Botany: An Introduction to Plant Biology - Mauseth, James D. 2017
Genetics and Evolution
Classification and Systematics

Chapter Opener Image: You probably have no trouble recognizing these as palm trees even though it is unlikely you have ever seen these plants before. They are Washingtonia filifera, native to seeps and artesian pools in the deserts of California. They are recognizable as palms because they have so many features we associate with palms we are already familiar with: a solitary unbranched trunk topped by many large leaves. Most palms have these features because they arose early in the evolution of the palm family and have been inherited for generation after generation ever since. But habitats are variable and plants diversify, so some palms are now shorter, others grow in clumps, some have smaller leaves. Features such as having one unbranched stem have evolved in other families; for example, many people assume that cycads are short, fat palms. When we attempt to classify organisms, we try to group together those that have similar features because they are closely related, and we try not to be fooled by characters that have evolved two or three times in different groups.
OUTLINE
✵ Concepts
✵ Levels of Taxonomic Categories
✵ Cladistics
- Understanding Cladograms
- Cladograms and Taxonomic Categories
✵ Other Types of Classification Systems
- Artificial Systems of Classification
- Classification Systems for Fossils
✵ Taxonomic Studies
✵ The Major Lines of Evolution
Box 18-1 Plants and People: Development of Concepts of Evolution and Classification
Box 18-2 Botany and Beyond: Cacti as Examples of Evolutionary Diversification
Box 18-3 Plants and People: Geneology Versus Clades: Your Family History Is the Opposite of a Clade
Box 18-4 Plants and People: Identifying Unknown Plants
LEARNING OBJECTIVES
After reading this chapter, students will be able to:
✵ Order the levels (taxa) of the natural classification systems.
✵ Explain synapomorphies and homoplasies.
✵ Summarize the way a cladogram shows evolutionary patterns.
✵ Define paraphyletic groups and polyphyletic groups.
✵ Recall two uses of an artificial classification system.
✵ Summarize the process for declaring a new plant species.
✵ Restate the importance of isotypes.
✵ Name the three domains of all organisms.
✵ List the major evolutionary lines that diverged within kingdom Plantae.
 Did You Know?
Did You Know?
✵ All plants evolved from one ancestral set of plants, so all are related to each other and have one common ancestor.
✵ Until the 1800s, anyone could make up their own name for a plant or animal; names were very confusing.
✵ In the mid 1800s, various scientific societies decided they would accept only one set of names, those based on the work of Carolus Linnaeus.
✵ Ancient manuscripts in the cuneiform of Sumerians and hieroglyphics of Egyptians name numerous plants, but without pictures, we do not know which plants these refer to.
![]() Concepts
Concepts
Starting approximately 400 million years ago, a long series of mutations and natural selection gave rise to the plants that are alive today. During the intervening eons since then, that evolutionary line has progressed and diversified, branching into more and more lines of evolution. Thousands of these have become extinct and are known only by fossils; thousands of other evolutionary lines are represented by the approximately 280,000 species of living plants. The hereditary relationships of any group of organisms constitute its phylogeny, basically the evolutionary history of each member of the group. Some phylogenetic studies attempt to create models of the evolutionary relationships of very large groups, for example, the steps that were involved as plants, animals, and fungi evolved from some early ancestor. Other studies focus on the relationships of smaller groups, such as the species of the legume family (e.g., peas, beans, and lentils). The goal of modern plant systematics is to understand each of these evolutionary lines and to have a system of names—nomenclature—that reflects their relationships accurately.
In the past, plants were given names merely so that people could communicate about medicinal plants, crops, poisonous plants, and so on. The task of taxonomists was to discover and identify new species and give them unique names, but with the discovery of evolution by natural selection, the basis of naming plants suddenly changed: Natural selection showed that all organisms are related to each other genetically, all are part of a large phylogeny. Taxonomists (also called systematists), scientists who specialize in classification and naming, immediately realized that the most scientifically valid system of assigning names to species would be one that reflected evolutionary relationships. At the end of the 19th century, taxonomists adopted the goals of (1) developing a natural system of classification, a system in which closely related organisms are classified together, and (2) assigning plant names on the basis of phylogenetic relationships. The nomenclature would reflect the natural system of classification. Because organisms range from closely to distantly related, taxonomists devised a classification and a nomenclature with numerous levels. In this system, closely related species are placed into a genus, and closely related genera are grouped together into a family, and so on.
Today, as in the past, there are many regions of Earth that have not been adequately explored. These probably contain thousands of plant species that have never been collected, named, or studied. Plant systematists still have the goals of searching for new species and using them to construct more accurate models of plant evolution. It is important to remember that the pattern of evolutionary diversification—the phylogeny—is a reality. Our classification systems are hypotheses, models that attempt to map that evolution. Because our knowledge is incomplete and imperfect, the current classification systems are only approximations. Most are probably close approximations, and we do not expect major changes; however, we do expect numerous smaller changes to be made periodically for many years.
![]() Levels of Taxonomic Categories
Levels of Taxonomic Categories
Plants have varying degrees of relatedness, and a natural classification system reflects this in its numerous levels.
The most fundamental level of classification is the species, which ideally and theoretically is a set of individuals closely related by descent from a common ancestor. (The word “species” is both singular and plural; the word “specie” refers to money.) Members of a species can interbreed with each other successfully but cannot interbreed with individuals of any other species (TABLE 18-1). Some species are not so predictable; they may not interbreed well with closely related species, but an occasional cross-pollination results in a viable seed that grows into a fertile adult. If this occurs frequently, the two plant groups may best be considered subspecies of a single species. The two subspecies must be so similar genetically that the chromosomes of the two parents can function in the same nucleus, and when the plant is mature, spore mother cells can undergo meiosis successfully. For all of this to happen, the organisms must be extremely closely related, having undergone divergent evolution from a common ancestor only recently so that few mutations have accumulated since they diverged. But as mutations continue to accumulate in each subspecies, finally either no viable seed results when the two are crossed or the seed grows into a sterile plant. The two then must be considered separate species.
Closely related species are grouped together into genera (singular, genus; pronunciation: JEAN us and GEN er uh; the first syllable is pronounced differently). Deciding whether several species are closely related enough to be placed together in the same genus is difficult. No objective criteria exist; the decision is entirely subjective and is often the cause of great dispute. Some taxonomists, generally referred to as “lumpers,” believe that even relatively distantly related species should be grouped together in large genera. Other taxonomists, called “splitters,” prefer to have many small genera, each containing only a few species that are extremely closely related. For example, some taxonomists believe that cranberries and blueberries are so similar that they should go into the same genus, Vaccinium; others think that cranberries are distinct enough that segregating them into their own genus, Oxycoccus, more accurately reflects evolutionary reality. Both groups of taxonomists agree that the two sets of species are closely related, but they have different opinions as to how much evolution has occurred since the time of the most recent common ancestor. The critical concern is that the genera are natural, that all of the species included in the genus are related to each other by a common ancestor, and that all descendants of that common ancestor are in the same genus. Such a group is monophyletic. In an unnatural, polyphyletic group, members have evolved from different ancestors and may resemble each other only as a result of convergent evolution. Whenever systematists discover that they have accidentally classified unrelated plants together in an unnatural, polyphyletic group, they immediately search for a more accurate classification composed only of monophyletic groups.
TABLE 18-1 The Taxonomic Categories
Plants and People
BOX 18-1 Development of Concepts of Evolution and Classification
Our modern classifications have their origins in ancient Greece. Theophrastus, a student of both Plato and Aristotle, wrote extensively about plants around 300 BCE and established many important concepts. He distinguished flowering and nonflowering plants, recognized sexuality in plants, and understood that fruits develop from carpels. Theophrastus described almost 500 species, and our genus names Asparagus, Narcissus, and Daucus (carrot) can be traced directly back to him. Pliny the Elder (Gaius Plinius Secundus; CE 23-79), a Roman lawyer and natural historian, wrote voluminously on almost every subject. His largest work, Natural History, was an attempt to describe everything in the world. Despite its inclusion of many fanciful creatures based only on folk tales, it served as the definitive, authoritative source of information on most subjects for more than 1,000 years. The most important book on plant classification from the ancient world is Materia Medica by Dioscorides. Written in the first century CE, it describes 600 plant species and how they can be used to treat disease. It was the best, often the only, source of information about preparing herbal medicines. Materia Medica was at last superseded by more accurate work during the Renaissance, but it is still published today for its immense historical value as a direct link to early Greek science.
Between the time of Dioscorides and the Renaissance, almost no works of great value in natural history were written. Changes began to occur in the 15th century, and Europe entered an age of exploration. Prince Henry of Portugal sent ships on expeditions down the west coast of Africa, and the explorers returned with new plants and animals and knowledge of new lands and peoples. Simultaneously, exploration of physics, chemistry, astronomy, and geology began. An important result of these explorations was the discovery that Pliny and other ancient authorities had been wrong in many subjects. Until then, people thought the ancient Greeks and Romans had represented a Golden Age and were basically infallible. The realization that the ancients had made errors meant that the answers to questions about the world had to be sought in the world itself, not in ancient books. Exploration—not only geographical but also scientific, philosophical, and religious exploration—became an obsession.
In botany, this new, independent thinking first became apparent in the publication of medically oriented plant books, called herbals. By the middle of the 16th century, they began to contain careful, precise descriptions of plants based on first-hand observation of plants, not by paraphrasing Materia Medica. Most herbals contained fewer than 1,000 plants, so a reasonably good botanist could become familiar with all of them; however, as exploration continued, especially after the discovery of the Americas, the number of plant species became too large for this type of familiar treatment. It became necessary to develop a classification system so that a person would have some means of identifying an unfamiliar specimen and finding it in the ever-larger herbals.
Several important ideas developed at this time. One was the concept of a genus as a group of similar species, established by Gaspard Bauhin (1560—1624). Although this may seem like an obvious notion, it was a profound breakthrough at the time. The concept of species had been easy—God was believed to have created all of the types of organisms—the species. But this concept did not explain why there should be groupings of species, as the concept of genus implies. Why would God create several types of roses, several types of mints? This was not a trivial question then. Until the theory of evolution by natural selection, the world was viewed as a reflection of the mind of God, who presumably had created it. If there were thousands of species of grasses but only three of cattails, this would reveal something about how God thinks. It was obvious to anyone familiar with the increasing number of plant and animal species that some species resembled each other very closely. But why?
Our system of nomenclature, of scientific names, can be traced directly to Carolus Linnaeus, a professor of natural history at the University of Uppsala in Sweden during the middle and latter part of the 18th century. He adopted the genus system of Bauhin, created a large numbers of genera, and placed every species into one genus or another. Every species had both a genus name and a species epithet, the basis of our present binomial system of nomenclature. Before Linnaeus, rather than names, plants had paragraph-long descriptions of their features, which was obviously cumbersome; after Linnaeus, each plant had a name of just two words.
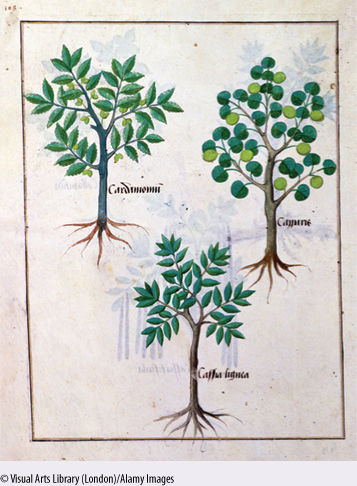
FIGURE B18-1A Prior to the Renaissance, most European botanists considered ancient books to be a better source of knowledge than plants themselves, so they merely copied drawings from preexisting books, and each drawing became less accurate and less useful. Left: cardamon; right: Capparis; bottom: cassia lignea.
In 1753, Linnaeus published Species Plantarum, a treatment of all plant species known in the world at that time. This book made botanical studies of all types much easier because specimens could be identified quickly; however, the system often placed unrelated species into the same genus on the basis of homoplasic (analogous) characters.
The idea of the evolution of species developed slowly over many years; it did not begin with Darwin and Wallace by any means. Bauhin recognized resemblance early, and an expansion of the idea of similarity occurred at the Jardin des Plantes in Paris. Antoine-Laurent de Jussieu wanted to organize the garden in a logical way so that plants resembling each other were planted together (the garden still exists, you can visit it). The accompanying catalog was Genera Plantarum (1789): It was the first major attempt at a natural system of classification, and people could not help but wonder whether some species within each genus had not evolved from others. Perhaps similar genera had changed, evolved. But how? Science was still dominated by the idea of divinely created types.
Unfortunately, J. B. P. de Lamarck at this time presented his theory of evolution by inheritance of acquired characteristics. This was the incorrect idea that all cells of the body produced fluids that diffused to the genitalia, where the fluids were concentrated and formed into sperm cells or egg cells. As an individual changed, the secreted fluids and therefore the characteristics carried by the gametes would change. The evolution of giraffes was explained as follows: Ancestors to giraffes had been born with short necks; however, as they stretched their necks to reach leaves in high trees, their necks lengthened—long necks in the adults would be the acquired characteristic—and subsequently, the genetic fluid passed to their gametes carried information about long necks. Consequently, their offspring had longer necks. Mendel showed that this could not be right because genes are nonvarying particles, not variable fluids. Evolution by natural selection explains giraffes by postulating that variability of neck size occurred among the earliest ancestors of giraffes; alleles in some animals produced short necks and in others slightly longer necks. Those with longer necks could reach more leaves; therefore, they had better survival and reproductive success. Over millions of years, various mutations produced new alleles that resulted in even longer necks, and these new alleles were increased by natural selection. The theory of inheritance of acquired characteristics was never widely accepted, and it gave evolution a bad reputation for many years.

FIGURE B18-1B During and after the Renaissance, people realized they could make accurate drawings and studies of plants only if they examined the plants themselves. Much more realistic images were produced. These are drawings of tobacco plants.
In 1859, Charles Darwin and Alfred Wallace each propounded the theory of evolution by natural selection, which gave natural systems of classification an immediate validity. Taxonomists quickly understood why some species within a genus resemble each other so strongly: They are descendants of a common ancestor and are related by evolution. Similarly, closely related genera constitute a family.
The theory of evolution by natural selection became accepted while two German botanists, A. Engler and K. Prantl, were working on a monumental classification of all the world’s plants, Die Natürlichen Pflanzenfamilien, published in 1915. It organized species on a phylogenetic, natural basis, as understood by Engler and Prantl. In those early days of evolutionary studies, organisms were assumed to have evolved from the simple to the complex (amoebae to humans). Thus, in flowering plants, the wind-pollinated species that have no sepals, and petals were placed first; the sunflowers, with their complex floral structures, were considered more advanced. It was quickly discovered that simplicity may also be the result of evolutionary simplification. Because no automatic correlation exists between simple and primitive or between complex and advanced, the Engler and Prantl classification has many errors, although as a compendium of information it is still an unmatched vital resource. Other major classifications were produced by C. E. Bessey (early 1900s) of the University of Nebraska, A. Takhtajan in Russia, and A. Cronquist (1980s) at the New York Botanical Garden.
The level above genus is family, each family being composed of one, several, or often many genera. Most families are well defined, with widespread agreement as to which species and genera belong in a particular family. As examples, consider how easy it is to recognize the following families: cacti, orchids, daisies, palms, and grasses. The reason for this probably is the age of most families. A line of evolution must be very old—usually at least several tens of millions of years—to diversify into several genera and many species. Even closely related families have been separate evolutionary lines for at least 20 or 30 million and often 50 million years. The common ancestor and many of the earliest species probably have become extinct. Therefore, although two families have many characters in common, enough features are unique to each family to make them recognizable as distinct from each other.
The levels above family are order, class, division, and kingdom. Although you might expect universal agreement at the level of kingdom, even the boundaries of the plant kingdom, kingdom Plantae, are disputed. Some believe that green algae should be included because they are almost identical biochemically to vascular plants. Others believe that, despite the biochemical similarity, green algae should be excluded because they are so different morphologically and anatomically.
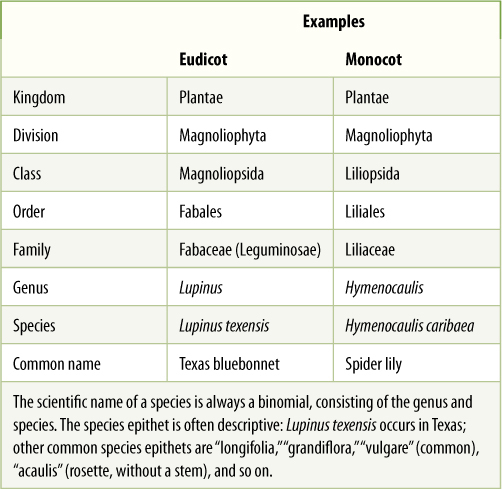
Except for kingdom, genus, and species, the names must have a certain ending to indicate the classification level. Division names end in -phyta, for example Chlorophyta (green algae), Coniferophyta (conifers), and Magnoliophyta (flowering plants) (Table 18-1). Class names end in -opsida, order names in -ales, and family names in -aceae (pronounced as if you were spelling the word “ace”). Genera and species names do not have standard endings. The scientific name of a species is its genus and species designations used together and either underlined or italicized; for example, tomato is Lycopersicon esculentum. Note that the species name is not “esculentum” but is Lycopersicon esculentum. Esculentum is the species epithet, the word that distinguishes this species only from the other species of the genus Lycopersicon. We cannot refer to tomatoes as “esculentum” because that species epithet is used for many different species in different genera: for example, buckwheat, Fagopyrum esculentum, and taro, Colocasia esculenta.
Some families have two names; the above rules were adopted to regularize family names so that each family is named after one of its genera, using the -aceae suffix. The rose family was named Rosaceae, based on the genus Rosa. This worked well for most families, but for some, the old name was so well known and so familiar that it was kept as well. Examples are Asteraceae (Compositae: daisies, asters, sunflowers), Fabaceae (Leguminosae, legumes: beans, peas, peanuts), Aracaceae (Palmae, palms), Poaceae (Gramineae, grasses: wheat, rice, corn), Brassicaceae (Cruciferae, mustards), and Apiaceae (Umbelliferae, umbels: celery, dill).
The word “taxon” (plural, taxa) is used to refer to any of the above groups in a general way. For example, some systematists study smaller taxa such as species and genera; others are concerned with higher taxa such as orders and divisions.
![]() Cladistics
Cladistics
The study of phylogeny centers on examining the similarity of one species to others. A species evolves into two species as different populations accumulate distinct alleles; even when enough divergence has occurred to create separate species, the two still resemble each other strongly. As they continue to evolve, each acquires its own mutations, and because they cannot interbreed, they cannot share the new alleles. Thus, they differ from each other more as time passes. Distantly related plants have been on separate lines of evolution for millions of years, time enough for so many mutations to accumulate that they resemble each other only slightly. Cladistics is a method of analyzing these phylogenetic, evolutionary relationships.
Cladistic studies are complicated by the fact that plants can resemble each other for two distinct reasons: (1) they have descended from a common ancestor, or (2) they have undergone convergent evolution. Features similar to each other because they have descended from a common ancestral feature are synapomorphies (homologous features). For instance, almost all members of the anthurium family are easily recognizable because of their spathe and spadix inflorescence (FIGURE 18-1). All members have these structures because all have inherited their inflorescence genes from a common ancestor. The spathe and spadix are homologous in all Anthurium species. Homologous features are the ones critically important for making taxonomic comparisons and the only ones that can be used to conclude that species are related.
The second cause of resemblance, convergent evolution, results when two distinct evolutionary lines of plants respond to similar environments and selection pressures. Under these conditions, natural selection may favor mutations in each line that result in similar phenotypes. Features like this are homoplasies (analogous features) and should never be used to conclude that plants are closely related. A striking example is the convergent evolution of cacti and the succulent euphorbias. Both occur in deserts where a succulent water-storing body is advantageous, and spines are selectively advantageous by deterring animals from eating the plants to get the water. The two families are not considered closely related on the basis of spines and succulence, however, because these features do not share a common ancestry; they converged evolutionarily because of similar selection pressure.
Determining whether a similarity is due to homology (common ancestry) or analogy (convergent evolution) can be extremely difficult. In the case of spines, it is easy: Cactus spines are modified leaves that are always smooth and never branched, occurring in clusters on an extremely short shoot (FIGURE 18-2). Euphorbia spines are modified shoots that branch and have small, scale-like leaves. Being modified shoots, they occur singly in the axil of a leaf, never in clusters. If we look beyond the analogous similarities, the dissimilar features are dramatic; flowers of cacti are large and have many parts, whereas those of euphorbias are small and unisexual, either staminate or carpellate. The wood of cacti is parenchymatous and has few fibers, whereas that of euphorbias is fibrous and hard. Most important, the plants that most closely resemble the succulent euphorbias in important homologous features are the nonsucculent euphorbias of Africa. Succulent cacti resemble a completely different set of plants, nonsucculent cacti of the tropical regions of the Americas. Thus, the evolution of spines and succulence occurred independently in each group.
A corollary to the assumption that similar plants are closely related is the assumption that dissimilar plants must not be closely related. Studying lack of similarity can also be difficult because in some cases a small genetic change results in dramatic phenotypic changes (FIGURE 18-3). Mutations that affect production, distribution, and sensitivity to hormones result in large changes of the phenotype between two closely related species. Also, mutations that affect early stages of development such as the embryo or bud meristems can cause closely related species to look deceptively dissimilar. In many evolutionary lines, species of large woody trees have evolved into herbs; the main transition involved is the ability to flower early and then not survive a harsh winter. It must be a simple evolutionary process involving few genes because it has occurred many times. For example, nearly half the species of the Violaceae are herbs in the genus Viola—violets and pansies—but the early members of the family were woody trees, as are the members of the relictual, tropical genus Rinorea. In the cabbage family (Brassicaceae), artificial selection by humans has resulted in one species that has dramatically different forms: Broccoli, brussels sprouts, cabbage, and cauliflower are all subspecies of a single species, Brassica oleracea.
For taxonomic studies, a mosaic of evidence is available—some valid and useful, some misleading. Systematists attempt to look at as many features as possible on the assumption that misleading evidence will be outweighed by valid characters. Also, certain features are considered more significant; those that result from numerous metabolic interactions and the influence of many genes tend to change more slowly than those with a simple metabolism controlled by just one or two genes. Loss of features, such as loss of sepals and petals in wind-pollinated flowers, is often rapid because deleterious mutations occur commonly; without the natural selection of insect pollination to eliminate these mutations, they are retained and cause a loss of perianth. After those genes have become so mutated that a flower lacks sepals and petals, the evolution of new ones would be slow and difficult.
Currently, taxonomists study virtually every aspect of plants using a wide variety of tools. Simple observation of major parts is still important, but scanning electron microscopy to study hairs, stomata, cuticle, and waxes is also used. Internal structure is studied with both light and transmission electron microscopy; the nature of plastids in the phloem has been found to be an extremely valuable character. Various aspects of metabolism are important, ranging from the types of pigments in flowers to the presence of Crassulacean acid metabolism (CAM) and C4 metabolism and specialized defensive, antipredator compounds.
DNA sequencing is a new tool for analyzing evolutionary relationships. Two plants are considered separate species only if they differ in significant, heritable ways; therefore, the sequence of nucleotides in the DNA of each species must differ from that of all other species. When plant phenotypes are studied, the actual objective is to determine differences in genotype. In the past, this could be done only by examining phenotypic features. Now DNA can be examined directly and mutations can be identified, even if they do not cause any detectable change in phenotype (TABLE 18-2).

FIGURE 18-1 Members of the family Araceae (the aroids, Jack-in-the-pulpit) all have a spathe (the collar-like sheath) and spadix (the central column), as in this Amorphophallus (A) and Anthurium (B). In all extant species, the genes that control the formation of these structures are descendants of the genes of the original, ancestral species. The genes that exist today have resulted from thousands of rounds of DNA replication during plant growth, crossing-over, syngamy, and then more rounds of DNA replication as the zygote grows. This has been repeated millions of times since spathes and spadices first evolved millions of years ago. During all this, mutations and natural selection have led to obvious varieties of spathes and spadices, but the basic relatedness, the homology, is still obvious.

FIGURE 18-2 (A) Cactus spines are modified bud scales, which makes them modified leaves. They occur in clusters as all bud scales do, and they never branch. Discocactus. (B) Euphorbia spines are modified branches. They bear small leaves that may themselves have axillary buds; therefore, the spine can branch, as in this E. meloformis. Although the spines in cacti and euphorbia are often called homoplasies, they are so different it is difficult to confuse the two. (C) In contrast to homoplasy, another problem arises when a structure evolves so much as to be almost unrecognizable. It took a great deal of study to be certain that cactus spines are modified leaves, and they continue to evolve. These flat, thin structures are spines of Tephrocactus papyracanthus; they protect the cactus by disguising it. This species grows among grasses and is almost impossible to find unless it is flowering. (D) The spines of this Tephrocactus strobiliformis have evolved to be glands, extrafloral nectaries: Each droplet is a secretion of dilute sugar water that attracts ants to the plant. The ants protect the plant as they defend their source of food (plants in (B), (C), and (D) were cultivated by B. Barth).

FIGURE 18-3 This small, leafless shrub (A, Acacia aphylla) may not at first glance appear to have much in common with this larger, leafy tree (B, Acacia drepanolobium), but careful examination reveals homologous similarities in many critical features of the flowers and stems, which allow assignment to the same genus, Acacia. In this case, genes controlling flower and stem morphogenesis have changed little during evolution, whereas genes controlling elongation growth of stems, formation of leaf primordia, and formation of bark have all changed dramatically.
Like traditional taxonomic data, DNA sequencing data have their own ambiguities and limitations. Any two species, even those closely related, must differ at many points (nucleotide pairs) of their DNA. Simply counting the number of sequence differences is not enough, because even individuals of a very uniform, homogeneous species may have hidden mutations that do not affect the phenotype. Furthermore, any two species in the same family probably share the great majority of their DNA sequences in identical form, with only a few genes mutated. How many differences are needed to conclude that two genomes represent two distinct species? There is no simple answer.
DNA sequencing permits a new type of study: the evolution of DNA itself. Sequencing allows us to investigate the evolution not only of taxa but of centromeres, telomeres, DNA replication start sites, enhancer elements, boundaries between introns and exons, and so forth.
Understanding Cladograms
A cladogram is a diagram that shows evolutionary patterns by means of a series of branches. Each point at which a cladogram branches, called a node, represents the divergence of one taxon into two, and all of the branches that extend from any particular point represent the descendants of the original group. That ancestor is their common ancestor (FIGURE 18-4). Any ancestor (any node) and all of the branches that lead from it constitute a clade (thus, this is a cladogram). Because each node represents a taxon dividing into two, it also represents some detectable change that creates a new group: After the divergence, one of the taxa differs from the other. For example, a plant in a species with red flowers might undergo a mutation that gives it white flowers. As long as the two can still interbreed, this is not especially significant; however, the change in flower color may cause different pollinators to visit it, and it might become reproductively isolated from its red-flowered ancestors. If the white-flowered individuals thrive and become numerous—but cannot interbreed with the red-flowered ones—this is a new species, and the divergence should be represented as a node with two branches coming from it. In this case, white flowers are a derived condition, an apomorphy, and red flowers are the ancestral condition.
TABLE 18-2 DNA Sequences Used for Phylogenetic Studies
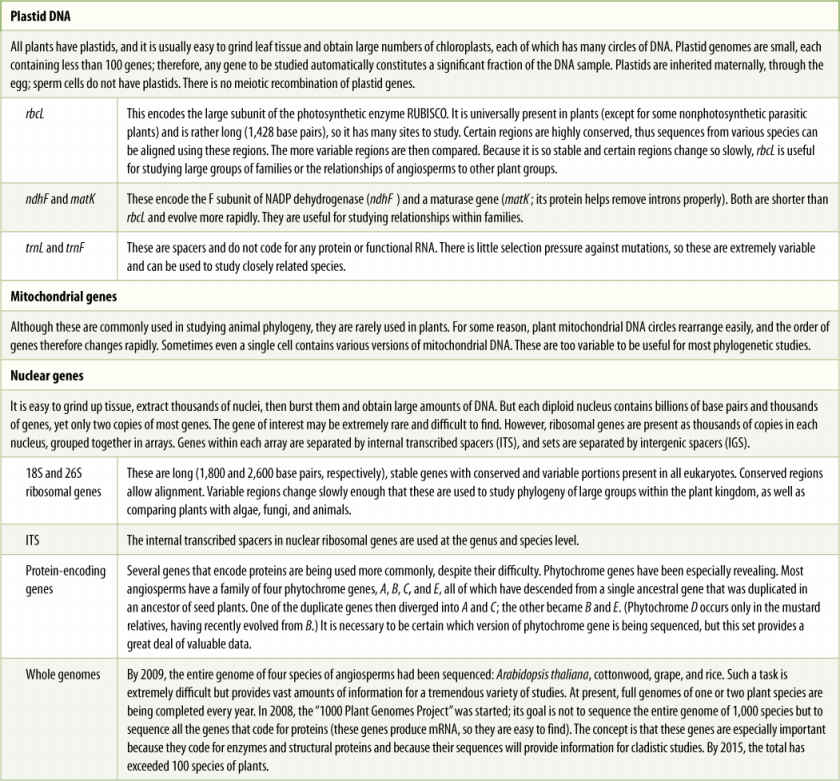
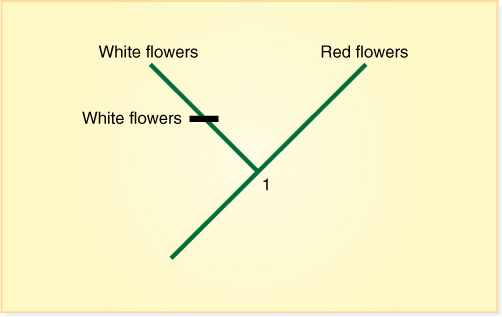
FIGURE 18-4 A simple cladogram showing the phylogenetic relationships between an ancestral group (at node 1) and its two descendants. The ancestral condition was to have red flowers. The group diverged into two; now one group has white flowers (an apomorphy), and the other still has red flowers. Be careful that this is treating only a very limited amount of information. It does not indicate whether we are dealing with two species and their ancestral species or whether we are comparing two genera or two families.
Both populations might continue to evolve and diverge, giving rise to a phylogeny like that shown in FIGURE 18-5. Node 1 here represents the same taxon as node 1 in Figure 18-4, but now with more time, both its lineages have diverged: At node 5, some that had continued to have red flowers have diverged into two groups, one continuing to have red flowers and the other mutating to have white flowers. White flowers have now originated twice. Synthesis of pigments requires many steps and many enzymes, and mutations in any of those genes could cause the same phenotype. The two sets of white flowers almost certainly have different genotypes. This is homoplasy. The original white-flowered lineage has diverged twice: After node 2, one group has trichomes on its leaves, and the other continues to have smooth leaves. At node 3, dwarfism originates in one line; at node 4 one group mutates to have narrow petals.
We can speak of many clades here: All of the species depicted, plus their common ancestor at node 1, form a clade. Species E and F plus node 5 are a smaller clade, as are the four species (A to D) that have descended from node 2. It is especially important to notice what is not a clade: There are now five species with white flowers (A, B, C, D, E), but we cannot classify E with the others—they do not form a clade. The most recent common ancestor of all white-flowered species is node 1, and a natural, monophyletic clade includes the ancestor and all of its descendants. If we would try to classify A—E as a group but leave out F, we would be creating a paraphyletic group, one that does not contain all the descendants of the most recent common ancestor (TABLE 18-3).
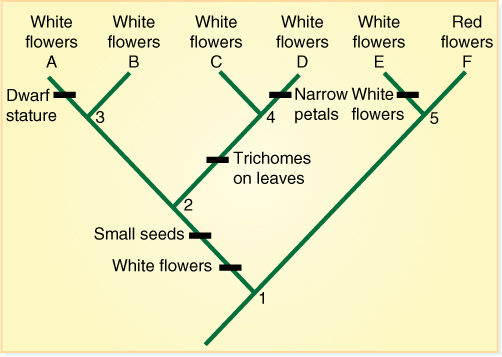
FIGURE 18-5 A more complex cladogram with more taxa and characters. We have data for the following characters: flowers (white or red), seeds (large or small), leaves (with or without trichomes), plant height (tall or dwarf), and petals (wide or narrow). In all taxa, the fruits are berries. Imagine that the ancestral group had red flowers, large seeds, leaves without trichomes, tall shoots, and broad petals (we probably would not know this beforehand; the cladogram is helping us determine what the ancestors were like). See the text for details.
Think about what a systematist is faced with. There are six species, some with white flowers, some with trichomes on their leaves, some with a dwarf stature, and some with narrow petals. In constructing a cladogram, we search for shared derived characters, synapomorphies. Species C and D both have trichomes on their leaves, but no other species does; therefore, this is a synapomorphy that shows these two are closely related. Clades A—D share the synapomorphy of having small seeds. Species A is the only one that is dwarf: No other species has this feature, so it does not help us understand the phylogeny—it is uninformative. All species still produce fruits that are berries. This has not changed anywhere; this shared ancestral condition is a symplesiomorphy, and it too is uninformative for arranging these species.
Given these six species and these characters, a systematist could create any number of cladograms. Between node 1 and species A we could imagine many steps in which flower color changed from white to blue to yellow and then back to white, with the blue and yellow forms having gone extinct. This might have happened: We can see the six living species and their characters; however, we cannot see any of the ancestors (they are all long since dead), and there may have been other descendants that died out and thus are missing from our cladogram. Given the information we do have, we use a principle called parsimony: We prefer the simplest possible hypothesis, and we do not make a hypothesis any more complicated than it needs to be. Parsimony is used in all fields of biology. The most widely accepted theory of water movement in vascular plants is that water moves upward through xylem. We could imagine all sorts of much more complicated mechanisms, but we focus on the simplest one and move on to a more complex hypothesis only if data indicate that the simple hypothesis is not accurate.
Botany and Beyond
BOX 18-2 Cacti as Examples of Evolutionary Diversification
The cactus family (Cactaceae) is a large group that has undergone extensive evolutionary diversification, with the modification of many characters. Many of the changes are easy to see and understand, so this is a good group to use as an example. First, several morphological and DNA features unite the entire family, the most obvious is that all have spines that evolved from modified bud scales. They are present in almost all members (Blossfeldia no longer makes spines), and no other family produces spines this way.
The first divergence at node 1 (see Figure B18-2O) produced one clade labeled Pereskia and another clade that leads to all the other cacti. The Pereskia clade actually branches into several species (not shown here), all of which lack many derived characters: These are large trees with ordinary broad, thin photosynthetic leaves, hard woody trunks, and no succulent tissue. These features are what we would expect in an ordinary eudicot tree; therefore, we think that many of these are ancestral characters, not derived ones. Pereskias are so similar to each other and so different from other cacti that they are grouped together as a subfamily, Pereskioideae.
Nodes 2 and 3 lead to Maihuenia and Opuntia. Plants in these clades are very similar to pereskias, but each has several derived characters. Plants in both are small, woody shrubs with small, cylindrical leaves that appear to consist of just a midrib without any lamina. Their cortex is thicker, with more water-storage parenchyma than that of pereskias. Opuntias differ from maihuenias and all other cacti in having hundreds of tiny bristle-like spines (called glochids) that are a synapomorphy distinguishing all opuntias from all other cacti. Maihuenias are distinguished from Pereskia and the opuntias by DNA sequence differences. Maihuenias (there are only two species) are placed in their own subfamily Maihuenioideae, and the opuntias (there are many genera and species) are in subfamily Opuntioideae (prickly pear cacti and chollas).
Node 4 is a real surprise. One branch leads to Blossfeldia, a monotypic genus (there is only one species, B. liliputana) of plants that have many derived characters. They are tiny dwarfs only about 1 cm tall when mature. Their foliage leaves are reduced to just leaf primordia. They have stomata in their stem epidermis, and their wood has an unusual type of tracheid but no xylary fibers. Until recently, B. liliputana was placed up by node 10 among several other genera that have similar, highly derived features, but DNA sequencing by several people indicates that it is a sister to all of the cacti from node 5.

FIGURE B18-2B Pereskia sacharosa, spines are modified bud scales. Notice the large green foliage leaf.

FIGURE B18-2C Maihuenia poeppigii. This is a cushion plant, a short, extremely compact tree approximately 20 cm tall; this is a common adaptation to cold, windy alpine habitats. Yellow structures are fruits.

FIGURE B18-2A Plant of Pereskia sacharosa.
Cacti from node 5 are extremely variable, but all are more “cactus-like” than pereskias and maihuenias. These really look like cacti: They are succulent, have water-conserving and water-storing adaptations. Those at node 8 (Leptocereus, Pachycereus, and their relatives) still have many ancestral features, especially their hard, fibrous wood that has few adaptations for desert life. Taxa from node 6 are extremely derived with fewer ancestral characters: They tend to have spherical or short cylindrical bodies (which have a low surface-to-volume ratio that helps reduce water loss by transpiration). Many have specialized spines that shade the body, disguise it, or secrete nectar. Also, this clade occurs only in North America, not South America, and geographic distribution is often an important character (closely related species tend to be found in the same area, not scattered all across the world).

FIGURE B18-2F Blossfeldia liliputana. This is a mature, full-grown plant, approximately 1 cm tall.

FIGURE B18-2G Mammillaria lasiacantha and many of its relatives are small, desert-adapted plants with many derived features that occur only in cacti.

FIGURE B18-2H Echinocactus horizonthalonius is one of many species of barrel cacti in North America.

FIGURE B18-2D Tiny but obvious foliage leaves of Maihuenia poeppigii.

FIGURE B18-2E Austrocylindropuntia schaferi has leaves without a lamina; its wood is hard and fibrous, and it has little succulent tissue.
All clades from node 9 are predominantly South American; only a few individual species have spread to the Caribbean, Florida, Central America, or Mexico. Hatiora and its relatives are epiphytic, living only in rain forests, not a habitat we typically associate with cacti. Of the remaining clades, Parodia, Gymnocalycium, and their relatives are small, globular South American cacti that greatly resemble the North American cacti of node 6: In fact, early taxonomists often classified them together. One of the original reasons for considering them distinct is that the two clades (node 6 and node 10) occur so far from each other, separated by thousands of miles of ocean and Amazon rain forest (both are significant isolation barriers). Later, certain flower, fruit, and seed differences were noticed, and more recently, DNA sequencing also supports the idea that North American small cacti and South American small cacti are remarkable cases of convergent evolution within the same family. The most recent common ancestor of node 6 and node 10 is node 5, and plants at that node must have lacked many derived features while having many relictual ones (shrubby, hard wood, little succulence).
This family has tremendous diversity. It contains trees, shrubs, and dwarfs (but no herbs—all still make at least a tiny bit of wood); succulents and nonsucculents; plants adapted to deserts, rain forests, grasslands, and alpine regions (some grow next to snowbanks); flowers pollinated by insects, bats, and hummingbirds. The monophyly of the family is not in doubt, however. All are universally accepted as being members of the same family-level clade.

FIGURE B18-2J Pachycereus pringlei has a very thick cortex and is capable of storing large amounts of water. Its wood, however, is like that of Pereskia—fibrous and strong enough to support tremendous weight.

FIGURE B18-2I Leptocereus weingartianus is the shrubby, brushy cactus sprawling out of the undergrowth; its stems are narrow, with little succulent tissue, and it is highly branched, like an ordinary noncactus bush. The tall, upright cacti are Pilosocereus polygonus.

FIGURE B18-2K Hatiora salicornioides is an epiphytic cactus with tiny, narrow stems; it never becomes a large plant.

FIGURE B18-2L Parodia maassii are small globular South American cacti whose body size and shape are similar to those of Mammillaria because of convergent evolution.

FIGURE B18-2M Gymnocalycium horstii has obviously diverged greatly from its Pereskia-like ancestors.

FIGURE B18-2N Oreocereus celsianus are adapted to high, cold alpine conditions on the border between Bolivia and Argentina.
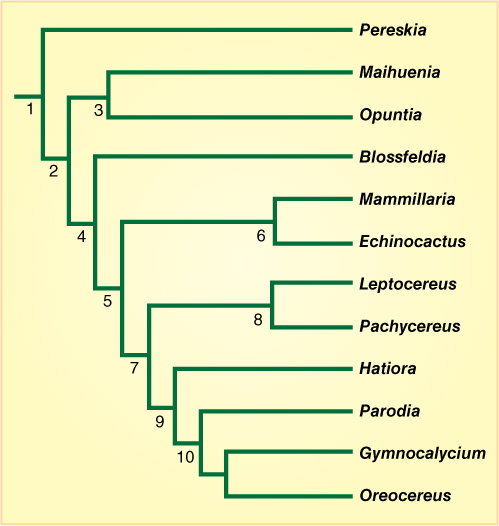
FIGURE B18-2O Simplified cladogram of several clades in Cactaceae.
In constructing cladograms, a great deal more data are used than those shown in Figure 18-5. For living species, we have DNA sequences in which each base pair is a data point, and thus, we might have dozens or hundreds of informative sequence differences when comparing six species. However, when a computer program uses the data to construct a cladogram, it almost always comes up with several that are equally simple—equally parsimonious—but that have the taxa arranged differently. When this happens, the various cladograms are analyzed to see if any clade appears repeatedly; if so, we have confidence that that particular clade might be an accurate depiction of the phylogeny. Sometimes there is simply not enough data, and then usually the various species are shown as arising from the same node. This is an unresolved polychotomy. These become the focus of more intense study.
As you study cladograms, keep in mind that they are hypotheses. Each is a model supported by some evidence, but we do not have all possible information about all species either living or extinct. DNA sequencing projects are progressing very rapidly and providing large amounts of data. These can be combined with morphological, anatomical, and physiological data to refine the cladogram and to understand not only how taxa are interrelated but also how the characters themselves have evolved.
TABLE 18-3 Cladistics

Cladograms and Taxonomic Categories
How are names or taxonomic levels assigned to a cladogram? The only taxonomic unit with an objective definition is species: Individuals that can interbreed. But there is no such objective definition for genus or family or so on. It has been proposed that every node and every clade in cladograms be names, but that would result in an unacceptably large number of names. Even worse, most cladograms are still not stable: They change as new species and data are added. Many systematists have decided to continue using the old names for genera, families, and orders unless they are shown to be definitely polyphyletic or paraphyletic. This has resulted in several well-known families being combined into one or divided into two. Informal names are temporarily being used for certain groups until we have more confidence that we truly understand their phylogeny. An especially important example for us involves angiosperms. Until recently, we believed that all angiosperms were either monocots or dicots, that the “monocot/dicot” divergence occurred extremely early (FIGURE 18-6); however, now there is strong evidence that early angiosperms diverged into several clades before that happened, and the living descendants of those early-diverging clades are no longer called dicots. They are just called the basal angiosperms and have not been given a formal taxonomic name. The rest of the dicots are called eudicots, and this too is an informal name. The old word “dicot” has to be abandoned because the group it referred to is now known to be paraphyletic (it included basal angiosperms and eudicots but left out monocots, which share a common ancestor with eudicots).
![]() Other Types of Classification Systems
Other Types of Classification Systems
Artificial Systems of Classification
The natural system of classification, which attempts to follow the evolutionary history of the organisms being categorized, is only one of several types of classification system. Another fundamental type is an artificial classification system, in which several key characters, often very easy to observe, are chosen as the basis of classification. Good examples are roadside floras and picture guides to plants, birds, and mammals of national and state parks. The botanical classifications in these are often based primarily on flower color: All plants with white flowers are grouped together, as are all those with red flowers, and so on. Within each category, the next classification category might be the plant’s habit: Trees are grouped together, as are shrubs, herbs, vines, and so on. These systems are described as artificial because many plants in a category are not closely related to each other by descent from a common ancestor; furthermore, they are separated from their close natural relatives in other categories simply because they differ in flower color or habit.
Artificial systems typically have the goal of easy plant identification by means of obvious characters such as flower color and plant habit. Alternatively, an artificial system may be designed to group together plants with economically or scientifically important features. From a practical standpoint, carpenters and woodworkers are more interested in color, texture, grain, and hardness of a wood than its phylogeny. Gardeners might classify plants according to their ability to tolerate shade, full sun, frost, or alkaline soils.
Plants and People
BOX 18-3 Genealogy Versus Clades: Your Family History Is the Opposite of a Clade
It is easy to confuse cladistics with genealogy. When tracing your family tree, you consider your parents, their parents, the next set of parents, and so on into your past. You are trying to locate your past relatives, but the real point of a genealogy is to map out all the people who might have contributed alleles to your own genome. You received all your alleles from your two parents, but those alleles came from your four grandparents, your eight great-grandparents, and your 4,096 great-, great-, great-, great-, great-, great-, great-, great-, great-, great-grandparents, and so on. At each generation you are considering the potential input of new alleles, and also you double the number of ancestors you are considering. Your family tree becomes broader as you go farther back.
A cladogram is just the opposite. We are interested in one single set of ancestors or founder individuals and how their progeny change with time. The ancestral group is genetically isolated from all other groups (remember, only members of the same species can interbreed), so their only source of new alleles is mutations within the descendants. A cladogram attempts to map out the origins of new alleles or of important new combinations of alleles that lead to detectable changes in phenotype.
A genealogy maps the genetic convergence of characters from many ancestors into one descendant over a few generations. A phylogeny maps the evolutionary divergence of characters and progeny over thousands or millions of generations.
Artificial classifications are only used as adjuncts to natural systems. As physiologists examine the phylogeny of all species with C4 metabolism, they find that C4 metabolism has evolved several times; therefore, all C4 metabolisms are not expected to be identical. Similarly, someone studying the metabolism of petal pigmentation might classify flowers artificially according to color, deciding to investigate the synthesis of red pigment first. But there are two very different types of red pigment: anthocyanins in most angiosperms and betalains in the order Caryophyllales (beets, cacti, portulacas). If some scientists were working with red rose flowers and others were studying red beets, they would get inexplicably conflicting results about the enzymes and pathways involved in pigment synthesis.
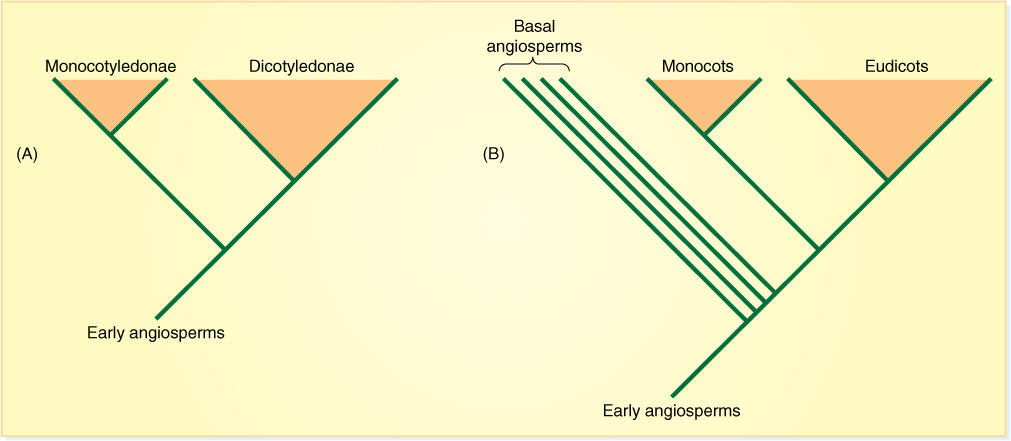
FIGURE 18-6 (A) A cladogram that represents the historical view that all angiosperms were either monocots or dicots. These had the formal taxonomic rank of class: Monocotyledonae and Dicotyledonae in some books, Liliopsida and Magnoliopsida in others (Table 18-1). (B) This very simplified cladogram summarizes a current hypothesis of evolutionary relationships within angiosperms. The “basal angiosperms” diverged from early angiosperms before the monocot/dicot split. All basal angiosperms had been known as dicots, as had all eudicots, but that concept of dicot would be a paraphyletic group, as it would leave out some descendants (the monocots) of the early angiosperms. We do not use paraphyletic clades, and therefore, we need a new name for the basal angiosperms (they are usually just called basal angiosperms) and for the dicots to the right of monocots (they are now called eudicots).
Classification Systems for Fossils
A third type of classification, used for fossil organisms, combines features of both artificial and natural systems. The goal is to understand the evolution of the fossil and to identify both its ancestors and its relatives that might have later evolved into other species. This requires a natural system. Typically, we do not know much about the fossil; thus, superficially similar ones are grouped together for convenience—a basically artificial system. The groupings are form genera: All fossils with the same basic form or structure are classified together. For example, a piece of fossil wood similar to the wood of modern pines, spruces, and larches is classified in the form genus Pityoxylon. If the piece of fossil wood was part of a branch with leaves and cones attached, there would probably be enough characters present to allow us to assign the wood to a natural genus, but if the fossil contains only wood, not enough characters are present to determine whether it came from an ancient pine, spruce, larch, or some other group that has since become extinct.

FIGURE 18-7 This Lipanthes orchid is fully grown; even in full flower, it could be overlooked easily. The photo is twice life size.
![]() Taxonomic Studies
Taxonomic Studies
People have been hunting for and discovering new plant types since before recorded history to find not only new food plants but also ornamental plants and beautiful woods. Exploration continues today as biologists enter new areas that either have never been explored or have been visited only once or twice. Tropical rain forests of Brazil, Central America, Africa, and Southeast Asia are so poorly explored that any serious expedition returns with previously unknown species of plants and animals. Some plants are less than 1 cm tall when fully grown and have an extremely limited range; it is easy to walk past them without noticing them (FIGURE 18-7). On the other hand, some large plants are too big to overlook, but they grow in only one small area and can be missed easily by explorers. Finally, a species may resemble others so closely that a scientist may ignore it, thinking it is already well known; the fact that it is a new species might be revealed only by careful study of certain details.
Botanical exploration is also concerned with discovery of new facts about already recognized species. No species is made up of absolutely identical individuals; therefore, numerous samples must be collected to gather information about the variability of its features as well as its geographical and ecological range. Exploration may be dedicated to gathering either seeds or live material so that the plant can be propagated for research, horticulture, or food. Now that recombinant DNA techniques make it possible to transfer genes from one species to another, live preservation of as many species as possible is particularly important.
Preliminary studies of newly collected plants include many diverse activities. Specimens must first be identified using diagnostic keys and personal knowledge. An experienced taxonomist often recognizes the family of an unfamiliar species almost immediately; even the genus may be obvious. If so, further identification is usually simple. In an unfamiliar region in which many species are unknown, identification can be more difficult, and numerous characters must be studied and compared with those in published descriptions of the plants of the area.
If the plant can be identified, it may be studied to see if it provides new information about the species. Even if it turns out to be absolutely typical in every way, that too is important. If the specimen cannot be identified (Box 18-4) or if it does not match any description, the taxonomist may suspect that it is a new species. Typically, the specimen is then sent to a specialist experienced with the species most closely related to the specimen.
Declaring a plant to be a new species is easy; proving it and describing it properly are difficult. Anyone can declare a plant to be a new species, name it, and write about it, even if it is obviously not a new species; this has caused countless problems. To overcome this, taxonomists from all over the world have established an International Code of Botanical Nomenclature that describes precisely the steps necessary for naming a new species. A valid name, one never previously used, must be declared and must be accompanied by a detailed description of the species in Latin and usually also in English, French, German, Spanish, or Russian. The name and description must be published in a widely circulated journal, a step that prevents many problems. The journal’s editors send the description to at least two independent specialists to verify that it is a previously unknown species and that the name has never been used before. The description must also include the designation of a type specimen; this is a single preserved plant that truly carries the name.
When new species are named, very little is known about them; as more research is carried out in the following years, enough variation may be discovered to warrant the recognition of a second species. For example, the range of leaf sizes, types of trichomes, and types of nectaries may warrant classification into two or three species. The original type specimen determines which type of leaf size, trichome, and nectary goes with the first name. When doubt arises as to which type of plant goes with which name, the type specimen must be checked. Type specimens are often kept in special fireproof cabinets and are not allowed out of the herbarium. During World War II, Allied bombing destroyed the herbarium in the Berlin Botanic Garden, and thousands of type specimens burned to ashes. Germans had been very active in plant exploration during the first part of that century and much of the 1800s, and the loss of those type specimens has caused tremendous problems. To prevent a recurrence of this disaster, other specimens, as similar as possible to the type specimen, are sent to many herbaria around the world; these are isotypes.
![]() The Major Lines of Evolution
The Major Lines of Evolution
All organisms are grouped into three domains: Bacteria (with cyanobacteria), Archaea, and Eukarya (FIGURE 18-8). The most significant event in evolution was the origin of life itself, probably about 3.5 billion years ago. The first organisms were simple, consisting of a cell membrane, protoplasm, and some means of inheritance, probably RNA; they almost certainly had no distinctive nucleus or other membrane-bounded organelles. Such organisms, either living or extinct, are prokaryotes. This line of evolution diversified rapidly into numerous clades, as evidenced by the presence of thousands of living species of bacteria, cyanobacteria, and archaeans. A significant step in prokaryote evolution was the development of a type of photosynthesis that liberates oxygen and is based on chlorophyll a. We do not know exactly which species of bacterium this occurred in, but their descendants are known as cyanobacteria, a clade within Bacteria, and one was the ancestor of chloroplasts.
The next major evolutionary event was the conversion of a prokaryote into a eukaryote, having a membrane-bounded nucleus. This must have been an extremely gradual procedure with many intermediates because many living species still have characteristics intermediate between prokaryotes and eukaryotes. A significant aspect was the origin of mitochondria; this was a gradual process in which a bacterium capable of aerobic respiration began living inside the protoplasm of an early eukaryote whose own capacity for aerobic respiration was less sophisticated. Both organisms would have benefited from their association; thus, this was a symbiosis, not a parasitism, and because one lived inside the other, it was an endosymbiosis. This is the hypothesis of the endosymbiotic origin of mitochondria, and there is a great deal of evidence to support it: Mitochondrial DNA, genes, and enzymes are more similar to those of certain bacteria than they are to the nuclei of the cells in which they are located. The most parsimonious (simplest) cladograms of mitochondrial genes are those that include bacterial genes as the early ancestral genes; trying to include eukaryotic genes into a mitochondrial gene tree requires many more nodes and steps and is much more complicated (less parsimonious).
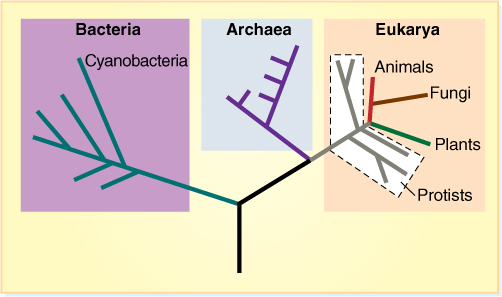
FIGURE 18-8 This cladogram represents our latest hypotheses about the interrelationships of the three domains: Bacteria, Archaea, and Eukarya.
Organisms with an organization similar to that of early eukaryotes were formerly classified in a “kingdom Protista,” but that classification was never satisfactory because it included members that were extremely diverse and obviously not closely related to each other. The criterion for classifying organisms in kingdom Protista was simply that they were eukaryotes but lacked the derived characters of plants, animals, or fungi; however, classification is based on shared derived characters (synapomorphies), not on a lack of them. Actually, the old classification of protista was a grade classification (as opposed to a clade): “Protistans” were placed together because they had a low level (low grade) of evolutionary advancement. Similarly, ferns and other plants that reproduce without seeds used to be classified together as the grade “Cryptogamae.” We no longer use classifications based on lack of characters, but the old names are often convenient; therefore, we use them informally, never capitalized, and occasionally in quotation marks to emphasize that “protists” do not form a clade.
After eukaryotes evolved to the level of having mitochondria, endoplasmic reticulum, and a true nucleus, numerous evolutionary lines emerged. One clade contained organisms that would later diverge into animals and fungi. Surprisingly, DNA studies are consistent with the hypothesis that fungi are more closely related to animals than they are to plants, despite superficial similarities such as lack of movement and reproduction by spores. The other early clade established an endosymbiosis with cyanobacteria, which then evolved into chloroplasts, producing the first algal cells. Just as with mitochondrial genes, phylogenies of plastid genes are most parsimonious if they go back to cyanobacterial genes instead of trying to include nuclear genes. The early algae continued to diversify, and approximately 400 million years ago, some became adapted to living on land, establishing the clade of true plants, kingdom Plantae. From these early pioneers, the major evolutionary lines that diverged were simple plants with neither seeds nor vascular tissues (mosses, liverworts, and hornworts), plants that do not produce seeds but do have xylem and phloem (ferns and similar plants), and the seed-bearing vascular plants (gymnosperms and angiosperms). The plants around us today represent thousands of evolutionary lines descended from early land plants, which in turn were derived from algae. Our classification system does not yet have every step identified, and not all relationships are clearly understood; however, the major features appear to be consistent with the data, and we expect that only relatively minor changes will be made as our knowledge increases.
Plants and People
BOX 18-4 Identifying Unknown Plants
When a plant must be identified, it can either be sent to an expert who will recognize it immediately, or it can be identified using a key, like the one shown here. Most keys are constructed of pairs of choices (couplets). Start at the first choice, and determine which description matches your plant; at the end of each line, there is either the name of the species or a number indicating the next set of couplets to use. The first choices are usually quite broad and easy, such as whether the plant is woody or herbaceous; this divides the plants into two completely artificial categories. This is acceptable because the key is used only for identification, not classification. As you proceed through the choices, you finally come to a name, meaning that you have tentatively identified your plant. The key should be accompanied by descriptions of all the species and, if possible, drawings or photos. After arriving at the name in the key, you should turn to the description to see whether it matches.
In this key, I have incorporated only the most common plants found in grocery stores, so you may try to identify a plant that is not in this key. If the plant does not resemble any plant in this key, you will soon run into a couplet in which neither description seems appropriate. In real life, this could mean that you have discovered a new species or a known species growing in an area where it had not been known to exist. Unfortunately, it most often means that you have made a mistake somewhere in the keying process.
Keys in floras or monographs usually are constructed in stages; a preliminary, small key identifies the plant to family or genus, and subsequent keys identify it further to species. As you go to each new key, you can check the description of the family or genus to see whether your keying has been correct to that point. With some experience and knowledge of the plants of an area, most people can recognize the family immediately and perhaps even the genus; therefore, they can skip the first keys and go directly to the appropriate level. The key shown here is really three keys: The first distinguishes between fruits and vegetables, and the other two are separate keys for fruits and vegetables; most of you should be able to identify your specimen as a fruit or a vegetable without having to go through a key. On the other hand, mushrooms present a problem. You do not know how I would treat a mushroom—it is reproductive like a fruit but has not developed from an ovary or associated structure; you have to use the first key for peculiar specimens like this.
As you use the key, keep in mind that you may have selected a plant that is not available in my store, so it will not be in the key.



 At the Next Level
At the Next Level
1. Mapping features onto cladograms. Constructing cladograms to display the evolutionary relationships among species, genera, and other taxa has been extremely informative. But many people treat this as only a first step. The cladogram is used as a map to study the evolution of the features of those taxa. For example, the pollinator of each species is listed on the cladogram’s line for each species, and we would expect that species pollinated by hummingbirds should be closely grouped together (indicating they are closely related) as would be those pollinated by bats, wind, butterflies, and so on. If we do this and find one hummingbird-pollinated species located among several bat-pollinated species, we have a dilemma. Either one species with bat-pollinated flowers evolved to have hummingbird-pollinated flowers (it is highly unlikely that large, white, open, night-blooming bat flowers will evolve to be small, red, tubular, day-blooming hummingbird flowers), so our original cladogram may have a mistake in it, and we need to correct it accordingly. Many characters can be mapped onto cladograms, including physiology, growth form, geographical location, and so on.
2. Cladistics at the next level. It is now extremely easy and inexpensive to extract DNA from specimens, then sequence it and create cladograms. The steps described in this chapter are only the basics. After sequences are obtained, they must be compared with each other and with related sequences stored in national and international online databases. In the olden days (less than 30 years ago), a person, usually a graduate student, would sit down with several sequences printed on paper and try to compare the As, Ts, Gs, and Cs of one with those of the others, using intuition and guesswork. Now this task is done by computer programs, and some programs (written by one set of people) make certain assumptions about data and rates of evolution, other programs (written by other people) make other assumptions. All programs make adjustments based on the statistical likelihood that errors are present and so on. Many review articles on cladistics will overwhelm you within the first few pages, but other articles are aimed at a an intermediate-level audience and should be appropriate for you, and you can learn a great deal about both biology and data handling by studying this. Having a lot of data is not the same as having an answer.
3. Horizontal gene transfer. A recently discovered phenomenon is horizontal gene transfer, in which a species picks up genes from an unrelated species, but not through sexual reproduction. Perhaps the transfer is caused by a virus that infects one organism, then some of its progeny infect a second while they are carrying genes from the first. In some cases, it even appears as if plants pick up genes from fungi that live with them. At present, we know little about horizontal gene transfer, we don’t even know if it is rare or common. If common, and if it really is possible for genes to somehow be transferred from fungi into plants, then we must be much more open-minded about what is occurring genetically in the world around us. Remember, one basic principle of the scientific method is skepticism: the willingness to accept new data, to keep an open mind.
SUMMARY
1. One of the goals of classification and nomenclature is to give each species a single unique name.
2. The classification system being developed at present is a natural one, attempting to reflect the actual evolutionary relationships—the phylogeny—of all species.
3. Artificial classification systems are now used only to identify plants or to categorize useful features but are never the basis for naming species.
4. Closely related species are classified in one genus, related genera are grouped into families, and so on through orders, classes, divisions, and kingdoms.
5. Each taxonomic category should be monophyletic, all of the organisms in it having evolved from the same ancestral group. Taxonomic groups discovered to have had two or more distinct origins are the result of convergent evolution; they are polyphyletic and the category is not a natural one; its members must be reclassified into monophyletic groups.
6. Homologous characters (synapomorphies) are those that have evolved from the same ancestral character, whether they now are similar to each other or not; these can be used as guides to phylogeny and the construction of a natural classification system.
7. Analogous characters (homoplasies) are those that resemble each other but have arisen independently; they are a part of convergent evolution and often cause confusion and misclassification because they appear to be homologous characters.
8. When a new species is declared, it must be given a name never before used, and it must be accompanied by a type specimen and a complete Latin description published in a widely circulated journal. The internationally recognized rules are occasionally not followed, leading to an invalid name.
9. All organisms are classified as Bacteria, Archaea, or Eukarya.
IMPORTANT TERMS
analogous features
ancestral condition
apomorphy
artificial classification system
basal angiosperms
binomial system of nomenclature
clade
cladistics
cladogram
class
common ancestor
division
equally parsimonious
eudicots
family
form genera
genus
grade classification
homologous features
homoplasies
inheritance of acquired characteristics
isotypes
key
kingdom
monophyletic
natural system of classification
node
nomenclature
order
paraphyletic group
parsimony
phylogeny
polyphyletic group
scientific names
species
species epithet
symplesiomorphy
synapomorphies
systematics
taxon
type specimen
uninformative
unresolved polychotomy
REVIEW QUESTIONS
1. What is the name of the science of giving things names?
2. One of the critical goals of biological nomenclature is to provide each species with a ______________ name. After the discovery of evolution by natural selection, taxonomists realized that the most scientifically valid name for each species would be one that reflected _________________.
3. Imagine that two completely unrelated species in very different families were accidentally given the same name. What kind of confusion would this cause? What if both were separately the subject of several scientific studies, and results were published in papers that did not mention the family name. Would it be possible to know which study concerned which species?
4. At the end of the 19th century, taxonomists adopted the goal of developing a natural system of classification. In a natural system, which kinds of organisms are classified together?
5. Originally, the main task of taxonomists was to discover new species and give them unique names. In a natural system of classification, they must also figure out the evolutionary relationships of the species they discover. Which do you think is more difficult: the original task or the modern one? Describe some of the reasons why it is more difficult.
6. Imagine trying to identify plants in your region by using a roadside flower guide. This type of a classification system is not a natural system. Instead, it is an ______________ classification system.
7. For fossils, we often must use a mix of artificial and natural classification. What is a form genus? If you found a specimen of fossil wood—but only wood, with no leaves or cones or bark—and it looked like wood of modern pines, what genus would you classify it in?
8. Theophrastus is often called the father of botany. When did he live? He had two very famous teachers. Who were they?
9. What is the book Materia Medica? Who wrote it and when? What kind of information does it contain?
10. In the 14th and 15th centuries, explorers brought to Europe information and specimens from Africa, the Americas and Asia, and this led to the realization that Pliny and other ancient Greek and Roman authorities were incorrect in many of their writings. Why was this significant? What did this realization lead to?
11. Linnaeus is well known for inventing the binomial system of nomenclature. In this system, each species has a ______________ name and a ______________ name.
12. Of all taxonomic categories, only species has an objective definition. What is it?
13. Closely related species are grouped together into _____________ (singular, _____________). By the way, what is the singular of “species”?
14. Study Table 18-1 and then, without looking at it, write all of the categories in order. Then write all the group names for Texas bluebonnet and for spider lily. Another bluebonnet, in the same genus as Texas bluebonnet, is Chisos bluebonnet, Lupinus harvardii. Write all of the group names for this species. Only one single word differs between it and Texas bluebonnet.
15. A critical concern of systematics is that the genera are __________________, that all the species included in the genus are related to each other by a common ancestor, and that (circle one: all, most, at least half) of the descendants of that common ancestor are in the same genus. Such a genus is (circle one: polyphyletic, monophyletic).
16. In an unnatural, ________________ group, members have evolved from different ancestors and may resemble each other only as a result of convergent evolution.
17. Give definitions for synapomorphy, symplesiomorphy, and homoplasy.
18. Sometimes two species appear similar because of convergent evolution not because they are closely related. Usually, it is safe to assume that if two plants do not resemble each other, they are not closely related. Is that always safe? Can two closely related plants be very dissimilar (Hint: Figure 18-3)?
19. Think about the apples we eat. They are in the genus Malus which is a member of the rose family, Rosaceae, along with roses, peaches, cherries, and even strawberries plus many other familiar plants. Almost all members of this family are trees or shrubs, and we are certain that the ancient ancestor was also a tree. If we are correct, is woodiness an apomorphy or plesiomorphy for the rose family? Being herbaceous is something new and rare in this family. Is it a plesiomorphy or an apomorphy? Apples, strawberries, and most other members of Rosaceae do not have a pit, but peaches and cherries do. We think this is a new character that originated just once, not in the ancestor of all the rose family but just in the ancestor of peach, cherries, apricots, and plums. If we are correct, is having a pit a symplesiomorphy, a synapomorphy, or a homoplasy? Rose bushes have spines and so do cacti, but the two families are not closely related. The spines of roses did not evolve from the same ancestral structure that cactus spines evolved from. The spines in these two families are a _______________________ and do not indicate that the families are closely related.
20. Have you tried using the Plants and People Box 18-4 to key out plants? Couplet 6 is used to distinguish between beets and turnips and relies on a character that is very easy to assess. What character is it? Couplet 4 distinguishes between carrots, beets, and turnips, even though only carrots are mentioned in couplet 4. If the plant being identified is a beet, what do you need to do at couplet 4 and in the next steps? Onions appear twice—first in couplet 11 and then in couplet 13. Why might onions be confusing enough to need to appear in two different places?
21. What are type specimens? Why are they important?
22. What is a monograph?
Design Credits: Hummingbird: © Tongho58/Moment/Getty; Green plant cells: © ShutterStock, Inc./Nataliya Hora; Purple tulip: © ShutterStock, Inc./Marie C Fields; Dandelion: © ShutterStock, Inc./danielkreissl; Poppy: © ShutterStock, Inc./Saruri; Plant icon: © ShutterStock, Inc./Vector; Digging man icon: © ShutterStock, Inc./Z-art