Botany: An Introduction to Plant Biology - Mauseth, James D. 2017
Genetics and Evolution
Genetics

Chapter Opener Image: As this bee gathers nectar and pollen from these flowers of Nolina parryi, it is setting up thousands of experiments in genetics. It carries pollen grains (each with two sperm cells) from some plants and deposits them on the carpels (each with egg cells) of other plants. Because of the crossing-over that occurs in meiosis, none of the sperm cells is identical to any other in the alleles they carry, and each egg also differs from all others. Therefore, each seed that results from this bee’s activity will differ genetically from all other seeds, and some will be more adapted to this environment than others. Chances are good that those that are more adapted will grow and reproduce better than those that are poorly adapted.
OUTLINE
✵ Concepts
✵ Replication of DNA
✵ Mutations
- Causes of Mutations
- Effects of Mutations
- Somatic Mutations
- DNA Repair Processes
✵ Monohybrid Crosses
- Monohybrid Crosses with Incomplete Dominance
- Crossing Heterozygotes with Themselves
- Monohybrid Crosses with Complete Dominance
- Test Crosses
- Multiple Alleles
✵ Dihybrid Crosses
- Genes on Separate Chromosomes: Independent Assortment
- Crossing-Over
- Genes on the Same Chromosome: Linkage
✵ Multiple Genes for One Character
✵ Other Aspects of Inheritance
- Maternal Inheritance
- Lethal Alleles
- Multiple Sets of Chromosomes and Gene Families
Box 16-1 Botany and Beyond: Botanical Philosophy and Popular Culture
Box 16-2 Botany and Beyond: Whose Genes Do You Have?
Box 16-3 Alternatives: Genetics of Haploid Plants
LEARNING OBJECTIVES
After reading this chapter, students will be able to:
✵ Explain the advantage of sexual reproduction over asexual reproduction.
✵ Discuss DNA replication.
✵ State the importance of transposable elements in DNA mutations.
✵ Recall why DNA repair mechanisms are necessary.
✵ Describe the process of making and analyzing a monohybrid cross with selfing.
✵ Explain the purpose of a Punnett square.
✵ Discuss how a test cross can reveal a plant’s genotype.
✵ Describe a complication of performing a dihybrid cross.
✵ Compare independent assortment to linked genes.
✵ Give an example of epistasis.
✵ Compare biparental inheritance to maternal inheritance.
✵ Discuss triploid and tetraploid cells.
 Did You Know?
Did You Know?
✵ Plants inherit one set of chromosomes through the sperm cell in pollen and one set through the egg cell in the ovule.
✵ Plants are promiscuous: Each plant will have hundreds or thousands of flowers, and it is possible that pollinators will bring pollen from so many other plants that each seed has a different “father” (pollen parent).
✵ All the seeds produced by a single plant have the same “mother” (ovule parent), the plant itself.
✵ Domesticated crop plants have been interbred with each other for thousands of years, but most still have wild relatives that can interbreed with them.
![]() Concepts
Concepts
Genetics is the science of inheritance. The chemical basis of genetic inheritance is the gene, the sequence of DNA nucleotides that guides the construction of RNA and proteins and also controls construction of more copies of the genes themselves. If all plants of a species had exactly identical nucleotide sequences in their DNA, then all those plants would be identical physically, but virtually all genes occur in multiple forms known as alleles, the alleles of a particular gene differing from each other in their sequence of nucleotides.
Alleles arise by mutation; if the nucleotide sequence is changed (mutated) in any way, the new sequence is a new allele. Mutations can occur in any gene in any individual, and thus, gradually a population of separate plants comes to have a variety of alleles. The types of alleles that a single individual has are called its genotype, and the expression of those alleles in the individual’s size, shape, or metabolism is its phenotype (FIGURE 16-1). As a result of mutations, the population of individuals has varying genotypes and phenotypes. They are not identical, as is apparent from considering humans.
An important concept of inheritance is the selective advantage of reproduction, which may be either sexual or asexual. In asexual reproduction, each offspring is identical to its parent and siblings, having exactly the same DNA and thus the same alleles. Although this might seem like the safest, most efficient mechanism for producing large numbers of offspring to carry the parent’s genes into future generations, the progeny are never more fit than the parent, and no progeny is adapted to any environment other than the parent’s environment. During a drought, plants with poor capacity to withstand water stress are outcompeted by plants whose alleles confer superior water stress resistance. During an infestation of pathogenic fungi, individuals without alleles that confer immunity are destroyed. There may be no individual resistant to both stresses (FIGURE 16-2).

FIGURE 16-1 Flower color in four o’clocks (Mirabilis jalapa) is controlled in part by a gene that has two alleles. The DNA sequence of one results in a protein whose primary and tertiary structures cause it to synthesize red pigment. The DNA sequence of the other allele codes for a protein whose tertiary structure is misformed; it has no enzymatic activity, and no pigment is produced. Therefore, flowers are white. If only a small amount of pigment is produced, the flower is pink. Flower color is the plant’s phenotype; the type of alleles present is its genotype.
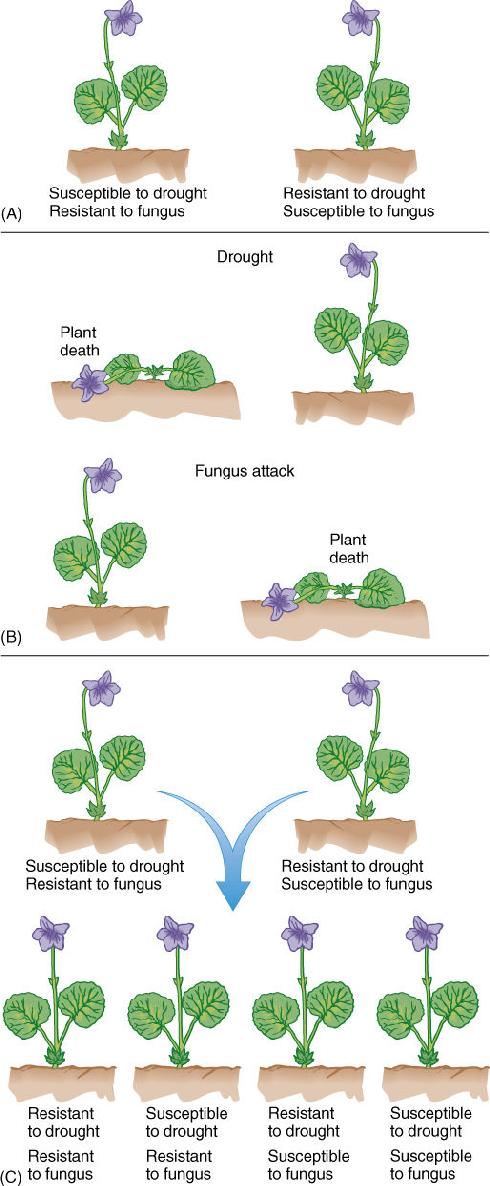
FIGURE 16-2 (A) A population probably contains a diversity of genotypes and phenotypes. (B) Without sexual reproduction, selectively advantageous alleles of one individual cannot be combined with those of another, and multiple stresses might kill all individuals. (C) With sexual reproduction, traits of one individual are combined with those of another, often producing individuals more fit than either parent.
Sexual reproduction is a mechanism by which an organism combines its alleles with those of other, possibly better adapted individuals, thereby increasing the probability that copies of its own alleles survive. If a plant is susceptible to drought, sexual reproduction is advantageous because some of its sex cells may fuse with those of a drought-resistant plant. If so, the new zygote should grow into an individual resistant to water stress, thus adding some protection to the alleles derived from the susceptible parent. Although the original plant may die during a drought, copies of its alleles survive in its progeny, protected by alleles from the resistant parent (Figure 16-2). The second parent should benefit as well, as it is almost certainly not superior in all attributes. The drought-sensitive parent may carry alleles for resistance to fungal attack, which, during times of fungal attack, protect the alleles of the drought-resistant parent in the offspring, making the progeny of this sexual reproduction resistant to both fungi and water stress.
Sexual reproduction involves a large degree of chance and risk. Fitness and survival capacity are not governed by just one or two genes, but by almost all genes, including those responsible for the proper construction of membranes, the functioning of organelles, the production and transport of hormones, and so on. One cannot say that any aspect is trivial and does not matter; therefore, the presence of one or two particular alleles in a sex cell is not the key feature, but rather the combination of all the alleles. Because of synapsis and crossing-over in meiosis, each plant produces thousands of types of sex cells, each with a unique genotype, which then fuse with the sex cells of many other individuals. For example, when plants bloom, they produce thousands or millions of pollen grains that are carried away and fertilize ovules on many other plants; simultaneously, its own ovules are receiving pollen from numerous plants (FIGURE 16-3). As a result, thousands of fertilizations may occur that involve this one plant’s alleles. Many may produce poor combinations of alleles that have little survival value, but at least a small percentage should have the best attributes of both parents and should produce healthy, genetically sound plants with a high capacity to survive all stresses. It is not necessary or even advantageous for all potential progeny to survive, only the fittest. The plants and animals around us today are the successful survivors of evolutionary experimentation.

FIGURE 16-3 Being diploid like most plants, this lupine (Lupinus) has one set of chromosomes from its paternal parent and one from its maternal parent, but the meiosis that precedes formation of each pollen grain and each ovule ensures that none of its many gametes is the same as any other. Insects will carry pollen to this plant from many other lupines, some of which may be better adapted than this plant is.
![]() Replication of DNA
Replication of DNA
Before a cell undergoes nuclear division, either mitosis or meiosis, DNA is replicated during S phase of the cell cycle. Replication doubles the amount of DNA, and each gene exists in at least two copies, one on each of the two chromatids, one of which goes to each daughter nucleus during anaphase.
As DNA replication begins, chromatin first becomes less compact, opening sufficiently to allow entry of the necessary replicating factors. DNA does not release from histones; instead, nucleosome structure remains intact. Next, one strand of the DNA double helix is cut, and the two strands separate from each other in a short region, forming a small “bubble” called a replicon (FIGURES 16-4 and 16-5). With the double helix open, free nucleotides diffuse to regions of single-stranded DNA and pair with its bases along both strands. These are ribonucleotides, not deoxyribonucleotides, and they are polymerized into short pieces of primer RNA approximately 10 nucleotides long. The primer RNAs then act as substrates for the DNA-synthesizing enzyme, DNA polymerase. It now enters and adds deoxyribonucleotides onto the end of the primer RNA using the open DNA as a guide. This method of replication, in which each strand of DNA acts as the template for making the complementary strand, is semiconservative replication because each resulting double helix contains one new molecule and has conserved one old one.
DNA polymerase adds deoxyribonucleotides only to the 3' end of the growing nucleic acid, and thus, one strand of the open DNA is copied in one direction and the other strand in the opposite direction (Figure 16-5: top strand toward the left, bottom strand toward the right; both are actually 5’ → 3’). As DNA continues to unwind and open in both directions, new pieces of primer RNA form and new pieces of DNA grow from them toward the existing fragments (called Okazaki fragments). The original primer RNAs are depolymerized, and new Okazaki fragments are joined to the first ones. On the other end of each strand, replication is continuous, one long chain forming from the original primer RNA.
Each chromosome contains only a single DNA double helix, and in each chromosome, hundreds or thousands of sites occur where replication is initiated. As DNA uncoils and then separates, it has a forked appearance; this region is a replication fork. As DNA continues to uncoil and open, replication forks at each end of each replicon travel along the DNA. Finally, each replication fork runs into one of the adjacent replicons; then all new pieces of DNA are ligated (attached to each other with covalent bonds) into two new, complete molecules. The large number of initiation sites allows numerous DNA polymerases to work simultaneously. Each is able to incorporate approximately 1,000 nucleotides every second into growing DNA fragments, and replicons extend at the rate of about 0.5 mm/min. Each replicon is about 45,000 to 180,000 base pairs long (45 to 180 kbp) and can be replicated in about 1 to 3 hours. Because S phase of the cell cycle is 3 to 10 hours long, only about one third to one tenth of the replicons are active at any particular moment. If only one DNA polymerase could act on each chromosome, S phase would require 40 to 60 days (TABLE 16-1).
As replication forks advance, DNA partially dissociates from the histone octamers, but as all enzymes migrate forward and the two new DNA double helixes are complete, they immediately reassociate with histones into complete nucleosomes. Some of the old histone octamers go to one double helix and some to the other, and new histone octamers are added to both. Any particular segment of DNA remains unpackaged for only a few moments.
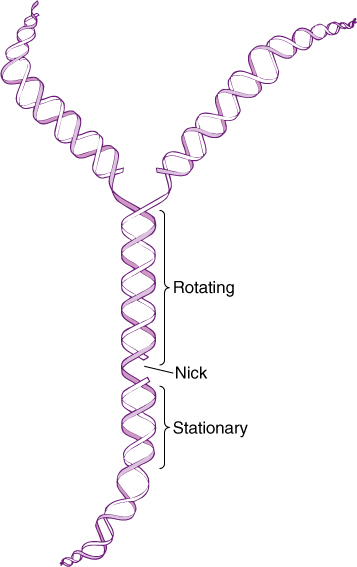
FIGURE 16-4 In order for DNA to unwind, one strand must be cut; otherwise, the entire unreplicated part would have to spin rapidly, which is impossible. With a single cut, only the region between the replicating enzymes and the cut must rotate.
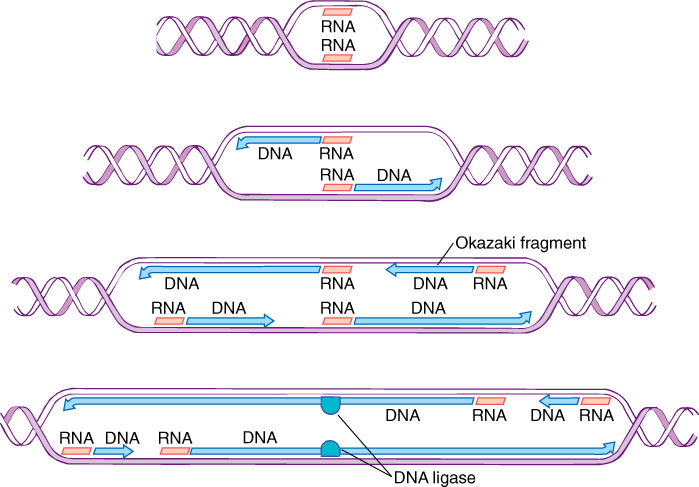
FIGURE 16-5 After a replicon opens, two pieces of primer RNA are formed; then DNA polymerase adds new DNA to the 3' end of the primers, and both strands elongate as the replicon unwinds at both ends. As the replicon continues to open, Okazaki fragments are initiated periodically; they grow to the downstream fragments, and the enzyme DNA ligase joins them into single molecules.
TABLE 16-1 Amount of DNA per Haploid Set of Chromosomes
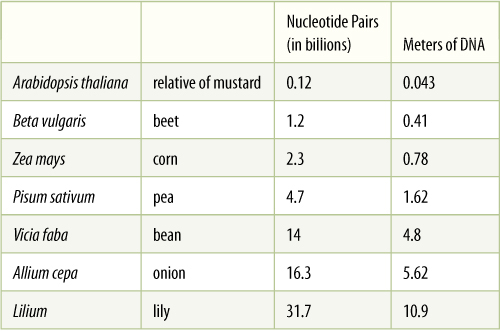
The values are known for only a few species at present, but more are being analyzed. Diploid nuclei have twice as much as shown in this table.
![]() Mutations
Mutations
A mutation is any change, however large or small, in DNA. The smallest mutation, affecting the least amount of DNA, is a point mutation in which a single base is converted to another base by any of various methods (FIGURE 16-6). If a piece of DNA is lost, the mutation is a deletion; the addition of extra DNA is an insertion. Under some conditions, a piece of DNA becomes tangled and breaks, and during repair, it is put in backward as an inversion.
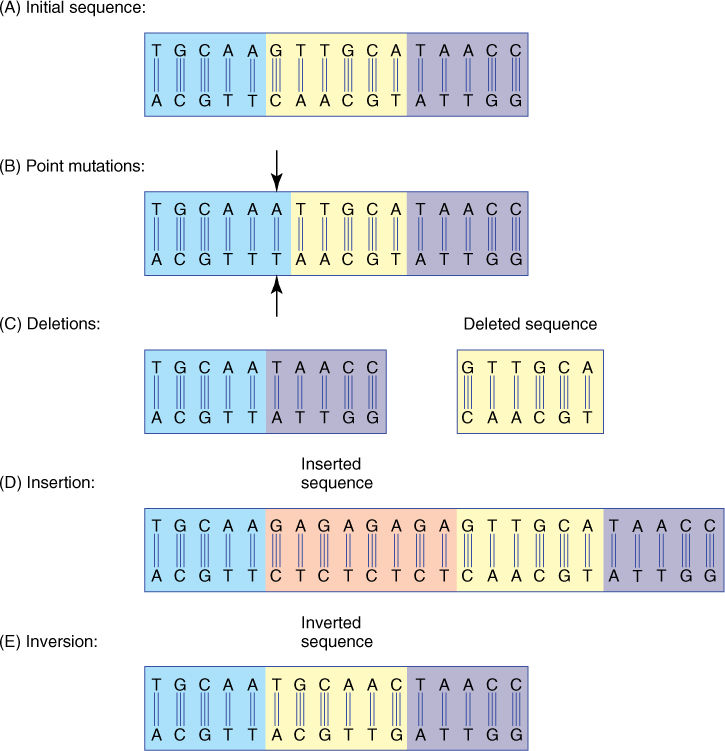
FIGURE 16-6 (A) Consider this initial sequence of DNA; the box is not important. (B) A point mutation consists of the change in just a single base pair—here G-C is converted to A-T. If this sequence is part of a gene rather than just spacer DNA, this change in sequence has resulted in a new form of the gene, a new allele. (C) The six base pairs in the yellow box of the initial sequence have been deleted. Deletions often remove hundreds of base pairs. On the upper strand, the right boundary of the deletion has removed a C and an A and left a T, changing CAT to AAT. The AUG mRNA start codon is coded by the DNA triplet CAT. Therefore, this may be an important mutation, destroying a start codon, but this may not be a triplet; the real triplets may have been GCA TAA or TGC ATA. The diagram provides too little information to predict the significance of the mutation. (D) An insertion is the addition of one or more base pairs. (E) In this inversion, the DNA has broken at two points and then flipped over and been reinserted; the original top strand of the insert is now the bottom strand.
Causes of Mutations
A mutagen is something that causes mutations. Several that are important are certain chemicals, ultraviolet light, X-rays, and radiation from radioactive substances. Many mutagenic chemicals are man made and are increasing in our environment. One mutagen, nitrous acid, reacts with cytosine and converts it to uracil, giving the DNA a G-U base pair in place of a G-C base pair (FIGURE 16-7). Numerous classes of DNA repair enzymes exist, one of which recognizes this G-U pair as abnormal and changes it back to G-C, but in meristematic cells that are replicating their DNA, DNA polymerase may arrive before the repair enzyme; if so, one new strand is formed with A complementing the U. In later replication, the A guides the incorporation of thymine, and the original G-C base pair becomes A-T.
Deletions can be caused in several ways; one method is due to short regions of a self-complementary sequence. As DNA unwinds ahead of the DNA polymerase, one strand with a self-complementary sequence may form a small loop. DNA polymerase may pass by this loop without reading any of the bases in it; the bases complementary to those in the loop are left out of the new DNA molecule, so it is shorter than it should be.
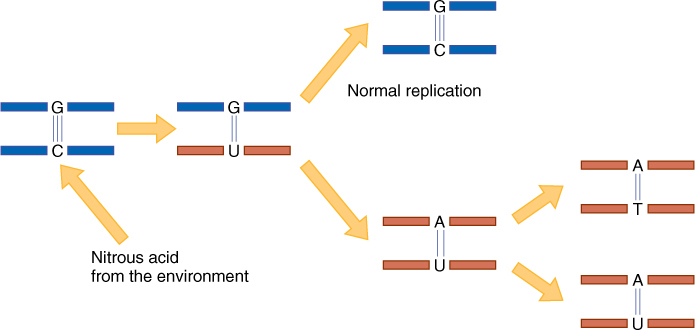
FIGURE 16-7 Nitrous acid converts cytosine to uracil. This strand is mutated, and all strands derived from it will be mutated, having A-T base pairs. The guanine is not affected and produces the original allele. One new allele has come into existence.
Insertion mutations can be caused by many methods because a variety of enzymes cut and rejoin DNA as part of repair processes. If a small piece of foreign DNA is present after cutting, it may accidentally be incorporated into the chromosomal DNA by DNA ligase.
Transposable elements are one of the most interesting and useful causes of insertion and deletion mutations. These are pieces of DNA that readily change their positions from one chromosome to another. Transposable elements have two basic forms—insertion sequences and transposons. Insertion sequences are only a few thousand base pairs long and contain the genes that code for the enzymes actually involved in cutting the insertion sequence out and splicing it into DNA somewhere else. A transposon is like an insertion sequence except that it may be much longer and carries genes that code for proteins not associated with transposition. The deletion and insertion mutations caused by transposable elements can vary in severity. If the element inserts into spacer DNA, the effect is not very important, but if it inserts into a gene, it totally disrupts either the promoter or the structural region. We suspect that transposable elements may actually be viruses, that they are a disease. Plants have mechanisms that fight them, but most plant nuclei contain transposable elements, sometimes in large amounts. More than half (58%) the DNA in every nucleus of Norway spruce (Picea abies) is transposon DNA; less than half of each nucleus is the plant’s own DNA.
Mutations caused by transposable elements may be one of our most powerful tools for genetic analysis and engineering. Often, a metabolic pathway is studied by exposing thousands of plants to mutagens such as X-rays or nitrous acid, then examining the plants to find any that have mutations disrupting the metabolic pathway of interest. These are then studied further to determine how the pathway was affected, but we never actually know which gene was disrupted. The gene is given a name (TABLE 16-2), but we do not know its DNA sequence, its location on a chromosome, or the nature of its promoter or structural regions. However, transposons are now being sequenced and engineered to contain markers such that the transposons can be applied to plants or cultured cells, where they cause insertion mutations. The mutants of interest are located (FIGURE 16-8), and DNA from the mutants is cut into pieces by restriction endonucleases, denatured, and combined with radioactive DNA complementary to the transposon. After the piece is found, it can be sequenced. Because we know the transposon’s sequence already, the DNA on either side of the transposon must be the gene of interest. Transposon mutagenesis offers the possibility of quickly correlating metabolic pathways, proteins, genes, and even promoters for virtually any aspect of an organism.
TABLE 16-2 Names of Genes in Pea Plants
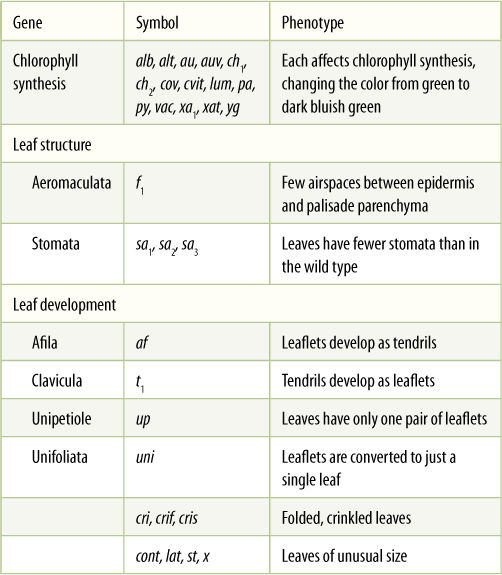
Although there are thousands of alleles in plants, only a few ever become obvious and only a small number of those are ever studied and given names. These are just a few of the named alleles in pea (Pisum).
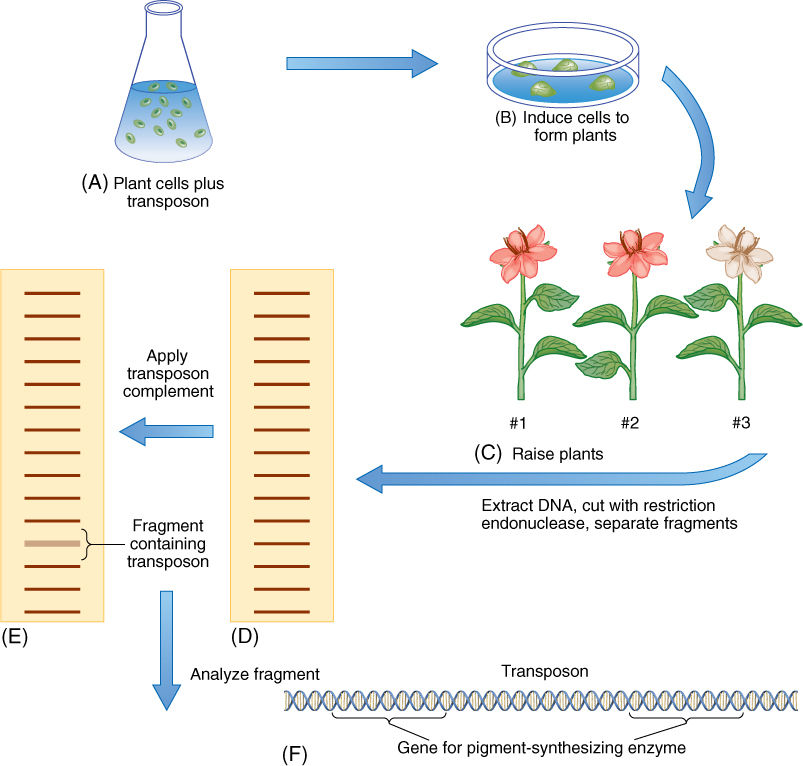
FIGURE 16-8 Transposon mutagenesis; the method is described in the text. In this case, flower pigment synthesis is of interest. Plant 1 is a normal plant. Plant 2 has a mutation for leaf shape, which is not our objective, and plant 3 has white flowers; thus, we have induced a mutation in some gene that is necessary for pigment synthesis.
Effects of Mutations
The effect and significance of a mutation depend on its nature, its position, and its extent. If it occurs in spacer DNA between two genes, it may have no effect whatsoever. Also, point mutations and small insertions and deletions in introns appear to be unimportant generally—they change a portion of hnRNA that will not be incorporated into mRNA. Within exons, a small mutation may not be important if it only changes a codon into another codon that specifies the same or a similar amino acid. Changing codons to ones that specify very different amino acids may not matter if they are located in a part that is not critical to the protein’s functioning.
On the other hand, a mutation in an exon may cause the gene to code for a protein whose active site is disrupted and the protein cannot function (Figure 16-6). An insertion mutation may cause the gene to code for protein so long that it cannot fold properly. Also, mutations in promoter regions can completely inactivate a gene or cause it to be active at the wrong time or place. Even point mutations can have profound effects; for example, nitrous acid conversion of a G-C base pair to A-T might cause the formation of a new start codon. Other simple changes can eliminate start codons, the recognition site for distinguishing the boundaries of exons and introns, TATA boxes, and so on. The larger the mutation is, the greater the probability that a critical part of the DNA is affected.
Statistically, mutations are almost always harmful. Enzymes tend to be approximately 300 to 400 amino acids long, and hundreds of trillions of proteins could possibly exist. Yet only a small fraction would be useful in living organisms. Any mutation that changes the structure of proteins, rRNA, or tRNA is more likely to produce a less useful than a more useful form. The majority are deleterious, the minority beneficial. Natural selection eliminates the deleterious mutations and preserves the beneficial ones.
Somatic Mutations
Mutations can occur at any time in any cell, but if they happen in cells that never lead to sex cells, they are called somatic mutations. For example, a gene in a leaf primordium cell may undergo a mutation, but because leaves are not involved in sexual reproduction, the mutation is somatic and is not passed on to the plant’s offspring, regardless of whether the mutation is advantageous or disadvantageous. When the leaf falls off in autumn, the mutated gene decomposes along with the rest of the leaf. The same is true of any mutation in roots, wood, or bark. In general, somatic mutations are not very important for most plants because they affect such a small portion of the plant and are not passed on to the offspring. A somatic mutation might not ever result in an altered phenotype. The mutation may occur in a leaf cell nucleus, but in a gene that is inactive in leaves, such as a gene that affects root hair growth or bark formation. In species that undergo extensive vegetative reproduction, such as blackberries, alder, and prickly pear cactus, somatic mutations can be important if they affect a part of a plant that gives rise to a vegetative offshoot. As the offshoot grows and reproduces vegetatively, the patch of mutated cells may finally include an axillary bud and then affect a flower and its sex cells. The mutation then becomes a sexual one.
DNA Repair Processes
Because most mutations are deleterious and occur frequently enough to be a significant problem, it is selectively advantageous for organisms to have DNA repair mechanisms that recognize and remove mutations. Certain mechanisms recognize base mismatches, loops, or other problems; other enzymes minimize the number of errors that occur in the first place.
The DNA repair rate must be neither too efficient nor too ineffective. In organisms that have very short genomes, such as prokaryotes and perhaps algae and protozoans, DNA polymerases can replicate a full genome without errors. Perfect replication does not occur every time, but nonmutated replication can sometimes occur. With the larger genomes of more derived animals and plants, one set of chromosomes can virtually never be replicated without mutations if no repair mechanisms are present. DNA polymerase may make an error only once in every 1 million nucleotides, but if the genome is several billion base pairs long, every replication produces numerous errors. Most angiosperms have about 10 to 100 billion base pairs per haploid set of chromosomes (we humans have only 3 billion base pairs). One of the smallest numbers known is in Arabidopsis, with about 120 million base pairs (containing an estimated 25,500 genes); every round of replication before a mitotic or meiotic division results in about 120 errors (Table 16-1).
As a zygote grows into an adult, every cell cycle introduces new mutations, and in the adult, no two nuclei are exactly alike; after several cell cycles, DNA would be useless if there were no repair mechanisms. It would be impossible to produce sperms or eggs that were not extensively mutated, and under such circumstances, complicated organisms could not exist. It is estimated that a minimum of 15,000 genes are needed to code for all the information required for a flowering plant. This is about 20 million base pairs, so DNA polymerase is not accurate enough to provide error-free replication for even the simplest angiosperm, but DNA proofreading and repair systems bring the error rate down to an acceptable level. Actual rates as low as one mutation per 500,000 genes have been measured in corn.
Mutations occur in the genes that code for repair enzymes, resulting in serious problems. In humans, the disease xeroderma pigmentosum is caused by an inability to repair mutations caused by ultraviolet light. People with this disease are sensitive to sunlight and develop skin cancers easily. Sunlight has the same effect on all of us, but most of us can repair the damage. Our bodies have mechanisms that repair other types of mutations as well, but our modern chemical society may be contaminating our environment with mutagens for which we have no repair mechanisms.
DNA repair mechanisms must not be perfect or even extremely efficient. If every replication were perfect and if no mutations ever arose, all cells of an individual would have absolutely identical nuclei, as would all sperm and egg cells. Sexual reproduction would be useless because all eggs and sperms of a species would carry identical genes. Mutations must occur at a low enough rate that they do not endanger every individual but rapidly enough that a species evolves as its environment changes. With no variation, there would be no differences for natural selection to act on; if all are identical, none has a selective advantage.
![]() Monohybrid Crosses
Monohybrid Crosses
Sexual reproduction between two individuals is called a cross. The meiotic divisions that precede a cross reduce the number of sets of chromosomes per cell from the diploid number to the haploid number. Consequently, each sex cell—that is, each sperm cell and egg cell—contains one complete set of genes. Furthermore, each sperm cell contains all the genes necessary to construct a new plant; the same is true of each egg cell. The zygote (the fertilized egg) has two complete sets of genes.
Within a population, mutations produce new alleles, and the genotypes of individuals within the population differ. Of the plants that grow in an area and that can interact sexually, many may have the same allele of a particular gene, but other individuals may have other alleles, other versions of the gene. Consequently, the alleles carried by a particular sperm cell may or may not be identical to the homologous alleles of the egg it fertilizes.
Monohybrid Crosses with Incomplete Dominance
In a monohybrid cross, only a single character is analyzed and studied; the inheritance of other traits is not considered. For instance, a plant with red flowers might be crossed with one that produces white flowers, and only the inheritance of that flower color trait is studied. Characters involving flower shape, leaf structure, and photosynthetic efficiency are also being inherited simultaneously, but in a monohybrid cross, only one is studied. This makes the analysis and understanding of the results much simpler. After basic principles of a particular trait are known, then its interaction with another factor (dihybrid cross) or two other factors (trihybrid cross) can be studied. Mendel gave his attention to monohybrid crosses, which are easy to understand; before him, people tried to analyze many characteristics at a time and became hopelessly confused.
Consider the cross just mentioned: A plant with red flowers is bred to one with white flowers. It does not matter which flower produces pollen and which produces ovules. After the cross is made, seeds and fruit develop; the seeds are planted, and when mature, the new plants are allowed to flower so that their flower color can be examined. The flowers of all plants in this new generation are pink, resembling each parent somewhat, but not exactly like either (FIGURE 16-9). The parents are called the parental generation; the offspring of their crossbreeding are the F1 or first filial generation, and if these interbreed, their offspring are the F2 generation.
The molecular biology of this monohybrid, flower color cross is easy to understand. Each parent is diploid and thus has two copies of the gene involved. In the red-flowered parent, both alleles produce mRNA that is translated into functional enzymes involved in synthesis of red pigment. In a white-flowered parent, both alleles are defective. It may be that each produces an mRNA that when translated results in a protein unable to perform the necessary reaction, or the promoter region may be mutated and can no longer interact with a chemical messenger. Whatever the cause, there is no pigment, and the flower is white.
Using genetic symbols, the red-flowered parent is RR and the white-flowered one is rr (Figure 16-9). Each parent is said to be homozygous because each has two identical alleles for this gene. The pink-flowered F1 has received an R allele from one parent and an r allele from the other, so its genotype is Rr; it is heterozygous because it has two different alleles for this gene. We use these symbols even though we have neither isolated the gene nor analyzed its nucleotide sequence; R and r are simply labels. Most genes are known only by their phenotypes and the labels given to them by geneticists (Table 16-2). With a genotype of Rr, the plant produces mRNA, half of which carries the defect; thus, only half the normal amount of enzyme is produced. This results in less pigment being formed, only enough to make the flower look pink, not red. Neither parental trait dominates the other, so this pair of alleles shows incomplete dominance: The heterozygous phenotype differs from both homozygous phenotypes.
When analyzing the possible outcomes of crosses and breeding, one must understand the types and quantities of gametes involved. All of the plants we are considering are diploid and form haploid spores by meiosis. The RR parent has chromosomes as shown in Figure 16-9B, and regardless of how chromosomes separate during meiosis, all spores receive an R allele. In the rr parent, all spores receive an r allele. Spores are not sex cells, but as they develop into gametophytes, they divide by mitosis—duplication division—so all cells of the microgametophyte (pollen) have the same allele, as do all cells of the megagametophyte (inside the ovule). Because the RR parent produces only R spores, it also produces only R sex cells, both sperms and eggs in typical bisexual flowers. Similarly, the rr parent produces, indirectly, only r gametes. When the two plants are interbred, an R gamete unites with an r gamete, establishing a heterozygous (Rr) zygote that grows into a heterozygous adult by means of mitotic cell divisions. No other outcome is possible; it does not matter which gamete is which, an R sperm and an r egg result in the same type of zygote as an r sperm and an R egg.
When the heterozygote matures and flowers, each spore mother cell produces two types of spores (FIGURE 16-10), not just one as is true of a homozygote. Because each cell has one R allele and one r allele, during the first meiotic division, one daughter cell receives R, and the other receives r. The second division of meiosis results in two R spores and two r spores. If this is occurring in the anther, half the pollen grains and hence half the sperms have R, and the other half have r. In the nucellus of many plants, only one megaspore survives. If this gene has no effect on spore metabolism (and a gene for flower color is probably inactive in spores), then half the time R cells survive and half the time r cells live. In heterozygote parents, the two types of sperms and eggs are produced in equal numbers.
Crossing Heterozygotes with Themselves
When a plant’s own pollen is used to fertilize its own eggs, the cross is a selfing. A plant can also be selfed by being crossed with another plant with exactly the same genotype. Selfing heterozygotes has interesting, instructive consequences; 50% (on average) of all sperms and eggs contain the R allele, and 50% have r (Figure 16-10). Not all zygotes are identical genotypically: Some are RR, having resulted from an R sperm and an R egg; others are rr (r sperm and r egg), and some are Rr (either R sperm and r egg or r sperm and R egg). Selfing a heterozygote produces three types of F1s, some of which (Rr) resemble the parents and others (RR and rr) the grandparents. Again, to analyze the results, we must wait for the zygote to develop into an embryo, then plant the seeds and wait until the new plants are old enough to flower. Because each genotype produces a distinct phenotype, the genotype of each plant is known simply by looking at the flowers. If a large number of heterozygotes (pink-flowered plants) are selfed and large numbers of F1 plants grown, approximately one fourth are red (RR), one half pink (Rr), and one fourth white (rr). This is an important ratio, typically represented as 1:2:1, and should be memorized immediately.
The reason for the proportion of these genotypes is explained in FIGURE 16-11. A Punnett square can be set up in which all types of one gamete, say, the egg, are arranged along the top of the square, and all types of the other gamete are arranged on the left side. The boxes are then filled in with the allele symbol above it and to the left. Because the gametes are produced in a ratio of 1 R: 1 r, listing them as in Figure 16-11 automatically represents their relative numbers in nature.
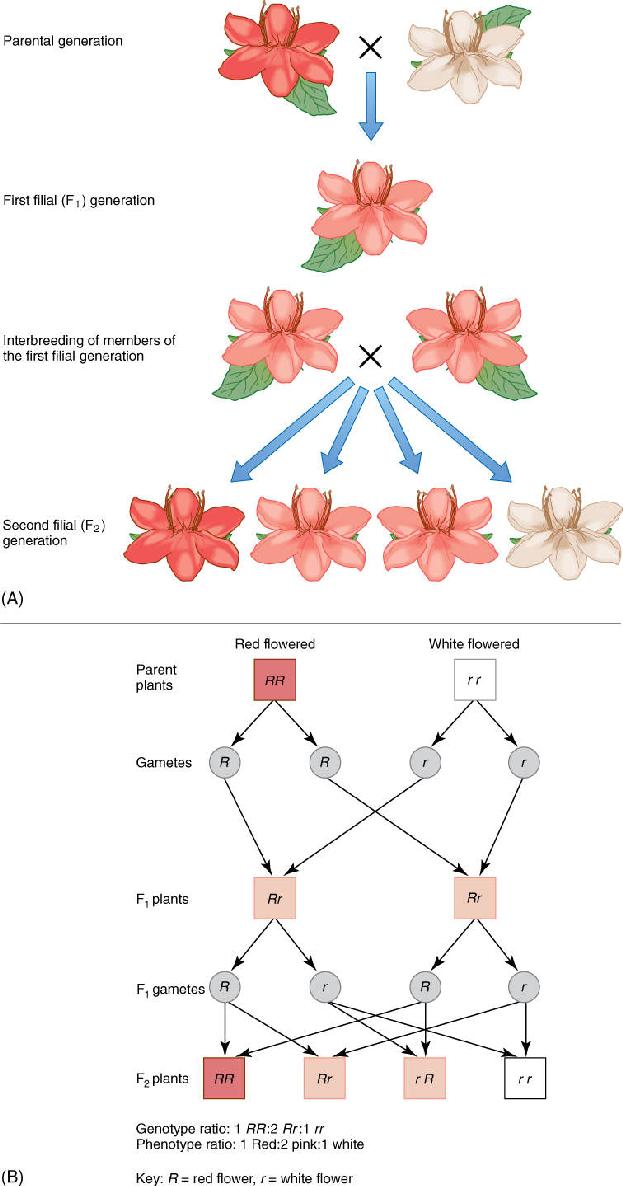
FIGURE 16-9 (A) A monohybrid cross analyzing the trait of flower color. Details are explained in the text. (B) The phenotypes of parents, gametes, and progeny.
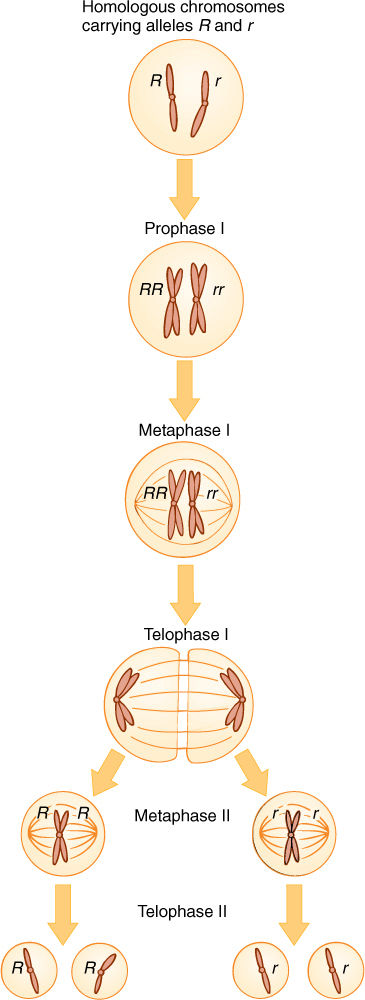
FIGURE 16-10 Production of haploid cells by meiosis in a heterozygote. Homologous chromosomes pair (synapse) during metaphase I, then the paternal chromosome is separated from the maternal chromosome during anaphase I: One type of allele is separated from the other. As sister chromatids separate after metaphase II, each daughter nucleus receives identical alleles.
The Punnett square does not represent the outcome of any one cross; if a single heterozygous flower is selfed, it may produce only one or two seeds. If you plant just one seed, there is one chance in four that it will have red flowers, two chances in four pink, and one chance in four white.
The 1:2:1 ratio was one of the great discoveries of Gregor Mendel (1822—1884), an Austrian monk who performed experiments with pea plants. His cross-breeding experiments became the basis for modern genetics. Mendel discovered that in a selfing of this type, the recovery of the parental types means that genetic material must be composed of particles, such that the R genetic material can be separated from the r genetic material in the Rr heterozygote. Before Mendel’s work, it was thought that the genetic material was a fluid. Two fluids cannot mix and then separate again perfectly. The constancy of the 1:2:1 ratio made it more logical to think in terms of discrete particles, genes, that never lost their identity regardless of the crosses in which they participated.
Furthermore, the 1:2:1 ratio can be realistically interpreted only in terms of each individual plant having two copies—being diploid—and each sex cell having one copy—being haploid. If each plant had only one copy, pink-flowered heterozygotes would be impossible, whereas if each had three, phenotypes such as dark pink (RRr) and light pink (Rrr) should also be present. Keep in mind the state of scientific knowledge in 1865 when Mendel was working. The concept that all organisms are composed of cells with nuclei had only recently been proposed; mitosis and cell division were very poorly understood, and meiosis would not be discovered for another 23 years. The concepts of chromosomes and homologous pairs would not be well established until the 20th century, and the existence of mRNA was not confirmed until the mid 1960s (BOX 16-1).
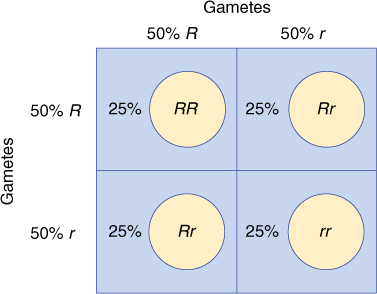
FIGURE 16-11 A Punnett square makes it easy to analyze and understand the results of a cross. Gametes from one parent are listed across the top; those of the other parent are listed on the left side.
Botany and Beyond
BOX 16-1 Botanical Philosophy and Popular Culture
When thinking about genetics and the contributions of Gregor Mendel, it is important to keep in mind the state of scientific knowledge at that time. Some of the greatest scientific geniuses of all time were discovering and documenting the fundamental concepts of biology. Only 27 years earlier than Mendel’s work, M. J. Schleiden and T. Schwann had proposed the cell theory, the concept that all organisms are composed of cells with nuclei and that all existing cells come from preexisting cells. We take this for granted today, but it means that all living organisms are fundamentally the same, that life may have arisen only once, and that all cells are descended from those first cells. In 1849, Wilhelm Hofmeister proved that during plant reproduction the new embryo develops primarily from the egg after receiving some “influence” from the pollen tube. This was revolutionary because until that time everyone firmly believed that in both plants and animals, the new embryo developed from the sperm cell—the female parent might provide nourishment and protection, but the new generation was really a continuation of the paternal parent (could this belief have been based not on observation of nature but on observation of society and history dominated by males?). Hofmeister’s conclusion that the female parent also contributes to the new offspring was both shocking and also very important for Mendel’s later observation that genes are passed equally from both parents.
In 1859, Charles Darwin published Origin of Species by Natural Selection, which postulated that organisms change through time, that they are not constant (A. R. Wallace came to the same conclusion at the same time). The discovery of evolution by natural selection caused a great deal of trouble for Mendel: Natural selection requires diversity and change, whereas Mendel’s theories of genetic inheritance showed that differences in phenotype were due only to the mixing and separating of unchanged, constant genes. The idea of evolution had aroused great controversy and serious philosophical battles between proponents of evolution and of biblical creation. This battle had been raging for 6 years when the paper by Mendel (a monk) appeared. Had he published 10 years earlier, he might have found instant acceptance, and Darwin would have had a more difficult time.
A further difficulty for genetics, however, was its postulation of genes as particles, not as fluids. This was at a time when fluids were considered the basis for most of biology, with sieve pores in the phloem and pits and perforations in the xylem considered to be effective by acting as filters that separated the various fluids that were thought to exist. Fortunately, in 1879, H. Fol showed that during angiosperm reproduction, a sperm nucleus (a particle) entered the egg, and in 1888, E. Strasburger described meiosis, giving a firm foundation to the concept of genes being particulate. If genes are on the chromosomes, then suddenly it is possible to see how Mendel’s ratios and independent assortment could occur. This was summarized in 1915 by T. H. Morgan in The Mechanism of Mendelian Heredity. Finally, in 1927 and 1928, H. J. Muller and L. J. Stadler showed that genes could be artificially changed (mutated) by the use of X-rays. Thus, genes could be stable enough to permit Mendelian genetics to operate and yet variable enough to allow natural selection to operate as well. It is important to keep in mind that these were not the only biologists working at the time, nor were these the only discoveries. A huge volume of information was being generated, much of which concerned unusual, exceptional, or aberrant cases and which thus made the fundamental discoveries more difficult to recognize as being important. Also, we must always try to appreciate the profound influence of the general philosophy of the time—a male-dominated society produced male-dominated theories of biology. It is likely that our own sincerely held beliefs are causing us to improperly evaluate some of our own biological observations.
Monohybrid Crosses with Complete Dominance
The situation in which only half as much product of an enzyme, such as the red pigment discussed above, is produced in a heterozygote is not universal. In certain species or with other traits, cytoplasmic control mechanisms may cause the enzyme to function until a specific amount of product is synthesized. The enzyme may have to work faster or longer, but the final amount of product is the same whether the plant has two functional alleles or only one. In other situations, the amount of enzyme might be monitored such that the nonmutant mRNA of the heterozygote is translated more frequently or rapidly.
In either case, the phenotype of the heterozygote is like that of the parent with two effective alleles. That trait is said to be dominant over the other version of the trait, which is recessive. An example is height. A tall plant with a TT genotype produces sex cells that are all T. A short plant, genotype tt, produces only t sex cells (FIGURE 16-12). When the tall plant is crossed with the short one, all F1 progeny have the Tt genotype, but they all have the phenotype of being tall, indistinguishable from their TT, homozygous dominant parent. We do not know what protein is produced by the T allele, but even with only one functional allele, enough product is made to permit normal growth. Tt plants are tall, and the tall character completely dominates the “dwarf” character.
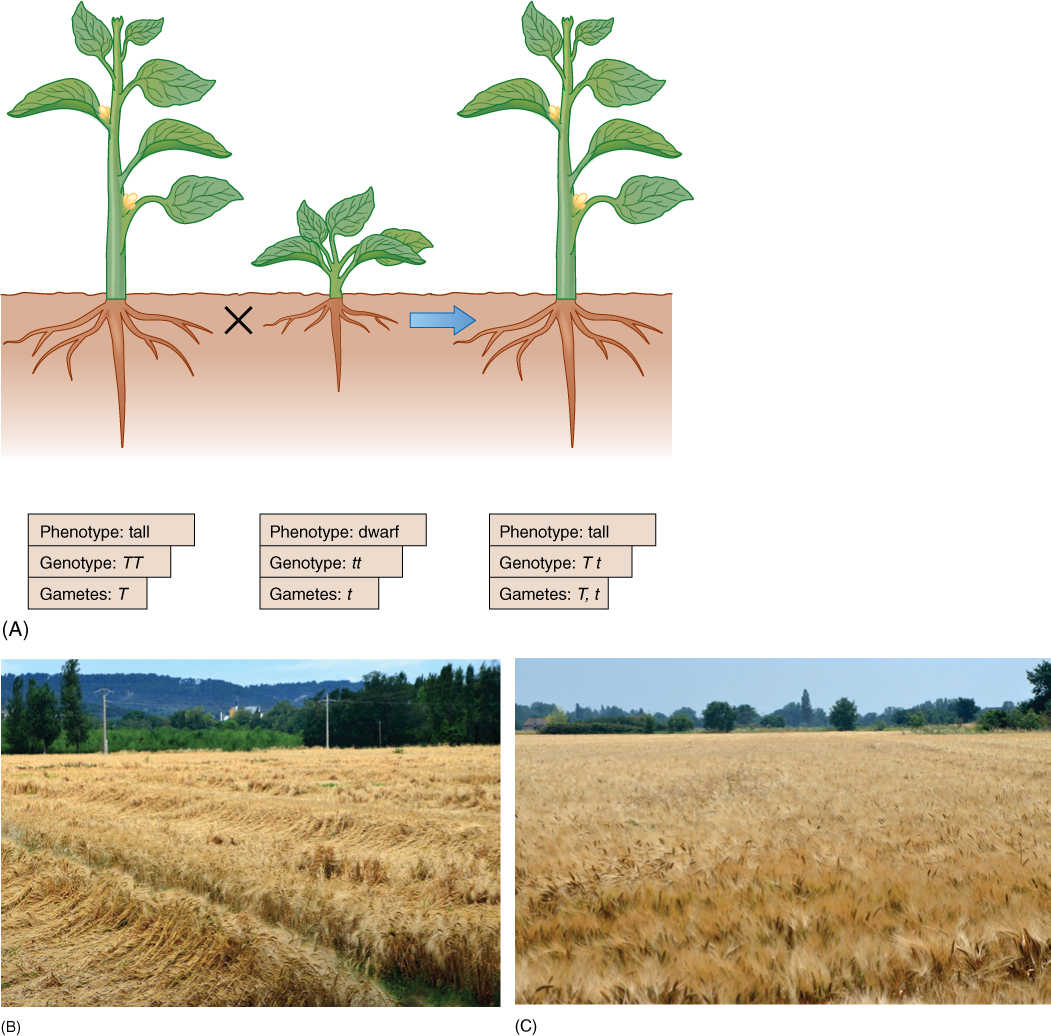
FIGURE 16-12 (A) In this monohybrid cross, the trait “tall” shows complete dominance over the trait “dwarf.” In a Tt nucleus, the T allele may be transcribed twice as much as each T allele in a TT nucleus. The resulting mRNA may be translated twice as much, or the protein may work twice as long or twice as fast. A TT plant may contain a level of T protein far above the threshold for responsiveness. (B) Most of the wheat plants in this field have blown over and are lying on the ground (we say the wheat has “lodged”). This is a tall variety of wheat: Its stems are too tall and slender to be strong enough to hold themselves upright. The harvesting machine (a “combine”) will have to cut the wheat so low to the ground that dirt and rocks may be brought into the machine, damaging it. Also, if it rains before harvest, the stems lying close to the soil may rot. (C) All the wheat plants in this field are still upright and will be easy to harvest; none of these plants has lodged. This is a short variety of wheat: Plant geneticists obtained the seeds for this crop by crossing plants that all had short, strong stems. This field can be harvested easily, and there is little chance any of the wheat will rot, even if it rains before harvest time.
Knowing the molecular biology of genetic systems, you can predict that when heterozygotes are selfed, two types of sperm cells (T and t) and two types of eggs (T and t) are produced. The Punnett square for the cross is as in FIGURE 16-13, and a genotype ratio of 1 TT:2 Tt:1 tt is expected. The prediction is correct, but what will the phenotype ratio be? One out of four plants will be tall due to a TT genotype, and two out of four will be tall due to a Tt genotype. Thus, three fourths have the tall phenotype, and one fourth have the short phenotype. Whenever a cross is made and a phenotype ratio of 3:1 is seen, we should suspect that the parents are heterozygous and the character shows complete dominance.
Test Crosses
Remember that we can only see an organism’s phenotype; to discover its genotype, we must perform crosses, and the test cross is one of the most useful. When traits with incomplete dominance are studied, the genotype of any plant is easy to determine from its phenotype. If the trait has complete dominance, it is difficult to know what the genotype of any particular plant is unless the plant shows the recessive trait. Imagine that you are studying the inheritance of tallness; you have selfed some heterozygotes and planted the resulting seeds. In your greenhouse or garden, there are now hundreds of plants, approximately 75% of which are tall and 25% short. You know the short ones are tt, and if you need a plant with the tt genotype for experimentation, you know automatically to choose short plants. But if you need to experiment on plants with the TT genotype, how can you tell which they are? A tall plant picked at random is more likely to be a Tt plant because there are twice as many of them as TT tall plants.
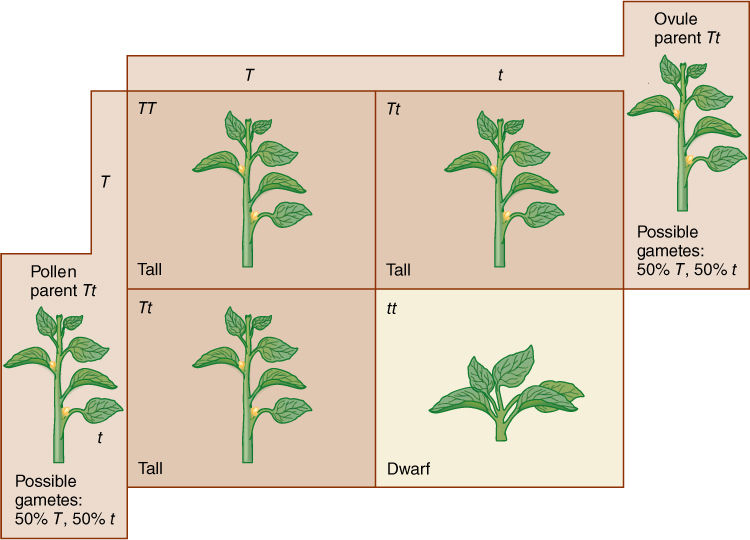
FIGURE 16-13 In setting up the Punnett square for a selfing of Tt plants, first establish the genotypes of the two parents. Then determine what types of gametes are produced and in what proportions, and fill in the squares with the genotypes. From the genotypes, the phenotypes in each square can be determined.
The genotype can be revealed by a test cross, a cross involving the plant in question and one that is homozygous recessive for the trait being studied. All gametes produced by a homozygous recessive parent carry the recessive allele, which is unable to mask the homologous allele in the resulting F1 zygote. If the plant being tested is actually homozygous dominant (TT), 100% of the progeny will be heterozygous (Tt) and tall (FIGURE 16-14). If the tall plant is heterozygous, half its progeny in the test cross will be tall (Tt) and half short (tt). If the cross produced thousands of seeds, you might not want to plant all of the seeds, just some, and if a large enough number of seeds are grown, the chance of accidentally not choosing any of the Tt seeds is very low, and if even one of the F1 plants is short, the test parent must have been Tt.
After the actual genotypes of plants are known, those that are homozygous dominant can be gathered and planted in special areas, kept free of all natural pollinators, and allowed to breed among only themselves. All of their progeny will be homozygous dominant and can be used in breeding experiments. The homozygous recessives can also be kept as a special line, being selfed and kept pure. Such groups are pure-bred lines and are both useful and valuable. It is not possible to maintain the heterozygotes like this because they do not breed true; that is, their progeny are not exactly like them. One fourth are homozygous recessive; one fourth are homozygous dominant and cannot be distinguished visually from the one half that are heterozygotes.
So far, we have considered only one trait, height, but for plants that are important crop or horticultural species, dozens or even hundreds of traits are cataloged. Seed companies maintain hundreds of different lines of corn, for instance, in which the genotype of many characters is known for each line (FIGURE 16-15). Many times, plants are collected from the wild, so nothing is known about their genotype unless it is immediately obvious from the phenotype. From looking at a few collected plants, however, it is not possible to tell which characters are dominant and which are recessive; carefully controlled and recorded crosses must be made.
When test crosses must be made on annual plants, the results are usually not known until after the plants have died. Their genotypes are then known, but the plants cannot be used for experimentation or breeding. In such cases, it usually is necessary to do both test crosses and experimental crosses simultaneously, not knowing which plants have the correct genotype for the experiment being performed. After the test cross results are complete, the parents that had the proper genotype can be identified, and the experimental crosses that involved them can be analyzed.
Multiple Alleles
Each gene may have many alleles, not just two as in the examples discussed so far (T and t, R and r). A protein of average size consists of about 300 amino acids, so the coding portion of its mRNA must have about 300 codons, each containing three nucleotides. The gene is therefore at least 900 nucleotides long, not counting introns and promoters. At least 900 sites exist at which point mutations can occur, and of course, any mutation may involve several nucleotides. Consequently, the gene may exist in many forms, called multiple alleles. When genes are polymorphic, having multiple alleles, numbers, such as X1, X2 , X3 , X4 , and so on, are used rather than capital and lowercase letters. Certain mutations still result in the production of a protein with the normal sequence, but most lead to altered protein structure. Some of these proteins are similar to the original protein, perhaps having similar or even identical enzymatic activity. However, many carry out the proper reaction more slowly or are not accurately controlled by regulatory mechanisms in the cytoplasm; a normal, wild-type phenotype may not be produced. With multiple alleles, the concept of dominance is more complex; one allele may produce the proper quantities of the functional protein, whereas a different allele produces more, another produces less, and a fourth produces one with altered activity. Many different phenotypes are possible.
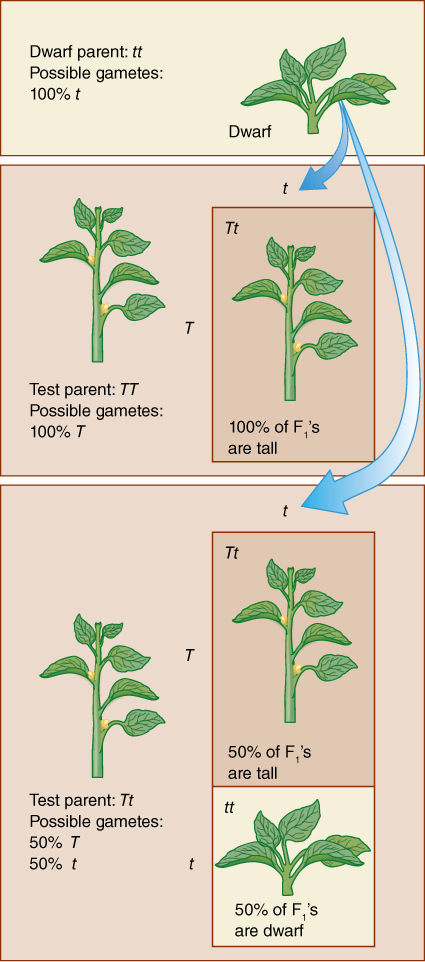
FIGURE 16-14 Test crosses to determine whether a plant with the dominant phenotype is heterozygous or homozygous. Of the data in this diagram, you would not know the genotypes of the test parents before the test cross; those data are what you are trying to discover. If you want to find only a single homozygote or heterozygote, you would need to do test crosses on only one or a few plants. Imagine, however, that you had done an experimental cross for a trait that you assumed would give you a 3:1 ratio; it would be necessary to do test crosses on a large number of progeny plants with the dominant phenotype just to confirm your experiment.
Within a population of plants, as many types of gametes can be produced as there are different types of alleles. A heterozygous X1X2 plant can be crossed with a heterozygous X3 X4 one, resulting in progeny such as in FIGURE 16-16: X1 X3 , X1X4 , X2 X3, and X2 X4. Four distinct types of F1 plant are produced, none of which has the genotype of either parent.

FIGURE 16-15 Maintaining pure-bred lines is relatively easy for perennial plants; after the plant is growing, it can produce pollen or ovules for experimentation for years. Annual plants are much more difficult because pollination, seed gathering, planting, and record keeping must be done annually. Here, wheat seeds are being frozen in liquid nitrogen; after several years, some will be thawed and planted. Their seeds will be frozen for further storage.
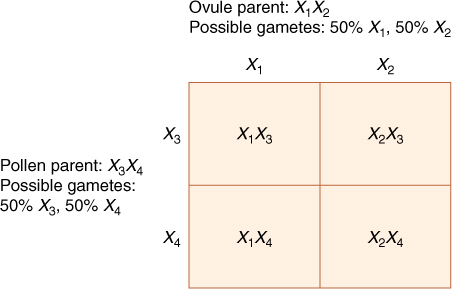
FIGURE 16-16 With multiple alleles for a single character, numerous types of crosses become possible; however, we still determine, from the parental genotype, all possible gametes and then construct a Punnett square in the usual fashion.
![]() Dihybrid Crosses
Dihybrid Crosses
A dihybrid cross is one in which two genes are studied and analyzed simultaneously, rather than just one, as in a monohybrid cross. Every cross involves all the genes in the organism, but the terms “monohybrid” and “dihybrid” refer only to the number being analyzed.
When two genes are studied, the results of the crosses depend on the positions of the genes on the chromosomes. If they are on different chromosomes, the alleles for one gene move independently of the alleles for the other gene, but when two genes are close together on the same chromosome, the alleles for one gene are chemically bound to the alleles for the other gene and move together. The situation in which the genes are on separate chromosomes is easier to understand and is explained first.
Genes on Separate Chromosomes: Independent Assortment
Consider a plant heterozygous for two traits, for instance seed coat texture and color, with a smooth seed coat (S) showing complete dominance over a wrinkled seed coat (s), and a yellow seed coat (Y) being dominant over a green seed coat (y). The plant’s genotype is SsYy, and its phenotype is smooth yellow seeds. We know that if we consider only the gene for color, the plant produces two types of gametes in approximately equal numbers, some carrying Y and some carrying y. Also, if we consider only texture, half carry S and half s. How do the alleles of the two genes relate to each other? As in monohybrid crosses, knowing the types of gametes that can be formed is the key to understanding the patterns of inheritance.
If the two genes are on separate chromosomes, the alleles of one gene move independently of the alleles of the other gene; this is called independent assortment. The chromosomes have duplicated during S phase and each has two copies of each allele (FIGURE 16-17A); all chromosomes align on the metaphase plate during metaphase I, where homologous chromosomes also pair with each other. There are two Y alleles, two y alleles, two S alleles, and two s alleles at the metaphase plate. During anaphase I, homologous chromosomes separate from each other, and both Y alleles move to one pole because they are on the two chromatids of one chromosome, still held together by its centromere. Both y alleles, located on the two chromatids of the homologous chromosome, move to the other pole. There is no way to predict which pole will receive which type of allele. Similarly, the S alleles separate from the two s alleles and move randomly to the poles. In some cells, the pole that receives the Y alleles at telophase I also receives the S alleles, but in other cells, the Y and s alleles end up together. During meiosis II, the two chromatids of each chromosome separate from each other, resulting in four types of haploid cells in equal numbers: SY, sY, Sy, and sy. Any single microspore or megaspore mother cell produces only two types of haploid cell: the set SY and sy or the set sY and Sy. All four types are produced by a single plant because some mother cells produce one set and some produce the other set.
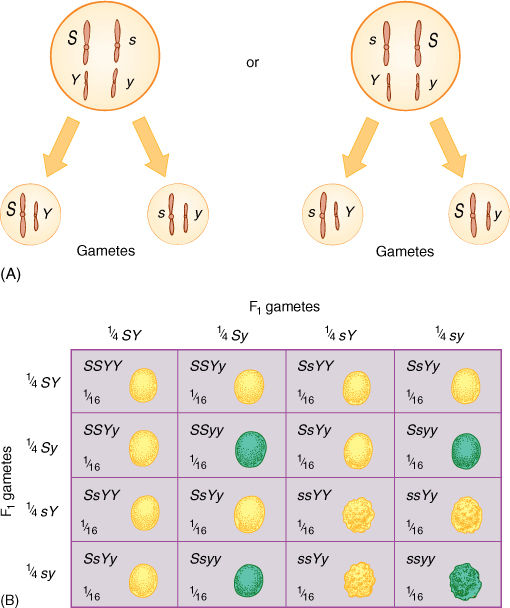
FIGURE 16-17 (A) During anaphase I, chromosomes move independently of each other, so one pole receives S and the other receives s; likewise, one gets Y and the other y. But in about half the cells, both S and Y move to the same spindle pole by chance, whereas in other dividing cells, one spindle pole receives S and y, again by chance. (B) A Punnett square for a dihybrid cross is set up just like one for a monohybrid cross; establish the types and relative abundance of gametes, and then fill in the squares. The table looks a little formidable, but it really consists of two 3:1 ratios intermingled.
After the possible types of gametes are known, the Punnett square can be set up, as in FIGURE 16-17B. Any single fertilization results from the syngamy of one sperm cell and one egg cell and produces only one of the 16 possible zygote genotypes shown in Figure 16-17B. All 16 types of zygote occur only if we study many fertilizations; to have the ratios come out accurately, we have to analyze hundreds of progeny. In plants, it is usually easy to obtain large numbers of fertilizations because pollen and ovules are produced in large amounts. After pollinated, most plants produce dozens or even thousands of seeds, enough progeny that all 16 zygote genotypes occur in about the expected ratios. But in large animals such as mammals, reproduction may be infrequent, and only one or two offspring are produced each year; a great deal of work is necessary to get enough progeny to verify the results of a dihybrid cross.
In a dihybrid cross involving independent assortment of two heterozygous genes, each gene showing complete dominance, a characteristic phenotype ratio occurs, just as is true of the 3:1 ratio in a monohybrid cross. The ratio is 9:3:3:1, with 9/16 of the plants having the dominant phenotype for both traits (in our example smooth yellow seed coats), 3/16 with the dominant phenotype of the first trait and the recessive phenotype of the second (smooth, green), 3/16 with the first trait recessive and the second dominant (wrinkled, yellow), and 1/16 in which the plants have the recessive phenotype of both traits (wrinkled, green) (TABLE 16-3). The 9:3:3:1 ratio results only if all four types of gametes are produced in equal numbers and have equal opportunity to participate in reproduction. The alleles Y and y must be independent of S and s during meiosis I; this automatically happens if they are on different chromosomes.
TABLE 16-3 Genotype and Phenotype Ratios of a Dihybrid Cross*
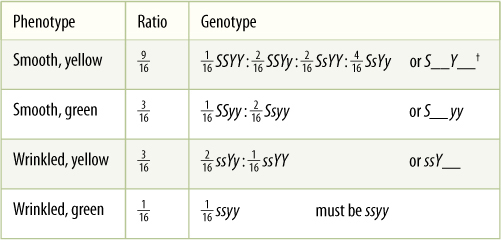
* The two individuals are heterozygous for two characters showing complete dominance and independent assortment.
† The notation ___ indicates that either allele may be present; e.g., S__yy represents either SSyy or Ssyy, both of which produce a smooth green phenotype.
Notice that if only one trait is considered, it behaves as in a monohybrid cross: Plants with smooth seeds outnumber those with wrinkled seeds by 3:1, and those with yellow seeds are three times more abundant than those with green seeds. Similarly, the monohybrid genotype ratios are also present—1 SS:2 Ss:1 ss and 1 YY:2 Yy:1 yy. Considering two genes simultaneously does not affect their inheritance at all.
Crossing-Over
Independent assortment can also occur if two genes are located far apart on the same chromosome such that crossing-over occurs between them during prophase I, after homologous chromosomes have paired and a synaptonemal complex is formed. Because no preferential sites for crossing-over seem to exist, the farther apart two genes are, the greater the possibility that crossing-over will occur between them. Most plant chromosomes are so long that crossing-over occurs several times within each chromosome during each prophase I. Consequently, the two ends act like separate entities; if the gene for seed coat color were at one end of the chromosome and the gene for seed coat texture were at the other, they would still undergo independent assortment. If the two genes are close together on a chromosome, however, crossing-over may not occur and the two alleles may move together during meiosis I, as described below.
Genes on the Same Chromosome: Linkage
If two genes occur close together on a chromosome, they usually do not undergo independent assortment during meiosis I; instead, the two genes are said to be linked. Consider a plant heterozygous for the two traits of the seed coat again, but now imagine the genes occurring close together on one chromosome. During meiosis, haploid cells that are either S or s, Y or y are formed, but a new complexity arises. Because the genes are linked, several types of heterozygote are possible: An SsYy individual may have the alleles S and Y on one chromosome and the alleles s and y on the homologous chromosome, but an SsYy individual may have s and Y linked together and S and y linked (FIGURE 16-18A). We must consider the types of gametes that can be formed. If crossing-over is ignored, the first individual produces haploid cells with the genotypes SY and sy only, whereas the second plant would produce sY and Sy gametes only. The result of selfing is not a 9:3:3:1 ratio, but something drastically different.
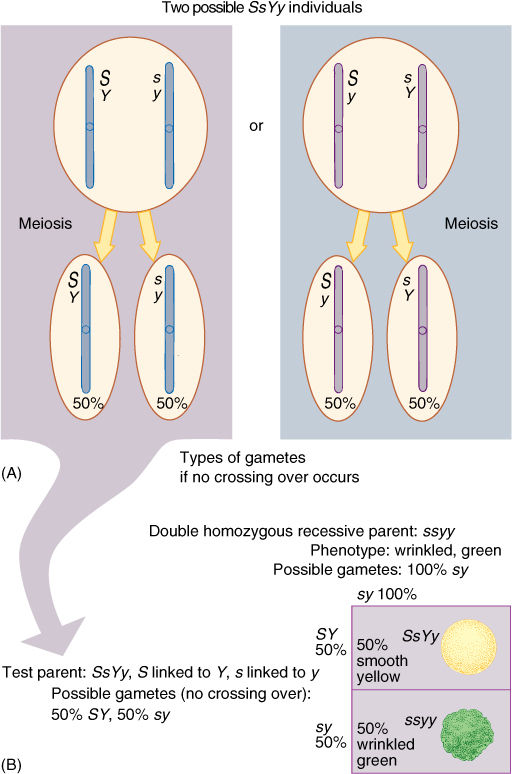
FIGURE 16-18 (A) Two types of SsYy individuals are possible, but in most instances, their phenotypes are identical. Only genetic tests can distinguish which is which, based on the unique gametes produced by each. (B) A test cross, with the plant on the left in (A) being the test parent. Try setting up the Punnett square for a test cross with the plant on the right in (A). A test cross of two closely linked genes produces results very different from those of a test cross involving nonlinked genes that assort independently. What would the Punnett square be like if S and Y, s, and y were not linked?
Botany and Beyond
BOX 16-2 Whose Genes Do You Have?
You received half your chromosomes from your father and the other half from your mother. So half of the DNA you had when you were a zygote—a single cell—was your father’s and half was your mother’s. But beyond that? One fourth of your genes came from your paternal grandfather (your father’s father), one fourth from your paternal grandmother, and so on for your maternal grandparents. But, can we say that one fourth of your DNA came from each of your grandparents? That is not so easy. Of the thousands of cells in your father’s testes, one in particular underwent meiosis and produced four sperm cells, one of which gave rise to you. During prophase I of that meiosis, the chromosomes your father got from his father paired (synapsed) with the ones he got from his mother (your grandmother), and they underwent crossing-over. If each homologous chromosome were to break in exactly the same place, then when they are repaired and meiosis continued, each would have exactly the same amount of DNA. Often, however, homologous chromosomes do not break at quite the same spot; after repair one might have a bit more DNA, the other a bit less, one might be mostly paternal, the other mostly maternal. As long as they are not too unbalanced, the chromosomes will be functional. But because of this, we cannot say that exactly one fourth of your DNA came from each of your grandparents: Some might have given you a bit more, and others a bit less.
With each generation we go back, the number of ancestors doubles and the number of genes we received from each drops by half. By the time we get to the 15th generation we have 32,768 great-great-etc.-grandparents (215), and each contributed 1 in 32,768 of our genes. We believe, however, humans only have somewhere between 30,000 and 35,000 genes. If we assume each generation is about 20 years (some people have children earlier, some later), then this 15th generation lived about 300 years ago, about 1700 CE, not long after the first microscopes were invented. Of course, each of us is descended from many more generations before that, but even so, we may not have actually gotten any of their DNA.
The most instructive cross is a test cross using a double homozygous recessive parent: ssyy. If we continue to exclude crossing-over (imagine that the two genes are extremely close together), the gametes from the first type of heterozygote will be SY and sy and the gametes from the homozygous recessive parent will be all sy. Two types of F1 will be produced, as shown in FIGURE 16-18B, smooth yellow and wrinkled green, and they occur in a ratio of 1 to 1. The two phenotypes are like those of the parents.
If the genes are not extremely close together, crossing-over may occur; for example, in 10% of the cells undergoing meiosis, a crossing-over might happen; therefore, the plant would produce four types of gametes, but not in equal numbers. Of the gametes for the first type of heterozygote, 47.5% would be SY, 47.5% sy, 2.5% sY, and 2.5% Sy (FIGURE 16-19A). The last two are recombinant chromosomes formed from a crossing-over of the homologous chromosomes and recombination of alleles. The first two are parental type chromosomes. In the majority of the cells, Y is still linked to S and y is linked to s, but in a minority, Y has become linked to s and y to S. A Punnett square alone is not sufficient to show the relative proportions of progeny, so the percentages of gamete must be added. A test cross now would result in 47.5% of the F1 progeny being smooth and yellow, 47.5% wrinkled and green, 2.5% wrinkled yellow, and 2.5% smooth green (FIGURE 16-19B). The first two types are like the parents, but the last two are recombinant types produced at the same percentage as crossing-over occurs. We cannot actually “see” crossing-over with a microscope, but we can infer that because 5% of the F1s have a recombinant phenotype, then 5% of the chromatids must have undergone crossing-over.
The rate of crossing-over is directly proportional to the physical spacing between the genes on a chromosome. Two genes that produce 6% recombinant F1s are closer together than two genes that produce 10% recombinant F1s. By analyzing as many mutant alleles as possible, we can measure “space” between them in recombination percentages, each 1% being called one map unit or one centimorgan; one map unit, on average, is approximately one million base pairs. By this means, a genetic map can be constructed. If enough genes can be mapped, we may find that A is linked to B, which is linked to C; even if A and C are so far apart that they assort independently, by knowing that both are linked to a common gene, B, we know that all three are on the same chromosome. On the other hand, if A, B, and C cannot be shown to be linked to another gene F, that does not mean that F is on a different chromosome; it might be that intervening genes have not yet been mapped (FIGURE 16-20). A set of genes known to be linked is called a linkage group; when all genes are mapped, there will be exactly as many linkage groups as there are chromosomes, but if only a few genes are mapped, there may be more linkage groups than chromosomes.
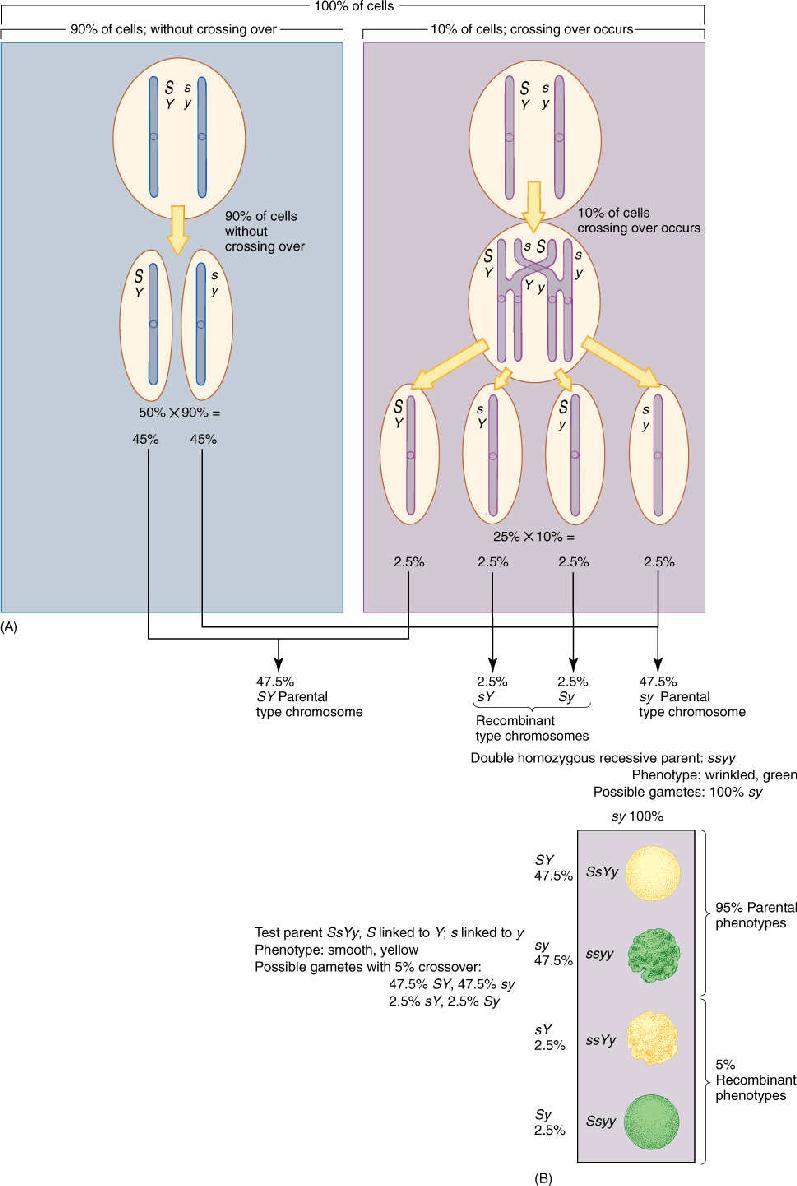
FIGURE 16-19 (A) With linkage, four types of haploid genotypes might be produced, but not in equal numbers. Recombinant types are less abundant than parental types—in this case 5% recombinant chromosomes and 95% parental type. These two genes are 5 map units apart. (B) This test cross, unlike that of Figure 16-18B, gives four types of progeny: two parental types and two recombinant types. The alleles S and Y show linkage rather than independent assortment.
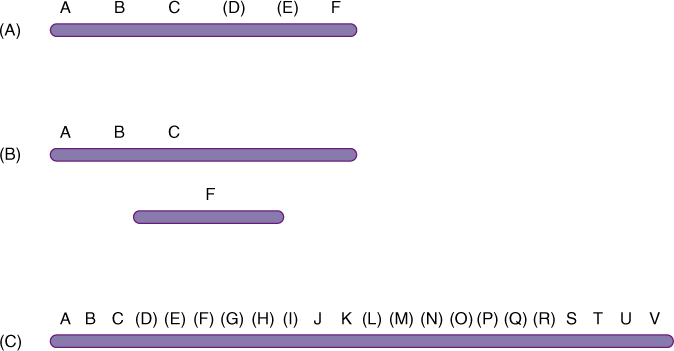
FIGURE 16-20 (A) A and C might be far enough apart for crossing-over to occur in virtually all nuclei, so they undergo independent assortment. If each can be shown to be linked to B, they must be on the same chromosome. If A, B, and C are not known to be linked to F, it might be because they are on separate chromosomes (B) or because they are on the same chromosome but intervening genes have not yet been mapped, as shown in (A). (C) Because of incomplete mapping, ABC, JK, and STUV are three linkage groups even though there is only one chromosome.
![]() Multiple Genes for One Character: Quantitative Trait Loci
Multiple Genes for One Character: Quantitative Trait Loci
Individual phenotypic traits are the result of complex metabolic processes involving numerous enzymes; therefore, many separate genes may affect any single trait. The gene R was described as affecting flower color by producing an enzyme that synthesized a red pigment, but that enzyme requires the proper substrate, which is present only if it is synthesized by a different enzyme controlled by a distinct gene. If this gene is present as an ineffective allele, there will be no substrate and thus no pigment, regardless of whether the first flower color gene is present as allele R or allele r. In Arabidopsis thaliana, abscisic acid (ABA) causes developing seeds to become dormant at maturity. In several mutants (TABLE 16-4), mature embryos continue to grow as if germinating and do not enter dormancy. One mutant, aba, is unable to produce enough abscisic acid. A second mutant, abi-3, produces plenty of abscisic acid but does not respond to it—perhaps its receptor is the affected protein. Although aba and abi-3 involve different genes, they both result in the same phenotype. They are multiple genes for one trait even though they are not part of one metabolic pathway.
TABLE 16-4 Multiple Genes for One Character
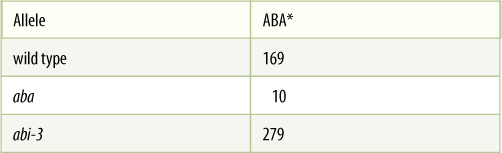
* ng/g fresh weight of seed tissue.
Most synthetic pathways involve at least four or five intermediates, and their four or five genes all affect the same trait. Complex traits such as leaf shape, cold hardiness, and general vigor can be the results of dozens of genes and rather than having phenotypes that are simply dominant and recessive, the trait may show a continuous gradient of values. Having multiple genes for each trait is referred to as epistasis. Some intermediates may be produced by several pathways, each with its own enzymes; a mutation in one is not necessarily particularly severe because the alternate pathway may increase its activity and produce sufficient amounts of the intermediate. If this happens, the mutation is masked and is not reflected in the phenotype, even if it is present in the homozygous condition. Extremely complex crosses, involving hundreds or thousands of progeny, may have to be performed to determine what fraction of a particular phenotype is correlated with a particular gene; the genes or other portions of DNA associated with such traits are quantitative trait loci. Conversely, when an intermediate is part of several metabolic pathways and is produced by only one enzyme, a mutation in that enzyme’s gene affects all of the pathways and alters several different traits. Multiple phenotype effects of one mutation are called pleiotropic effects. For example, any mutation that affects the protein portion of phytochrome affects all developmental processes controlled by phytochrome, and mutations that alter the level of pyruvate affect the citric acid cycle, amino acid synthesis, and C4 metabolism.
![]() Other Aspects of Inheritance
Other Aspects of Inheritance
Maternal Inheritance
In the crosses described above, the alleles of both parents are transmitted equally to the progeny, a situation called biparental inheritance. All genes in the nucleus undergo biparental inheritance. An unexpected feature of both plants and animals is that during fertilization, the sperm cell loses most of its cytoplasm and only the sperm nucleus enters the egg. Consequently, the zygote obtains all its plastid and mitochondrion genomes from the maternal parent; this is uniparental inheritance, more specifically maternal inheritance.
Mitochondrion genetics are difficult to study for several reasons. First, each cell has many mitochondria, up to hundreds, and each mitochondrion has several circles of DNA. Each cell has multiple copies of each mitochondrion gene and may have received diverse types of alleles from its maternal parent. Simple Mendelian ratios are not produced. Second, the presence of many of the mitochondrion DNA mutants does not result in easily recognizable phenotypes. The plants usually simply grow somewhat more slowly, or if the mutation has a severe effect, they may die as embryos. Consequently, the presence of mitochondrion mutants may not be detectable by examination.
Plastid genetics are somewhat easier if alleles affecting chlorophyll synthesis are studied. Consider a plastid whose DNA has mutated so that the plastid can no longer produce any chlorophyll; it may be red or orange because carotenoids are no longer masked by chlorophyll, or it may be white if no pigment at all is produced. If a zygote receives only plastids like these, the embryo and seedling will be red, orange, or white. Such seedlings are rare, but in large commercial nurseries that grow millions of seedlings, they appear from time to time. The plants can be kept alive artificially if they are grafted onto a normal plant that supplies them with carbohydrates (FIGURE 16-21). Grafted plants can then be used for genetic experiments.
When an achlorophyllous plant is crossed with a green, chlorophyll-bearing plant, the outcome depends on which plant is the pollen parent and which is the ovule parent. If the achlorophyllous parent provides the pollen, 100% of the zygotes will be green because they receive all of their plastids from the normal parent by way of the egg; all mutant plastids are destroyed during sperm differentiation and syngamy. But if the achlorophyllous plant is the ovule parent, all progeny also have mutant plastids and are achlorophyllous. For some reason, conifers are an exception among all plants: Conifer eggs destroy their plastids but conifer sperms do not. Therefore in conifers, plastid genes are inherited paternally, mitochondrial genes are inherited maternally, and nuclear genes have biparental inheritance.

FIGURE 16-21 Plastid mutations in the cactus genus Gymnocalycium seem to be common; quite often totally red or orange seedlings appear in cactus nurseries. These lack chlorophyll but can be kept alive by grafting them onto a green cactus that supplies them with sugar. Many plants like Coleus and maples have bright red leaves; they do have chlorophyll, but it is masked by large amounts of other pigments.
Plastid inheritance is also responsible for certain types of variegation in plants, the presence of spots or sectors that are white, red, or orange on a plant that is otherwise green (FIGURE 16-22). Because plant cells have many plastids, only rarely do all the plastids of an egg have the same alleles unless the parent has only one type, as in the cactus of Figure 16-21. More often, each cell has a mixture of plastid types. During cell division, some plastids move into each end of the dividing cell; usually random chance alone results in each daughter cell getting some of both types of plastid (Figure 16-22A); however, occasionally, one daughter cell by chance receives only mutant plastids. It is white, red, or orange and is able to survive because surrounding normal cells supply it with sugar. If this occurs in a leaf primordium or a young internode, the cell continues to grow and divide along with the surrounding cells, resulting in a patch of cells that lack chlorophyll (Figure 16-22A). If it occurs in a very young leaf, the cells grow quite a lot and produce a large spot; if it happens in an older leaf that has reached almost full size, the patch is small.
When variegation affects stems, the achlorophyllous patch may include an axillary bud. If so, the entire bud and the branch that grows from it are achlorophyllous. Any flowers it produces can be used for genetics experiments, providing either pollen or ovules. Experiments on both corn and morning glory have shown the achlorophyllous character to be maternally inherited; thus, it is a feature of the plastid DNA as just described. Recombinant DNA technology is now greatly accelerating our efforts to understand plastid and mitochondrion genetics. In both organelles, the DNA is a circular double helix, one of its ends being attached to the other, and no histones are present. Plastids contain many circles, 200 in each spinach plastid and 1,000 in each of those of wheat, so rather than being haploid or diploid, wheat plastids are 1,000 ploid. Plastid DNA may constitute up to 21% of the total DNA content of a cell. In angiosperms, plastid DNA tends to be about 150 kbp long. In plants, mitochondrion DNA circles are very large, well over 200 kbp (up to 2,400 kbp), more than ten times larger than mitochondrion DNA in animals.
Genomes of plastids contain genes for numerous enzymes as well as a complete set of plastid ribosomal RNAs and 30 of the transfer RNAs. Currently, it is estimated that 80 to 100 enzymes and membrane proteins are coded by plastid DNA, and many of these are essential: the large subunit of RuBP carboxylase, components of the proton channel/ATP synthetase complex, cytochromes, and proteins that bind chlorophyll a (FIGURE 16-23). Mitochondrion DNA also contains genes for rRNA and tRNA, as well as those that code for proteins involved in ATP synthetase complexes, electron transport, and NADH dehydrogenase complex. In many cases, organelle genes must work in coordination with nuclear genes. The interaction of these three genomes within one cell is now an area of very active research.
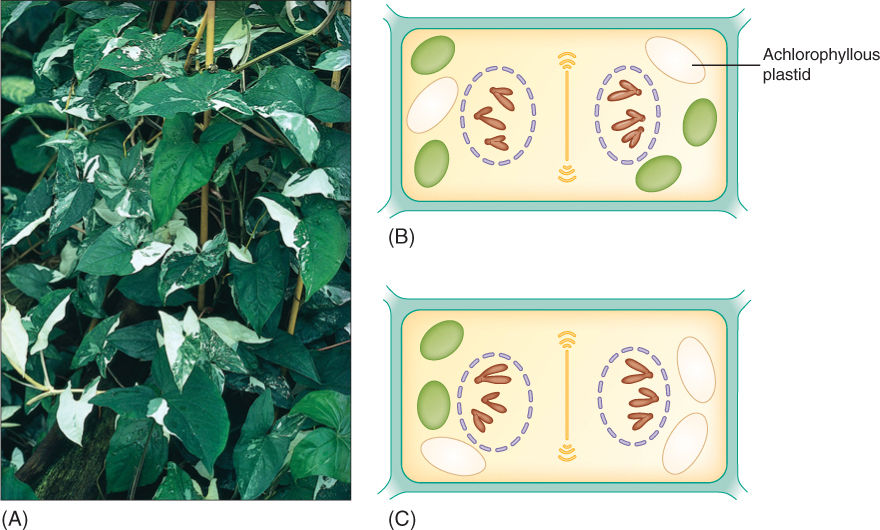
FIGURE 16-22 (A) Variegated plants of Syngonium podophyllum. Each green cell probably has a mixture of normal chloroplasts and plastids with a mutation that prevents chlorophyll synthesis. Cells in the white area have only mutant plastids. (B and C) Cells usually have many plastids and mitochondria, so random distribution of the organelles is usually sufficient to ensure that both cells receive some (B). If mutant plastids are present, then occasionally when cytokinesis occurs one end of the cell may by chance have only mutant plastids (C). Most cells have only two sets of (nuclear) chromosomes; therefore, all of the elaborate mechanisms of mitosis are necessary if each daughter cell is to receive one complete set.
As mentioned in the beginning of this chapter, recombination of genes during meiosis and sexual reproduction creates greater genetic diversity, which is the raw material of natural selection: Sexual reproduction results in more rapid evolution. But plastids and mitochondria, being derived from endosymbiotic prokaryotes, have no sex, no genetic recombination. However, there are mechanisms that transfer genes from the organelles into nuclei where they become nuclear genes. We think this happens very rarely (once every several million years), but gradually, most of the prokaryotic ancestral genes of plastids and mitochondria are now located in the nucleus (they are recognizable in DNA sequencing projects because they lack introns and other features of eukaryotic genes), and the DNA circles within the organelles themselves have only a few genes left. The organelle-derived genes that are now located in the nucleus have all of the benefits of nuclear genes: They are not destroyed as sperm cells mature; syngamy of sperm and egg brings two sets of genes (possibly with different alleles) together into one nucleus, and then during meiosis, homologous chromosomes pair and undergo crossing-over, generating genetic diversity. These genes can evolve rapidly, whereas those still located in the organelles evolve more slowly.
Lethal Alleles
The phenotypic result of a mutation can vary in severity from almost undetectable to lethal; that is, its presence can kill the plant. Genetically inherited lethal mutations are most often recessive and are fatal only if present in the homozygous condition; a heterozygous plant has enough normal protein to survive. The reason for the recessive nature of most lethals is simple: A dominant lethal would kill the plant, even in the heterozygous condition; therefore, it would rarely be passed on to any offspring.
The presence of a lethal allele can be difficult to detect if its effect occurs early. If it affects basic metabolic functions such as respiration, DNA replication, cellulose synthesis, or the structure of histones, plants homozygous for it probably die while still very young; in some instances, the gametes die even before fertilization. The only seeds that complete development are heterozygotes or dominant homozygotes. This results in unusual genotype and phenotype ratios. A selfing of a heterozygote for a lethal condition, Ll, produces zygotes in the typical 1 LL:2 Ll:1 ll genotype ratio, but the ll zygotes die. The seeds and plants that live are 1 LL:2 Ll genotypically; such a ratio should always lead one to suspect a recessive lethal allele.
FIGURE 16-23 This is a map of the plastid genome of cotton, Gossypium hirsutum. Look first at the smaller central circle; it has four parts: the Large Single Copy, the Small Single Copy, Inverted Repeat a, and Inverted Repeat b. IRa and IRb are duplicates of each other, but with the genes of one running in the opposite direction (inverted) as those of the other. The two single copy regions do not duplicate each other or the repeat areas. The large outer circle displays the locations of all the genes. The plastid genomes of hundreds of species have been sequenced, and by comparing the maps of the various species, we can tell which species are more closely related and which are more distant.
Plants have an alternation of heteromorphic generations, the diploid sporophytes alternating with the haploid gametophytes. In gametophytes, each gene is present only as a single allele. No homologous chromosomes occur, and recessive and lethal alleles are not masked by the presence of a dominant homologous allele. In animals, this haploid phase is restricted to the sperm and egg, which are simple and have many inactive genes. As a result, numerous genes with deleterious mutations are not active during the brief haploid condition; they are turned on only during growth and differentiation from a zygote to an adult, when their effect is mitigated by their diploid condition. In plants, however, the haploid phase consists of at least a pollen grain with three cells and a megagametophyte with seven cells. This phase is still quite simple and most genes are inactive; however, in both gametophytes, many types of central metabolism are occurring, and any highly deleterious allele causes that gametophyte to develop poorly or not at all. If the gametophyte dies without producing a sperm or an egg, the allele is eliminated.
Multiple Sets of Chromosomes and Gene Families
Whereas most animals have diploid body cells and haploid gametes, plants are much more diverse. Rarely, a spore is formed without undergoing meiosis, resulting in diploid gametophytes and diploid gametes. If these fertilize a normal haploid gamete, a triploid zygote results. Although this would be instantly lethal in almost all animals, plants tolerate it well, and a triploid sporophyte develops from the zygote. Triploids, however, are sterile because they cannot undergo pairing of homologous chromosomes during prophase I; chromosomes come together in threes, and the rest of meiosis is aberrant.
Occasionally, cells fail to undergo mitosis after S phase DNA replication, and the cell becomes tetraploid (or diploid if it happens in a gametophyte). Tetraploid cells are usually perfectly healthy; if they produce a part of the plant that initiates flowers, diploid pollen and eggs are produced, again raising the possibility of triploid zygotes if fertilized by haploid gametes. These processes can occur in virtually every conceivable combination, and plants that are 3n (three sets of chromosomes, triploid), 4n, 5n, 6n, . . . up to several hundred n are common. All plants with more than two sets of chromosomes are polyploid. All plants with an even ploidy level can undergo meiosis and are fertile; all odd ploidy levels are sterile like triploids.
A change in chromosome number per nucleus in plants can also come about by nondisjunction: During the second division of meiosis, the two chromatids may remain together (fail to disjoin), so one daughter cell receives both copies of the chromosome (the two chromatids), whereas the other receives none. The cell with the extra chromosome probably survives quite well and forms a functional sperm or egg cell. If this gamete, which is diploid for one chromosome, is involved in fertilization, the new zygote will be triploid for that chromosome. It received a normal, haploid chromosome set from one parent and a haploid set plus the extra chromosome from the parent in which nondisjunction occurred. In plants, this condition is tolerated often, and only by studying the karyotype can it be discovered. In animals, this is almost invariably fatal. The human congenital disease Down syndrome is caused by nondisjunction of the very small chromosome 21. Down syndrome individuals are triploid for just a few genes yet have severely disrupted metabolism. Fetuses triploid for longer chromosomes abort spontaneously.
Plants with even ploidy levels grow and reproduce successfully, often more vigorously than diploid individuals of their species. Initially, these have complex genetics. For example, a tetraploid can have any of the following genotypes for red/white flower color: RRRR, RRRr, RRrr, Rrrr, or rrrr. Its gametes can be RR, Rr, or rr. Notice that polyploids have more copies than needed for every gene. In all diploid plants, even heterozygotes with only one functional allele are usually healthy. For nonessential genes, a complete absence of functional alleles may not be harmful, such as when rr produces white flower color. Consequently, the two or three extra copies of each gene in a tetraploid plant are surplus DNA. Mutations that completely incapacitate one of the four alleles of a tetraploid usually are not deleterious, as one, two, or three functional alleles remain in every nucleus. For polyploid plants, deleterious mutations tend to have little effect on phenotype; therefore, they do not affect survival as much as they would in a haploid cell or a diploid plant. As a result, deleterious mutations are not eliminated quickly by natural selection, and the extra copies of a gene may rapidly become nonfunctional; as long as the genes are still so similar to the wild-type allele that they can be recognized as having originated as duplicates of it, they are called paralogs. Before many generations have passed, so many genes have changed that polyploids act like diploids, their “extra” copies having mutated. Almost half of all flowering plant species are actually polyploids; those that now appear to be perfectly normal diploids probably underwent conversion to the polyploid condition so long ago that all their extra alleles have mutated extensively and can no longer be considered alleles of those that are still functional. Nondisjunction, insertions, and deletions may have changed the numbers and sizes of the chromosomes so much that the four originally homologous chromosomes now appear as just two different pairs. Conversion back to the diploid state is an uneven process, and some species act like diploids for certain genes but tetraploids for others.
What do the extra alleles mutate into? At first, they may simply be alleles that allow the cell to produce a particular protein more rapidly or produce slightly different versions of the same protein. For example, the five histone proteins that make up nucleosomes are needed in such abundance that each nucleus may have up to 600 paralogs of the genes, not just two as for most traits; these multiple copies constitute a gene family (FIGURE 16-24). Similarly, there are at least five versions for phytochrome genes, all with such closely related nucleotide sequences that we are certain they are a gene family that evolved from one ancestral gene. The receptor-like kinases (RLKs) constitute a very large gene family with dozens of members already known and probably many yet to be discovered. Each paralog will evolve independently of the others, and some may ultimately produce defective proteins (but some must continue to produce the proper protein). Mutations in the promoter regions may prevent them from ever being activated, so they turn into “junk,” acting as nothing more than spacer DNA. Deletions, inversions, and movements by transposons may break them up. DNA sequencing has now revealed that there are large families of genes in which one gene codes for a useful protein, whereas the other genes have nucleotide sequences that are obviously related to the useful gene but are inactive and apparently code for nothing. For example, the genome of Arabidopsis thaliana contains 27,416 genes that code for proteins and 4,827 “pseudogenes” that are related to genes but which now are so altered they are not transcribed into mRNA and thus can no longer guide the translation of their information into proteins.

FIGURE 16-24 As corn seeds mature, they fill with the storage protein zein. Each nucleus has two families of zein genes, each with about 25 copies of the gene, and thus, zein mRNA can be produced very rapidly. It is not essential that all zein molecules have identical amino acid sequence (they will merely be digested by the embryo when the seed germinates); therefore, zein paralogs evolve independently of each other, but still, each produces a zein protein 90% identical to those produced by any other member of the gene family.
This extra DNA is actually extremely valuable—it is the raw material for the evolution of new genes. Some may mutate into genes that produce enzymes almost identical to those being coded by the original form of the allele. The original enzyme may work best at low temperature, whereas the new form may have a higher optimum temperature. The plant can now produce two types of enzymes and function well in both warm and cool days, or the species may be able to extend its range, the new enzyme allowing it to survive in hot deserts and the original enzyme allowing it to live in its original habitat (FIGURE 16-25). Mutations in the promoter region instead may allow the structural portion to produce the same protein as the original gene but at a different time or place in response to a different chemical messenger.
In other cases, mutations in the extra DNA may result in totally new genes that produce proteins whose function is not at all related to that of the original gene. Mosses, ferns, conifers, and flowering plants have evolved from green algae. Because algae have no genes for flower color, lignin synthesis, and so on, the evolution of these species has involved the evolution of entire new metabolic pathways. All of these genes had to arise by the gradual mutation of surplus alleles into nucleotide sequences that code for useful proteins.
FIGURE 16-25 After a plant becomes tetraploid, mutations in the extra copies of a gene are tolerable as long as (1) at least one allele is left functional, and (2) the extra copies do not mutate into a lethal allele. In this example, two chromosomes have changed morphology, becoming shorter; they could have become longer by picking up pieces of DNA by unequal crossing-over. Several copies have mutated into spacers and some into junk that still has much sequence homology with functional alleles. Others have evolved into new alleles of the original genes (d3 to d5 ; a2 to a3 ; f3 to f4), and some have evolved into entirely new genes (a1 to g1; d4 to h1).
Alternatives
BOX 16-3 Genetics of Haploid Plants
This chapter discusses diploid zygotes formed by syngamy, the fusion of a haploid sperm with a haploid egg. Each zygote has two chances of obtaining at least one functional allele for every gene in its nucleus. If one gamete happens to supply a nonfunctional allele for a particular gene, the zygote will probably survive if the other gamete provides a functional allele: The zygote will be heterozygous for that gene, but often a single functional copy is enough. Almost all plants familiar to us benefit from the safety net of having two sets of genes in each nucleus.
Haploid plants also exist, however. Each has only a single parent, not two, and thus, each nucleus has only a single set of chromosomes, not two. Mosses are a familiar example: Each moss plant is initiated when a haploid spore germinates and grows. Spores differ from gametes in being able to undergo mitosis and growth but being unable to fuse with another cell. A gamete, on the other hand, undergoes syngamy with another gamete but can not grow or divide (unfertilized eggs of bees are exceptional and develop into haploid workers). After a haploid moss spore germinates, it grows as a branching network of green parenchyma cells, and later, some cells take on the shape of a shoot apical cell and produce upright green, leafy little moss plants. Every nucleus in every moss plant is haploid.
How does this affect the genetics of these plants? Imagine a gene that has two alleles, A and a. Some plants in the moss population might be A and others a, but none is ever AA, Aa, or aa. If a produces a nonfunctional enzyme, then all of the a mosses will be in danger and probably die before they can reproduce. When they die and decompose, the nuclei and all of the genes—including the a allele—break down; consequently, this allele will be very rare in the moss population. In diploid plants and animals, nonfunctional alleles can persist for many generations in the heterozygous condition: It is usually when they occur in homozygotes, aa, that natural selection eliminates them. Because mosses are haploid, nonfunctional alleles cannot be protected by heterozygosis and natural selection eliminates them quickly. Moss populations tend to have “healthy” genomes in which almost all alleles are functional.
Is this good or bad, or more accurately, what are the consequences of quickly eliminating mutant alleles? Evolution by natural selection operates only if there is genetic diversity. Mutations produce new versions of genes, versions that often are less functional than the original versions but which may actually be better in a different habitat, one that is drier or wetter, warmer or cooler, or which has different animals and fungi. If the plant population has both the original and the new allele, it may be able to adapt to a wider range of habitats, or as the climate changes, a more diverse set of alleles for many genes may allow the population to adapt rapidly enough to survive the new climatic conditions. Imagine a new allele arising in some diploid plant. If it occurs in a somatic cell, it cannot be passed on to another individual, but if the mutation occurs in a sex cell, then a copy of the new allele will be incorporated into some of the sperms and eggs and then into zygotes. Almost certainly, the diploid zygote has gotten a functional allele from the other parent, and thus, the zygote will be heterozygous and will survive—and so will the new allele. Every time this plant reproduces, more copies of the allele will be duplicated and passed on. At some point, a seed carrying this allele might land in a microhabitat where this allele is adaptive, not useless, and the allele will then help the population adapt to a broader range of habitats.
Now imagine a similar allele being formed by mutation in a haploid plant. Again, if it forms in a somatic cell, it does not matter; however, if it forms in a spore, then it will soon find itself as the sole allele for that gene in a haploid plant, and almost certainly the plant will suffer from not having a functional wild-type allele. Under very lucky circumstances, the spore with this new allele might land in a habitat where the allele is adaptive; if so, this may be beneficial, but the chances are low that a spore with the very first copy of new, mutant allele would land in a suitable area. It is more likely that the spore will land near the parent, in a habitat similar to the parent’s habitat and the spore with the mutant allele will develop poorly or die, eliminating the allele within a few days or weeks after it formed. The gene pool of the moss population will return to being the same as it was before the mutation and will have no new genetic resources for helping it adapt to new or changing conditions.

FIGURE B16-3 All of the short plants beside this stream are mosses, and all are haploid. Each grew from a haploid spore, not from a diploid embryo in a seed.
 At the Next Level
At the Next Level
1. Gene families. Sequencing the entire genome of a species is now so quick, easy, and inexpensive that we already have complete genomes for dozens of species. Very often this is speeded up by not bothering to sequence every single nucleotide, but to instead ignore difficult-to-sequence regions such as telomeres, centromeres, and regions of short repeats. Also, instead of sequencing all the genome, people often focus on just one particular gene. For example, the gene that codes for the protein portion of phytochrome exists in several forms, and we are as interested in its evolution as we are in the evolution of whole species. We can use one known sequence of the phytochrome gene as a probe to find related genes in other species. We now know the evolutionary history of many genes, many of which involved duplication and diversification into gene families. The phytochrome gene family is well-known, important, and instructive. You can study it in detail, and then search for information on other gene families.
2. Doubling of the entire genome. In the evolutionary history of plants, the ancestors of certain lineages underwent a doubling of the whole genome. A plant and its immediate descendants were tetraploid, and quickly the extra genes mutated and evolved to have new functions. As more and more of the extra genes evolved to have new functions, the clade would begin to act as if it were diploid again, and with enough gene conversion, it finally truly would become diploid. But then genome doubling might occur again. Whole genome doubling has occurred several times in the history of certain members of the angiosperms. The consequences of this are important and interesting.
3. Recommended text. Plant genetics is such an important and large topic, several advanced textbooks are available. If you wish further study now on your own, it might be best to use one of these books to provide focus and an integrated approach. I recommend Principles of Plant Genetics and Breeding, Second edition by George Acquaah (2012, Wiley-Blackwell).
SUMMARY
1. Most genes occur as slightly different forms, alleles, in which the DNA sequence varies. Alleles arise by mutations.
2. An organism’s physical and physiological features constitute its phenotype, whereas its suite of alleles is its genotype.
3. During sexual reproduction, two sets of alleles, maternal and paternal, are brought together into one cell. During prophase I of meiosis, alleles of homologous chromosomes are rearranged into new chromosomes.
4. Mutations are any changes in the sequence of nucleotides in DNA. Larger mutations are often more significant than small ones, but any type can potentially affect the phenotype. The great majority of mutations are selectively disadvantageous.
5. DNA proofreading and repair enzymes keep the mutation rate low enough to allow complex organisms to live, but they are imperfect enough to allow a low rate of mutation, permitting evolution by natural selection.
6. If a diploid organism has two identical alleles for a gene, the organism is homozygous for the gene; if it has two distinct alleles, the organism is heterozygous for that gene. If the phenotype of heterozygotes is indistinguishable from that of one of the homozygotes, the trait shows complete dominance. If the heterozygote’s phenotype differs from that of both homozygotes, the trait shows incomplete dominance.
7. When heterozygotes with complete dominance are selfed, the F1 generation should have a phenotype ratio of three dominant to one recessive, and a genotype ratio of one homozygous dominant to two heterozygotes to one homozygous recessive.
8. A test cross is one in which one parent is known to be homozygous recessive for the trait of interest; this allows the phenotype of the other parent to be expressed in every progeny, thus revealing the genotype of the parent.
9. Almost any gene has multiple alleles; that is, there are many mutant forms of the gene. Any particular phenotype trait is probably the result of the action of many enzymes or other proteins; the alleles for all these genes may affect the expression of the phenotype.
10. A dihybrid cross is one in which two traits are studied simultaneously; if the responsible genes are on separate chromosomes or if they are widely separated on the same chromosome, they show independent assortment.
11. Two genes that occur close together on a chromosome are linked. Crossing-over during prophase I breaks up linkage groups.
12. In sperm cells of both plants and animals, organelles are destroyed, so sperms do not contribute mitochondrion or plastid genes. All organelle genes are provided in the egg and show maternal rather than biparental inheritance.
13. Lethal genes cause such a severe disruption of metabolism that the individual dies. Many kill the organism before it reaches reproductive maturity, so they can be passed on only rarely, masked in the heterozygous condition.
IMPORTANT TERMS
alleles
biparental inheritance
complete dominance
dihybrid cross
DNA polymerase
dominant (allele)
F1 (first filial) generation
gene family
genotype
heterozygous
homozygous
incomplete dominance
independent assortment
lethal allele
linked genes
map unit
maternal (uniparental) inheritance
monohybrid cross
mutagen
mutation
paralogs
parental generation
phenotype
polyploid
Punnett square
pure-bred lines
quantitative trait loci
recessive (allele)
recombinant chromosomes
selfing (self-fertilization)
semiconservative replication
somatic mutations
test cross
transposon
REVIEW QUESTIONS
1. What is an allele? Do all genes come in only one form such that every plant that has that gene has exactly the same nucleotide sequence? Look at the gene in FIGURE 16-26. How do the various alleles differ from each other?
2. What is the phenotype of a plant? Can you see it? Measure it? Describe it. (Hint: Could you describe the phenotype of a geranium well enough to distinguish it from a maple tree?) What is the genotype of a plant?
3. As DNA replication begins, chromatin becomes less compact. Why?
4. When DNA is read by RNA synthase, mRNA is made. What is the name of the enzyme that reads DNA and makes more DNA?
5. Describe DNA replication. What are Okazaki fragments? Why does each chromosome have thousands of replication start sites instead of just one?
6. Name the types of mutations that may occur and describe how some of them happen. With regard to UV-induced mutations, think about the fact that most leaves last for just a few months and then are replaced by new leaves the next spring. The next time you are getting a suntan, remember that your dermis must last as long as you live.
7. Statistically, are mutations almost always beneficial or harmful? Why?
8. What is the name of a mutation that occurs in a cell that never leads to sex cells? Why is this type of mutation less important than one that affects gametes?
9. Imagine that as a leaf is growing, one of its cells is hit by ultraviolet light, which causes a mutation in a gene that is involved in producing flower pigments. Will that be an important mutation? Will it ever actually affect a flower’s color? When the leaf is abscised in autumn, will the gene have ever been transcribed to make mRNA?
10. Examine the value for lily in Table 16-1. How many billion pairs of nucleotides are there in a diploid nucleus? If DNA synthase is very accurate and if DNA repair mechanisms are so good that, on average, there is a mistake made once in every ten million nucleotides, how many mistakes are made on average every time a lily nucleus replicates? If a meter is 39 inches long, how many inches of DNA are there in every diploid nucleus of every cell in the body of a lily?
11. There are pieces of DNA that readily change their position from one chromosome to another. These are called.____________________________________________ When one of these moves to a new position, it might move into a gene and cause a mutation. Describe why this may be one of our most powerful tools for genetic analysis and engineering.
12. What are monohybrid crosses? Is it possible to make a cross in which only one single character is actually involved, or is it just that one single character is being analyzed and the others are being ignored?
13. Imagine that a plant with red flowers is bred to one with white flowers and when they make fruits, the seeds are collected and grown into new plants. The new plants, when old enough, have only pink flowers. Which plants are the parental generation? Which are the first filial generation? If the pink-flowered plants are interbred with each other and the resulting seeds planted, what would that generation be called?

FIGURE 16-26
14. Assume the plants in Question 13 are like those in Figure 16-9. What is the genotype of the plants with red flowers? Those with white flowers? Those with pink flowers? When a plant with red flowers makes pollen grains, will they all carry the R allele? Will any carry the r allele? Or will half have R and half have r? Will any of the pollen grains be Rr? Explain your answer.
15. In plants like those of Questions 13 and 14, with red, pink, and white flowers, does flower color show complete or incomplete dominance?
16. What do homozygous and heterozygous mean? In the plants of Questions 13 and 14, which plants are which?
17. Imagine you are crossing two plants that have these two genotypes—Tt and tt. What are the two genotypes of the gametes that the Tt plant can make? What is the one genotype of the gametes that the tt plant can make? Set up a Punnett square to analyze the cross. Does the Punnett square need to have four boxes or just two? If you were crossing a TT plant with a tt plant, how many boxes would be necessary in a Punnett square?
18. Imagine you are given two tall plants, but you do not know their genotype (see Figure 16-12). There are three possibilities—they might both be TT or both Tt or one TT and the other Tt. Imagine you cross the two plants, wait for them to produce seeds, and then plant the seeds. If all resulting plants are tall, can you then determine what the genotypes of the two parents are? If approximately half the resulting plants are tall and about half are short, can you then figure out the genotypes of the parents?
19. In a field you find three types of the same plant: some with long leaves, some with short leaves, and some of intermediate length. What would you suspect to be the genotypes of each? You measure hundreds of plants and find that about half have leaves of intermediate length, whereas one fourth have long leaves and one fourth have short leaves. Does this support your estimate of the genotypes? What kinds of crosses would you do to test it further? In each cross, what types of gametes would each parent produce and in what ratios?
20. What is a test cross? Why do we have to use it? When we just look at a plant, can we see its genotype? Can we see its phenotype?
21. When traits show complete dominance, the genotype can be revealed by a test cross. In the example of tall and dwarf plants (Figure 16-14) why is it necessary to use the dwarf parent? In general, why is it necessary to use a parent that is homozygous recessive?
22. What does the term “multiple alleles” mean? Think about flowers such as roses that have many different colors. Do you think that flower color genes in roses might have multiple alleles? Now think about a protein like RuBP carboxylase in which all RuBP carboxylase molecules have the same amino acid sequence. We can extract this protein from almost any plant, and the structure will always be the same. Do you think that there are multiple alleles for RuBP carboxylase?
23. We say that genes that are close together on the same chromosome are linked. What does that mean? Imagine that in a diploid nucleus, one chromosome has the allele R and right next to it on the same chromosome is the allele T for a different gene. Are these linked? Imagine that r is right next to t on the homologous chromosome. If this cell never undergoes meiosis, does this matter at all? If it does undergo meiosis, what are the two genotypes that the gametes are most likely to have? Look at Figure 16-17 for help if you need it.
24. In Question 23, if one chromosome has R and T and the homologous chromosome also has R and T, does linkage matter? What is the single type of genotype that this plant can produce in its gametes?
25. The answer to Question 23 will be different if R and T are very far apart on their chromosome (and if that is true, then automatically r and t will also be very far apart on their chromosome). If it undergoes meiosis, there are four genotypes that can be produced, not just two. Why? What are the four genotypes? Which represent recombinant chromosomes? Which represent parental chromosomes? Look at Figure 16-18 for help if you need it.
26. Think about the same two genes that were mentioned in Question 23—R and T, present in the heterozygous condition as R and r, T and t. Now imagine that the gene for flower color (the alleles R and r) is not on the same chromosome as the gene for plant height (the alleles T and t). Because the two genes are on separate chromosomes, they show independent assortment. What does that mean? When this nucleus undergoes meiosis, it can make gametes with four different genotypes. Why? A very important aspect is that the four different genotypes will be formed in almost exactly equal numbers. Why?
27. Figure 16-17 shows a dihybrid cross but it can be analyzed as a monohybrid cross of just smooth seed coat (S) and wrinkled seed coat (s) with complete dominance that should show a 1:2:1 ratio. Examine Figure 16-17 and see if you can find these results. Likewise, it can be analyzed as a monohybrid cross of yellow seed coat versus green. Can you find these in the expected ratio?
28. What is maternal inheritance? Which characters show this type of inheritance?
29. Why is it easier to study plastid inheritance than mitochondrial inheritance? What is unusual about both? Do plastids and mitochondria undergo sexual reproduction? Syngamy?
30. Imagine that you have a plant with no chlorophyll. It is completely white because of a mutation in a plastid gene. Imagine that you cross this plant with an ordinary green plant. In one cross, you transfer pollen from the white plant onto the stigma of the green plant. After seeds form and you plant them, what color do you expect the seedlings to be—white, green, or pale green? At the same time as the first cross, you also transfer pollen from the green plant onto the stigma of the white plant. After you obtain these seeds and plant them, what color do you expect the seedlings to be? Will they be the same as in the first cross? Explain the results.
31. What is a lethal allele? Would you expect most lethal alleles to be recessive (that is, only ll is fatal), or would you expect many of them to be dominant (LL and Ll are both fatal)? Do you think a plant carrying a dominant lethal allele would survive very long? Do you think such a plant could become abundant in the population?
32. If a cell has one set of chromosomes, it is said to be haploid. If it has two sets, it is diploid. What term is used if it has three sets? Multiple sets?
33. Plants with multiple sets of chromosomes (polyploid plants) have more sets than they need. They are often healthy and vigorous. What might happen to their extra chromosomes and genes over long periods of time?
Design Credits: Hummingbird: © Tongho58/Moment/Getty; Green plant cells: © ShutterStock, Inc./Nataliya Hora; Purple tulip: © ShutterStock, Inc./Marie C Fields; Dandelion: © ShutterStock, Inc./danielkreissl; Poppy: © ShutterStock, Inc./Saruri; Plant icon: © ShutterStock, Inc./Vector; Digging man icon: © ShutterStock, Inc./Z-art