Botany: An Introduction to Plant Biology - Mauseth, James D. 2017
Genetics and Evolution
Population Genetics and Evolution

Chapter Opener Image: This small area contains many thousands of individuals of plants with yellow flowers (Physaria fendleri in the mustard family); millions more exist throughout the American Southwest. Each is very similar to many others, but no one is exactly identical to any other. We might decide to select one as a representative, but it will not represent the individuals that are taller or shorter, or those that have thicker leaves or thinner, or those whose flowers differ in color. Each individual has a particular set of features, but the species has all the features of every individual living now, that has lived, or that will live in the future.
OUTLINE
✵ Concepts
✵ Population Genetics
- Factors that Cause the Gene Pool to Change
- Situations in which Natural Selection Does Not Operate
- Multiple Selection Pressures
✵ Rates of Evolution
✵ Speciation
- Phyletic Speciation
- Divergent Speciation
- Convergent Evolution
✵ Evolution and the Origin of Life
- Conditions on Earth Before the Origin of Life
- Chemicals Produced Chemosynthetically
- Formation of Polymers
- Aggregation and Organization
- Early Metabolism
- Oxygen
- The Presence of Life
Box 17-1 Botany and Beyond: Species Are Populations, Not Types
Box 17-2 Plants and People: Zoos, Botanical Gardens, and Genetic Drift
LEARNING OBJECTIVES
After reading this chapter, students will be able to:
✵ Describe how the discovery of geological evolution affected our understanding of the Earth.
✵ Define population genetics.
✵ Discuss the relationship of a gene pool to sexual reproduction.
✵ List four factors that can cause a gene pool to change.
✵ Name the two conditions that must be met for natural selection to occur.
✵ Compare phyletic speciation to divergent speciation.
✵ Name three ways gene flow occurs.
✵ Identify the two fundamental causes of reproductive isolation.
✵ Give an example of a biological reproductive barrier.
✵ Explain convergent evolution.
✵ Summarize the conditions that made life on Earth possible.
 Did You Know?
Did You Know?
✵ People used to think of species as being a set of uniform individuals, just as all hydrogen atoms are identical.
✵ But we realize that even though all people are the same species, Homo sapiens, there is variability of individuals; the same is true in plants. Individuals of a species are neither identical nor interchangeable the way hydrogen atoms are.
✵ A mutation is usually important only if it affects a cell that produces spores or gametes such that the mutation can be passed to subsequent generations.
✵ The individuals of a species may be spread over thousands of square kilometers, and a mutation in one area has no immediate impact on individuals located far from it, even if the mutation is beneficial or harmful.
![]() Concepts
Concepts
Evolution is the gradual conversion of one species into one or, in some cases, several new species. It occurs for the most part by natural selection: Mutations cause new alleles or new genes to arise that affect the fitness of the individual, making it less or more adapted to the environment than other individuals without the new allele or gene. If the mutation is deleterious, the individual may grow or reproduce slowly or even die early without reproducing. Either way, the new allele has less chance of increasing in the population and is more likely to be eliminated as the individuals that carry it die out. If the mutation is beneficial, the individual should grow and reproduce better than other individuals, producing a greater number of gametes and ultimately a greater number of seeds than the other individuals. The abundance of the new allele increases relative to the original alleles (FIGURE 17-1). As evolution by natural selection continues, the types and abundances of alleles present in the species change, and consequently, the phenotype also changes.
Evolution is an extremely slow process that may require thousands of generations and millions of years to produce obvious changes in a species. Because it is so slow compared with the length of a human lifetime, it is easy to understand that it went undetected until recently, just as continental drift, mountain building, and valley formation by erosion went unnoticed. A human lifetime is too short to perceive changes in these processes; even the full history of accurate observation and detailed record keeping is not long enough to detect the fact that, since the times of the ancient Chinese, Sumerians, and Egyptians, continents have moved closer together, mountains have risen higher, and species of plants and animals have changed. It is not surprising that people thought in terms of a constant, unchanging world composed of specific mountain ranges, rivers, lakes, continents, plants, and animals.
During the explorations of Africa, the Americas, and Australia near the time of Columbus, so many plants and animals were discovered that people could not help but notice remarkable similarities between many species. Various types of roses resembled each other more than they resembled other plants, just as did many new types of lilies, orchids, and so on. People began to realize that giant basins of sediment, such as the deltas at the mouth of the Nile, the Ganges, and the Mississippi, held enough eroded material to account for entire mountain ranges; the concept of a changing landscape—geological evolution—began to develop (FIGURE 17-2). An important corollary was the realization that in order for so much sediment to have formed, Earth must be millions (actually billions) of years old, not merely 6,000 years old, as had been calculated from the genealogy of the Book of Genesis in the Old Testament (see the timeline diagram on the inside of the back cover). This is one of those discoveries that is easy for us to underappreciate; the quantity of sediments proved that either Earth had a very long history or that God had created sediments. This gave theologians a difficult dilemma and caused many people to be even more skeptical of priestly interpretations of natural phenomena. One of the areas of doubt was the idea that all species had been created at once. Everyone was vividly aware that the Church already had made a grave error by censuring Galileo, insisting that Earth was the center of the universe and that the sun, planets, and stars all revolved around it (FIGURE 17-3).
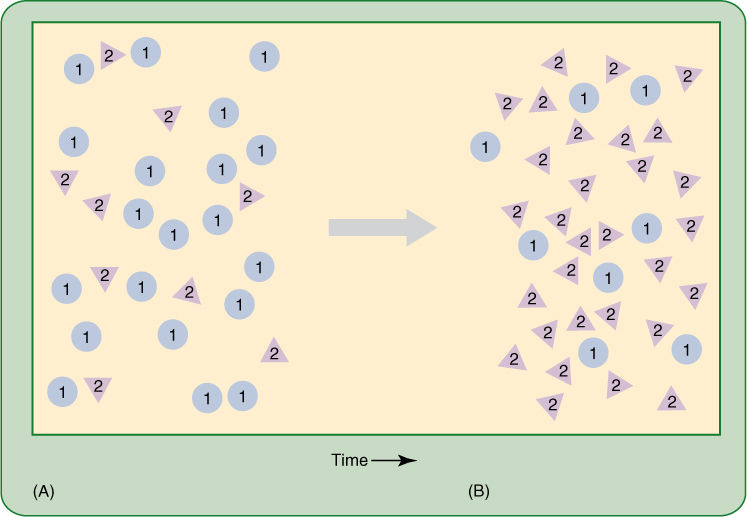
FIGURE 17-1 (A) The population in this diagram originally consisted of 29 individuals, 20 of which carried allele 1 and 9 of which carried allele 2. Allele 2 produces individuals that are more vigorous, and thus, allele 2 has increased in the next generation (B) from 9 of 29 (31%) to 30 of 38 (78.9%). Allele 2 produces a phenotype (triangle) distinct from allele 1 (round); therefore, as the allele frequency of the population changes, so does the phenotype. In this diagram, the extra vigor of allele 2 is allowing the population as a whole to enlarge, from 29 to 38 individuals, perhaps by crowding out other species or by entering new habitats.
As careful observations of nature became common after the Renaissance and as the scientific method proved to be an accurate method for analysis, the concept of the evolution of species became widespread. Scientists searched for an understanding of the mechanism by which it occurred. Finally, Alfred Russel Wallace and Charles Darwin independently discovered the basis—natural selection—in the mid-1800s, the critical explanation being given in Darwin’s Origin of Species published in 1859. The concept of natural selection—survival of the fittest—had an electrifying effect on all biologists and led to a revolution in all aspects of thinking. Suddenly, many observations had a rational explanation; every character could be interpreted in terms of whether it made a species less or more adapted to its environment. And because so many environments exist, it is logical to expect that many types of plants and animals should also exist, each having genotypes and phenotypes that make it particularly adapted to its own environment. Before the discovery of natural selection, flowers were thought to have been created by God for the delight of the human eye and nose; that philosophy did little to explain bilaterally symmetrical flowers, inferior ovaries, or wind pollination. Science had been greatly hampered by trying to interpret the world in terms of the mind of God and what had been created to feed, clothe, and house the descendants of Adam and Eve.

FIGURE 17-2 As the science of geology developed, it was soon realized that river deltas, such as this delta of the Mississippi, contained so much sediment that they must represent the transformation of landscapes, that mountain ranges must have eroded and no longer exist. This means that the world is not constant, rather it is changing and evolving.
Fortunately, the discovery of evolution by natural selection happened at about the same time as other critical discoveries—the discovery of genes and chromosomes, cell theory, proving that spontaneous generation does not occur, sophisticated hydroponic cultures of plants, the discovery of enzymes and the carrying out of some metabolic steps in a test tube without living cells, and the artificial synthesis of biological compounds. All of these combined to move biology firmly out of the realm of metaphysics/theology and into that of scientific analysis and interpretation.

FIGURE 17-3 Galileo Before the Inquisition. Kepler, Newton, and Galileo proved that the sun is the center of the solar system, but church doctrine considered this heresy; as the home of the one organism created in the image of God, the Earth should be the center of the Universe. The Inquisition forced Galileo to recant his belief in a sun-centered solar system and placed him under house arrest for the rest of his life. Attempts were made to confiscate and burn all copies of his book; only a few survived. Less famous scientists were frequently punished more severely. Misinterpretation of the physical world by religious doctrine is not a fundamental aspect of religion, but rather the rejection of observation and experimentation in favor of speculation without confirmation.
For those of you who believe in divine creation rather than evolution by natural selection, keep in mind that all of our experience has shown the scientific method to be greatly superior to theological interpretation of scriptures for understanding and predicting the nature of the physical/biological world. Equally important, only ethical/philosophical systems allow us to understand and solve moral problems; science cannot do that. It is not logical or consistent for a person to accept certain discoveries of science (photosynthesis, DNA, respiration, vessels, sieve tubes) while rejecting others (evolution) if one type of discovery is just as well documented as the other.
![]() Population Genetics
Population Genetics
In this chapter we discuss the genetics of groups of individuals of one species. Population genetics deals with the abundance of different alleles within a population and the manner in which the abundance of a particular allele increases, decreases, or remains the same with time. The genetic recombination that occurs during sexual reproduction is important only if the two sexual partners have differing genotypes. A cross between two plants that are A1A1B1B1 × A2A2B2B2 produces offspring that have the genotype A1A2B1B2, which is different from that of either parent. Crossing-over also increases genetic diversity in populations.
Botany and Beyond
BOX 17-1 Species Are Populations, Not Types
Our concept about what a species is has undergone a major change. We now think of a species as being all the individuals that can interbreed with each other. For example, think of our own species, Homo sapiens: It consists of every living human individual on Earth. We could all interbreed if given the chance; we can all exchange genes. No single person is more human than any other. Now think of all the people who have lived recently, and those who lived centuries ago: We are all similar to each other, we all together make up the species Homo sapiens.
What are the characters of our species? For each character, we must give a range of values. Adult height: from 1 m to 2.5 m. Skin color: white, brown, black, yellow, and more. Blood type: A, B, AB, O. Similar ranges of variation occur in all species, and it’s just as true of plants as it is of us. This is the population concept of species.
Until the late 1800s, our concept of species was based on types, the type concept of species. Biologists and others thought in terms of one single ideal specimen—the type—that would represent a species. The concept had several foundations. One was the biblical creation story in Genesis, which states that God created all the types of organisms. Another was Plato’s concept that every type of thing on Earth was an imperfect manifestation of a perfect ideal. For example, we cannot draw a perfectly straight line, but we can conceive of a hypothetical line that is perfectly straight. Every straight line we draw is a variation of the ideal line. The third foundation is our tendency to summarize and generalize, to think of an average as representative. Clothes, cars, bicycles, and computer keyboards are all designed for people with average dimensions.
After the Dark Ages, naturalists began studying nature avidly. They collected and named plants, animals, mushrooms, rocks, almost everything. Obviously, every slight variation of poppies could not be given a separate name, so each name came to represent an ideal but variable species. This variation was perplexing. Some people postulated that God had created variable species. Others thought that God had created only invariable species but allowed variation to occur. Still others believe that variation was due to the imperfect nature of the material world, as opposed to the perfect nature of an ideal world, but if so, why are chemical elements and compounds invariable? Poppies are variable but sodium chloride is not. Variation even perplexed Gregor Mendel and Charles Darwin: Mendel’s experiments in genetics seemed to show that genes do not change during sexual reproduction, and thus living organisms should not change with time. Darwin’s theory of natural selection required variability and change.
As biologists studied the conflict between Mendel’s theory and that of Darwin, the answer became clear. Genes are almost as stable as Mendel had postulated, but they occasionally undergo mutations, and these mutations produce the variation that Darwin’s natural selection requires. Also, there are vastly more genes then people had realized at first: If each individual is the product of thousands of genes, and if each gene has several alleles, then a species can be extremely variable. Furthermore—and this is the crucial point—with all this variation, every individual matters, every individual is just as representative as any other. There is no ideal, there is no type. With this population concept of species, it is easy to see how a species evolves as some of the millions of individuals survive a bit better than others. Evolutionary change will be gradual. In contrast, under the type concept of species, each species was pictured as being a discrete entity distinctly separate from every other species, so the evolution of one species into one or several others would require sudden jumps, sudden large changes in many characters.
This change of our concepts from types to populations affects all aspects of biology. People had thought in terms of discrete genes and enzymes, for example, of one single gene for amylase and therefore of one single form of that enzyme. But now we know genes are variable not only because of the different alleles they have, but also because they occur in gene families that have evolved from ancestral genes. Similarly tracheids vary in length, width, and wall thickening, as do vessel elements, and it is easy to see how some tracheids could evolve into vessel elements. But if instead all tracheids in all plants were absolutely identical, if all were exactly the same as some hypothetical ideal tracheid, then it would be difficult to see how any could ever change or evolve and give rise to vessel elements.
The total number of alleles in all the sex cells of all individuals of a population constitutes the gene pool of the population. Imagine that gene A has four alleles: A1, A2, A3, and A4. If the population consists of 1 million individual plants, each with 100 flowers and each flower producing on average 100 sex cells (sperm cells and egg cells), the gene pool contains 10 billion haploid sex cells. The alleles are probably not present in equal numbers; for instance, 60% of all gametes may be A1, 20% A2, 15% A3, and 5% A4. Will this ratio be the same for the population next year or in the next generation? Early in the 20th century, two biologists, G. H. Hardy and G. Weinberg, demonstrated mathematically that if only sexual reproduction is considered, the ratio remains constant over time; even if an increase or decrease occurs in the total number of individuals or gametes, the ratios do not change. Sexual reproduction alone does not change the gene pool of a population; if no other factors were involved, the gene pool would remain constant forever.
Factors that Cause the Gene Pool to Change
Although it is possible theoretically for a gene pool to remain constant, in reality, changing allele frequencies are the rule because populations are always affected by factors other than sexual reproduction.
Mutation
All genomes are subjected to mutagenic factors, and mutations occur continually. Because of mutation, existing alleles decrease in frequency, and new alleles increase. Whether or not mutation is significant depends in part on the population’s size. In the population described earlier, there were 10 billion copies of gene A, and 60% were the allele A1. Therefore, there were 10,000,000,000 × 0.6 = 6,000,000,000 copies of A1; if one mutates to become a new allele, A5, then the frequency of A1 drops to 5,999,999,999, which is still pretty close to 60%. A5 exists as just one copy out of 10 billion, and thus, its frequency is extremely small; however, the presence of one copy is an infinite increase over zero copies, and the existence of even a single copy should not be ignored.
Accidents
Accidents are events to which an organism cannot adapt, such as the collision of a large meteorite with Earth. When a meteorite of sufficient size strikes Earth, a large region of Earth’s surface is destroyed, killing all life in the area. All organisms, along with all their alleles, are eliminated. If by chance the area of impact had included the single individual that had just obtained A5 by mutation, then A5 would no longer exist. If the population in the impact area has the same gene frequencies as the general population, the alleles are eliminated in the same proportions as they exist generally, and no change in allele frequency occurs. If the impact area had an unusually high number of a particular allele, for instance A3, then A3 is affected more than the other alleles; therefore, the allele frequency of the gene pool of the survivors is altered. This change of allele frequency, however, does not make the species any more able to survive another hit by a meteorite.
Many phenomena qualify as accidents. A volcanic eruption produces poisonous gases and molten rock that destroy everything within a limited area. Infrequent floods, hailstorms, or droughts can act as accidents for plants too small and delicate to become adapted to those events; all individuals in the affected area are killed. For other species that consist of plants with larger bodies, such natural phenomena act as selective forces, removing the weaker, less well-adapted individuals but not affecting the more well-adapted members (FIGURE 17-4). The continental drift of Antarctica southward from a temperate region to the South Pole was an accident for all the plants living on it. While located in the temperate latitudes, Antarctica had a rich flora with abundant plant life; as it drifted southward, it entered a region too cold and severe for any plant life to survive. All individuals and their alleles were eliminated (FIGURE 17-5).
Accidents can be small events as well as large ones. After an allele is formed, A5 for instance, its numbers may increase as it is used in sexual reproduction; after a few years, there may be 10 individuals with the A5 allele. The individuals are closely related and probably located close together because most seeds do not travel very far. It is possible for a local disturbance to eliminate all 10: an avalanche, a herd of grazing animals, or the construction of a highway.

FIGURE 17-4 Severe drought and heat across all of Texas in 2011 resulted in many wildfires in all parts of the state. (A) A fire in a pine forest near the East Texas town of Bastrop killed almost all trees, none of which appears to have been adapted to survive fire. Because all trees were killed, this fire was not a selective force; there was no differential survival. (B) A fire near the West Texas town of Fort Davis damaged many trees, shrubs, and succulents but seems to have killed only a few. These plants with long leaves are sotols (Dasylirion leiophyllum), and although all were burned severely, many actually survived; however, others were killed completely. If the ones that survived differed genetically from those that died (perhaps they had genes for thicker bark or more heat-resistant organs), then the fire was a selective force: The gene pool after the fire will be different from what it was before the fire.

FIGURE 17-5 In earlier times, Antarctica was located in temperate latitudes and supported many plants, such as this Glossopteris. Continental drift moved Antarctica to the South Pole, into conditions for which no alleles are able to produce an adapted phenotype. All plants perished.
Artificial Selection
Artificial selection is the process in which humans purposefully change the allele frequency of a gene pool. The most obvious examples are the selective breeding of crop plants and domestic animals (FIGURE 17-6). Plant breeders continually examine both wild populations and fields of cultivated plants, searching for individuals that have desirable qualities such as resistance to disease, increased protein content in seeds, and the ability to survive with less water or fertilizer. When plants with beneficial qualities are found, they are collected and used in breeding programs to produce seed for future crops. Consider just wheat, rice, and corn: Almost the entire world populations of these three species consist of cultivated plants; very few of the natural ancestors still exist in the wild. Consequently, the gene pool for each is made up almost entirely of alleles that have been artificially selected for thousands of years.
Artificial selection is also used to produce ornamental plants that flower more abundantly or for a longer time. Artificial selection has also been used to alter flower color and size and to make the plants hardy in regions where they otherwise could not grow. The trees cultivated for lumber and paper are also subjected to artificial selection.
Artificial selection is often carried out in conjunction with artificial mutation. Plants are exposed to mutagens such as acridine dyes or irradiation with ultraviolet light or gamma rays to increase the number of new alleles that come into existence. The plants are allowed to grow to see how the new alleles affect the phenotype, and those plants with the desired phenotypic traits are used in selective breeding programs.

FIGURE 17-6 Artificial selection by crop breeders has resulted in increased frequency of certain alleles and elimination of others. The allele frequency of the population changes dramatically from year to year, depending on whether farmers decide to grow one variety or the other. Cabbage (A), broccoli (B), and cauliflower (C) are all the same species (as are brussels sprouts), and all evolved from the same ancestor; however, their evolution was controlled by artificial selection for certain traits in each variety. Recently, artificial selection has produced the variety Violet Queen that stores large amounts of pigment (D).
Natural Selection
Natural selection, which is the most significant factor causing gene pool changes, is usually described as survival of the fittest: Those individuals that are most adapted to an environment survive, whereas those less adapted do not. However, natural selection is such an important factor in evolution that it must be given careful attention. Two conditions must be met before natural selection can occur:
1. The population must produce more offspring than can possibly grow and survive to maturity in that habitat. This condition is almost always valid for plants anywhere on Earth. Most plants produce hundreds of seeds, which often germinate near the parent plant (FIGURE 17-7). Even in species with wind-dispersed seeds, such as maples or milkweeds, most seeds do not travel far. Consequently, the ground can be covered with hundreds of seedlings crowded closely together, and there simply is not enough room to accommodate the physical bulk of so many plants as they grow.
Besides limited resources, the number of individuals that can survive in a particular habitat is affected by predators, pathogens, and competitors. All plants are faced with attack by herbivorous animals, ranging from almost microscopic mites and nematodes to much larger beetles, reptiles, birds, and mammals. Animals not only eat plants but may also lay eggs in them, bore into tree trunks for nesting sites, walk on them, and rob nectar without carrying out pollination. Pathogenic fungi and bacteria are similarly harmful. Competitors are other organisms that use the same resources the plant needs to survive. When root systems grow together, the two plants compete for the same water and nutrients. If two species are pollinated by the same species of insect or bird, they must compete for the attention of the pollinators. In a forest, plants compete for light: Those that can grow tallest receive the most light; those that have shorter trunks receive only dim light. All of these activities adversely affect the plant’s ability to reproduce and may cause the plant’s death.

FIGURE 17-7 Even if animals carry away many of the fruits of this red elderberry (Sambucus racemosa), most fall and germinate in its immediate neighborhood, and there are too many seedlings to fit into the space physically. Also, the tree will produce seed abundantly for many years.
2. The second condition necessary for natural selection is that the progeny must differ from each other in their types of alleles (FIGURE 17-8). If they are all identical, all are affected by adversity in the same way and to the same degree. Under crowded conditions, probably all are stunted similarly, all grow poorly, and finally, none reaches reproductive maturity. If all individuals of a species are equally susceptible to a pathogenic fungus, no increase in survivability, and fitness occurs as the result of a fungal attack. Even if some survive and reproduce, they are identical genetically to those that died, so no change occurs in allele frequency; natural selection has not occurred.
When genetic diversity exists among individuals, differential survival can occur. If some members of the population have an allele that gives them increased resistance to fungi, those plants should fare much better during an outbreak of fungus in the population than those lacking the allele. If the fungus is so virulent that it often kills the plants it attacks, the allele frequency of the population is changed radically after infection—the resistance allele constitutes a much greater percentage of the gene pool. Natural selection operates even if the fungus only weakens plants but does not kill them outright; the weakened plants should produce fewer seeds than do the resistant, healthy plants.

FIGURE 17-8 The summer of 1988 produced one of the most severe droughts on record in the American midwest, our primary corn-growing area. Under natural conditions, such a drought would have resulted in the natural selection of drought-resistant plants; all corn plants of 1989 would have been the progeny of the resistant plants that survived, and all would have carried alleles for drought resistance. Natural selection did not occur, however, because there was no genetic variability among the corn plants; corn is one of our most highly inbred crops, and all seed corn is produced by careful crosses of absolutely uniform parents. Consequently, of the trillions of plants affected by the drought, none was more or less resistant than any other. Any that survived did so not by genetics, but by accident: receiving a little extra rain or being in a valley where extra moisture collected.
Competition for water and nutrients among crowded root systems also acts as a selective force; alleles that allow roots to absorb water and nutrients more effectively have an advantage. If most roots can no longer absorb water when the soil has a water potential of —0.8 MPa, an allele that alters root metabolism such that it can extract water even at —1.0 MPa enables the plant to grow when others cannot. The mutation has a selective advantage—the plant is more fit or adapted to conditions of crowding.
Natural selection refers to the differential survival among organisms that have different phenotypes. Natural selection can act only on preexisting alleles; it does not cause the mutations. The presence of a fungus does not cause plants to become resistant; if none had been resistant before, all individuals would be adversely affected. The advantageous allele must exist first. But if an allele for resistance does exist, natural selection can cause the population to become resistant by the preferential survival of resistant individuals, even though it cannot cause an individual to become resistant. A population or species evolves, but an individual does not. Similarly, a population may become more adapted but a single individual does not change; it cannot become more adapted during its lifetime. An attack by a fungus may stimulate a plant to activate genes that produce antifungal chemicals, but this is a case of causing a plant to express genes it already has; it is not creating new genes or alleles.
Natural selection does not always result from the action of an agent outside the organism. Although many selection pressures are external—pathogenic fungi and dry, hot climate—any factor that causes one plant to produce more progeny than other plants is a selective factor (FIGURE 17-9). If an allele causes chloroplasts to photosynthesize more efficiently, plants with that allele can produce carbohydrates more rapidly than plants that lack the allele; the former plants grow faster and produce more seeds, at least half of which carry the advantageous allele.
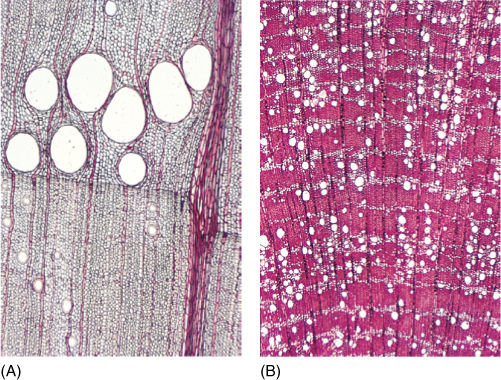
FIGURE 17-9 (A) This oak wood contains wide vessels that can conduct water with relatively little friction but which cavitate easily (×80). (B) This Casuarina wood has vessels so narrow that the water in them is strongly reinforced and cavitation is almost impossible under natural conditions. Even though vessels are completely internal, they still affect the plant’s survival relative to its environment (×80).
Factors that Are Not Part of Natural Selection
Natural selection can be understood more clearly and accurately if you realize that certain factors are not part of natural selection at all: Natural selection does not include purpose, intention, planning, or voluntary decision making. Whenever we say that “plants do something in order to . . . ,” we are suggesting that the plants can plan their activities and have purpose, which is not true. It is not correct to say that certain plants have disease-resistance genes in order to protect themselves against fungi. The resistance of a population of plants to a fungus is nothing more than the result of the preferential survival of the plants’ ancestors because they had an allele for resistance, whereas their competitors did not. Although plants that have this allele in the presence of the fungus have a selective advantage, the plants do not have the allele in order to protect themselves. Similarly, plants do not produce nectar in order to attract pollinators; those plants in the past that did produce nectar happened to be pollinated more often than others that did not produce nectar, so the alleles for nectar production were increased in the population. Currently, those plants that secrete nectar are visited by pollinators, but there is no purpose, intent, or planning by the plant. Only humans and other primates act with intent and purpose, and this applies only to our voluntary actions. Even humans do not digest food in order to have an energy supply; rather, our autonomic nervous system and cell metabolism have automatic responses to the presence of food in the small intestine that cause the secretion of digestive juices, the absorption of monomers, their distribution through the hepatic portal system, and their respiration by cytoplasm and mitochondria. We have no control over this result of our evolutionary history. This may seem to be just a trivial problem of semantics because everyone knows what we mean, but statements should be accurate, not sloppy. If we are not meticulous in how we express our ideas, we will not be meticulous in how we think, and important details will be lost.
Situations in which Natural Selection Does Not Operate
Further understanding of natural selection can be gained by considering several cases in which it does not operate. As already mentioned, it cannot operate if all individuals of a population are identical genetically or if it is impossible to become adapted to a certain condition. Competition does not occur in a habitat that can support the growth and reproduction of all individuals; if survival is universal, natural selection does not occur. Situations like this occur in newly opened habitats such as a plowed field. All seeds present may germinate and grow vigorously, even the ones not well adapted for competition. Because all survive, no natural selection has occurred. Other examples are the sides of a road cut, a recently burned area, or a recently flooded plain covered with rich sediments. We must be careful here; if the road cut passes through a heavy, dark forest, the newly exposed sides may be too bright and exposed for seedlings from plants adapted to the forest shade. In this case, the environment favors those plants that can tolerate full sunlight and suppresses those that require shade; many selection pressures, but not all, have been eliminated, and some natural selection can still occur.
Multiple Selection Pressures
In many cases, the loss of individuals and reduced reproduction are not caused by a single factor, such as a pathogenic fungus. Instead, the plants are also affected by insect attack, drought, cold, need for pollinators and need for a mechanism to disperse their seeds, as well as the efficiency of their own metabolism, such as the ability of their membranes to pump ions, the capacity to reduce nitrogen, or the efficiency of producing just enough P-protein in the phloem without a wasteful, nonuseful excess.
A mutation that produces an allele that would result in improved fitness is potentially advantageous selectively, but it may never have the opportunity to improve the fitness of the plant or the species. A mutation that results in improved cold hardiness may be eliminated from the gene pool if the plant carrying the new allele is killed by fungus or drought or cannot reproduce because of poor competition for pollinators. Such a loss of this allele is simply an accident.
However, if the new allele for cold hardiness does survive it may be able to improve the species. If cold winters are common, this allele greatly improves fitness, and its frequency may increase rapidly. If cold winters are infrequent, they do not exert a strong selection pressure, the allele does not improve fitness very much, and its frequency may remain low for years until a harsh winter does occur. Until that time, the allele’s frequency is determined by several factors. It may be tightly linked to an allele that is strongly advantageous for an important condition. For instance, the cold hardiness allele may be on the same chromosome as the allele for resistance to fungi. If the two genes are so close on the same chromosome that crossing-over virtually never occurs between them in prophase I, the presence of fungi selects not only for the antifungal allele but also for the cold hardiness allele, just by coincidence.
The cold hardiness allele may affect the plant in various ways besides the ability to withstand cold; that is, pleiotropic effects may operate. If these are also advantageous under common conditions, the allele increases in frequency, but often many of the “side effects” are disadvantageous. Many improvements to phenotype have some negative aspects, at least in terms of cost. Increased cold hardiness may be due to thicker, more sclerified bud scales with a thick layer of wax. These require the input of increased amounts of nutrients and energy that could have been used to produce more seeds. Thus, whereas this allele may be strongly advantageous in an environment with frequent cold winters, it may be disadvantageous in an environment where winters are always mild. Whether a particular allele is beneficial or not depends entirely on the habitat, which may change with time. As the environment changes, the selection pressures change and certain features become more or less advantageous.
![]() Rates of Evolution
Rates of Evolution
From the examples given, it seems that the allelic composition of a population could change rapidly, within a few generations, but that is not typically the case. Most populations are relatively well adapted to their habitat, or they would not exist. Very few mutations produce a new phenotype so superior that it immediately outcompetes all other members of the population. At the extremes, there are many species of seedless plants (lycopods, Equisetum, ferns) that have persisted for tens of millions of years without diverging into new species (FIGURE 17-10). In contrast, very rapid speciation is occurring in a group of asters in Hawaii: Shortly after Kauai formed 5.2 million years ago, an aster seed arrived, thrived, reproduced, and spread rapidly in the unpopulated island. Its descendants have diversified into three distinct genera, each with many species, and on average, a new species arises in this group once every 500,000 years.

FIGURE 17-10 Plants of Equisetum have jointed stems, often with whorls of branches at each node; they are easily recognizable, even as fossils. This group originated millions of years ago and had many species at one time, most of which are extinct. There are now only 15 species of living plants, and even though many are widespread and well-adapted to many areas, they do not appear to be evolving into new species.
It is difficult to identify the presence of particular alleles in a population unless they result in an easily identifiable effect on the phenotype. Consequently, most studies of evolution concern the changes in gross structures such as flowers, leaves, fruits, shoots, and trichomes. But these complex structures are the product of the developmental interaction of many genes. Any new mutation results in a more adaptive structure only if the effects of the new allele fit into the already existing highly integrated mechanism of morphogenesis without causing serious disruptive effects. As systems become more intricate, the probability decreases that any random change is beneficial.
Evolutionary changes that result in the loss of a structure or metabolism can come about quickly, however, and for the same reason: complexity. If a feature becomes selectively disadvantageous, many of the mutations that disrupt its development become selectively advantageous. Because disruptive mutations outnumber constructive mutations, loss can occur relatively rapidly. For example, the ancestors of cacti lived in a habitat that became progressively drier; large, thin leaves were advantageous because they carried out photosynthesis but were disadvantageous because too much water was lost by transpiration (FIGURE 17-11). Mutations that disrupted formation of the lamina were advantageous, and cacti lost their leaves in perhaps as little as 10 million years, whereas the evolutionary formation of leaves in seed plants had required over 200 million years. Leaves could not be lost too quickly, however, because the plants would be left with virtually no photosynthetic surface area. Mutations that caused the complete absence of leaves could not be selectively advantageous until other mutations had occurred that permitted the stem to remain green and photosynthetic, that prevented the early formation of an opaque bark, and that slowed the metabolism of the plant to a level compatible with reduced photosynthesis. The loss of leaves could occur only simultaneously with or after these modifications of the stem.

FIGURE 17-11 (A) The ancestors of the cactus family were large, woody trees with rather ordinary dicot leaves. The cactus genus Pereskia still contains members quite similar to the ancestors, as shown here. Apparently few genes had been modified by the time Pereskia appeared. (B) This Gymnocalycium is also a cactus, but its phenotype is significantly different from the ancestral condition; apparently all critical genes involved in leaf production have mutated so much that they are nonfunctional or absent. Genes involved in stem elongation now produce short stems, and this species contains genes for succulence that were not present in the ancestral Pereskia-like species; these new genes probably are highly mutated forms of “extra” genes from a tetraploid ancestor or arose by other methods of gene duplication.
![]() Speciation
Speciation
As natural selection operates on a population for many generations, the frequencies of various alleles and consequently the phenotype of the population change. At some point, so much change has occurred that the current population must be considered a new species, distinct from the species that existed at the beginning. Natural selection has caused a new species to evolve, a process called speciation.
At what point can we conclude that a new species exists? It is not possible to give an exact definition of species that is always valid, but generally, two organisms are considered to be members of distinct species if they do not produce fertile offspring when crossed. Many exceptions exist, however. The individuals of maples in the western United States cannot naturally reproduce sexually with those in the eastern United States, but that is only because the pollen does not travel that far; if an eastern plant is brought close to a western one artificially, they can cross-fertilize and are, therefore, considered the same species. On the other hand, many orchid species grow together in nature without producing hybrids, but artificial cross-breeding may produce healthy, fertile hybrids. Nevertheless, they are considered distinct species because they look different from each other. So, if two plants freely interbreed in nature, they are members of the same species; if they do not interbreed even when manually cross-pollinated, they are separate species, and if they do not interbreed naturally but they do artificially, it is a judgment call on the part of the scientists who work with the plants and are most familiar with them.
Speciation can occur in two fundamental ways: (1) phyletic speciation, in which one species gradually becomes so changed that it must be considered a new species (FIGURES 17-12A and B), and (2) divergent speciation, in which some populations of a species evolve into a new, second species while other populations either continue relatively unchanged as the original, parental species or evolve into a new, third species (FIGURES 17-12C and D).
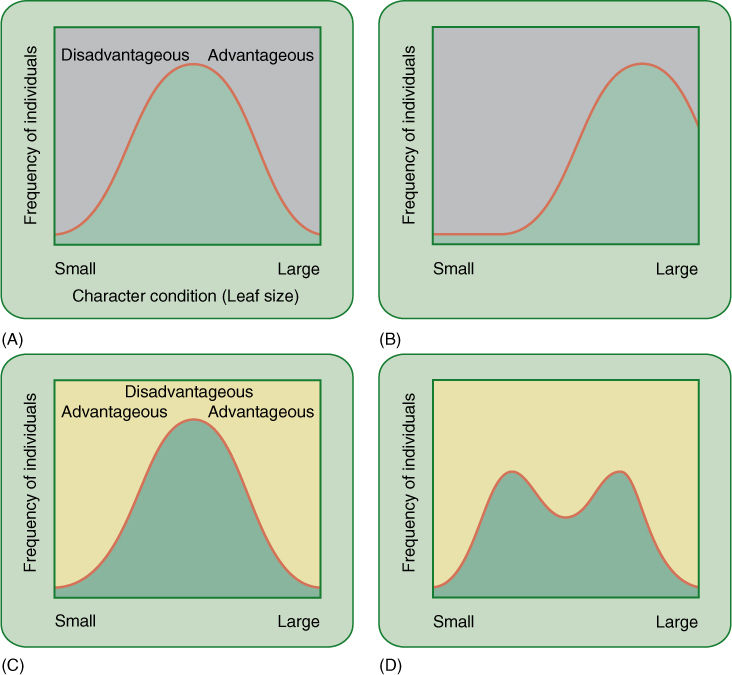
FIGURE 17-12 (A) and (B) In phyletic speciation, all of a species gradually changes because one particular aspect of a character is advantageous for all individuals. Here, all leaves become larger; perhaps the climate is becoming more humid or herbivorous insects are less of a problem. (C) and (D) In this scenario of divergent evolution, both extremes of the condition are more advantageous than intermediate values; the climate may become drier in some areas, favoring the smaller leaves, but moister in other areas, favoring plants with larger leaves.
Phyletic Speciation
Millions of years are often required for a species to evolve into a new one. The critical feature is that as new beneficial alleles arise and are selected for, they become spread throughout the entire population. This movement of alleles physically through space, called gene flow, occurs in many ways, such as by pollen transfer, seed dispersal, and vegetative propagation.
Pollen Transfer
Pollen grains each carry one full haploid genome, and all alleles of a plant are present in its pollen grains. Wind-distributed pollen, such as that of ragweed, grasses, and conifers, can travel great distances. If a new allele is carried by some of the pollen grains, it can move to very distant plants; if the pollen grain’s sperm cells fertilize an egg, a new seed is formed whose embryo contains the new allele.
Animal-mediated pollination also contributes to gene flow; both birds and insects tend to spend most of their time in a small area; therefore, although birds can fly long distances, they usually spread pollen through a smaller area. Nevertheless, allele movement can be rapid.
Seed Dispersal
The fruits and seeds of some plants fall close to the parent, but many species have long-distance dispersal mechanisms. Seeds and fruits can be carried by wind, floods, and stream flow. They can be carried to islands by rafting, in which they are trapped above water on a tangled mat of floating debris. Seeds or fruits that are spiny or gummy stick to the fur or feathers of animals; migratory animals can be especially important in dispersing seeds (FIGURE 17-13). Most birds reduce their weight before migration by preening themselves of all adhesive seeds, but if just one seed is overlooked, its alleles are transferred; as the seed germinates, grows into a new plant, and reproduces, the new alleles can be spread throughout the new site.
Vegetative Propagation
If a species produces small, mobile pieces that reproduce vegetatively, these too contribute to gene flow.
If these various mechanisms are sufficient to enable alleles that arise in one part of the species’ range to travel to all other parts, the species remains relatively homogeneous, even as the entire species evolves into a new species. Alleles that arise at various geographic sites ultimately come together by gene flow; then meiosis, crossing-over, and genetic recombination rearrange them into thousands of combinations (FIGURE 17-14).

FIGURE 17-13 After the hooks on this cocklebur fruit tangle in fur or feathers, the fruits will be carried far from the parent plant. When the seeds inside germinate and grow into new plants, their genes can combine with the genes of other cockleburs that are also in the new location.
Divergent Speciation
If gene flow does not keep the species homogeneous throughout its entire range, divergent speciation may occur; if alleles that arise in one part of the range do not reach individuals in another part, the two regions are reproductively isolated.
Reproductive isolation can occur in many ways, but the two fundamental causes are abiological and biological reproductive barriers.
Abiological Reproductive Barriers
Any physical, nonliving feature that prevents two populations from exchanging genes is an abiological reproductive barrier. The original species is physically divided into two or more populations that cannot interbreed; if speciation results, it is called allopatric or geographic speciation. Mountain ranges are frequently reproductive barriers because pollinators do not fly across entire mountain ranges while feeding or gathering pollen. Seeds may occasionally be carried across mountains by birds or mammals, but probably too rarely to be significant. Rivers are often good barriers for small animals, but they rarely prevent plant gene flow by means of seed dispersal. Deserts and oceans are effective barriers. Plants adapted to the harsh conditions of mountain tops are reproductively isolated from plants on adjacent mountain tops by the intervening valleys because pollinators and seed dispersers do not often travel from mountain to mountain (FIGURES 17-15). Any seeds dropped into the valley would not be able to compete with lowland plants and would die. Ultraviolet (UV) light and dry air are barriers to very long-distance wind dispersal of pollen; during the ride on wind, pollen is damaged by UV light and dry air and often dies before it travels far.
Biological Reproductive Barriers
Any biological phenomenon that prevents successful gene flow is a biological reproductive barrier. Differences in flower color, shape, or fragrance can be effective barriers if the species is pollinated by a discriminating pollinator. A mutation that inhibits pigment synthesis and causes a normally colored plant to have white flowers might prevent the pollinator from recognizing the flower. The flower is not visited, and gene flow no longer occurs between the mutant plant and the rest of the species even though the individuals grow together. Timing of flowering can be important: If some flowers open in the evening, they probably do not interbreed with those that open in the morning. Differences in flowering date are critical because of the brief viability of pollen after it leaves the anthers. When two groups become reproductively isolated even though they grow together, the result is sympatric speciation.
For example, one species of monkey flower, Mimulus lewisii, has wide pink flowers and a small amount of concentrated nectar, which are ideal for bumblebees. The closely related M. cardenalis flowers are narrow and dark red and have large amounts of dilute nectar, a hummingbird pollination syndrome. The two species grow together but do not hybridize because bumblebees do not visit hummingbird flowers (they cannot even fit into them) and vice versa, but the two species can be crossed artificially and produce healthy hybrids. Apparently just a few floral genes have major effects in Mimulus, so a few mutations resulted in strikingly different flowers, completely different pollinators, and thus biological isolation.
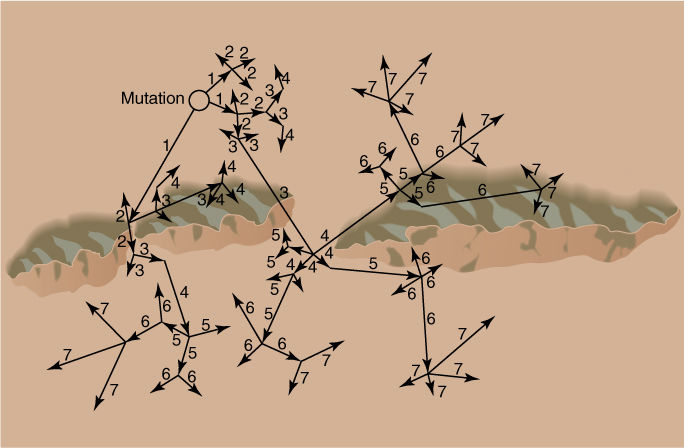
FIGURE 17-14 Gene flow. This range is almost divided in two by a series of mountains over which pollen and seeds cannot travel; this slows down gene flow and homogenization of the population. The numbers indicate the generation in which the allele flows: 1, gene flow in the first generation; 2, gene flow from plants that grew from seeds produced by the gene flow of the first generation; and so on. Here, three generations were necessary for the new allele to become distributed to the lower side of the mountains, seven generations to become widespread throughout the range. In a species of annual plants, each generation is 1 year long; for biennials, a generation is 2 years long, and gene flow is slower. In trees that do not produce their first flowers, pollen, or seeds until they are many years old, gene flow can be very slow.

FIGURE 17-15 (A) These small flowering herbs (Caltha) and shrubs (willow, Salix) grow in high alpine habitats near snow. A mountain range, such as the Tetons in Wyoming (B), provides many suitable patches of habitat near their summits, but each patch is separated from others by valleys that are warm or by peaks that are too dry. Seeds are only rarely carried from one patch to another, so gene flow is restricted: The Caltha in this photo is part of a small population that can interbreed but is relatively isolated from other populations on other peaks of this same mountain range.
Evolutionary changes in pollinators can also act as reproductive barriers for plants. If a plant population covers a large area, some parts of the range probably have characteristics different from those of other parts, such as elevation, temperature, and humidity. These variations may be important to pollinators and seed distributors if not to the plant. If so, the animals may diverge evolutionarily into distinct species, each limited to a small part of the plant’s range. Little or no flow of the plant’s genes occurs because of the restricted movements of pollinators and seed dispersers. These examples of biological reproductive barriers prevent pollen from moving from one plant to another, and thus, neither pollination nor fertilization occurs. Consequently, these are called prezygotic isolation mechanisms: They act even before a zygote can be formed.
The environmental diversity of a large geographic range can lead directly to divergence of the plants themselves. Although a plant species may occupy an extensive, heterogeneous range, mutations will gradually arise that are particularly adaptive for certain aspects of specific regions of the range. When these new alleles arrive at that part of the range by gene flow, their frequency there is increased by natural selection. When they arrive at other parts of the range, their frequency remains low or they are eliminated there if they are neutral or selectively disadvantageous for conditions in these sites (FIGURE 17-16). Even with active gene flow and interbreeding, different subpopulations of the plant species emerge, each adapted to its own particular portion of the total range. As this process continues, each subpopulation becomes progressively more distinct and may be recognized and named as a subspecies. Finally, because of the large number of unique, characteristic alleles, each may become sufficiently distinct phenotypically that the two subpopulations can no longer interbreed: Their genomes are too different. At this point, postzygotic internal isolation barriers are in place, and the two subpopulations must be considered separate species.
One of the earliest postzygotic barriers to arise is hybrid sterility: The two populations occasionally interbreed or are artificially cross-pollinated and produce viable seed, but the seed grows into a sterile plant. Spore mother cells in anthers and ovules in the sterile hybrid are unable to complete meiosis because of a failure of synapsis: “Homologous” chromosomes are no longer truly homologous, and they fail to form pairs and synaptonemal complexes during prophase I. Without fertile pollen or ovules, no seed is formed, and the mixture of the two genomes ends when the hybrid plant dies. The two populations continue to diverge and become even more distinctly separate species. If cross-pollination occurs, alleles from one parent may code for proteins incompatible with those coded by alleles from the other, and not even a sterile hybrid can result; instead, the zygote or embryo dies early in development. This is called hybrid inviability.
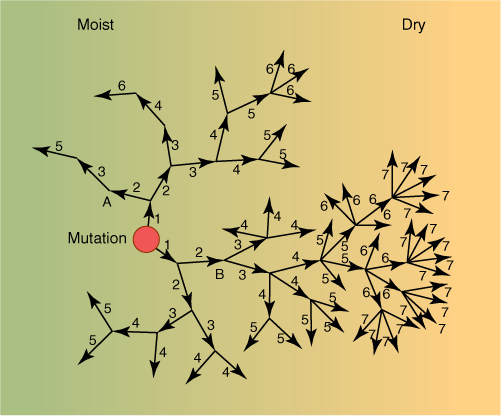
FIGURE 17-16 A mutation has produced an allele highly adaptive in dry parts of the range but neutral or deleterious in moist parts. Because it originated in a less than optimal environment, in generation 1, only two offspring survive. In generation 2, each of these produces two more that survive, one in a moister habitat (A) and one in a drier region (B). The allele is deleterious at A, and only a single progeny survives in generation 3; at B, two survive, both in drier habitats, and in generation 4, each produces three surviving progeny. As the alleles flow, by chance, into regions that are more optimal (drier), reproductive success is greater, and the allele frequency increases. As they flow, by chance, into unsuitable habitats, reproductive success is low and the allele never becomes abundant.
After internal isolation barriers are established, evolutionary divergence should be even more rapid because new alleles cannot be shared; gene flow between the two populations comes to a stop. Each species becomes more easily recognizable as new characters evolve in each, characters which are not present in its sibling species. The two species are initially so similar that they are obviously related; such closely related species are grouped together into genera by taxonomists.
Divergent evolution may result in numerous types of new species; in some cases, one subpopulation changes into a new species, while the remaining part of the population continues relatively unchanged as the original species. In other cases, both subpopulations change so much that two new species emerge and the original species no longer exists. The original species has become extinct, although it has numerous progeny that form the members of two new species.
Adaptive Radiation
Adaptive radiation is a special case of divergent evolution in which a species rapidly diverges into many new species over an extremely short time, just a few million years. This usually occurs when the species enters a new habitat where little or no competition or environmental stress exists. The best examples are the colonization of newly formed oceanic islands such as the Hawaiian or Galapagos Islands. After being formed by volcanic activity, they are initially devoid of all plant and animal life, but eventually a seed arrives, carried by a bird, wind, or ocean currents. After the seed germinates and begins to grow, it is free of danger from herbivores, fungi, bacteria, or competition from other plants. It must be relatively adapted to the soil, rainfall pattern, and heat/cold fluctuations, but otherwise, its life is remarkably free of dangers. If this plant is self-fertile or if it reproduces well vegetatively, it successfully colonizes the area.
All offspring greatly resemble the first, founder individual(s) because the initial gene pool is extremely small; if just one seed is the founder, the original gene pool consists of its two sets of alleles. This homogeneity may last only briefly because, with the lack of competition, pathogens, and predators, fewer forces act as selective agents. Consequently, new alleles build up in the population much more rapidly than can occur in the parental population on the mainland (FIGURE 17-17). Although the population is small, it is more subject to accidents, and thus, the gene pool can change rapidly and erratically; this is genetic drift, and the island population soon becomes heterogeneous. Natural selection for adaptation to soil types, drainage, climate, and metabolism is still operative; as a result, divergence is based largely on physical factors in the environment.

FIGURE 17-17 One of the pioneer plants that arrived and colonized the Hawaiian Islands was an aster, related to dandelions. Because of the low levels of competition and because so few individuals existed initially, each mutation became a significant part of the gene pool even while only one or a few copies existed; therefore, genetic drift was rapid. Argyroxyphium, silverswords, are basically giant, perennial dandelions. Their leaves grow in a rosette, and their many “flowers” are like dandelion “flowers,” actually being groups of tiny flowers. Silverswords grow only in Haleakala National Park in Hawaii. Goats ate them almost to extinction before the park was established.
If the island is very far from any seed source (the mainland or other islands), like Hawaii is, hundreds of years may pass before the arrival of more seeds of either the same or another species. Even if it is relatively close to the mainland, like the Galapagos, seeds arrive only rarely. For thousands of years the only animals might be birds or airborne insects, typically spiders; land reptiles (lizards) and mammals (mice) arrive by rafting, but this can be extremely rare.
Adaptive radiation can also occur in mainland populations if the environment changes suddenly and eliminates the dominant species of a region. With the absence of these species, competition changes and other species that had not been able to compete well before can now occupy the new areas; while few in number, they undergo genetic drift, rapidly producing many unusual genotypes. Within a short time, many new species are recognizable, and adaptive radiation has occurred.
Convergent Evolution
If two distinct, unrelated species occupy the same or similar habitats, natural selection may favor the same phenotypes in each. As a consequence, the two may evolve to the point that they resemble each other strongly and are said to have undergone convergent evolution. The most striking example is the evolutionary convergence of cacti and euphorbias (see Figure 17-11, FIGURES 17-18, and 17-19). Cacti evolved from leafy trees in the Americas; as deserts formed, mutations that prevented leaf formation were advantageous because they reduced transpiration. Other selectively advantageous mutations increased water storage capacity (succulent trunk) and defenses against water-seeking animals (spines). In Africa, the formation of deserts also favored a similar phenotype, and the euphorbia family became adapted by these means. As a result, the succulent euphorbias are remarkably similar to the succulent cacti, even though the ancestral euphorbias are quite distinct from the ancestral cacti. Two groups cannot converge to the point of producing the same species; only the phenotypes converge, not the genotypes. For example, cactus spines are modified leaves, whereas euphorbia spines are modified shoots.
Plants and People
BOX 17-2 Zoos, Botanical Gardens, and Genetic Drift
Genetic drift, adaptive radiation in small populations, and artificial selection are important factors in conserving endangered species. With only a small number of individuals maintained in zoos, botanical gardens, and national parks, the gene pools of the captive plants and animals are so small that rapid fluctuation and drift can cause them to evolve rapidly. An endangered species protected in captivity may be lost because it evolves into one or several species adapted to highly artificial, humanly maintained conditions. Even if it still looks like the ancestral species that was placed into the zoo, it differs in its resistance to infections, parasites, and predators and its ability to understand and perform mating interactions. Most zoologists now believe that habitat preservation is the only true means of maintaining an endangered animal species. Zoos cannot accommodate the numbers of individuals necessary and cannot provide realistic habitats that act as agents of natural rather than artificial selection. Many of the progeny born in zoos would be poorly adapted for survival in the wild; if they are kept alive by antibiotics and special diets, the presence of nonbeneficial alleles is maintained in the population. Natural populations have a tremendous death rate among juvenile animals, a factor necessary for natural selection; with this eliminated in zoos, the gene pool of the captive population rapidly diverges from that of the natural population, being greatly enriched in deleterious mutations. The zoo population may become incapable of surviving without human care. The same is true of plant species maintained in botanical gardens, where often the most unusual varieties are given special attention, increasing the presence of exotic alleles that would be selectively disadvantageous in natural conditions. Artificial and natural selection result in very different gene pools. This is not a condemnation of zoos or botanical gardens: They have provided an essential service in saving many species from extinction, which at present would be the only alternative for them, but it should be remembered that a species in the wild consists not only of its individuals but of its genetic diversity interacting with its environment.
In order to provide truly safe sanctuary for endangered species, each park or habitat preserve may have to be expanded to encompass a population large enough to be stable against genetic drift. Even Yellowstone National Park is not large enough to maintain a genetically stable population of grizzly bears, although the buffalo seem to be doing well. Many individuals of a plant species may survive in a small area, such that 1 km2 may hold a large enough number for their genetic stability on a theoretical basis. An important aspect, however, is to preserve enough habitat to maintain the pollinators and seed dispersers as well. The habitat must also be diverse enough to contain realistic selection pressures, including pathogens, herbivores, and so on. If these are eliminated owing to insufficient habitat preservation, the plant species experiences unnatural selection. Recent forest fires are a good example. Fire is a natural part of many ecosystems and is an agent of natural selection for many features; if fires are suppressed, nonresistant individuals survive, and the allele frequency of the population is altered.
Although artificial selection improves our cultivated plants, it can have disastrous effects on natural populations. For many species, individual specimen plants are valuable commercially, such as orchids, bromeliads, and cacti with particularly beautiful shapes or flowers. In many cases, it is easier and cheaper to collect these plants from natural populations than to cultivate them in nurseries. Extensive plant collecting actually threatens some species with extinction, but more often it has the very serious impact of removing most of the healthy plants from a population. Because the plants that are left are those that are weak, unhealthy, or misshapen, the remaining population has an increased frequency of alleles that produce unhealthy, poorly adapted plants. Even though the collector does not wipe out an entire population, he or she may do significant damage to the population’s genetic resources. We inflict the same condition on parrots, toucans, and tropical fish.
![]() Evolution and the Origin of Life
Evolution and the Origin of Life
The species present today have evolved from those that existed in the past, which evolved from those that existed before them. It is appropriate now to consider how life arose and what it was like initially.

FIGURE 17-18 Cacti and euphorbias have undergone extensive convergent evolution: Many early euphorbias were leafy trees (A) like early cacti (see Figure 17-11A), but some have evolved to be succulent (B) like many cacti. Both (A) Sapium sebiferum and (B) Euphorbia gymnocalycioides are members of the Euphorbia family, Euphorbiaceae.
The most seriously considered hypothesis about the origin of life on Earth is that of chemosynthesis. Basically, the chemosynthetic hypothesis attempts to model the origin of life using only known chemical and physical processes, rejecting all traces of divine intervention. It was first proposed by the Russian scientist A. Oparin in 1924 and then by J. B. S. Haldane in England. They postulated that before the origin of life, the surface of Earth was different from the way it is now, and the chemicals present then could react spontaneously, producing more complex chemicals that could in turn continue to react. Over millions of years, reactions might produce all the molecules necessary for life, and these might aggregate into primitive protocells. From the protocells, natural selection would guide the evolution of true, living cells. Four conditions would have been necessary for the chemosynthetic origin of life: The primitive Earth would have to have had (1) the right inorganic chemicals, (2) appropriate energy sources, (3) a great deal of time, and (4) an absence of oxygen in its destructive molecular form, O2.

FIGURE 17-19 Both cacti and euphorbias are parasitized by distinct types of mistletoe that have also undergone convergent evolution. Flower buds of Viscum minimum are shown here emerging from the body of Euphorbia lactea. Tristerix aphyllus infects a cactus called Trichocereus chilensis (see Box 2-2). Both mistletoes are dwarf plants that spend most of their life buried deep inside the host’s succulent body.
Conditions on Earth Before the Origin of Life
Chemicals Present in the Atmosphere
Earth condensed from gases and dust about 4.6 billion years ago; it was initially hot and rocky and had an atmosphere composed mostly of hydrogen. Because hydrogen is such a light gas, most of this first atmosphere was lost into space. It was replaced by a second atmosphere produced by release of gases from the rock matrix composing Earth and from heavy bombardment by meteorites. Both sources would have provided gases such as hydrogen sulfide (H2S), ammonia (NH3), methane (CH4), and water. All of these are found in volcanic gases and in meteorites that still strike Earth. Molecular oxygen was absent; it had already combined with other elements, resulting in compounds such as water and silicates. The early second atmosphere was a reducing atmosphere due to the lack of molecular oxygen and the presence of powerful reducing agents (FIGURE 17-20).
Energy Sources
There must have been a complex chemistry in the early second atmosphere because it was exposed to powerful sources of energy. Foremost was intense UV and gamma radiation from the sun. These radiations have energetic quanta that knock electrons from atoms, creating highly reactive free radicals. Part of the ammonia would have decomposed to hydrogen and nitrogen, and some of the methane would have converted to carbon monoxide and carbon dioxide, increasing the complexity of the atmosphere.
Heat was another source of energy available to power reactions. One heat source was the coalescence of gas and dust to form Earth: As the particles fell toward the center of gravity, they accelerated and then collided, converting kinetic energy to heat. A second heat source was radioactive decay of heavy elements like uranium and radium; this decay was extremely intense 4.5 billion years ago. Even today, enough radioactive decay remains to keep Earth’s core molten. The chemicals present in the early second atmosphere were also dissolved in the ocean’s water, and whenever they came into contact with hot or molten rock, endergonic reactions could proceed, resulting in the formation of more complex chemical compounds.

FIGURE 17-20 The outer planets, except for Pluto, are so massive that their gravity has retained their original hydrogen atmosphere. In addition, meteor bombardment and other activities have added ammonia, methane, and other components that make these atmospheres similar to Earth’s early second atmosphere. Reactions in the atmosphere may be producing organic compounds.
Electricity was abundant on a gigantic scale; much of Earth’s water was initially suspended in the atmosphere because of the high temperature of the air and the planet’s surface. When sufficient planetary cooling occurred, rains fell, but the rainwater evaporated upon hitting the hot surface. Rainstorms must have been immense, lasting for thousands of years and generating tremendous amounts of lightning. As each lightning strike flashed through the atmosphere, it triggered more chemical reactions, and the resulting products were washed downward by rain: On the hot surface, they would have boiled, producing even more chemical reactions. After further planetary cooling, the surface temperature dropped to less than 100°C, and liquid water began to accumulate as streams, lakes, and oceans.
Volcanoes also produce lightning around their throats as they erupt (FIGURE 17-21). Considering the immense amount of volcanism that occurred, this lightning would have supplied a significant amount of energy. Also, volcanic lightning occurs through the clouds of venting gases where hydrogen sulfide, methane, ammonia, and water are most concentrated.

FIGURE 17-21 Rapid movement of gases during a volcanic eruption generates the electrical potential necessary for lightning. Electrical discharges through the volcanic gases produce organic compounds.
Time Available for the Origin of Life
The time available for the chemosynthetic origin of life basically had no limits, simply because of the lack of free molecular oxygen. Without oxygen, no agent was present to cause the breakdown and decomposition of the chemicals being created. If molecular oxygen had been present, the chemicals either would have not formed or would have oxidized soon after formation; without oxygen, they could accumulate for millions of years. The ocean of that time has been called a “dilute soup” or a “primordial soup” containing water, salts, and numerous organic compounds that became increasingly complex as time went on. As much as 1.1 billion years may have elapsed between the time Earth solidified and life arose.
Chemicals Produced Chemosynthetically
After the writings of Oparin and Haldane, the first experimental tests of the chemosynthetic hypothesis were performed in 1953 by a graduate student, S. Miller, at the University of Chicago. He constructed a container that had boiling water in the bottom and a reducing atmosphere in the top; electrodes discharged sparks into the gases, simulating lightning: As the water boiled, steam rose, mixed with the atmosphere and was acted on by the electrical sparks and then condensed and fell back into the water to be cycled again (FIGURE 17-22). Miller let his first experiment cycle for a week and noticed that the solution had become dark from the accumulation of complex organic compounds that had formed. When he analyzed their composition, he found that many different substances were present, including amino acids. Since then, this type of experiment has been performed numerous times, testing the effects of varying atmospheric compositions, using several types of energy sources, or including metal ions in the water. Virtually all of the small molecules essential for life can be formed this way: amino acids, sugars, lipids, nitrogen bases, and so on.
These experiments tell us what is theoretically possible; direct analysis of meteorites and lunar samples reveals what has actually happened in nonliving environments. Rock samples brought back from the moon by the Apollo astronauts contain various organic compounds, including amino acids. The interiors of meteorites, uncontaminated by the fall through the atmosphere or contact with soils, have contained alcohols, sugars, amino acids, and the nitrogenous bases that occur in nucleic acids. With regard to the formation of monomers, the chemosynthetic hypothesis represents a plausible model.
Formation of Polymers
Monomers present in the early ocean had to polymerize if life were to arise, but polymerization required high concentrations of monomers. Given enough time, the oceans would have changed from a dilute soup to a concentrated one, but that probably was not necessary. Numerous mechanisms would have produced pools of highly concentrated reactants.
An obvious method of concentration is formation of seaside pools at high tide that evaporate after the tide goes out. With intense sunlight the pools would have been warm, perhaps even hot, and polymerization reactions could occur. With the return of high tide, the polymers would be washed into the sea and accumulate. Monomers could also have accumulated when ponds and seaside pools froze; ice is relatively pure water and the monomers become increasingly concentrated in whatever water has not yet frozen. This might produce a class of polymers distinct from those formed by evaporation at high temperature.
Absorption by clay particles could have concentrated monomers, and clays are receiving great attention now. Because clay particles are tiny fragments of rock, they have a regular, crystalline surface; organic molecules adhere to them in a particular orientation, not simply at random. Thus, binding to a clay particle is similar to binding to an enzyme. Furthermore, the crystalline matrix of clay contains contaminating ions of iron, magnesium, calcium, phosphate, and other charged groups that are typically present at the active sites of enzymes. Considerable experimentation is being done to determine whether clays might have both concentrated monomers and acted as the first primitive catalysts.
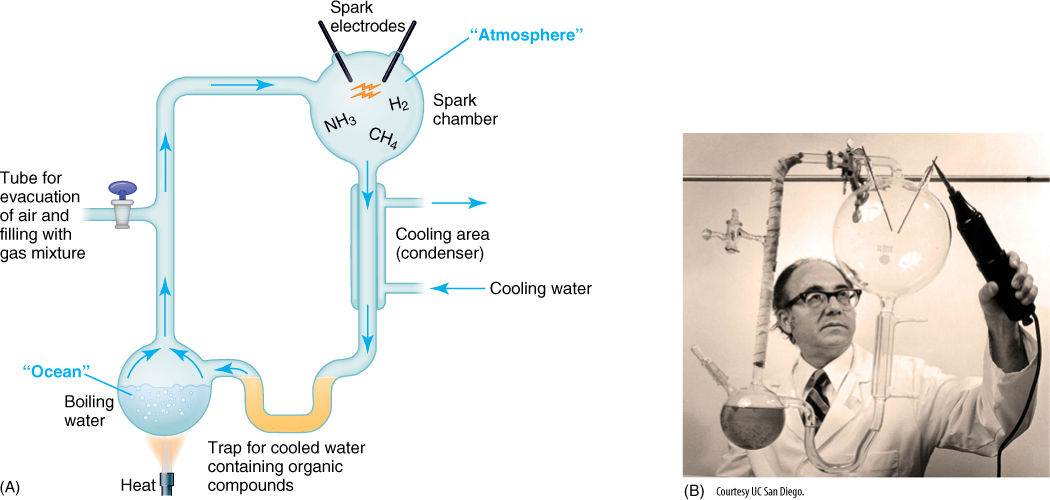
FIGURE 17-22 (A) Diagram and (B) photograph of apparatus used by Miller to show that the first steps in the chemosynthetic origin of life were possible.
Aggregation and Organization
The next step in the possible chemical evolution of life would have been aggregation of chemical components into masses that had some organization and metabolism. Fatty, hydrophobic material would have accumulated automatically as oil slicks in quiet water or as droplets in agitated water. Fatty acids would have occupied the outermost layer, accompanied by other molecules such as proteins that had a hydrophobic portion and a hydrophilic one. The interior may have been mostly hydrophobic, but proteins that had a hydrophobic exterior and hydrophilic interior would have added complexity. Aggregation of certain types of proteins would have resulted in large regions of hydrophilic sites.
These first aggregates would have formed basically at random, controlled only by relative solubility. If some of the proteins had some enzymatic activity by chance, the aggregate would have had some simple “metabolism”—perhaps the conversion of some molecule, absorbed from the sea, into another molecule; the aggregates would have been heterotrophs completely.
These aggregates are not postulated to have been alive or even to have been early stages of life because at that point no means of storing genetic information existed. Without genetics, natural selection cannot occur. Countless numbers of these aggregates might have formed and disassociated throughout all of the oceans over millions of years. Their existence may have had a significant effect on the chemistry of the oceans. Some may have provided appropriate conditions for certain proteins to be very active enzymes that might have produced quite complex products. However, others may have been able to degrade such products, and an equilibrium may have been established between formation and destruction of ever more elaborate molecules and polymers.
At some point, presumably an aggregate formed that did have a heritable information molecule able to direct the synthesis of products useful to the aggregate. By “useful” we mean that the product helped the aggregate persist longer, without being broken up by wave action or dissipating by diffusion. It may have helped the aggregate grow or even helped the information molecule replicate. With the presence of heredity, everything changed; any mutation that caused the production of a more efficient, more advantageous enzyme or structural protein provided a strong selective advantage and could be passed on to progeny aggregates. Currently, attention is focusing on RNA as the first heritable information molecule. As the aggregate increased in size, perhaps largely by absorbing material from the ocean, its information molecule would replicate. If the aggregate divided into two by either wave action or surface tension and if each half contained an information molecule, reproduction occurred.
Early Metabolism
The aggregates would have been complete heterotrophs, absorbing all material from the ocean and modifying only a few molecules. As aggregates continued to consume certain nutrients, however, scarcity occurred; any aggregate that could produce an enzyme capable of synthesizing the scarce molecule from an abundant one still available in the ocean would have had a strong selective advantage. With this, there would have been a metabolic pathway two steps long involving two enzymes (FIGURE 17-23). Being able to synthesize a valuable, scarce molecule from an abundant, free one would have given that aggregate great advantage over the others; it would have had a more rapid metabolism and would have grown and reproduced more rapidly.
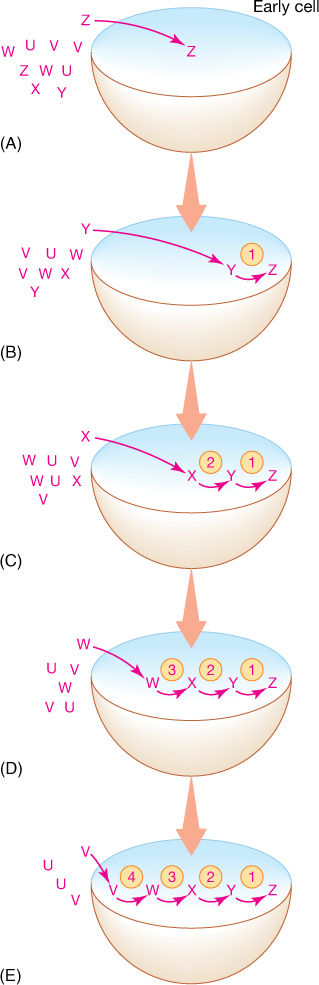
FIGURE 17-23 (A)—(E) We believe that many metabolic pathways evolved backward as raw materials in the environment became scarce. Currently, the pathway for photosynthesis has gone as far as it can, requiring only water and carbon dioxide.
As its abundance increased, it and its descendants would finally have become so numerous as to use the second chemical faster than it could be formed chemosynthetically, and scarcity would have occurred again. Natural selection would again favor any genetic system that could extend the metabolic pathway another step to include an abundant precursor. Recall that in photosynthesis in modern plants, the only two precursor molecules that need to be drawn from the environment are water and carbon dioxide—both are abundant and cheap. From them, metabolic pathways extend and ramify such that all carbon-containing compounds are derived from them. No preformed organic molecules are necessary.
Energy metabolism must have been important also, and glycolysis must have evolved early because it is present in virtually all organisms. The aggregates and first cells may have absorbed some ATP and generated more by fermentation with glycolysis for millions of years. At some point, electron transport systems and hydrogen ion pumping systems would have evolved. The first would have been powered by light, and an early form of photosynthesis would have evolved. Higher plant photosynthesis is elaborate, but much simpler types occur even today in certain photosynthetic bacteria. The evolution of photosynthesis would not have been as complex or as difficult as chloroplasts would lead us to believe. Oxidative electron transport, as occurs in mitochondria, could not have evolved because free molecular oxygen still did not exist.
Oxygen
The evolution of chlorophyll a and photosynthesis that liberates oxygen had two profound consequences: (1) it allowed the world to rust, and (2) it created conditions that selected for the evolution of aerobic respiration. Until that time, photosynthesis had involved the bacterial pigment bacteriochlorophyll, which liberates sulfur from hydrogen sulfide rather than oxygen from water. Earth retained its reducing atmosphere, and chemosynthesis of complex precursor molecules could continue. However, after chlorophyll a evolved, the raw material for photosynthesis became water, and free molecular oxygen, O2, was released as a waste product. We know this evolutionary step occurred 2.8 billion years ago because the oxygen rapidly combined with iron and formed ferric oxide—rust. Sedimentary rocks of this age contain a thick red layer of rust, indicating when oxygen-liberating photosynthesis arrived and how long the oxygen/iron reactions continued. Only after all of the free iron in Earth’s oceans had oxidized did oxygen finally begin to accumulate in the atmosphere. The atmosphere present today was derived from the early second atmosphere by this addition of oxygen from photosynthesis. It is an oxidizing atmosphere.
The period of rusting was critically important for all life because it kept the concentration of free oxygen very low. It can be difficult for us humans—obligate aerobic animals—to appreciate just how toxic free oxygen is; we need it to live, but our bodies have numerous mechanisms to keep it under control and to prevent it from reacting at random in our bodies. The same is true of plants, fungi, algae, and many bacteria. Had iron not been present, the concentration of free oxygen in water and air might have increased rapidly and killed everything; possibly there would have been no survivors. With iron, however, oxygen concentration remained low for millions of years; it was an environmental danger but was not instantly, universally lethal. Any mutation that produced a mechanism that could detoxify oxygen had great selective advantage. Of course, the best way is to add two electrons to it and let it pick up two protons and turn into water, which is exactly what cytochrome oxidase does. This is the last electron carrier in the mitochondrion transport chain, and with its evolution, aerobic respiration became possible. Oxygen was transformed from a dangerous pollutant to a valuable resource. The evolution of aerobic respiration did not occur in mitochondria but rather in bacteria, and some species seem to retain very relictual, early versions.
The buildup of atmospheric oxygen had other important effects; under the influence of UV light, oxygen is transformed into ozone, O3. Ozone is extremely opaque to UV light and thus prevents most of it from penetrating deep into the atmosphere and reaching Earth’s surface. This immediately removed UV light as an energy source and must have greatly slowed the chemosynthetic formation of organic molecules. Biological photosynthesis became the main means of bringing energy into the living world. The lack of UV radiation made the surface a safer place to live; UV radiation damages nucleic acids, and while the atmosphere lacked ozone, probably nothing lived near the top of the oceans or on land. With the ozone shield, organisms could move higher in the oceans and onto the mud of seashores. The transition to terrestrial life became possible.
The Presence of Life
The chemosynthetic theory postulates a long series of slow, gradual transitions from completely inorganic compounds to living bacteria. At which stage can we say that life came into being? Can the aggregates be considered alive? This is a difficult question because the theory delineates no absolute demarcation between living and nonliving objects (FIGURE 17-24). The chemistry of living creatures is more complex than that of nonliving objects, but it does not possess any unique properties. The physics of living and nonliving systems is identical. As so often in the living world, we are dealing with a continuum, not simply two mutually exclusive alternatives. To look for a dividing line would be simplistic; it is more important for us to understand the processes in their complexity.
FIGURE 17-24 The fossil remains of an early prokaryote that lived approximately 3.5 billion years ago.
 At the Next Level
At the Next Level
1. Isolation barriers. This chapter introduced the concept of abiological and biological reproductive barriers. The concept is simple: Anything that prevents two members of the same species from interbreeding is a reproductive barrier (an isolation barrier). But think about this for a minute: Numerous phenomena can prevent two individuals from reproducing, and these phenomena might differ for long-lived plants versus annuals; for ones that live in sparse populations versus dense populations; for animal-pollinated plants versus wind-pollinated plants, and so on. There is a large literature about isolation barriers, so be careful to focus on those that pertain to plants.
2. Refugia. From time to time, conditions change such that most members of a species are killed, or at least they fail to reproduce. This might be climate change, drought, or the outbreak of a new fungal pathogen or an insect pest. During some of these changes, some small groups of individuals might be located in several safe areas, areas called refugia (sing.: refugium), where they are protected from the altered conditions. The group of individuals may be isolated in the refugia for just a few years until the fungus disease is over, but they may be isolated as long as 100,000 years during an ice age. Either way, once the harmful conditions are over and this species can spread and survive away from the refugia, an important concept is that most of the genetic diversity of the species was lost, all except that of the members of the refugia. Consequently, as a species spreads out of refugia, it will be very different genetically than it was before the harmful conditions began. In some cases this can lead to rapid evolution, in other cases it results in extinction.
3. Molecular clocks and the rates of evolution. It is natural to wonder when particular organisms evolved, or when particular structures or physiologies came about. In some cases, we are lucky enough to find fossils that give us some clue, but in many cases, there are no appropriate fossils and we have to estimate from other evidence. Many people have assumed that certain sequences of DNA—such as introns and the spacers between genes—have no function. If that is true, mutations in these regions are neither beneficial nor harmful, and these mutations do not cause a group of organisms to evolve more rapidly or slowly: The mutations merely come into existence at some neutral rate. Thus, older sequences would have more mutations than younger sequences, and by comparing related non-coding sequences in different species we would know which species are younger and which are older. This is called a molecular clock sequence, and although some people have great faith in molecular clock sequences, other people see many serious problems.
SUMMARY
1. When considering the reproductive effort of an entire population, it is important to consider all possible alleles for each gene, their relative frequency in the gene pool, and their effects on reproductive success.
2. Sexual reproduction alone does not cause relative abundance of alleles in the gene pool to change. Mutations bring about new alleles but do not greatly affect allele frequency.
3. Both artificial and natural selection increase the fitness of a population to its habitat by preventing certain individuals from breeding and passing their alleles on to the next generation. Artificial selection often increases the plant’s fitness for artificial habitats.
4. In order for natural selection to operate, a population must produce more offspring than can possibly survive in the environment, and offspring must be genetically diverse. These two conditions occur almost everywhere in nature at all times.
5. Natural selection operates only on the phenotypes of preexisting alleles.
6. Plants and most other organisms never do anything with purpose or intent. Only humans and higher primates have purpose, intent, and will and then only with regard to voluntary actions.
7. Organisms face multiple selection pressures and must be adapted to all aspects of their environment. Certain aspects are more important than others, and it may be less dangerous to be poorly adapted to one factor than to another.
8. Most adaptations have negative consequences as well as advantageous ones; retention or loss of these alleles from the gene pool is related to the relative benefit versus cost of the adaptation. As an environment changes, the selective value of the phenotype may also change.
9. In phyletic speciation, all of a species evolves into one new species. In divergent speciation, part of a species evolves into a new species while the rest of the population continues as the original species or evolves into a different new species.
10. Gene flow is inhibited or completely prevented by reproductive barriers, either abiological aspects of the environment or biological aspects of the organisms themselves or of their pollinators and seed dispersers.
11. Life is believed to have originated on Earth by the process of chemosynthesis. This hypothesis postulates that reactions of inorganic compounds in Earth’s early second atmosphere resulted in formation of organic compounds that coalesced into simple aggregates with rudimentary metabolism. After a system of heredity developed, evolution by natural selection made it possible for truly living cells to come into existence.
IMPORTANT TERMS
abiological reproductive barrier
adaptive radiation
allopatric (geographic) speciation
artificial selection
biological reproductive barrier
chemosynthesis
convergent evolution
divergent speciation
founder
gene flow
gene pool
genetic drift
hybrid inviability
hybrid sterility
natural selection
phyletic speciation
population genetics
reducing atmosphere
speciation
sympatric speciation
REVIEW QUESTIONS
1. Evolution is the gradual conversion of one species into one or several new species. It occurs for the most part by __________________ ________________. Mutations cause new __________________ or new __________________ to arise, which affect the ___________________ of the individual.
2. Relative to a human lifetime, is evolution a process too slow or too fast for us to see it easily?
3. As the idea of geological evolution developed, it had an impact on biological thinking. Describe the impact. What was the importance of river deltas?
4. The discovery of evolution by natural selection happened at about the same time as other critical discoveries. Name some of these discoveries. Can you describe how they might have affected biological thinking?
5. Genes and inheritance have an impact on the lives of parents and their progeny, the F1 and F2 generations. Population genetics also deals with genetics, but in a different way. What is the definition of population genetics? How can the abundance of an allele be different in a population as compared with an individual of the population? (Hint: How can the phenotype of a population differ from that of an individual? Can a population have red flowers, pink flowers, and white flowers? Can one individual have all three types of flowers?)
6. What is the gene pool of a species? Why are some alleles more common than others?
7. Many mutations can be corrected by DNA proofreading/repair enzymes. What would happen to the further evolution of a species if a perfect repair mechanism evolved such that no mutation ever went uncorrected?
8. How does an accident differ from natural selection? Would an unusually severe drought with no rainfall at all for 50 years be a selective or a nonselective destructive force?
9. How does artificial selection differ from natural selection? Do you think the ancestors of lettuce had such soft, nonbitter leaves? Could today’s lettuce plants survive in natural conditions?
10. One of the conditions necessary before natural selection can occur is that the population must produce more offspring than can possibly grow and survive to maturity in that habitat. Do you think this is true for most plants or most animals? (Do not think about humans. We are an exceptional case.) Think about some tree that you see on campus or near your home. Estimate the number of seeds it makes every year and the number of years it might live. How many seeds (offspring) will it produce in its lifetime? Do you think it would be possible for them all to germinate, grow, and reproduce?
11. Another condition necessary for natural selection to occur is that progeny must differ from each other in their types of alleles. Do you think this is true for most plants or animals? Be careful for some familiar crop plants. Bananas have no seeds. They must be propagated by taking cuttings from buds at the base of the plant. All of the bananas in a store are clones—they are all the same plant. Can natural selection affect these bananas? Most apple varieties are also clones. Are they being affected by natural selection?
12. Many people assume that natural selection depends on something outside the plant in the environment affecting the plant, but is that true? Wood is always inside the plant. It is never in contact with the environment. Can the phenotype of wood affect a plant’s fitness? Its survival (Hint: Figure 17-9)? If a mutation caused a plant to not be able to make either vessels or tracheids, just fibers, in its wood, do you think that would affect the tree’s fitness compared with all of the other trees in the population?
13. Name several factors that are not part of natural selection.
14. Change each of the following sentences into ones that are true: (a) Plants produce root hairs in order to absorb more water. (b) Plants close their stomata at night to conserve water. (c) DNA produces mRNA in order to keep ribosomes and potentially harmful high-energy enzymes away from itself. (d) Stems and roots produce bark to keep pathogens out. (e) Cacti and euphorbias have succulent bodies in order to store water. (f) Certain plants have C4 metabolism in order to reduce photorespiration. (g) Some plants time their flowering by night length in order to avoid damage caused by blooming too early in an unusually mild spring.
15. Are plants subjected to just one type of selection at a time or are there multiple selection pressures affecting a plant? The text gives an example of a mutation that improves cold hardiness but that might be eliminated from the gene pool despite being beneficial. Why might it be eliminated?
16. Why is it that evolutionary changes that result in the loss of a structure or metabolism can come about quickly?
17. Modern cacti evolved from ancestors that were leafy trees (Figure 17-11), and in the early stages of cactus evolution, leaves were lost; however, the capacity to produce leaves was not lost quickly. Why not?
18. Look at the two cacti in Figure 17-11. Both the leafy Pereskia in part a and the very desert-adapted Gymnocalycium in part b evolved from the same ancestors—leafy trees that were not themselves cacti. In which genus, Pereskia or Gymnocalycium, has natural selection favored more of the new alleles produced by mutations? In which genus has natural selection retained more of the original alleles and eliminated almost all of the new alleles produced by mutation?
19. What is phyletic speciation? What is divergent speciation?
20. What is gene flow? How does it occur?
21. The Hawaiian Islands were formed as underwater volcanoes that became so tall that they protruded above the ocean’s surface. Being formed from molten rock, they were absolutely sterile at first, but now they are home to many types of plants and animals. How did the ancestral plants and animals get to Hawaii? Was this a type of gene flow?
22. What is an abiological reproductive barrier? Name several. Is something that is an abiological reproductive barrier for a plant necessarily also one for an animal? Would you guess that small herbs have more abiological reproductive barriers than do birds?
23. Differences in flower color, shape, or fragrance are listed as being potential biological reproductive barriers. Do you think they would matter at all in a wind pollinated species? If a plant species has several flower colors (for example, red, white, and pink), could a mutation in the plant’s pollinator—but not in the plant species itself—be a biological reproductive barrier? For example, a mutation that makes it unable to see red?
24. One of the earliest postzygotic barriers to arise is hybrid sterility. Two plants can be crossed, but the resulting seeds grow into sterile plants that cannot reproduce. Is this as important in plants as it is in animals (Hint: How many types of hybrid flowers and fruits are there, compared to hybrid animals)?
25. What is genetic drift? What is a founder? Are these important in large populations or in small ones? How are these phenomena important when we create national parks and preserves to protect endangered species?
26. What is convergent evolution? Euphorbias and cacti are often used as examples of convergent evolution. Have their genotypes converged? Have their phenotypes converged? Both have spines, but if we look carefully we can tell that those of cacti evolved from ____________________ and those of euphorbias evolved from ____________________. They have converged, but not so much that we cannot tell them apart.
27. What is the theory of chemosynthesis?
28. Describe the conditions present on Earth at the time of its early second atmosphere. What chemicals and energy sources were present? What chemical in particular was absent? Can you speculate about what conditions would be like on Earth today if meiosis, crossing-over, and sexual reproduction had never evolved?
29. Life originated on Earth so long ago that erosion and weathering and metamorphosis of rock have destroyed most traces of the processes that occurred, but oxygen is an interesting exception. What is the source of all the oxygen in our atmosphere? What happened to the first oxygen to be produced? Why was this important to the survival of all other life forms?
Design Credits: Hummingbird: © Tongho58/Moment/Getty; Green plant cells: © ShutterStock, Inc./Nataliya Hora; Purple tulip: © ShutterStock, Inc./Marie C Fields; Dandelion: © ShutterStock, Inc./danielkreissl; Poppy: © ShutterStock, Inc./Saruri; Plant icon: © ShutterStock, Inc./Vector; Digging man icon: © ShutterStock, Inc./Z-art