane, detector proteins bind to them because the proteins have shapes that match the bits and pieces of cellulose and hemicellulose. Such proteins are called pattern recognition receptor proteins. Once a pattern recognition receptor protein binds to a cell wall fragment, it initiates a signal inside the cytoplasm, informing the cell that its wall has been damaged.
Second, pathogenic fungi and bacteria attack plants with digestive enzymes such that the microbial cells can penetrate the cell wall. Such an attack often produces damage-associated molecular patterns that alert the cell, but just as importantly, the microbes have their own chemicals that a plant can perceive. Fungus cell walls contain chitin, and fragments of chitin have distinctive patterns known as microbe-associated molecular patterns (MAMPs). The plant cell’s plasma membrane has pattern recognition receptor proteins whose shapes match that of chitin, and which inform the plant a fungus is attacking whenever the protein binds a chitin fragment. Fragments of chitin are small, so they diffuse toward the plant cell’s plasma membrane more rapidly than the fungus can grow there. Similarly, many plants have pattern recognition receptor proteins that recognize bits of bacterial flagella. The evolution of pattern recognition receptor proteins as part of a plant’s self-defense mechanisms has been possible because of the ancient origins of chitin, flagella, cellulose, and hemicellulose. These have been reliable indicators of trouble that have not changed for hundreds of millions of years.
![]() Responding to Environmental Stimuli
Responding to Environmental Stimuli
Plant responses to the diverse types of information present in the environment can be grouped into four simple classes (TABLE 14-1).
Tropic Responses
A tropic response is a growth response oriented with regard to the stimulus. For example, growth toward a bright light is a phototropic response (FIGURE 14-7 and TABLE 14-2). A positive tropism is growth toward the stimulus; a negative one is growth away from the stimulus, and plagiotropism is growth at an angle (FIGURE 14-8). Most tap roots are positively gravitropic, growing downward in response to gravity, whereas shoots are negatively gravitropic, growing upward in response to gravity. Branches and secondary roots grow horizontally or at an angle, plagiogravitropically.
When touch is the stimulus, the response is thigmotropism. Positive thigmotropism occurs when a tendril touches an object and, by growing toward it, wraps around it. Pea tendrils are extremely sensitive to touch: After a brief rubbing with a light-weight thread, tendrils gradually coil toward the stimulated side, even if a physical object is no longer present.
Pollen tubes of flowering plants are suspected of displaying positive chemotropism, growing along the style to the ovary by following a gradient of chemical released from the ovule, probably from the synergids. However, this is still uncertain: The chemical responsible is not known, and pollen tubes would have to be sensitive to extremely slight variations in its concentration.
Tropic responses often involve a change in direction of growth. Tendrils grow in a slow spiral, called circumnutation, until they touch an object, and then they change to oriented growth. Changing direction involves differential growth: One side of the tendril must grow more than the other. This can occur by (1) growth on the contact side slowing, (2) growth on the opposite side accelerating, or (3) both. After a tendril starts differential growth, it continues growing differentially until it is mature a few hours or days later. In contrast, if a plant is tilted, its shoots begin differential growth until they are again oriented properly, and then they grow straight ahead with equal growth on all sided, perhaps for years.
TABLE 14-1 Types of Plant Responses to Stimuli


FIGURE 14-7 (A) Construction of porch steps trapped these plants, but by growing toward light (positive phototropism), their shoot tips found spaces between boards and emerged from a suboptimal dark environment into a more suitable sunny environment. (B) Sunflowers (Helianthus annuus) have an unusual type of phototropism: Their “flower” (it is really a set of many small flowers) follows the sun all day long, so they turn from east to west every day for several weeks until finally the stalk becomes too tough and woody to move. Notice that every sunflower is facing the same direction, almost directly toward the sun.

FIGURE 14-8 The central shoot of this tree grows vertically, probably due to both positive phototropism and negative gravitropism, but the branches grow at an angle to vertical, plagiotropically.
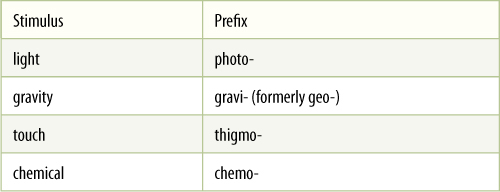
TABLE 14-2 Prefixes for Stimuli
Nastic Responses
A nastic response is a stereotyped nongrowth response that is not oriented with regard to the stimulus. For example, the trap-leaf of a Venus’ flytrap has six large, sensitive trichomes. If a fly or other insect touches any two of these, the trap closes. It does not matter if the fly was moving north or south or up or down, the trap always closes in the same manner in this thigmonastic response. Furthermore, the trap does not grow shut; it closes as motor cells on the midrib upper side suddenly lose turgor. “Positive” and “negative” are not used because the response is not oriented with regard to the stimulus.
Many pollinators are active only at night or during the day, and the flowers they pollinate are open only at the appropriate time. Sepals and petals spread open when the sun rises in diurnal species (active during daylight) and as it sets in nocturnal ones (active at night). Although presence of the pollinator is the critical factor for pollination, the cue that stimulates flower opening is the presence or absence of light, not the presence or absence of pollinators. The opening and closing always happen in the same manner, even if light is given artificially from the west, north, or south, above, or below. This response is photonastic.
Nastic movements are based on changes of turgor pressure rather than growth; thus, movements can be repeated. This occurs in many leaves that undergo sleep movements with the blade elevated in the day, lowered at night. Such leaves often have an enlarged area, a pulvinus (plural, pulvini) at their petiole base, and it contains motor cells that cycle between being turgid and flaccid.
Morphogenic Responses
A morphogenic response, sometimes called a morphogenetic response, causes a change in the “quality” of the plant; that is, a fundamental change occurs in the metabolism of a tissue or even the entire plant. Because day length is such an excellent indicator of season, photomorphogenic responses are numerous: the induction to form flowers (which later open photonastically; FIGURE 14-9), the induction of dormant seeds to germinate (FIGURE 14-10), and the induction of buds to become dormant. An example of a gravimorphogenic response is the formation of fibrous wood when a stem or branch is tilted and becomes stressed by gravity. Thigmomorphogenic responses include formation of extra bark where branches rub against an object and formation of a suture when petal or carpel primordia grow against each other (see Figure 14-5).

FIGURE 14-9 Conversion from the vegetative to the floral condition is a common photomorphogenic response. Day length is controlled in these commercial greenhouses to ensure that all the poinsettias bloom simultaneously at Christmas. The growers could just as easily make them bloom on the 4th of July by controlling day and night length.
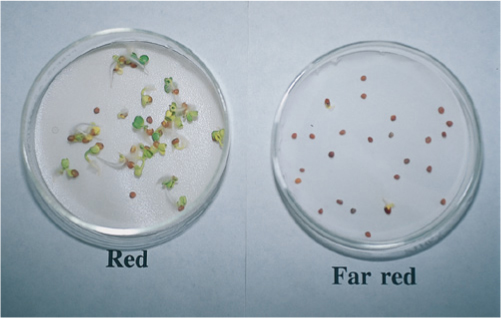
FIGURE 14-10 In these seeds and many others, a two-part mechanism ends dormancy: Cold winter temperatures and rain destroy or wash out an inhibitory chemical, and then light triggers germination. The light-dependent mechanism ensures that the seeds do not germinate while deeply buried under leaf litter and soil. Red light induces germination, but far-red light (infrared) blocks germination.
Taxis
Taxis is a response in which a cell swims toward (positive taxis) or away from (negative taxis) a stimulus. Even in plants like mosses, ferns, cycads, and maidenhair tree (Ginkgo), sperm cells swim to egg cells by following a chemical gradient (chemotaxis). In algae, chemotaxis is similarly important for reproduction, and in many species, phototaxis allows them to swim toward light for photosynthesis or away from light that is too intense.
![]() Communication Within the Plant
Communication Within the Plant
Perception and Transduction
Many, possibly most, responses occur in tissues or organs different from those that sense the stimuli. The site of perception is not the site of response, so a form of communication must exist. In plants, most sites of perception and response are not specialized for those functions but seem to be rather ordinary cells. Day length is probably perceived by all living leaf cells; no specialized region of cells has been discovered. Low temperatures for vernalization appear to be detected by buds, which do not contain a particular group of cells specialized just for temperature perception. In root caps, certain cells called statocytes do have large starch granules, statoliths, that sink in response to gravity; statoliths are too dense to float in cytoplasm and always settle to the bottom of the cell, thereby distinguishing “down” from “up” (FIGURE 14-11). This is our best example of a set of specialized perceptive cells. The trigger hairs on Venus’ flytrap leaves are also a discrete perceptive mechanism, but it is not known which cells within the hairs are responsible.
The site of perception is tentatively assumed to be the site of transduction, where the stimulus is converted into a form that can be transmitted and can trigger a reaction at a response site (see Figure 14-3). Transduction is still a complete mystery in almost all plant responses; we do not know how changes in temperature, light, weight, or humidity are converted into chemical signals.
Two factors are important in perception and transduction: presentation time and threshold. Presentation time is the length of time the stimulus must be present for the perceptive cells to react and complete transduction. Presentation time for root gravitropism is easy to understand: A root must lie on its side long enough for statoliths to sink to the new bottom of the cell. If the root is returned to vertical before they can settle, no perception occurs. In many tropic responses, only a brief touch or unilateral lighting is sufficient to cause curvature; presentation times are often only a few seconds.
After the stimulus has acted long enough to fulfill the presentation time, a response occurs even if the stimulus is removed. For example, the vernalization of many biennial plants has a presentation time of only one or a few days; after this, the plants still flower at the proper time even if kept in warm, nonvernalizing conditions. Tendrils of peas do not bend thigmotropically in the dark, but if they are rubbed for several seconds—their presentation time—in the dark, they bend when placed in light even though they are no longer being touched.
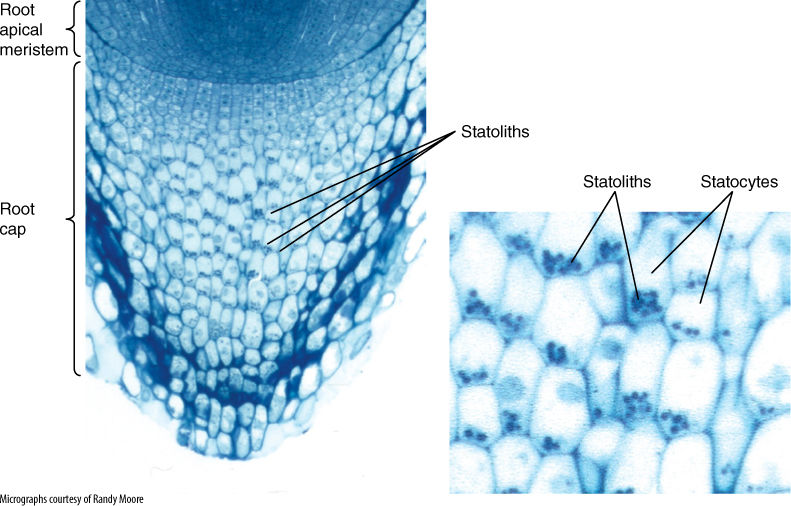
FIGURE 14-11 Cells located centrally in this root cap are statocytes, and their starch grains are statoliths. Regardless of vertical or horizontal position, the statoliths are located at the gravitationally lower side. It is necessary to distinguish between the gravitational and the morphological bottom in gravity-sensing systems.
Threshold refers to the level of stimulus that must be present during the presentation time to cause perception and transduction. In phototropism, plants are extremely sensitive to very dim unilateral light if they are in an otherwise dark environment; the threshold for curvature is low. In bright conditions, the threshold is higher, and the unilateral light must be much stronger to trigger curvature. In Venus’ fly-traps, the threshold for stimulating leaf closure is moderate: The trigger hairs must be firmly bent. This is advantageous in preventing wind or rain from triggering trap closure; the moderate threshold almost guarantees that the trap contains an insect every time it closes (FIGURE 14-12).
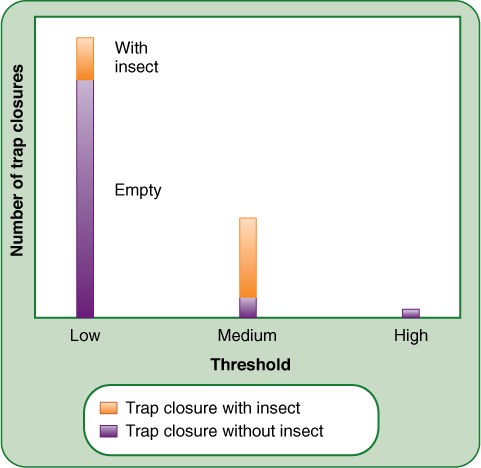
FIGURE 14-12 Threshold must be appropriate to the amount of change a stimulus can cause. If Venus’ flytrap hairs had an extremely low (sensitive) threshold, they would be capable of detecting and catching every insect, but wind and rain would cause so many useless closings that the leaf would be inefficient because it would miss insects whenever it was closed unnecessarily. With a medium threshold, it captures more insects because it is open and ready much of the time. Small insects may escape because they cannot bend the trigger hairs enough to meet the threshold. If the threshold were too high, no insects would be caught because none could bend the hairs. The trap would close only when larger animals brushed against it, but these animals are too big to be enclosed in the trap; thus, all closures would be unproductive. We can hypothesize that natural selection results in a threshold appropriate for the most abundant size of insects.
Related to threshold is the level of response relative to the level of stimulation; the alternatives are all-or-none responses and dosage-dependent responses. In an all-or-none response, after the threshold and presentation time requirements are met, the stimulus is no longer important; the response is now completely internal. Individuals respond identically whether they received strong or weak stimuli, regardless of whether the stimulus was brief or long lasting. For example, many species are induced to flower by environmental conditions; after the minimal threshold and presentation time requirements are met, the plants flower fully, limited only by their general health, vigor, and nutrient reserves. Until they receive the proper stimulus, they produce no flowers; their flowering is all or none. Examples are poinsettia, chrysanthemum, Hibiscus syriacus, and oats.
In dosage-dependent responses, the amount or duration of the stimulus affects the amount or duration of the response. In species of this type, individuals that receive only minimum stimulation flower poorly, even if the plant is quite healthy. Those that receive longer or stronger stimulation produce more flowers. Examples are turnip, marijuana (Cannabis sativa), and some varieties of cotton and potato.
Chemical Messengers
Almost all plant communication is by a slow mechanism: transport of hormones through the plant. Hormones are organic chemicals produced in one part of a plant and then transported to other parts, where they initiate a response. A critical aspect is that hormones act at very low concentrations. Hormones are synthesized or stored in regions of transduction and are released for transport through either phloem or mesophyll and cortex cells when the appropriate stimulus occurs. At the site of response, hormones bind to receptor molecules, usually located in the plasma membrane, and thereby trigger a response. Hormones appear to be released into general circulation and are not carried specifically to the target. Many regions that are not target regions are exposed to the hormone but do not respond because they do not have the proper receptor molecules. In some instances, a plant hormone acts directly on the cells that produce it.
At one time, plant hormones were believed to carry in their structure much of the information necessary for the response. We now know that plant hormones are quite simple in structure. The receptor cell and its nucleus contain almost all of the information necessary for proper response, and hormones serve only to activate the response. An analogy is a computer, its programs, and commands. The computer is capable of carrying out numerous functions and processes, but only if properly controlled; the same is true of cells, tissues, organs, and entire plants. Computer programs contain the information needed to run the computer, just as the nuclear, plastid, and mitochondrial genes contain the information needed to run cells. Both computer programs and genomes contain numerous sets of information. On a computer, commands such as OPEN, PRINT, SAVE, and COPY select subroutines or programs. Hormones are thought to act as commands that activate programs within the target cells (FIGURES 14-13A and 14-13B).
In higher animals, because so many systems and responses must be activated, many distinct hormones are necessary. Because plants are much simpler, their responses can be controlled by fewer hormones; even so, the handful of known plant hormones seems inadequate. It is likely that many plant hormones are still unknown to us. Also, many responses are activated not by one hormone but by a combination or a sequence of several hormones (FIGURE 14-13C), or a particular hormone elicits different responses when present at different concentrations. The following are examples of the most well-studied hormones.
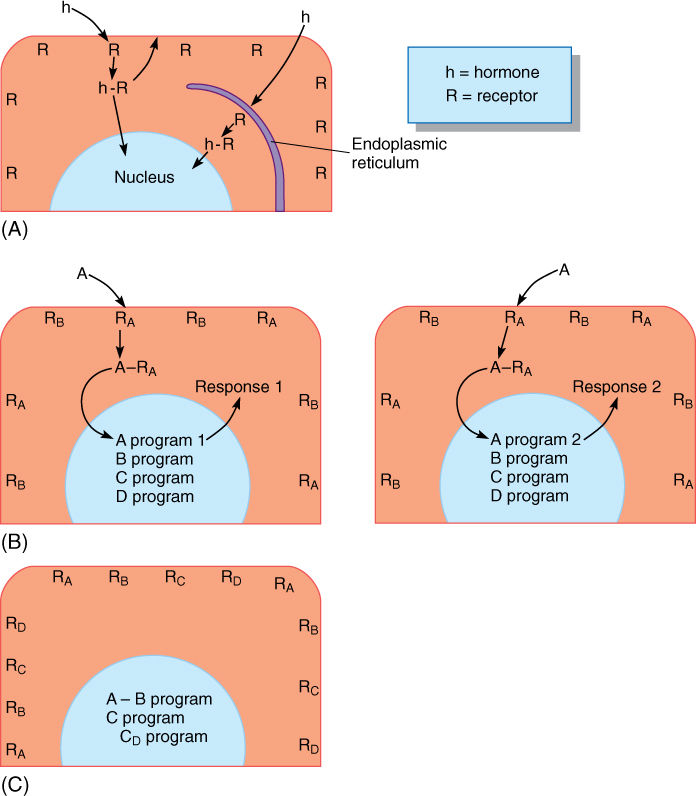
FIGURE 14-13 A cell responds to a hormone only if it has receptors for that hormone. (A) Evidence suggests that some receptors (R) are in the plasma membrane and that others are in membranes such as the endoplasmic reticulum. Once bound, the hormone-receptor complex (h-R) may cause a metabolic change immediately, or the complex may migrate to another site, such as the nucleus. (B) In many responses, some nuclear genes are activated and others are repressed. Cells may have receptors for several hormones (RA and RB); if hormone A is present, it binds and activates (or represses) program A. Other programs are unchanged. One of the results of program A might be to withdraw the receptors of either A or B from the membrane or to add receptors for C or D, thus changing the sensitivity and type of response possible. A second cell (the cell on the right) may have a different program (A program 2) activated by hormone A; the response is cell specific, not just hormone specific. (C) The effects of a hormone are often quite different when the hormone is applied alone or with or after a second hormone. In this hypothetical cell, development occurs only if hormones A and B are applied simultaneously or if hormone D is applied after hormone C.
Auxins
The first plant hormone discovered was auxin. In 1926, it was identified as the chemical messenger involved in positive phototropism in oat seedlings. Identifying it chemically at that time was impossible because it is present in such low concentrations. Experiments had to be performed by allowing auxin to diffuse out of a seedling leaf tip into a small block of agar, which was then used as if it were a small dose of auxin (see Figures 14-25G and 14-25H). The auxin was later identified as being indoleacetic acid (IAA), which could be synthesized artificially and applied to plants under various conditions to find other responses that IAA might either mediate or inhibit (FIGURE 14-14). The search was successful—dozens of responses were found (TABLE 14-3). Many compounds chemically related to IAA were also found to be as effective or even more so. It was hypothesized that IAA was only one of many natural auxins, each with its own effect and role, but further findings were not consistent with that hypothesis. Analysis of IAA metabolism showed that the compounds were converted to IAA by the plant’s enzymes.
Many synthetic compounds mimic the effect of auxin or the other hormones; for clarity, only natural products are called hormones. The term “plant growth substance” is used for any hormone-like compound, natural or artificial (Figure 14-14). Naphthaleneacetic acid, an artificial compound, produces effects in plants that are for the most part indistinguishable from those of IAA. 2,4-Dichlorophenoxyacetic acid (2,4-D) is auxin like but so powerful that it disrupts most normal growth and development even in low concentrations, making it valuable as an herbicide.
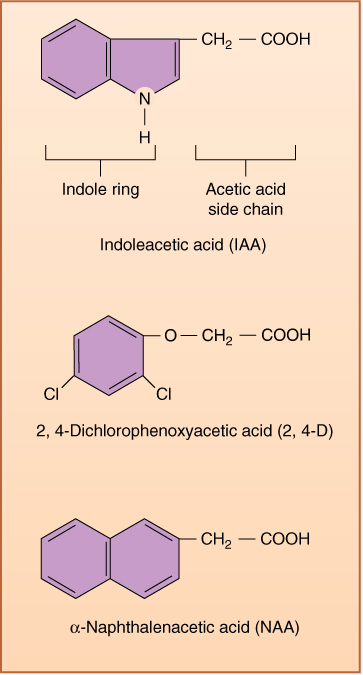
FIGURE 14-14 IAA is the natural hormone auxin; the others are synthetic. They are called plant growth substances.
IAA is related to the amino acid tryptophan; so far, at least four separate metabolic pathways are known that convert tryptophan to IAA. Some plants, corn, for example, synthesize IAA by different pathways at different stages of development, and each pathway has its own characteristic set of controls. The most active centers of auxin synthesis are shoot apical meristems, young leaves, and fruits. IAA is present in root tips, but is believed to be transported there from the shoot rather than being synthesized there.
The concentration of substances as powerful as hormones can be controlled not only by synthesis but also by destruction and by conversion to an inert storage form. Two pathways for IAA destruction are known: removal of the side group and oxidation of the five-member ring. IAA is converted to an inactive form by conjugating (attaching) it to various compounds. In the conjugated form, IAA is safe from destruction; it can be stored indefinitely in seeds, and it can be transported from cotyledons to the epicotyl during germination. Conjugation allows rapid regulation of the level of free IAA. Up to 80% of the IAA in oat seeds is conjugated, but this can be deconjugated, releasing free IAA during germination more quickly than synthesis could.
TABLE 14-3 Examples of Responses Involving Hormones
In addition to hormone transport by phloem, a second mechanism exists for auxin only: polar transport. In shoots and leaves, auxin moves basipetally—from the apex to the base of the plant, and in roots it moves acropetally toward the root apex. Movement is about 11 mm/hr regardless of whether the tissue is in a vertical, horizontal, or upside-down orientation. By means of the polar transport system, auxin movement through the plant can be maintained independently of the variation in phloem transport caused by changing sinks and sources for carbohydrates and minerals.
Cytokinins
Cytokinins were named for the fact that their addition to a tissue culture medium containing auxin and sugar stimulates cell division—cytokinesis (FIGURE 14-15). The first one discovered, kinetin, is an artificial cytokinin; however, two natural ones, zeatin and isopentenyl adenine, have been found, and more are suspected to exist (FIGURE 14-16). Cytokinins are purines, related to adenine; extensive testing of adenine analogues has been done to determine which aspects of its chemical structure are critical to the molecule’s ability to act as a cytokinin. The most active compounds have a side group containing four to six carbon atoms attached to C6. If this side group is longer or if complex groups occur at other areas, the molecule does not act like a cytokinin, apparently lacking the proper shape and charge to bind with the cytokinin receptor molecule.
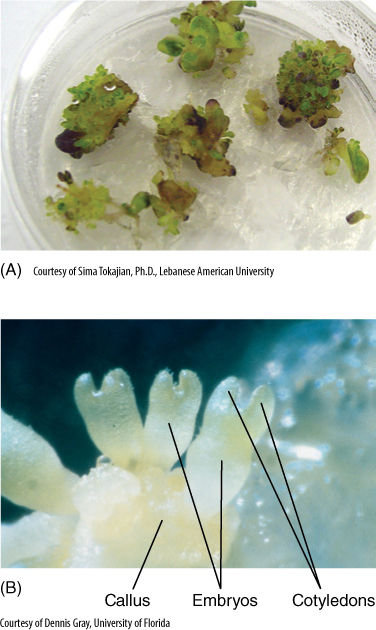
FIGURE 14-15 Cells of most dicots, such as this tobacco, can be grown in culture if provided with auxin, cytokinin, some vitamins, minerals, and sugar. The ratio of auxin to cytokinin is important, as are the absolute concentrations. (A) At one ratio, the cells proliferate as a callus composed of parenchyma. (B) At another ratio of auxin to cytokinin, buds form in the callus and then grow into embryos.
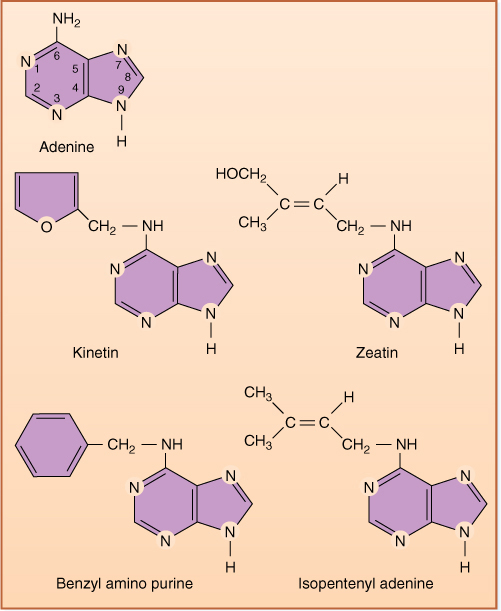
FIGURE 14-16 Both natural and artificial cytokinins are related chemically to adenine. The size and chemical nature of the group on C6 is critical.
Like auxin, cytokinins are involved in dozens of responses in all parts of the plant (Table 14-3). One important response is root-shoot coordination. As roots begin to grow actively in the spring, they produce large amounts of cytokinins that are transported to the shoot, where they cause dormant buds to become active and expand. The richest concentrations of cytokinins generally occur in endosperm and are apparently involved in controlling the development and morphogenesis of the embryo and seed.
Gibberellins
At least 125 gibberellins are known, and rather than being named, they are just numbered: GA1, GA2, . . . GA125 (FIGURE 14-17). GA3 has the name gibberellic acid, and at first it received the greatest amount of study because it could be obtained easily from fungi and used in experiments; however, it now appears that GA1 and GA19 are more active and important in plants. Gibberellins have diverse functions but a unifying structure, the gibberellane ring system. This class of hormones is thus defined by structure: A compound cannot be a gibberellin if it does not have the gibberellane ring system.
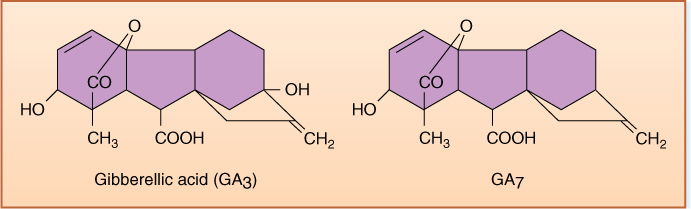
FIGURE 14-17 All gibberellins are based on gibberellane; the most common forms are GA3 and GA7. Not all of the 125 gibberellins occur in plants; at least 15 have been found only in fungi.
Gibberellin metabolism is complex. Only a few gibberellins are known to be active as hormones; others are precursors or intermediates in transforming one active form into another. Relative concentrations of the various gibberellins change in response to environmental signals. When spinach is exposed to long days (summer conditions), the level of GA19 undergoes a fivefold decrease; GA20 and GA29 increase drastically, but GA17 and GA44 do not change. As a result of these changes, spinach stems begin to elongate: long days are the stimulus; shoot elongation is the response; and alterations of gibberellin levels is the mechanism linking the two. Gibberellin metabolism appears to occur in all parts of the plant, but seeds, roots, and leaves are especially important. Gibberellin transport is studied by giving plants synthetic, radioactive gibberellins and then tracking the movement of the radioactivity. Both xylem and phloem sap become radioactive, and it is believed that the large amounts of gibberellins synthesized by active root tips are loaded into xylem. Gibberellin movement from leaves and seeds appears to correlate more closely to phloem sap dynamics. Like all other hormones, gibberellins are involved in numerous responses (FIGURE 14-18).
Abscisic Acid
This class apparently contains the single compound abscisic acid (ABA) (FIGURE 14-19). As its name suggests, it was thought to play a role in the abscission of fruits, leaves, and flowers, but currently, we do not know whether that is true of many species or only sycamore, the species in which it was discovered. ABA is widely regarded as a growth inhibitor, possibly involved in inducing dormancy in buds and seeds; however, dormancy is not just inhibition of growth, but a complicated set of changes that prepare the plant or seed for adverse conditions.
ABA appears to be especially involved in many types of stress resistance. Heating leaves, waterlogging roots, chilling, and high salinity have all been found to cause sudden increases in ABA. If healthy plants are pretreated with ABA, they become much more resistant to stressful conditions. When plants begin to wilt, the concentration of ABA in leaf cells increases dramatically from about 20 μg/kg fresh weight to 500 μg/kg, and guard cells close stomatal pores. Wilt-induced production of ABA overrides all other stomatal controls; other mechanisms that normally cause stomata to open are ineffective when ABA is present. The stimulus for ABA production appears to be soil dryness, which is detected by roots. When soil dries to some critical level, ABA production is dramatically increased and the ABA moves upward through the xylem to the leaves, causing stomatal closure.
ABA can be removed by being converted to phaseic acid, which has no known hormonal activity.

FIGURE 14-18 Many biennial plants grow as a rosette in their first year and then elongate rapidly (bolt) in their second year, producing a tall shoot that bears numerous flowers. The bolting is controlled by gibberellic acid; if it is absent, the plants remain short. The plant shown here, Arabidopsis thaliana, completes this cycle in just a few months, not 2 years: All its large leaves are arranged in a rosette at ground level; after it was several months old, it bolted and produced the flowering stalks with long internodes.
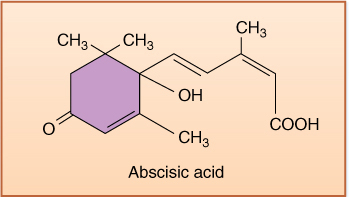
FIGURE 14-19 ABA is transported rapidly between cells and through phloem; therefore, its presence in a tissue is not proof that it was produced there. ABA can be synthesized from mevalonic acid in roots, stems, leaves, fruits, and seeds of various species.
Alternatives
BOX 14-1 Simple Bodies and Simple Development in Algae
Angiosperms have complex bodies, which require them to control many aspects of development, but there are alternatives. Green algae have much simpler bodies. Unicellular algae such as Chlamydomonas have bodies consisting of just a single cell. No middle lamella is formed during cell division; therefore, nothing holds the two daughter cells together and they just swim away from each other (they have flagella). Each individual has a body (the single cell) but no tissues or organs. They develop polarity with a front end that faces forward as they swim, and a rear end.
Colonial green algae are slightly more complex. These cells also lack a middle lamella but instead produce a gelatinous material that holds cells near each other: As a zygote divides and multiplies, all daughter cells remain loosely bound together. There is so little interconnection and interdependency that each group is considered to be a colony of individuals rather than the body of a single individual. In Gonium, each colony contains only a few cells (4, 8, 16, or 32), and the only sign of organization is that all flagella beat in a coordinated fashion. Pandorina is slightly more derived because it shows a trace of differentiation; the colony swims in one direction, and anterior cells are slightly different from posterior ones. Volvox colonies contain up to 50,000 Chlamydomonas-like cells and are easily visible without a microscope (FIGURE B14-1A). Slight differentiation exists in that up to 50 cells in the posterior half of a colony are specialized for reproduction only.
In green algae that do produce a middle lamella, daughter cells remain bound together and form a single integrated individual with a multicellular body. After the zygote of Ulothrix divides, one of the two daughter cells becomes a colorless adhesive “holdfast” cell that glues itself to a stone or seashell. The other daughter cell produces no adhesive, and its plastids develop into chloroplasts: Already the body has polarity, differentiation of two cell types, and can detect which cell is in contact with a solid substrate. The chlorophyllous cell divides repeatedly but always with cross-walls parallel to each other, and thus, the body grows as a radially symmetrical filamentous body one cell thick but many cells long. This requires morphogenetic mechanisms that precisely control the orientation of cell division. In some filamentous species, a few cells undergo a longitudinal division, and then one of the two daughter cells grows out as a filamentous “branch.” Laminar bodies, such as those of Ulva (sea lettuce; FIGURE B14-1B), are slightly more complex: A cell settles down and begins to grow into a Ulothrix-like filament, but its cells divide in two directions and thus form a sheet. Then all cells divide once in a third direction; as a result, the sheet becomes two layers thick. Both filamentous and laminar bodies remain simple with regard to cell differentiation, having only holdfast cells and chlorophyllous cells. Also, their bodies are soft and flexible, being supported and moved by water, so they do not have phototropism or gravitropism.
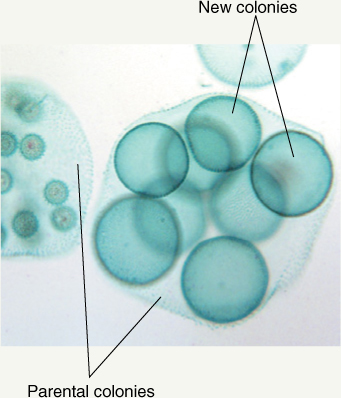
FIGURE B14-1A Colonies of Volvox; all cells are almost identical; a few are specialized for reproduction. Little differentiation occurs.

FIGURE B14-1B Individuals of the laminar alga Ulva are two cells thick. The holdfast is not visible here.
Several groups of green algae have parenchymatous bodies. Chara has a stem-like body up to 1 m long, divided into “nodes” and “internodes,” with whorls of branches arising in a precise pattern at internodes. The body is several cells thick, with large multinucleate cells in the center of each internode and smaller uninucleate cells surrounding them as well as composing the nodes and branches. All constitute a true parenchyma tissue because they are derived from an apical meristem that contains a prominent apical cell that divides in all three planes. Reproduction in Chara is significant. Its sperm cells are produced in a truly multicellular structure (a gametangium) whose outer cells are sterile; only the inner cells convert to sperm cells (FIGURE B14-1C). At maturity, the outer cells separate slightly and the motile sperm cells swim away. The egg is formed as the terminal cell of a short filament three cells long, but the subterminal cell subdivides, and those cells grow upward and surround the egg, differentiating into a variety of specialized cells. After fertilization, the sterile cells surrounding the fertilized egg deposit thickenings on their inner walls, those adjacent to the zygote. The resting structure thus consists not only of a thick-walled zygote but also protective sterile cells. Both sperms and eggs are produced in gametangia consisting of two types of tissue (fertile and sterile): Reproduction requires the integrated functioning of several types of cells. This is an organ-level of differentiation. Furthermore, germination of the zygote is reported to be promoted by cold temperatures and by red light; development responds to environmental cues.
Although all algae are much simpler than true plants, all undergo at least some development and morphogenesis, and Chara and its relatives have numerous sophisticated developmental metabolisms.

FIGURE B14-1C Bodies of Chara are several cells thick and are true parenchyma, formed by an apical meristem. Bright orange round structures are about to release sperm cells, yellow, elongate structures contain eggs; each of these gametangia consists of some fertile cells surrounded by several types of sterile cells. Individuals of Chara undergo considerable differentiation.
Ethylene
Ethylene is the only gaseous plant hormone, and it has the simplest structure (FIGURE 14-20). Its most commonly studied effects occur during fruit development. Fruits such as apple, avocado, banana, mango, and tomato are climacteric fruits: They ripen slowly as they mature, but in the final stages, numerous developmental changes occur rapidly. Starches are converted to sugars. Cell walls break down and soften. Flavors and aromas develop, and color changes. Ethylene stimulates these changes, but at first, so little ethylene is present that the changes occur slowly; however, one effect of ethylene in these fruits is the production of more ethylene, which constitutes a positive feedback system: The concentration increases exponentially and rapidly. The sudden burst of ethylene and the rapid completion of maturation of the fruit are known as a climacteric; at this time, ethylene production can be as high as 320 nL/g/hr (one nL = one nanoliter = one billionth of a liter). In nonclimacteric fruits, such as cherry, lemon, and orange, ethylene does not stimulate its own production; therefore, ethylene levels remain stable and no sudden change occurs just before maturity.
Because ethylene controls the ripening of most of our important food fruits, it is important commercially. It is used in harvesting cherries, cotton, and walnuts by causing their uniform abscission; it also synchronizes flowering and fruiting in pineapple, making harvesting easier. Ethylene can be drawn out of unripe fruits by storing or transporting them in a partial vacuum. When they reach market, air pressure is returned to normal; ethylene accumulates, and ripening occurs. Fruits may also be treated with 2-chloro-ethylphosphonic acid (commercial trade name Ethrel), which breaks down and releases ethylene.
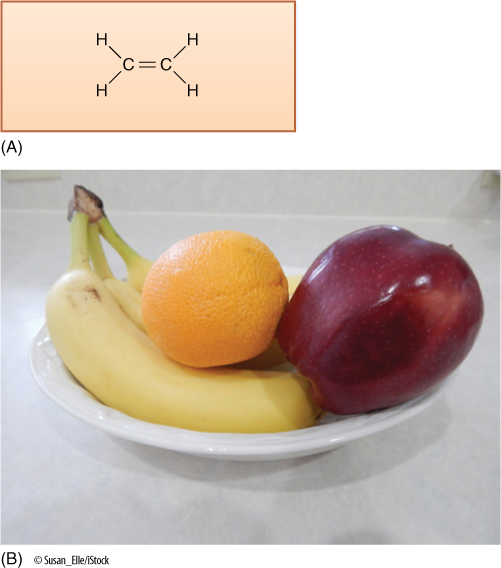
FIGURE 14-20 (A) Ethylene is a simple, small molecule. Many of its effects are blocked by carbon dioxide, whose size and shape are similar enough to those of ethylene that it can bind to ethylene’s receptor and block normal response. (B) Bananas produce large amounts of ethylene as they ripen. This diffuses away and stimulates other types of fruits (called climacteric fruits) to ripen rapidly. People often place a single banana, or even just a banana peel, into a plastic bag with other fruits: The plastic traps the ethylene, which then causes the other fruits to ripen faster and more fully. Similarly, a bruised apple produces large amounts of ethylene, which can cause other apples to overripen, thus, the old saying “One bad apple spoils the barrel.”
Being a gas, ethylene moves rapidly through tissues by diffusion rather than by specific transport mechanisms. In many cases, it acts as a final effector for auxin. Arrival of auxin at a target site often causes that site to produce ethylene, which diffuses rapidly and triggers responses in the adjacent area more quickly than the auxin itself could. Being a gas also allows ethylene to escape from plants through stomata and lenticels, which makes it a means of detecting flooding. When rivers flood, ethylene builds up in submerged stems and induces the formation of aerenchyma in certain species.
Other Hormones
Several other compounds show hormone-like activity but are either not present in all plants or are much less well studied.
Brassinosteroids are complex chemicals involved in leaf morphogenesis, root and stem growth, and vascular differentiation. If plants of Arabidopsis thaliana are artificially mutated to disrupt brassinosteroid synthesis, the plants develop abnormally but can be rescued by applying small amounts of brassinosteroids.
Jasmonic acid is involved in defense against animals and fungi. When animals chew into plants, membrane lipids are released, and one of them, linolenic acid, is converted to jasmonic acid. This travels to healthy cells and induces them to activate the genes necessary to synthesize alkaloids and chemicals that inhibit protein-digesting enzymes; both of these interfere with an animal’s ability to eat and digest. When fungi attack plants, they secrete small peptides and oligosaccharides, and several of these somehow cause jasmonic acid to be produced; it then stimulates plant cells to form an antifungus compound. Plants can attach a methyl group to jasmonic acid, creating methyl jasmonate, which diffuses away from the plant and stimulates adjacent plants to become resistant to pathogens.
Salicylic acid, related to aspirin (acetyl salicylic acid), is involved in resistance to pathogens, especially viruses. Artificial inoculation of leaves with viruses causes salicylic acid levels to rise, and these in turn activate disease resistance genes. The resistance mediated by salicylic acid is not confined to just the site where the viruses were applied; instead, all parts of the plant become resistant. This is known as systemic acquired resistance. As with methyl jasmonate, methyl salicylate is volatile and is carried by wind to other plants, alerting them to the presence of pathogens in the area. Both chemicals are part of a method of plants communicating with each other. Also, staminate flowers of voodoo lilies (Sauromatum) produce salicylic acid, which then travels downward through the thick inflorescence axis until it reaches the base; there it induces certain cells to undergo thermogenic respiration, heat up, and give off foul-smelling fragrances that attract pollinators.
Signal Amplification Cascades
Most chemical messengers are not transported specifically from the tissues that produce them into the tissues that will be controlled by them. Instead, the cells that produce them release them to be moved symplastically (from cell to cell by plasmodesmata) or apoplastically (they move through cell walls) or into phloem. Consequently even though a source might provide quite a bit of hormone, the concentration decreases as the hormone spreads through the plant, and only a few hormone molecules might actually bind to receptor molecules on the target cell. The signal is very weak and must be amplified by the target cell.
Most plants and animals are diploid, so most nuclei have only two copies of each gene. Imagine that a hormone needs to activate a particular gene, perhaps one for cell growth. Each diploid nucleus has approximately 60,000 genes (two sets of 30,000 each) and several billion nucleotides. The chances are very low that a few hormone receptor-complexes would diffuse from the plasma membrane into the nucleus and somehow encounter the correct genes. It is more likely that some would break down in the cytoplasm or be absorbed by the vacuole or spend days going from one gene to another, never finding the correct two.
In all cases that have been studied, these weak signals are amplified by the target cell. When a hormone binds to its receptor, the receptor usually changes shape and becomes an active enzyme, typically a kinase, an enzyme that transfers a phosphate from ATP on to some other molecule. Because the hormone-receptor complex is an enzyme, it functions repeatedly: The single act of one hormone molecule binding to its receptor is amplified into hundreds of molecules being phosphorylated. What do the second, newly phosphorylated molecules do; do they go into the nucleus and bind to DNA? No. Typically they also become active kinases and thus phosphorylate thousands of a third type of molecule, amplifying the signal even further. At this point there are now thousands of messenger molecules, not just the original one or two or three, so chances are very high that many will enter the nucleus and some will encounter the proper genes quickly. This is called a signal amplification cascade.
An example will make this clearer. One of the most important signal amplification cascades is the MAP kinase cascade. It plays an important role in signal amplification for many types of development in plants and animals, and is believed to be to have arisen very early in the evolution of eukaryotes (MAP stands for mitogen activated protein; it is involved in mitosis). Hormone binding converts the receptor into a kinase called MAP kinase kinase kinase (MAP 3K). This strange name means that there are three steps in this amplification cascade: MAP 3K phosphorylates and activates hundreds of molecules of MAP kinase kinase (MAP 2K), each of which phosphorylates thousands of molecules of MAP kinase (MAPK). MAPK then finds and activates various factors needed to control gene expression and development in the target cell. This cascade allows one hormone molecule to trigger the activation of thousands of important regulatory molecules within just a few seconds or minutes. It also turns out that MAP 3K phosphorylates several factors in addition to MAP 2K, and similarly MAP 2K activates various other factors besides MAPK. At each step, more and more types of response factors are being activated.
Many other signal amplification cascades exist, some triggered by certain hormones, others being found only in certain types of response. The various cascades interact in some situations. For example, two hormones might both activate the same cascade such that the two hormones reinforce each other’s effect. In contrast, the effect of one cascade might be to shut down or modify another cascade such that one suppresses or alters another.
![]() Activation and Inhibition of Shoots by Auxin
Activation and Inhibition of Shoots by Auxin
Auxin is often described as a growth hormone, whereas ABA is considered an inhibitor; unfortunately, such characterizations are confusing. Hormones simply carry information about the status of a particular region, nothing more; whether the elicited response is inhibition or activation depends on the site of response. An example of the complexity is provided by shoot tips.
As shoot apical meristems grow and initiate the new cells of shoots and leaf primordia, they also produce the auxin, IAA. Young leaves are also a rich source of this hormone. No external signal must be perceived to initiate auxin production; instead, this is a means of integrating the plant during ordinary growth. Large quantities of auxin indicate to cells that shoots are elongating and producing new leaves. Although neither signal perception nor transduction occurs, transport takes place. In stems auxin undergoes basipetal, polar transport at a speed of 5 to 20 mm per hour, perhaps by means of molecular pumps in plasma membranes. This downward flow of auxin surrounds all stem cells, and at least three cell types are set to respond to it, each response unique to the particular cell type.
Cell Elongation
In cells of the young internodes just below the apical meristem, auxin triggers cell elongation. When IAA contacts these responsive cells, which are prepared for growth, it binds to a receptor, thought to be a small protein called ABP1 (Auxin Binding Protein 1). The cells begin to transport protons actively out across the plasma membrane (FIGURE 14-21). This has the effect of acidifying the cell wall. The protons break some of the chemical bonds that hold one cellulose microfibril to another and activate enzymes that weaken other bonds so that the wall becomes weaker. If the protoplast is turgid and pressing against the wall, it exerts enough pressure to stretch the weakened wall and growth results. Immature cells neither excrete protons nor grow if auxin is lacking. At lower internodes, fully grown, mature cells apparently lack the proper auxin receptors because auxin does not cause them to extrude protons or grow.
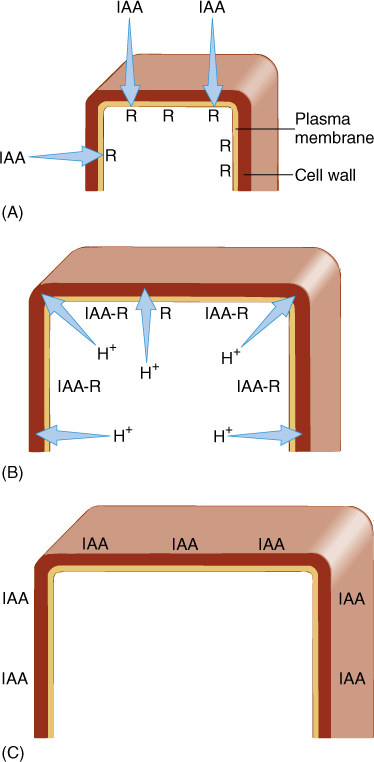
FIGURE 14-21 Cells in the subapical region have auxin receptors. If the apex is growing and producing auxin, IAA is present to bind to the receptors (A). After stimulation by auxin binding, the plasma membrane pumps protons from the cytoplasm into the wall (B), weakening it and allowing turgor pressure to stretch it. (C) Cell elongation stops once the maximum cell size is reached, and adding more auxin does not cause any more elongation; perhaps the receptors have been removed from the membrane.
Apical Dominance
The second site of response to apically produced auxin is the buds located in leaf axils; their response is not cell elongation but rather inhibition of growth. Apically produced auxin induces dormancy in these axillary buds, the result being that each shoot tip has only one active apical meristem, a phenomenon called apical dominance (FIGURE 14-22). This is a threshold response: As the terminal shoot apical meristem grows away, the concentration of auxin around an axillary bud gradually decreases until at some point it drops below the threshold. Inhibition cannot be maintained, and the axillary bud becomes active and grows out as a branch or flower. As the axillary bud grows, it produces auxin but does not inhibit itself, although it does inhibit all its own newly formed axillary buds.
Differentiation of Vascular Tissues
The third site of response to auxin produced in shoot tips is the vascular cambium; the response is cell division and morphogenesis. In springtime, as air temperatures rise and buds become active, their auxin moves basipetally, activating the dormant vascular cambium. Auxin not only stimulates cambial cells to begin mitosis and cytokinesis but also causes new daughter cells to differentiate into xylem cells. If an apical meristem is destroyed, by insects or a late frost for example, the basipetal flow of auxin stops, vascular differentiation is interrupted, internode elongation ceases, and apical dominance is broken. Some axillary buds, now free of apical dominance, become active and re-establish a flow of auxin that maintains the vascular cambium and any other cells that depend on it.
Three separate target tissues give three distinct responses, not because there are three separate chemical messengers—there is only auxin—but because part of their previous differentiation was preparation to respond to auxin in a particular way. It is important to realize that auxin carries no information except that the shoot apex is healthy and active. Each target site must have receptor molecules that interact with IAA and therefore detect its presence or absence. It is not known whether all three targets have the same receptors or whether each has a unique type, nor is it known how the interaction of IAA with the receptor triggers the response.
![]() Interactions of Hormones in Shoots
Interactions of Hormones in Shoots
In some species, apical dominance may involve only the presence or absence of auxin; in others, there is an interplay of two or three hormones. Active roots synthesize cytokinins that are transported to the shoot and stimulate axillary buds. Whether buds become active or remain dormant depends on the relative amounts of the two hormones. If a plant is growing vigorously, its roots are active and cytokinin levels are high; many buds at a distance from a shoot apical meristem have a low auxin/high cytokinin ratio and become active. Such a mechanism is adaptive because if a plant is growing well, activating more dormant buds increases the rate of new leaf production. The role of ABA in apical dominance is uncertain. It is present in quiescent buds but does not decrease either just before or as buds are becoming active and growing out.
Apical dominance in prickly pear cacti (Opuntia polyacantha) is more elaborate. The spine clusters are short shoots, and the spines are highly modified leaves. If the spine cluster is excised and placed in a tissue culture with cytokinin, the dormant short shoot apical meristem grows out as a long shoot—a new “pad” similar to a normal branch. If the culture medium contains gibberellins instead of cytokinin, the short shoot apical meristem produces more spines—it acts as a rejuvenated short shoot.
In the last few years, we have made great progress in understanding the ways in which hormones and peptides interact to control shoot apical meristems. Cells in the meristem itself divide and grow, producing more cells for the shoot: The rate at which new cells are produced must match the rate at which some cells stop dividing and differentiate into mature cells of epidermis, cortex, xylem, and so on. If too few cells differentiate, the meristem would enlarge; if too many differentiate, the meristem would shrink.
In the center of the shoot apical meristem, some cells express a gene named WUSCHEL (WUS), which produces a protein that stimulates growth and division of meristem cells (Box 14-2). If only WUS were active, the meristem would constantly grow larger. But another group of cells expresses the gene CLAVATA3 (CLV3), which limits meristem size. The two make a negative feedback loop because each affects the other: WUS activates CLV3, which inhibits WUS. If WUS becomes too active, it stimulates CLV3, which then inhibits it. But if CLV3 is too active, WUS becomes too quiet and the cells are not stimulated to make CLV3. Other genes and their peptide products are involved, but the interaction of WUS and CLV3 represents a common type of feedback loop that controls many aspects of differentiation and morphogenesis.
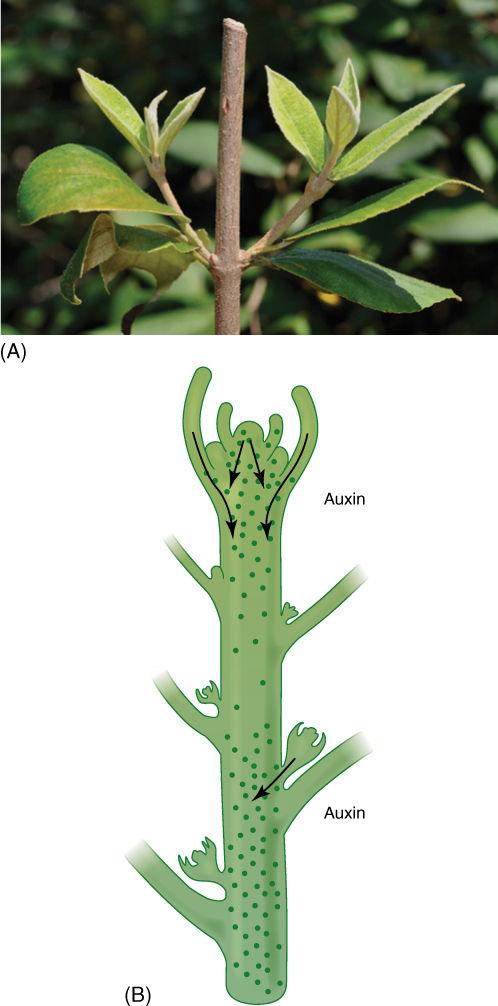
FIGURE 14-22 (A) This twig of Viburnum was pruned 2 months before the photo was taken. The axillary buds had been completely inhibited by the shoot apical meristem at the time of pruning, but then both became active once pruning stopped the flow of auxin from the shoot apex. (B) The concentration of auxin is greatest at the shoot apex and less at lower levels; at sites where it drops below the threshold level, it can no longer suppress axillary buds, and one or several become active. Other factors must be involved because it is not always the lowest bud that becomes active first, even though it should be the first to encounter sufficiently low auxin concentrations.
Botany and Beyond
BOX 14-2 Names of Genes
Plant scientists working with Arabidopsis thaliana have agreed that the wild-type form of the gene (the form found in most natural populations) should be written in ITALICIZED FULL CAPS, whereas any mutant allele is written in italicized lower case; if the protein itself is discussed, it is in ordinary font. Often, genes are discovered by exposing thousands of seeds to mutagenic chemicals, then sowing the seeds and looking for any aberrant plants with interesting characters. After such a procedure one plant of A. thaliana had many more stomata than is typical; that plant was cultivated to maintain the mutated form of the gene. The gene was named after its mutant form: TOO MANY MOUTHS (TMM) (full caps indicates this is the wild-type allele; italics indicates it is the gene rather than the protein), with the newly-produced allele being too many mouths (lowercase indicates this is the new mutant allele), the protein coded by the wild-type gene is TOO MANY MOUTHS, and the protein produced by the mutant allele is too many mouths.
TABLE B14-2 Names of Genes
Notice that an odd thing about this system is that genes are named after mutant, aberrant characters: The wild-type, ordinary TMM version of the gene produces just the right number of stomata, but its name suggests it produces too many.
If the sequence of the TMM allele is known, then the equivalent genes can be searched for in other plants whose development and genomes are well known, for example snapdragon (Antirrhinum majus), tobacco (Nicotiana tabacum), and other plants commonly used in developmental studies. If they are found, then the species initials are used to distinguish them: atTMM, amTMM, ntTMM. These three genes all descended from an ancestral TMM gene.
The WUS/CLV3 interaction changes both during development of an individual plant and during evolution. In many species, the shoot apical meristem of a germinating seed is very small, but it gradually grows to be a larger size and then stabilizes. By some unknown mechanism, the WUS gene in seedlings is temporarily able to expand its influence and causes a greater number of cells to remain meristematic without triggering CLV3 to inhibit it. When the proper adult size is reached, WUS and CLV3 balance each other and the meristem stops enlarging (FIGURE 14-23).
Species differ in the typical size of their shoot apical meristems. The typical diameter in Arabidopsis thaliana is about 80 μm, but in other species, meristems are as narrow as 50 μm, and 200 μm is probably the most common diameter among angiosperms. A few species of cycads and cacti (especially Echinocactus platyacanthus and E. grusonii) have gigantic shoot apical meristems with diameters up to 2500 μm (2.5 mm, they are visible with the naked eye) in diameter. This may mean that during evolution of these species, the WUS gene has evolved such that it is less sensitive to inhibition by the CLV3 gene, or perhaps it is CLV3 that is less sensitive to stimulation by WUS. Of course, it is also possible that other, unknown factors are involved.

FIGURE 14-23 Each cluster of spines on a cactus is an axillary bud, so even though the foliage leaves are too small to be visible, the spine clusters reveal the pattern of leaf initiation by the shoot apical meristem. While this saguaro (pronounced sa WAH row; Carnegiea gigantea) was very young, its shoot apical meristem was so small and narrow that only a few rows of leaf primordia could fit in the meristem’s leaf initiation zone. Consequently, there are only a few rows of spine clusters at the base of the shoot. But as the shoot became older and taller, the meristem increased in diameter so much that there was room for another row of leaf primordia; this happened several times. Arrows indicate where the new rows were initiated. At some point in the future, this shoot’s apical meristem will stop enlarging and after that, the number of ribs will remain constant.
A second interaction of hormones occurs in the vascular cambium. Auxin alone activates the cambium and elicits differentiation of xylem, but gibberellin is also present in a healthy stem and causes some of the new cells to differentiate as phloem. Without the interaction of both auxin and gibberellin, a normal, functional vascular system would not develop.
![]() Hormones as Signals of Environmental Factors
Hormones as Signals of Environmental Factors
Leaf Abscission
Whereas normal growth of shoots and roots results in large flows of auxin and cytokinin, respectively, environmental factors also influence hormone concentrations. Hormones communicate to various parts of a plant the information that a particular part has encountered an environmental change. Export of ABA by wilted leaves has been mentioned, and another example involves abscission of leaves and fruits. A young leaf produces large amounts of auxin, but production falls to a low but steady level in a mature leaf. As long as auxin flows out through the petiole, activity in the abscission zone is inhibited (FIGURE 14-24). If the leaf is damaged by animal feeding or water stress, auxin production drops to such a low level that its flow through the petiole does not keep the abscission zone quiescent. Perception and transduction in this case may be simply that insect or wilt damage makes it impossible for the impaired cells to produce enough auxin to inhibit the abscission. Old age of the leaf may also result in the lack of sufficient auxin, but evidence suggests that autumn conditions stimulate production of ethylene, which then suppresses auxin production and transport in time for abscission before winter.
Fruits are prevented from abscising prematurely by the presence and export of sufficient amounts of auxin through the pedicel. Fruit ripening is under the control of both auxin and ethylene, at least in edible, fleshy fruits. Initial transformation of the ovary wall into a fruit is a response to auxin synthesized in developing embryos and transported to the ovary wall. Auxin stimulates many changes, including cell enlargement and differentiation; there is usually surprisingly little cell division during the formation of fruits, even large ones. One effect of auxin is release of ethylene by the developing fruits, which leads to other aspects of ripening in both climacteric and nonclimacteric fruits. At maturity, the high concentration of ethylene stimulates the pedicel abscission zone, overriding the presence of auxin.
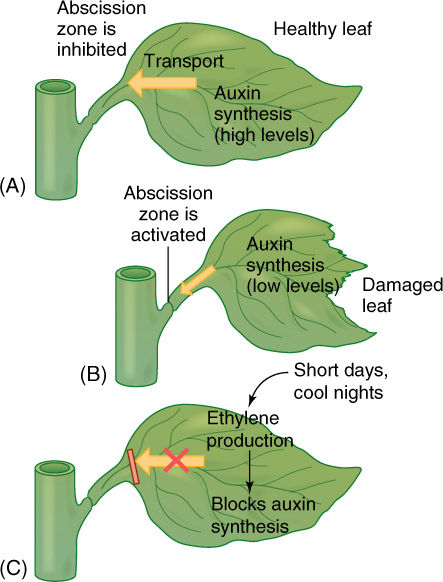
FIGURE 14-24 (A) A healthy leaf produces and transports enough auxin to suppress activity in the abscission zone. (B) A damaged leaf produces less auxin, insufficient to prevent abscission. (C) Autumn stimuli may cause the production of ethylene, which then suppresses auxin synthesis and transport. An alternative hypothesis postulates that autumn conditions directly suppress auxin production and ethylene is not necessary.
Tropisms
Light is the stimulus in phototropism; therefore, a pigment is needed, and its absorption spectrum must match the action spectrum (just as the spectra of chlorophyll and photosynthesis must match each other). The action spectrum indicates that blue is the most effective wavelength for phototropism, and the pigment whose absorption spectrum matches that is a small protein, now called phototropin, with two flavin mononucleotides attached as chromophores. Phototropin is a component of a cell’s plasma membrane and becomes phosphorylated when exposed to blue light. Blue light stimulates other aspects of plant development also (it inhibits hypocotyl elongation once a seedling has emerged from underground; it stimulates chlorophyll and carotenoid synthesis and causes opening of stomata at dawn), and together these are called blue-light responses.
Oat seedlings are often used to study phototropism. They are grown in the dark (surviving on their endosperm), and if illuminated briefly with blue light from one side, they bend toward it. Plants grown in darkness do not have chlorophyll so that pigment is not present to interfere with studies of absorption and action spectra or studies of presentation time and threshold amount of light needed. Oat seedlings have an outermost protective leaf called a coleoptile (pronounced coal ee OP tile—the p is pronounced), which shows a strong positive phototropic response; oat coleoptiles are the organ studied most often.
The site of perception of blue light is the tip of the coleoptile, and if it is covered with an opaque hood, light direction is not detected. Similarly, if the coleoptile tip is cut off before unilateral illumination, the rest of the coleoptile does not perceive the light. We do not know the steps of transduction, but the result is an asymmetric redistribution of auxin: Auxin is transported from one side of the coleoptile to the other. Neither synthesis nor destruction of auxin is affected. After the differential redistribution of auxin is established, the darker side of the coleoptile, or the stem in other plants, receives extra auxin, and thus it grows more rapidly and bends toward the light. When the coleoptile or stem points directly at the light, neither side is brighter nor darker; as a result, differential auxin transport stops, and the organ grows straight ahead. The site of response, where differential growth causes bending, is approximately 5 mm below the coleoptile tip (FIGURE 14-25).
In positive gravitropism in roots, the root cap acts as the organ of perception (see Figure 14-11). After the lower side of the root is detected, a growth inhibitor is transported to that lower side of the root cap and then into the root, where it slows growth on the lower side. We know almost nothing about how the position of the statoliths is transduced, nor are we certain of the growth inhibitor. Much circumstantial evidence indicates that it is auxin. How can auxin be a growth stimulator in coleoptiles and stems but an inhibitor in roots? It depends on the concentration and the tissue’s sensitivity. Small amounts of auxin stimulate growth, and larger amounts inhibit it (FIGURE 14-26). Coleoptiles and stems typically have suboptimal amounts, so redistribution causes the side with more auxin to have a more nearly optimal level and that side grows faster. Roots are more sensitive to auxin and ordinarily have optimal or excess amounts; therefore, redistribution causes the side with more auxin to have such an excess it is inhibitory.
![]() Flowering
Flowering
Ripeness to Flower
Almost all plants must reach a certain age before they can be induced to flower; only a few species (Chenopodium rubrum, Pharbitis nil) can be induced as seedlings. Annual plants need to be only several weeks old before they become competent to respond to a floral stimulus, but many perennials must be 5 or 10 years old; beech trees (Fagus sylvatica) must be approximately 40 years old. Before this time, conditions that should induce flowering have no effect. Virtually nothing is known about the metabolic difference between the juvenile phase, when plants are incapable of being induced to flower, and the adult phase, when they are sensitive to floral stimuli. The conversion from juvenile to adult is called phase change.
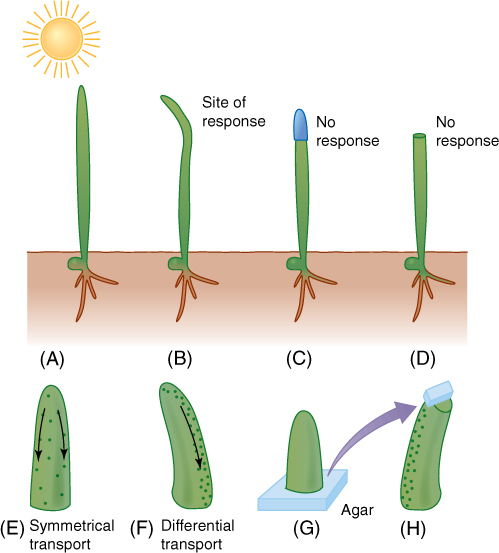
FIGURE 14-25 (A) When illuminated from directly above, oat seedlings grow upward. (B) When a young oat seedling is exposed to light from one side, its outermost sheathing leaf, the coleoptile, bends and grows toward the light. If the coleoptile apex is covered (C) or cut away (D), no response occurs to unilateral illumination, and thus, the tip is the site of perception. If the site of response is covered, bending occurs and the site of response is not involved in perception at all. (E) In dark conditions or with overhead lighting, auxin is transported symmetrically down the coleoptile, causing equal amounts of growth everywhere. With unilateral illumination (F), auxin is redistributed, with the darker side transporting more auxin than the lighted side, so the darker side grows faster, resulting in curvature. (G) and (H) Auxin can be collected in small blocks of agar or other absorptive material and then placed asymmetrically on a decapitated coleoptile; the side receiving auxin grows, but the other side does not.
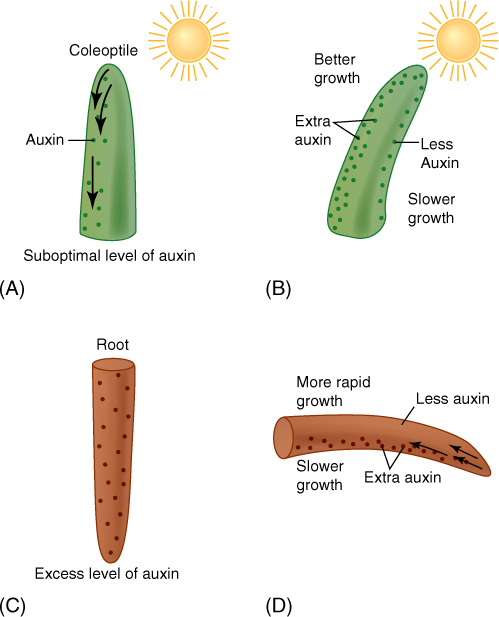
FIGURE 14-26 (A) and (B) Shoots may contain slightly too little auxin for fastest growth. Thus, redistribution in a coleoptile apex caused by unilateral light causes the side with more auxin to have a level that is more nearly optimal, whereas the other side, which receives less, has even poorer growth than before. (C) and (D) Roots may contain slightly too much auxin for fastest growth. In root positive gravitropism, the horizontal position of a root cap causes statoliths to fall to the side of statocytes, causing a downward redistribution of auxin. The lower side receives extra auxin and becomes even more inhibited, whereas the side that receives none has a more optimal amount and faster growth.
Cold temperatures are the stimulus responsible for phase change in biennial plants, and the process is vernalization (see Figures 14-6 and 14-18A). The site of perception is the shoot apex itself; if it is cooled while the rest of the plant remains warm, vernalization occurs; however, if the rest of the plant is cooled while a small heater keeps the apex warm, no vernalization occurs. Presentation time is as short as 1 day in some plants. The transduction process is known to require oxygen, carbohydrates as an energy source, and an optimal temperature just above freezing, between 1°C and 7°C. Vernalization results in a stable change. If the plants are returned to warm conditions but are not given the floral stimulus of short days and long nights, they continue to grow vegetatively, without flowering, year after year, but they retain their vernalization and flower whenever the floral stimulus is finally given.
We know little about what stimulates phase change in perennial plants. Often a tree that is capable of flowering looks the same as one that cannot flower: Leaves, twigs, phyllotaxy, and bark all seem similar. Ivy (Hedera helix) is a common plant with distinct juvenile and adult morphology: Juvenile ivy has palmately lobed leaves and climbs (it is the form you see on walls), whereas adult ivy has entire, ovate leaves and upright stems that do not climb. Ivy can persist in the juvenile condition for years, growing extensively over walls and fences, and then individual branches may undergo phase change and start to flower while adjacent branches continue in the nonflowering juvenile phase.
Several genera of cacti have a much more pronounced difference between juvenile and adult phases. Juveniles of all species of Melocactus and Backebergia grow as cacti with ordinary bodies for many years, but when they undergo phase change, the adult shoot has a narrower pith and cortex, the cortex is not green and not photosynthetic, a different kind of wood is produced, phyllotaxy changes, epidermis is immediately replaced by bark, and the size, shape, and abundance of spines are different. The adult phase differs so strongly from the juvenile phase that a single plant appears to be two entirely different plants grafted together (FIGURE 14-27). In Espostoa and Facheiroa, when a phase change occurs, it affects only one narrow strip of the shoot: As the shoot apical meristem continues to function, one side of the shoot it produces is adult (with characters like that of adult Melocactus) and able to produce flowers, and the other sides of the same shoot are juvenile and cannot flower (FIGURE 14-28).

FIGURE 14-27 The green base of this Melocactus is its juvenile phase, and the red top is its adult phase; the two are parts of a single shoot produced by just one apical meristem. While young, the plant grew for years as a juvenile, with a body resembling that of many cacti, but it was unable to flower. Once old enough, it underwent phase change and the apical meristem switched to producing the narrower, very spiny adult phase (called a cephalium), which can flower. The adult portion cannot photosynthesize because of the dense covering of spines. For the rest of its life, this plant will continue to grow as an adult, with the cephalium becoming taller and the juvenile base remaining as it is—no new photosynthetic tissues will be formed.
Photoperiodic Induction to Flower
The conversion of an adult plant from the vegetative to the flowering condition may be the most complex of all morphogenic processes. This is not one process—different mechanisms exist in different species. In certain annual species, size appears to be the only important factor: Peas and corn initiate flowers automatically after a particular number of leaves has been produced, regardless of environmental conditions; flowering is controlled by internal mechanisms. In many species, perhaps most, transition to the flowering condition is triggered by photoperiod—day length—which acts as a season indicator (TABLE 14-4). One subclass of these plants blooms when days are short (spring or fall) and are short-day plants. Another subclass, long-day plants, is induced to bloom when days are long, in summer. Plants that do not respond to day length are day-neutral plants.

FIGURE 14-28 The green parts of this Espostoa are its juvenile body, and the brown strips are its adult phase. While young, it grew as just a juvenile that had about 20 branches, but as each branch became old enough, each underwent a phase change. Only a strip of each branch became adult, however; the rest of each branch continued to develop with juvenile characters. Flowers are produced only from the brown adult regions. Unlike Melocactus, new photosynthetic tissues are produced every year in Espostoa.
TABLE 14-4 Photoperiodic Species*
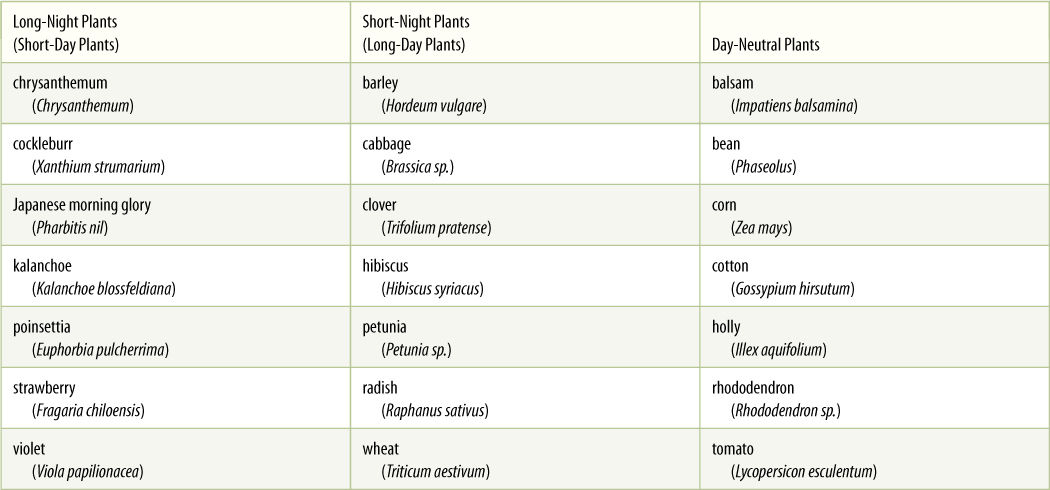
*All plants fall into one of these three photoperiod categories; this table lists only a few familiar examples.
We know much about how plants measure day length. First, the pigment phytochrome detects the presence or absence of light (TABLE 14-5 and FIGURE 14-29). Phytochrome has a light-absorbing portion (a chromophore) attached to a small protein of about 125,000 daltons. Phytochrome occurs as dimers in cells. When phytochrome absorbs red light with a wavelength of approximately 660 nm, the chromophore changes shape, and this forces the protein to change its folding. This affects many of its properties, one of which is its hydrophobicity; in the refolded state, it is more hydrophobic and binds to membranes. A second altered property is its absorption spectrum; it now absorbs not at 660 nm but in the far-red (almost infrared) region of 730 nm. When this form absorbs far-red light, however, it refolds back to the red-absorbing form and releases from the membrane. The two forms are called Pr (red absorbing) and Pfr (far red absorbing). Also, Pfr reverts to Pr in darkness. Apparently Pr is inactive metabolically but becomes morphogenically active and exerts its effect when it absorbs red light and is converted to Pfr: Pfr is the active form and brings about metabolic responses. Pfr is transported into the nucleus and binds to one or more proteins, and the combination of the two apparently activates particular genes. After a plant is given red light, converting phytochrome to the active Pfr form, exposure to far-red light converts phytochrome back to the inactive Pr form. If far-red light is given quickly enough after red light, phytochrome does not have enough time to affect cell metabolism, and no effect is seen; however, if far-red comes long enough after red for the presentation time to be fulfilled, Pfr is able to complete the transduction process and far-red light can no longer cancel the red light stimulation (see Figure 14-10).
FIGURE 14-29 The chromophore or light-detecting portion of one particular phytochrome. The rest of the molecule is a protein. The two portions are joined by a sulfur atom. We now know that there is a family of at least five closely related phytochromes; some have overlapping functions, some have distinct functions. Many details are being investigated now.
TABLE 14-5 Photomorphogenic Responses for which Phytochrome Is the Photoreceptor

A plant experiencing a short day in nature automatically receives a long night (FIGURE 14-30). Similarly, long days are always accompanied by short nights. Night length is actually the critical factor. A long-day plant is in reality a short-night plant. It can be placed in a growth chamber and artificially given both long days and long nights—for example, 16 hours of light and 16 hours of dark in a 32-hour “day.” If day length is the important factor, the plant should flower because it has long days; if night is the critical factor, then it should not flower because it does not have short nights. When the experiment is done, the long-day plant does not flower, indicating that night length is critical, not day length. Similar experiments have shown that short-day plants really are long-night plants: If given a 16-hour cycle (8 hours light/8 hours dark—both day and night are short), they do not flower, but if given long nights, even accompanied by artificially long days, the plants are induced to flower.
Each species has its own particular requirements for long or short nights; that is, not all “long nights” have to be the same length. Instead each species has a critical night length; if a short-night plant receives nights shorter than this critical length, it flowers, whereas a long-night plant must receive nights longer than its own critical night length. Because critical night length varies from species to species, it is possible for a long-night plant and a short-night plant to bloom under the same conditions if the critical night length for the short-night plant happens to be longer than that for the long-night plant (TABLE 14-6).
TABLE 14-6 Concurrent Flowering of Long-day and Short-day Plants
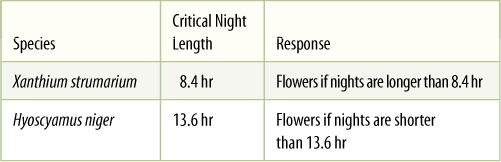
Although Xanthium strumarium is a short-day plant and Hyoscyamus niger is a long-day plant, both flower if given days 14 to 12 hr long; those days have nights 10 to 12 hr long, longer than the 8.4-hr critical night length of Xanthium but shorter than the 13.6-hr critical night length of Hyoscyamus.
Day length/night length control of flowering and other processes such as the initiation or breaking of dormancy is more common at locations farther from the equator. Away from the equator, nights become progressively shorter from winter to summer and then progressively longer from summer to winter. The greater the distance from the equator, the greater the length of the longest winter night and the shorter the length of the shortest summer night. Thus, if two species are to bloom just after the beginning of May, a species in the southern United States or Mexico must have a critical night length shorter than that of a species in the northern United States or Canada.
Presentation time varies considerably; in some morning glories, one photoperiod of the proper length induces flowering, whereas at least 1 or 2 weeks of proper photoperiods are necessary for other species. The accuracy with which night lengths can be measured varies, but the most accurate species known is the long-day (short-night) plant henbane (Hyoscyamus niger); it must have nights shorter than 13 hours, 40 minutes. If the nights are even 20 minutes too long, 14 hours long, for example, it does not flower.
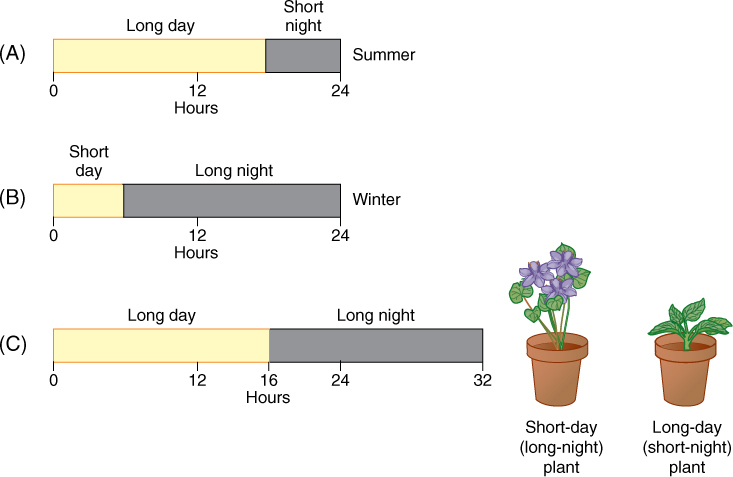
FIGURE 14-30 (A) and (B) Natural 24-hour light/dark cycles must have short nights any time the days are long (summer) and long nights when days are short (winter); thus, plants could detect season by measuring either day or night. When photoperiodism was first discovered, it was assumed that day length was important; therefore, all our terminology was based on that. (C) With electric lights, it is possible to create a long photoperiod—32 hours long here—with both long days and long nights. Under these conditions, short-day (long-night) plants bloom, but long-day (short-night) plants do not.
When phytochrome was discovered to be responsible for measuring night length, it was hypothesized that most phytochrome was converted to Pfr by the end of a day and then reverted slowly back to Pr during the night. It was postulated that if it could completely revert in the dark, before the next sunrise, metabolic changes would be triggered; however, it is now known that virtually all Pfr converts back to Pr within 3 or 4 hours, a time far too short to be a night-measuring clock by itself. It must instead interact with another type of clock that occurs in plant cells (a circadian rhythm, discussed later here).
The sites of perception for night length are young leaves. It is possible to stimulate one leaf with a spotlight and induce the plant to flower, even if the apical meristem, the site of response, is not illuminated (FIGURE 14-31). Because the site of perception is not the site of response, a chemical messenger must be transmitted between the two. If a leaf is photo induced and then immediately cut off the plant, no flowering occurs; if it is allowed to remain attached for several hours, the flowering stimulus is synthesized and transported out of the leaf. If the leaf is then removed, the plant still flowers. An obvious experiment is to induce a leaf by giving it the proper night length, then collect the sap that is transported through the petiole and assay it for the hormone that acts on the apical meristem. This has been done hundreds of times by many people, without any repeatable success; the process is not as simple as we had at first thought.
An extremely interesting set of results has been obtained by grafting together plants with different photoperiod requirements. Individuals of the tobacco species Nicotiana silvestris are long-day plants. The species N. tabacum has two types of individuals: those of the cultivar N. tabacum cv. Trabezond are day-neutral plants, and those of N. tabacum cv. Maryland Mammoth are short-day plants. When long-day and day-neutral plants are grafted together and given long days, the day-neutral plant is also induced to flower, presumably by a floral stimulus that passes through the graft union. Similarly, day-neutral plants are induced to flower if grafted to short-day plants and given short days. Grafting one day-neutral plant to another does not increase flowering, so the grafting by itself has no effect.
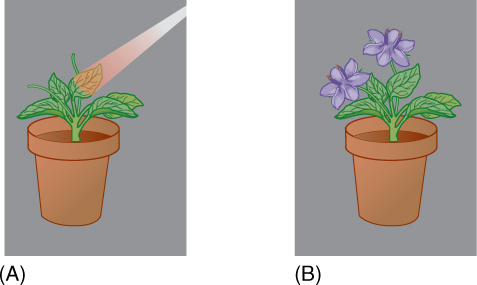
FIGURE 14-31 If a short-night plant is given long nights, it does not flower, but it is possible to cause flowering by illumination with 15 minutes of dim red light; the plant acts as though it has received two short nights separated by a 15-minute day. (A) The red light “night break” does not have to be given to the entire plant; if a narrow beam of red light shines on a single leaf while all of the rest of the plant remains in darkness, the plant flowers (B).
When the combination of short-day plants grafted to day-neutral plants was given long days, the short-day plants did not flower as expected, and the day-neutral partners flowered about the time they would have if not grafted to anything. When long-day plants grafted to day-neutral plants were exposed to noninductive short days, however, not only did the long-day plants not flower, but the day-neutral partners were also prevented from flowering. Apparently, under noninductive conditions, long-day plants actually produce an inhibitor of flowering. Flowering in these long-day tobacco plants seems to be controlled by a switch from inhibitor production to promoter production, whereas in the short-day tobacco plants, flowering is controlled only by the presence or absence of a promoter. Neither the promoter nor the inhibitor has been isolated, but gibberellins are suspected to be involved in the synthesis or activation of the promoter.
A new advance in our understanding of the control of flowering is the discovery that when leaves are given the proper day length, a gene called FT is activated. This gene codes for a protein called FT that is carried through phloem to the apical meristems. This is an exciting breakthrough, but control of flowering is complex and we must anticipate that there are many other steps to be discovered with many variations involving numerous genes other than just FT.
Endogenous Rhythms and Flowering
Plants contain endogenous rhythms; that is, certain aspects of their metabolism cycle repeatedly between two states, and the cycle is controlled by internal factors, by an internal clock. The most obvious example of this is in the “sleep movements” of the leaves of plants like prayer plant (Oxalis). In the evening, leaflets drop down, and in the morning, they raise themselves to the horizontal position as motor cells increase their turgor. It is easy to assume that this is a photonastic response, but if the plants are placed in continuous darkness, the leaflet position continues to change: They return to the up position about every 24 hours and move down about 12 hours later, controlled not by dawn or dusk but by their own internal clock. In many flowers, the production of nectar and fragrance is also controlled by an endogenous rhythm and occurs periodically even in uniform, extended dark conditions.
The underlying mechanism that constitutes the clock is poorly understood but is known to involve a negative feedback loop. Imagine just two proteins, A and B, with A activating B, and with B inhibiting A. As A works, B becomes more active and thus inhibits A, but as A becomes inhibited, it can no longer activate B. Consequently, B’s activity lowers, which allows A’s activity to increase. The system can cycle rhythmically indefinitely (compare with the positive feedback of ethylene in climacteric fruits). In Arabidopsis thaliana, the negative feedback loop involves at least three genes rather than just two: TOC1 (and light) activate LHY and CCA1, but one of the effects of these latter two is to inhibit TOC1.
When placed in uniform conditions (constant temperature and either constant darkness or constant light), the endogenous cycle typically differs slightly from 24 hours, being either somewhat longer or shorter (FIGURE 14-32); however, in nature, the rhythm is exactly 24 hours long because light is able to entrain (reset) the rhythm. The pigment responsible for detecting the light for entrainment is phytochrome. Each morning, the red light of sunrise resets the rhythm so it can never get out of synchronization with exogenous light/dark cycles.
Many types of endogenous rhythms have a period that is not 24 hours long. Cytoplasmic streaming and the spiraling motion of elongating stem tips have periods of only a few minutes to a few hours; these are ultradian rhythms. If a period is approximately 24 hours long, it is a circadian rhythm, the most common kind (TABLE 14-7). The release of gametes in brown algae is controlled by a 28-day lunar rhythm. Some seeds have an annual rhythm of germinability: If stored in uniform conditions and periodically provided with moisture and warmth, they germinate only at times of the rhythm that correspond to springtime.
The involvement of endogenous circadian rhythms in flowering was discovered during dark interruption experiments: A short-day (long-night) plant can be prevented from flowering by interrupting long nights with a brief (15 minutes or less) exposure of red light. This is detected by phytochrome, and the plant acts as though it has received two short nights separated by a 15-minute day. Short-day plants given a very long night—continuous darkness—have an endogenous rhythm of sensitivity to light breaks. If the light break is given at 6 hours into the dark period, or at 30 hours (24 + 6), 54 hours (24 + 24 + 6), and so on, the light break prevents flowering (FIGURE 14-33). These times correspond to darkness in a normal environment. But if the light break is given at a time when the endogenous rhythm would be “expecting” normal daylight conditions, such as at 16, 40 (24 + 16), or 64 (24 + 24 + 16) hours after the beginning of the dark treatment, the light break does not stop flowering. Plants kept in uniform, dark conditions undergo an endogenous cyclic sensitivity and insensitivity to red light interruption of the critical night length. Just how the endogenous rhythm and the critical night length work together to stimulate flowering is not known.
TABLE 14-7 Metabolic Processes that Undergo Endogenous Rhythms in Plants
A brief descriptions of photoreceptors is presented in TABLE 14-8.
ABC Model of Flower Organization
All flowers share the same basic developmental organization. Most have four types of “appendages”: Outermost are the sepals, then petals, stamens, and finally carpels occur in the center. There are many variations: Wind-pollinated flowers often lack sepals and petals, and in many species certain flowers lack carpels and others lack stamens (in corn, for example, the flowers in the cob have carpels and develop into the fruit we eat, whereas the tassels at the top of the corn plant have stamens but no carpels). Many of the plants we cultivate in flower gardens are “double” flowers, which means they have extra petals but no stamens (FIGURE 14-34). It is typical for eudicot flowers to have five of each appendage: five sepals, five petals, five stamens, and five carpels (and monocots typically have three of each), but some flowers have dozens of petals, 100 or more stamens, and numerous carpels (FIGURE 14-35).
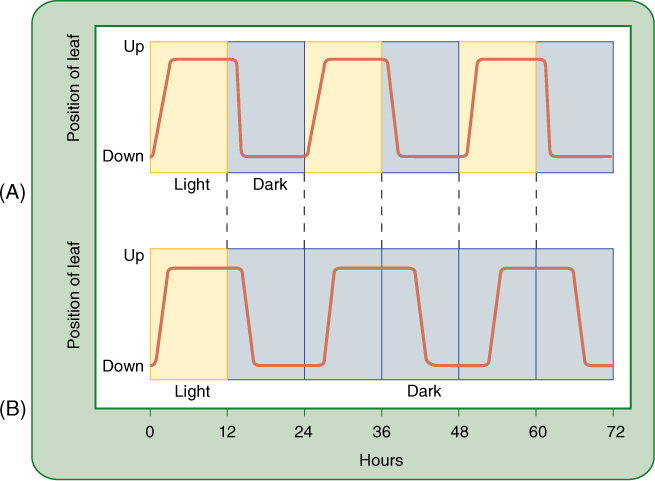
FIGURE 14-32 (A) Under natural conditions, a circadian rhythm matches the cycle of light and dark, being exactly 24 hours long. (B) In continuous darkness, most circadian rhythms have periods slightly longer than 24 hours. In nature, sunrise resets the clock by acting on phytochrome every morning.
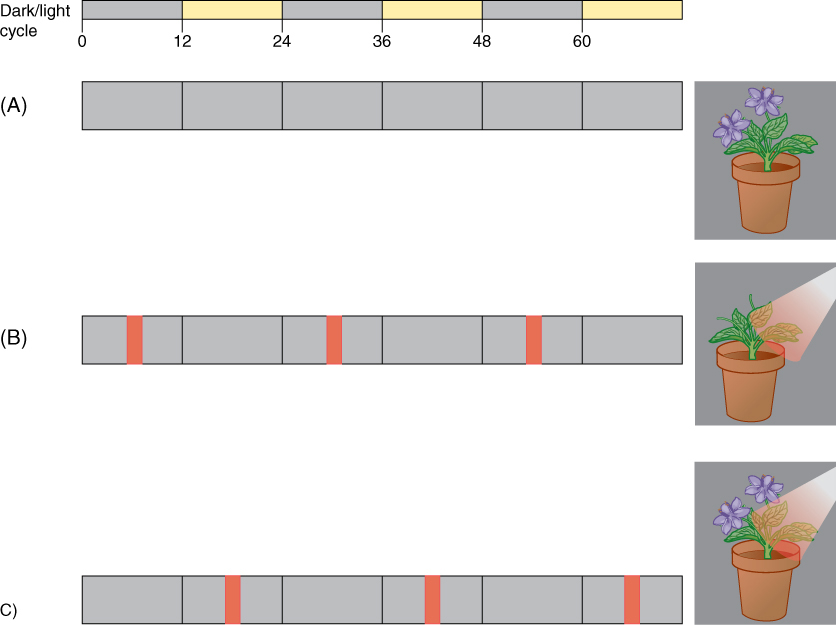
FIGURE 14-33 (A) If a short-day (long-night) plant is placed in continuous darkness, it will flower. (B) The plant can be prevented from flowering by giving it red light night breaks, but these are effective only if given at those times of the endogenous rhythm when the plant is “expecting” darkness. (C). If given when the rhythm is “expecting” light, the light breaks are ineffective and the plant will bloom. Whatever metabolism red light interrupts does not occur continuously in prolonged darkness, but rather periodically, controlled by the internal clock.
Despite the diversity it appears as if the basic organization of most flowers can be explained by a hypothesis called the ABC model. This model postulates that three fundamental genes, A, B, and C, interact to control the basic aspects of flower organization. Gene A affects the outermost, lowest regions of a flower shoot apical meristem, gene C controls the uppermost, central region, and gene B partially overlaps the other two (FIGURE 14-36A). In the outermost, lowest, doughnut-shaped region where only gene A is expressed, the primordia (similar to leaf primordia on a vegetative shoot apical meristem) develop into a whorl of sepals (flower appendages usually occur in whorls, not in spirals like leaves). Primordia in the next region upward and inward are influenced by both genes A and B, and they develop as petals. The combination of genes B and C produces stamens, and in the very center of the flower where only gene C is expressed, primordia develop as carpels.
TABLE 14-8 Photoreceptors
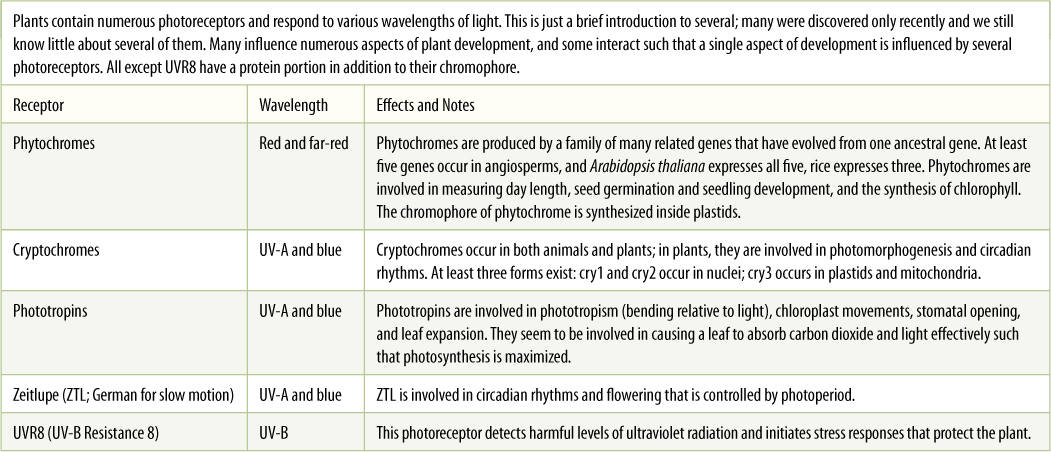

FIGURE 14-34 (A) This wild rose has only a few petals and is a “single” flower. (B) Many cultivated roses have been bred to be “double” flowers, with a mutation that causes them to produce extra petals in the place of stamens.

FIGURE 14-35 (A) This Graptopetalum is the type of flower often thought of as being a typical eudicot flower: It has five sepals, five petals, five stamens (this actually has 10, which is quite common), and five carpels. (B) This flower of Epiphyllum (in the cactus family) differs from that of a typical eudicot by having dozens of petals (36 are visible here), almost 100 stamens (many are too short to be seen here), and 11 carpels. Whereas each floral apical meristem of most species produces only a few primordia, those of cacti produce hundreds.
This model was proposed and is supported by many types of aberrant flowers. For example, if gene B is mutated and nonfunctional, sepals are produced in the outer part of the flower and carpels occupy the inner parts. There are no petals or stamens. More importantly, when these flowers are examined closely, they have an outer whorl of sepals, as is normal, and then an inner whorl of sepals where we would expect petals. Then there is a whorl of carpels where we would expect stamens. Finally, there is the innermost set of normal carpels. Although such a flower has just two types of appendages, they are still arranged in four whorls. The important thing is that normal appendages are produced in the “wrong” regions (sepals in the petal zone and carpels in the stamen zone; FIGURE 14-36B). A mutation that causes an organ to develop in an unexpected site is called a homeotic mutation. Mutations in gene A result in carpels in whorl 1 (where sepals would be expected) and stamens in whorl 2 (where petals would be expected).
The ABC model has been so successful that it is now the basis for numerous experiments. Many plant geneticists induce mutations in well-studied species such as Arabidopsis thaliana and Antirrhinum majus (snapdragon), searching for more details about the ways in which these genes are controlled and how they work together. Because there are so many types of flowers, we expect to discover not only many versions of A, B, and C but also many other genes that subtly or dramatically influence A, B, and C. An example of modified flowers whose development still is unknown is those that consist of just stamens or carpels without any other appendages (FIGURE 14-37).
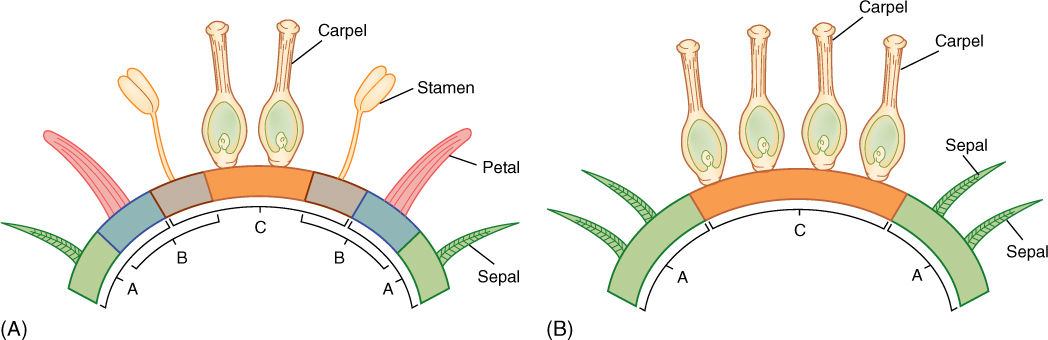
FIGURE 14-36 (A) The ABC model proposes that the outer part of a floral apical meristem is controlled by gene A, the inner part by gene C, and gene B overlaps the other two. Notice the outermost area is controlled only by gene A and produces sepals; the innermost is controlled only by gene C and produces carpels. Petals are produced by the zone where both A and B are active, and stamens occur where both B and C are active. (B) If a mutation inactivates gene B, then the floral apex has only two zones; the one controlled by gene A produces two whorls of sepals, and the one controlled by gene C produces two whorls of carpels. If gene B is inactivated, then the floral apex does not produce either petals or stamens.

FIGURE 14-37 (A) Each cluster on this slender, pendant stalk is an oak (Quercus) flower, consisting of just stamens (you are most likely to notice these staminate flowers in springtime, after they have been shed and are lying on sidewalks, streets, and cars). Elsewhere on the oak tree are flowers that consist of just carpels. (B) Cattails (Typha) produce two types of extremely reduced flowers: At the top are thousands of flowers that consist of just stamens, and at the bottom are flowers that consist of just carpels. Both types of flower are extremely simple, consisting mostly of either stamens or carpels without sepals or petals. A few hairs may be present. In this photograph the carpellate flowers have been pollinated and have developed into fruits; you are unlikely to ever see carpellate flowers of cattails unless you dissect them from the plant before they emerge above the leaves.
Plants and People
BOX 14-3 Environmental Stimuli and Global Climate Change
Global climate change is causing our world to rapidly become warmer and wetter, but plant mechanisms for detecting and responding to environmental stimuli are changing more slowly, if at all. As we burn oil, coal, and natural gas and as we convert forests into pasture for cattle, we increase the amount of greenhouse gasses in the atmosphere, causing the air, soil, lakes, and oceans to become warmer. As ocean temperatures rise, their surface waters evaporate faster, making the atmosphere more humid and increasing the amount of rain and snow that later fall on land. Temperatures do not increase uniformly everywhere; instead, circulation patterns in the atmosphere and oceans are affected, so some areas become warmer, others cooler, some wetter, others drier.
Changing climate will have profound effects on all plants, not only on those that respond to temperatures but also on those controlled by day length. Increasing temperatures affect two critically important events for temperate plants: The date of the last frost in spring occurs earlier, and the time of the first frost in autumn comes later. The frost-free growing season in many areas starts earlier and ends later: Plants have a longer growing season.
Plants that germinate or bud out solely based on temperatures can take advantage of this longer growing period, and many seem to be thriving. But for plants controlled by photoperiod, their critical night length does not change, they germinate or bud out at the same time in spring as they have for centuries, and they go dormant at their typical time in autumn. They are not able to take advantage of the extra days of warmth in spring and autumn; instead, they are dormant when they could be photosynthesizing, growing, and reproducing. And, just as bad, their respiration during dormancy is higher than before because it is controlled by environmental temperature: Not only are the plants not producing sugars photosynthetically as long as they could, they are now respiring away their carbohydrates faster. They will have less reserve nutrients available when they resume growth in springtime.
Now consider the interaction of photoperiodic plants and temperature-controlled plants. They occur together in the same habitat and compete with each other for water, minerals, room for their roots, and so on. As warm temperatures occur earlier in spring, the temperature-controlled plants get a head start over the photoperiod-controlled ones, and the same is true in autumn. It is likely that the photoperiod-controlled plants will suffer in this competition, and the ratio of the two types of plants in the ecosystem will change.
As temperature in general increases, the snow-free habitable zone in alpine areas gradually rises to higher elevations. Similarly, habitable zones are expanding northward in the Northern Hemisphere, southward in the Southern. Areas near the North and South Poles are more hospitable. Again, temperature-controlled plants may benefit from this: If their seeds happen to occur in the newly warmer areas, they should be able to grow and reproduce. But the same is only partially true for photoperiod-controlled plants. These should be able to grow higher on any mountain on which they exist already: The critical night length is the same up and down the mountain. But close to the poles, a few days at the beginning of summer (June 20 or 21) have sunlight for 24 hours: There is no night at all for a few days. And at the beginning of winter (December 20 or 21), several days have no sunlight. From early winter to early summer, day length increases from 0.0 to 24 hours. At the equator daylight always lasts for 12 hours and night is also 12 hours, all year long. Between these two extremes, days get slightly longer each day in lower latitudes and much longer each day in high latitudes. For plants that need very long days to bloom (for example, 17 hours, with 7 hours of night), that occurs in May or June in the northern part of the United States and southern Canada, but it occurs in March in northern Alaska and Canada. If a longer growing season would allow that species to grow that far north, its critical night length would occur too early (March) while the plant is still a seedling: It could grow in the new habitat but not reproduce there.
It is important to remember that plants and their control mechanisms do evolve. Hundreds of different plant species differ in their critical night length, and this variation came about through evolution by natural selection. The important question is whether these mechanisms will evolve rapidly enough to allow plants to adapt to the changing climate. We do not know the answer, but, in general, such evolution is slow and we are causing the climate to change rapidly.
 At the Next Level
At the Next Level
1. Regulatory cascades. When a hormone binds to its receptor, the first reaction is the activation of one or several enzymes, often by the addition or removal of a phosphate group. These enzymes activate others, which in turn activate others. Through the control cascade, one event—hormone binding—affects many different aspects of cell development. We know a great deal about control cascades, and there is much more that we do not know. This is a complex topic for independent study, so it is best to find one particular hormone you are interested in, or one particular development pathway, and focus only on those.
2. Perception of environmental signals. After many years of frustration, we are finally constructing a clear, unambiguous model of many types of signal perception in plants. Light signals (red, far-red, blue) have been useful because we can experimentally control color (wavelength) and intensity very precisely and we know that light must be detected by a pigment. I recommend light perception as a topic for independent study, but any aspect of signal perception will be rewarding. Just keep in mind that this is a very active area of research and many gaps remain, as there are many experiments in which the results are not well understood.
3. The Arabidopsis book. The American Society of Plant Biologists provides a wonderful free service to all botanists and other biologists by maintaining an online compilation of information about the model organism Arabidopsis thaliana (www.arabidopsisbook.org). This has illustrations, reviews, summaries, essays, and original research articles. It is updated regularly so it is worthwhile to visit the site frequently.
4. Control of the initiation of flowering. For almost a century, the term florigen was used as a code word for the unknown flowering messenger. It was known that for plants whose flowering is controlled by day length, the stimulus had to be given to a leaf, but the response would occur by inducing shoot apical meristems to change from leaf to flower production. It was assumed that “florigen” was a mixture of chemicals because no single chemical could be extracted from induced leaves that would have the desired effect when applied to a shoot apical meristem. In the last several years, it has finally been discovered that the messenger RNA for a gene called FLOWERING LOCUS (abbreviated FT) is either the flowering signal or the dominant part of a complex of several chemicals. Because this is a crucial step in reproduction, it is well worth independent study.
SUMMARY
1. It is selectively advantageous for organisms to be able to sense and respond to significant aspects and changes in their environments.
2. Communication between body parts of an organism is essential to the integration and coordination of the organism’s metabolism and development. Certain parts may need to respond to environmental or metabolic changes that they cannot sense themselves.
3. Plants must perceive important environmental information, transduce it to a communicable form, and respond to the transduced information.
4. Four ways in which plants respond to stimuli are tropic responses (oriented growth), nastic responses (stereotyped turgor changes), morphogenic responses (changes in quality), and taxis (oriented swimming).
5. In general, sites of perception are different from sites of response and must be linked by a means of communication. Presentation time and threshold are important elements in perception. Most communication appears to be by hormones.
6. Currently, the known classes of plant hormones are auxins, cytokinins, gibberellins, abscisic acid, ethylene, jasmonic acid, and several others.
7. The response to a hormone depends on which hormone is acting, the preparation of the responding cell, and the simultaneous or sequential presence of other hormones.
8. Flowering may involve the following steps: competence (“ripeness”) to be induced, occurrence of inductive conditions, sufficient health to produce flowers, and later stimuli to induce flowers to open.
9. Flowering and many other seasonal responses are controlled by night length in many species. Phytochrome is involved in measuring night length.
10. Plants contain endogenous rhythms, cyclic changes in their metabolism. The rhythms most frequently are circadian, having a period of approximately 24 hours. These rhythms affect numerous aspects of plant metabolism.
IMPORTANT TERMS
ABC model (of flowers)
abscisic acid (ABA)
adult phase
all-or-none response
apical dominance
auxin
blue-light responses
circadian rhythm
climacteric
critical night length
cytokinins
day-neutral plants
differential growth
differentiation
dosage-dependent responses
endogenous rhythms
ethylene
gibberellins
homeotic mutation
hormones
indoleacetic acids (IAA)
juvenile phase
long-day plants
morphogenic (morphogenetic) response
nastic response
negative feedback loop
pattern establishment mechanism
perception of a stimulus
phase change
photoperiod
phytochrome
polar transport (of auxin)
polarity
positive feedback system
presentation time
response to a stimulus
short-day plants
signal amplification cascade
statocytes
statoliths
symmetry
taxis
threshold (for a stimulus)
transduction of a stimulus
tropic response
vernalization
REVIEW QUESTIONS
1. Most plants are so large their bodies exist in several different environments. What does this mean? Give examples for a single plant.
2. For a stimulus to cause a plant to change its metabolism or development, there must be three actions: _____________ ________________, __________________________, and _______________________.
3. Before a plant can respond to environmental factors, they must be perceived. Give examples of several environmental cues that plants perceive.
4. After a plant perceives a stimulus and before it can respond, there is another step called _____________ ___________. What happens in this step?
5. Name five environmental factors that plants detect as stimuli. In each case, what types of information are provided to the plant?
6. Different prefixes are used for various stimuli. What is the prefix for each of the following:
a. Light
b. Gravity
c. Touch
d. Chemical
7. How does a positive tropic response differ from a negative tropic response? What is a diatropic response?
8. How does a nastic response differ from a tropic response? What is a morphogenic response?
9. In the perception of a stimulus, what are presentation time and threshold?
10. Responses can be either of two types: __________________-_________________-_______________ and __________________-_________________.
11. What is an all-or-none response? Give some examples of plants that flower with an all-or-none response.
12. What is a dosage-dependent response? Name some plants that flower this way.
13. What are the four ways that a plant can respond to a stimulus? Define and give examples of each. Why are some responses classified as “positive,” “negative,” or “plagio-,” whereas others are not?
14. Almost all plant communication is by a slow mechanism. What is the mechanism?
15. Name the known classes of plant hormones. What are the characteristics of plant hormones?
16. Name one natural auxin: _______________________.
17. The concentration of substances as powerful as hormones can be controlled by three methods in plants. What are the three methods?
18. Auxin, like many substances, can be transported through the phloem; however, there is a second mechanism that exists just for auxin transport. What is it?
19. Like auxin, cytokinins are involved in dozens of responses in all parts of the plant. Name one important response.
20. About how many gibberellins are known to exist? Are all of them active hormones?
21. What is the only gaseous hormone? How is it involved in fruit ripening?
22. What is apical dominance? How is auxin involved?
23. Describe the mechanism by which auxin appears to control leaf abscission.
24. Bending of plant parts toward or away from stimuli requires _______________ ___________________.
25. Studies of phototropism often use which part of an oat seedling: _________________?
26. True or false: Despite years of intensive research, we still do not have even an outline of the mechanism that actually causes plants to begin producing flowers.
27. Almost all plants must reach a certain age before they can be induced to flower. Before that point, they are said to be in the __________________ stage. Afterward, when they can be induced, they are in the ________________ stage.
28. Light is often the most important environmental factor for inducing a plant to flower, but it is not the intensity of light. What it the important aspect?
29. What are long-day plants? Short-day plants? Day-neutral plants? What is the critical night length?
30. Which pigment is responsible for detecting presence or absence of light? What are its two forms?
31. We know that short-day plants do not really need short days, but actually long nights. Describe the type of treatment that proves this. Is some kind of light given during the night or some period of darkness given during the day?
32. What is the difference between endogenous and exogenous rhythms? Give several examples of each type, and be certain to include several that have either short periods or long ones.
33. List eight metabolic processes that undergo endogenous rhythms in plants.
34. If an endogenous rhythm has a period of about 24 hours long, we say it is a _________________ rhythm.
35. Imagine a short-day plant (really a long-night plant) being given continuous darkness. If it is given a flash of red light at a time when its endogenous rhythm would be “expecting” daylight, will that red light inhibit flowering? What about if the red light is given at a time when the plant’s endogenous rhythm is “expecting” a long night?
Design Credits: Hummingbird: © Tongho58/Moment/Getty; Green plant cells: © ShutterStock, Inc./Nataliya Hora; Purple tulip: © ShutterStock, Inc./Marie C Fields; Dandelion: © ShutterStock, Inc./danielkreissl; Poppy: © ShutterStock, Inc./Saruri; Plant icon: © ShutterStock, Inc./Vector; Digging man icon: © ShutterStock, Inc./Z-art