Botany: An Introduction to Plant Biology - Mauseth, James D. 2017
Genetics and Evolution
Nonvascular Plants: Mosses, Liverworts, and Hornworts
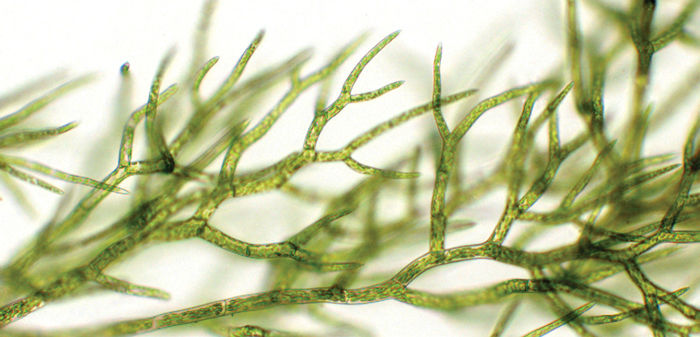
Chapter Opener Image: Is this a plant or an alga? It certainly looks like a green alga at first glance, but one hint that it is a plant is that it has many small chloroplasts per cell whereas most algae have only one or two chloroplasts per cell. When spores of mosses, liverworts, and hornworts germinate, the first cells that grow out strongly resemble filamentous green algae; this was one of the characters that led early botanists to suspect that green algae are the closest relatives of true plants. After this moss grows slightly larger, some of the tip cells will convert themselves into apical cells in the shape of an inverted pyramid. After this, all growth will be obviously plant-like with stems and leaves.
OUTLINE
✵ Concepts
✵ Characters of Nonvascular Plants
✵ Classification of Nonvascular Plants
✵ Division Bryophyta: Mosses
- The Gametophyte Generation
- The Sporophyte Generation
- Metabolism and Ecology
✵ Division Hepatophyta: Liverworts
- The Gametophyte Generation
- The Sporophyte Generation
✵ Division Anthocerotophyta: Hornworts
- The Gametophyte Generation
- The Sporophyte Generation
LEARNING OBJECTIVES
After reading this chapter, students will be able to:
✵ Summarize the changes and adaptations that allowed for the evolution of nonvascular plants.
✵ List four characteristics of nonvascular plants.
✵ Explain the morphology, water transport, development, and reproduction of mosses.
✵ Recall how water availability and retention affect moss sustainability.
✵ Describe liverwort gametophores.
✵ Compare liverwort sporophytes to moss sporophytes.
✵ Recall four characteristics of hornworts.
✵ Note how hornwort sporophytes differ from liverwort and moss sporophytes.
 Did You Know?
Did You Know?
✵ Many species of mosses and liverworts, when viewed with a hand lens, are as easy to recognize as wildflowers.
✵ The green moss plants you see are haploid; the small tan or brown capsules and stalks they sometimes have are diploid.
✵ Many mosses are adapted to deserts; dew forms on their tips on cool mornings and is conducted downward along the exterior of the moss.
✵ Many of the characters of hornworts and some liverworts are very similar to those of the first true plants.
![]() Concepts
Concepts
In this chapter the origin and evolutionary diversification of true plants—embryophytes—are discussed. Several critical events occurred in the history of true plants: their origin from green algae, their adaptation to terrestrial habitats, the development of the vascular tissues xylem and phloem, and the origin of seeds, leaves, and woody growth. Plants are traditionally divided into those that (1) have neither vascular tissues nor seeds, the nonvascular plants (often called “bryophytes”); (2) have vascular tissue but not seeds, the vascular cryptogams; and (3) have both vascular tissue and seeds, spermatophytes (FIGURE 20-1 and TABLE 20-1). Nonvascular plants arose first, and some of their later members were the ancestors of the vascular plants. Many of the fundamental aspects of plant biology were established in the early land plants.
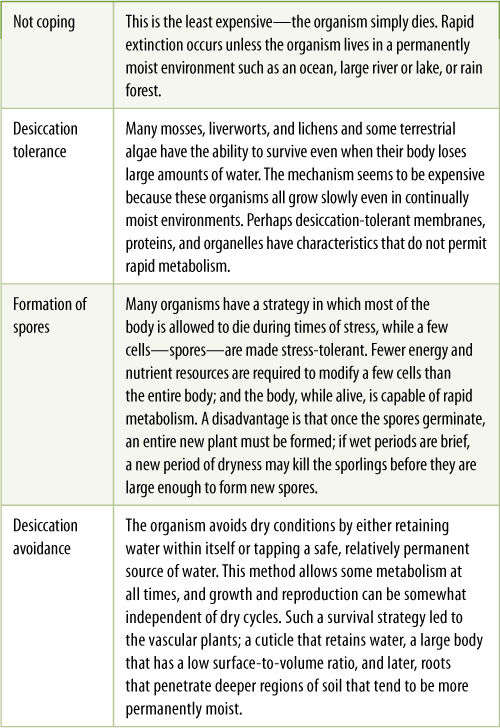
Beginning at some point earlier than 450 million years ago, certain green algae known as charophytes began to adapt to living on land. They evolved to survive the occasional drying of their streams, smaller lakes, and ocean-side mud flats. In environments where long, dry periods alternated with long, moist periods, the optimal survival strategy probably was formation of dormant, drought-resistant spores, but spore production requires major metabolic conversions; if the dry periods were brief, the ability to continue active metabolism by conserving water and avoiding desiccation would have been advantageous (TABLE 20-2). A large, compact, multicellular body would have had a low surface-to-volume ratio and would have automatically retained water better than a small unicellular or filamentous body. In addition, a water-proofing cuticle would have been selectively advantageous. Reproduction would have had to change to coordinate gamete production with periods of moisture because sperms had to swim. Also, gamete and spore mother cells would have needed protection from dryness by having a jacket of one or several layers of cells around them (FIGURE 20-2 and FIGURE 20-3). Such reproductive modifications resulted in the grouping and protection of spore and gamete mother cells into the sporangia and gametangia characteristic of all embryophytes—more massive than those of algae and protected by a layer of sterile cells.
TABLE 20-1 Major Groups of Plants
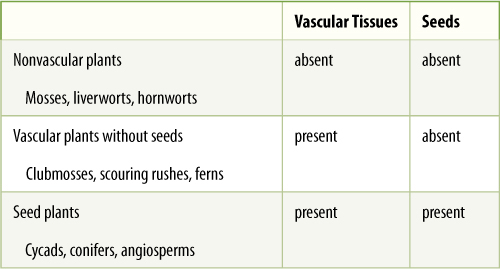
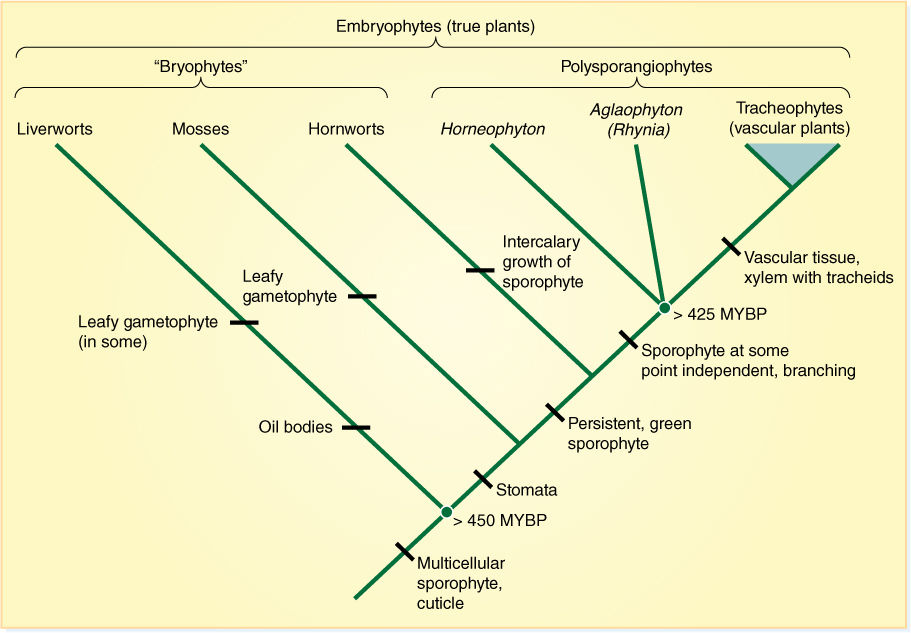
FIGURE 20-1 Recently proposed phylogeny of true plants. (MYBP = million years before present)
TABLE 20-2 Means of Coping with Dry Periods
These simple modifications probably allowed the algal ancestors of plants not only to survive but also to be metabolically active during short dry periods; however, an automatic consequence of much greater advantage was that the algae were safe from predators while out of the water. Tremendous selective benefit resulted from mutations that enabled the algae to be active for even longer periods out of water. These mutations probably involved increasing the size of the body and the impermeability of the cuticle, but there had to be the simultaneous evolution of stomatal pores and guard cells because a more protective cuticle also prevents the entry of carbon dioxide.
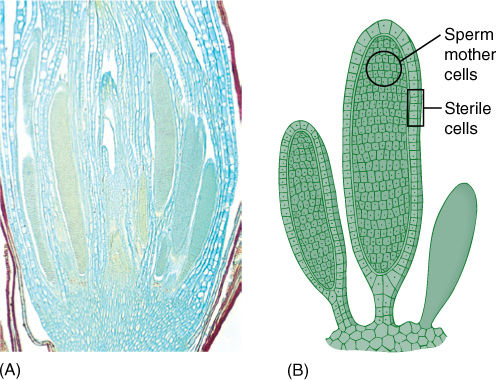
FIGURE 20-2 In true plants, gametangia and sporangia always have an outermost layer of cells that do not become gametes or spores. In these microgametangia of mosses, the sterile layer is only one cell thick, and each microgametangium produces numerous sperm cells (×25). Compare this with an algal gametangium.
As a truly terrestrial existence became more successful, the mud flats and stream banks would have become crowded, and some plants must have grown over others and shaded them. With such shading, the environment became selective for mutations that produced an upright body that could grow into brighter light. The nonvascular plants mostly use turgid parenchyma cells and thus cannot grow very tall, but in the group of plants that would become vascular plants, the strengthening material that evolved was xylem, which is also good at conducting water. As phloem evolved, the basal part of the plant that remained in the shade could be nourished. Phloem permitted roots to extend deep into the dark soil, and xylem permitted transport upward of the water and nutrients those roots encountered. The presence of xylem, phloem, and roots had profound consequences: They freed plants from their muddy habitats and allowed them to grow anywhere moisture was available. The presence of the two vascular tissues, necessary for individuals even a few centimeters tall, suddenly gave plants the potential to become 100 meters tall, to branch, to form leaves, and to put sporangia high into the air, allowing the spores to be blown great distances, carrying the plant’s genes over huge areas, and permitting the colonization of distant habitats.
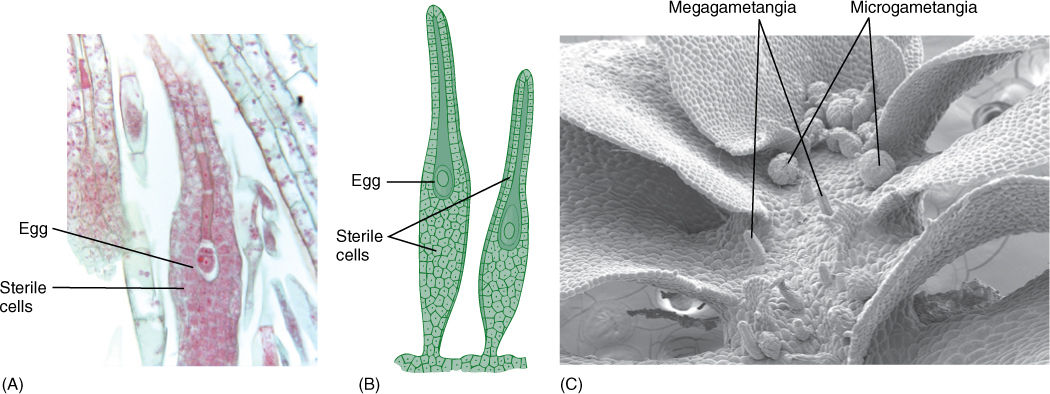
FIGURE 20-3 Megagametangia of true plants, such as these of a moss (A), also have a one-layered sterile jacket (B), but each produces only one egg, not four (×25). (C) This scanning electron micrograph shows the gametangia of a bryophyte (these are of a liverwort, but they are similar to those of mosses). The spheres are microgametangia in which sperm cells are produced, and the tubular structures are megagametangia, each with a single egg inside. The large flat sheets of cells are the leaves of this leafy liverwort (Fossombronia) (×50).
Vascular tissue, especially phloem, also made feasible the evolution of truly heterotrophic tissues—roots, meristems, and organ primordia. Without phloem, each part of a plant can grow and develop only as rapidly as its photosynthesis permits, but phloem allows mobilization of sugars, minerals, and hormones throughout the entire body and their transfer to a shoot apical meristem or a group of sporangia, thus permitting a more vigorous, robust growth than could otherwise occur.
One obstacle to the total invasion of land was that plants were still reproductively amphibious. They had terrestrial bodies but aquatic reproduction with swimming sperms. At some point in the life cycle, the environment had to be sufficiently wet that sperms could swim from one gametophyte to another. This requirement is a handicap, but not an insurmountable one: All plants need water to grow; no plant ever lives where there is never rain, fog, or dew (FIGURE 20-4).
Production of pollen and seeds eliminates the need for environmental water for reproduction. What changes must occur for seeds to evolve? Gametophytes must become so reduced that they mature completely within the walls of the spores (FIGURE 20-5). After this happened, retention of the megaspore and megagametophyte inside the parental sporophyte was feasible. The gametophytes functioned basically as tissues of the sporophyte and benefited from the land adaptations that had evolved; this is what occurs in ovules of seed plants. The microgametophyte, inside the microspore (known as pollen in seed plants), could be transferred by wind to the vicinity of the megasporangium, where it released sperms into fluids secreted by the megagametophyte. The sperms would swim in this tiny artificial pond and effect fertilization; only the last step of reproduction was still wholly aquatic. In the most derived seed plants, a pollen tube carries nonmotile sperms even further, to the egg itself.
Some gametophytes—for example, those of all nonvascular plants—do not need any protection by the parental sporophytes: In mosses, liverworts, and hornworts, gametophytes are the main photosynthetic phase of the life cycle (FIGURE 20-6), but even so, these gametophytes remain rather small and delicate, limiting the entire plant to environments mild enough for themselves (a plant can live and reproduce only in environments that are suitable to all essential aspects of its full life cycle). Plants should have been able to develop a life cycle in which gametophytes were just as large, tough, and woody as sporophytes, even having bark, leaves, roots, and so on, but this never happened. Some early land plants did have rather large gametophytes, but in all lines of evolution, gametophytes became simpler and smaller.

FIGURE 20-4 This rock is covered with a moss (Grimmia laevigata, the gray-green material) and lichens (the blue-gray material). Mosses typically grow in moist, shady habitats; however, there are also large numbers adapted to dry, sunny conditions as here at Inks Lake State Park in central Texas. This moss thrives side by side with cacti and yuccas in the blazing sun and with only sporadic, infrequent rains. But the important thing is that it does rain at some point, and there is dew or fog at other times…that is true of absolutely every place where plants grow: No plant ever grows where there is never liquid water at some time. Mosses always have small, simple bodies, and rather than being a handicap, this is an adaptation that allows them to live in small, rocky areas that have only a bit of occasional water, too little water to sustain a large tree. Mosses are confined to moist areas because their sperm cells must swim from one plant to another; the sperms do swim, but there is always liquid water at some time, so sexual reproduction is possible. It is important to learn the characteristic features and habitats of plants, but it is also important to be aware of alternative adaptations and their consequences.
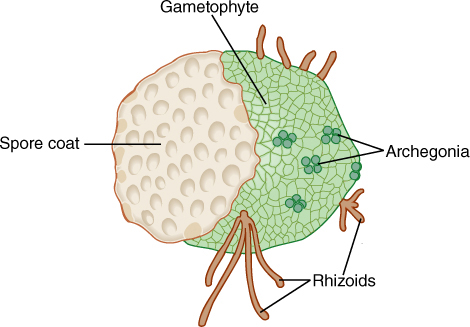
FIGURE 20-5 The megagametophyte of Selaginella, which has developed almost completely within the original wall of the megaspore. This is not part of a seed plant but is a necessary first step in seed evolution. Small megagametophytes can be protected and nurtured by their parent sporophyte. By doing so, the gametophyte generation benefits from all of those mutations that make the sporophyte adapted for life on land.
![]() Characteristics of Nonvascular Plants
Characteristics of Nonvascular Plants
Most people are at least somewhat familiar with nonvascular plants because mosses are well known, and many people have heard of liverworts (FIGURE 20-7 and FIGURE 20-8). The third group, hornworts, is relatively unfamiliar, even to most botanists (Figure 20-6 and FIGURE 20-9). It is important to understand clearly what nonvascular plants are not: Spanish moss of the southeastern United States is the flowering plant Tillandsia usneoides of the pineapple family, not a true moss. Club mosses are lycophytes, not mosses, and the slimy, bright green “mosses” of ponds and slow-moving streams are green algae, usually Spirogyra. Several types of lichens, especially Alectoria, Bryonia, Usnea, and “reindeer moss,” are frequently mistakenly thought to be mosses or liverworts.

FIGURE 20-6 The tiny leafy shoots are gametophytes of a small moss. The numerous plate-like structures are gametophytes of a hornwort, Phaeoceros. Plants of both remain small and thin, never more than a few cells thick, and never elaborate.

FIGURE 20-7 Although mosses occur in hot, arid areas, they are most abundant in cool, moist regions. What you think of as the typical moss plant—green or gray-green and leafy—are masses of gametophytes; the slender stalk and capsules you may have noticed are sporophytes.
Technically, nonvascular plants are embryophytes that do not have vascular tissue. Being embryophytes, they have multicellular sporangia and gametangia: Reproductive cells are always surrounded by one or several layers of sterile cells (Figures 20-2 and 20-3). Their bodies are not composed of filaments as in many algae, but rather of true parenchyma derived by three-dimensional growth, usually from apical meristems: All mosses and many liverworts have leafy stems that look remarkably like small versions of flowering plants. Nonvascular plants are almost exclusively terrestrial and have a cuticle over much of their bodies, and many have stomata.

FIGURE 20-8 This is Marchantia, the most abundant greenhouse liverwort; unfortunately, it is one of the least typical. As in mosses, the large green plant is the gametophyte, not the sporophyte. The cup-shaped structures contain clumps of cells that can be splashed out by rain and then grow into new plants.

FIGURE 20-9 Hornworts such as this Phaeoceros are quite rare, and few people ever see them; they are easily confused with liverworts unless the tall “horns”—sporophytes—are present.
*A assumes that they are less closely related than B does.
Like all plants, nonvascular plants have a life cycle with an alternation of heteromorphic generations; the sporophyte and gametophyte differ from each other structurally. Recall that sporophytes in flowering plants are the large plants with leaves and roots, whereas gametophytes are tiny and occur inside pollen grains and ovules. In nonvascular plants, the gametophyte is the larger, more prominent generation, and the sporophyte is much smaller, more temporary, and often very inconspicuous. Any green moss plant that might be familiar to you is the gametophyte, the haploid phase, and it is photosynthetic, perennial, and collects mineral nutrients; it typically can spread rapidly by asexual reproduction. The sporophytes it produces are usually very small. They absorb minerals only from the gametophytes and always remain attached to them. Sporophytes carry out so little photosynthesis that they could not support their own growth and sporogenesis. Unique sporophyte features in all species of nonvascular plants are that sporophytes are never independent of the gametophyte, never branch, and never have leaves. You will see sporophytes only if you examine mosses closely: They look like green or brown “hairs” standing up on the green gametophyte, but sporophytes are only present at certain times of the year.
Nonvascular plants can never grow to be really large, but being small and simple provides great selective advantage in certain habitats. The tiny parenchymatous bodies of mosses and liverworts permit them to thrive in microhabitats such as stone walls, fences, and bare rock, microhabitats that have too little water or soil for the larger, more complex vascular plants.
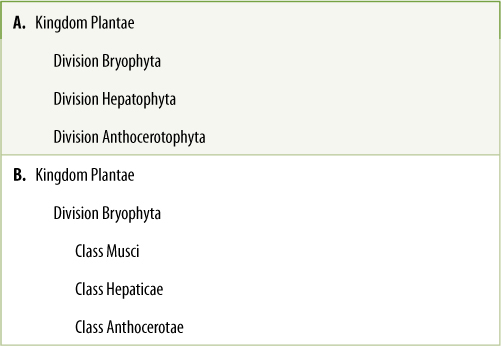
TABLE 20-3 Alternative Methods of Classifying Nonvascular Plants*
![]() Classification of Nonvascular Plants
Classification of Nonvascular Plants
It is not known how closely related mosses, liverworts, and hornworts are (Figure 20-1). They have many features in common but also differ in significant respects. They are often treated as three distinct divisions: liverworts, division Hepatophyta; mosses, division Bryophyta; and hornworts, division Anthocerotophyta (TABLE 20-3). In the past, all three were often grouped together in division Bryophyta as three classes: mosses, Musci; liverworts, Hepaticae; and hornworts, Anthocerotae. Even today, they are informally referred to as “bryophytes.” The three are treated here as divisions, and the generic term “bryophyte” is used only for mosses. All groups together are called “nonvascular plants.”
![]() Division Bryophyta: Mosses
Division Bryophyta: Mosses
The Gametophyte Generation
Mosses are ubiquitous, occurring in all parts of the world and in almost every environment (TABLE 20-4). They are perennial and thrive in many places within cities (Tortula on walls and Mnium on soil).
Morphology
The leafy stems, technically known as gametophores, of many moss plants grow close together, tightly appressed and forming dense mounds (Grimmia, Pohlia). In other species, particularly those of cool wet areas, the plants are more open and loose (Anacolia, Climacium, Platygyrium; see Figure 20-7). Scouleria gametophores grow as ribbons up to 15 cm long, submerged in rapidly flowing water. All moss stems have leaves (FIGURE 20-10), but because they are parts of a gametophyte, not a sporophyte, they are not homologous with those of vascular plants; that is, leaves of mosses did not evolve from the same structures as vascular plant stems and leaves did.
TABLE 20-4 Classification of Division Bryophyta
Moss gametophores grow from an apical meristem that contains a prominent apical cell (FIGURE 20-11). Derivative cells subdivide, producing the tissues of stem and leaves in rather precise arrangements. Leaves are aligned in three rows at least while young, and most have a midrib (costa). Leaves of almost all mosses are only one cell thick except at the midrib and along the margin (FIGURE 20-12A). In the family Polytrichaceae, common along roadsides in forested areas of the northern United States, genera such as Polytrichum and Atrichum have thin lamellae on the leaf upper surface, greatly increasing the photosynthetic tissue (FIGURE 20-12B). Cuticle occurs only on the upper surface of most moss leaves, the underside being uncutinized and capable of absorbing water directly from rain, dew, and fog. The lack of a cuticle means that mosses have little protection against desiccation; when the microhabitat dries, so do the leaves. No stomata occur on moss leaves; they would be useless because the leaves are unistratose.
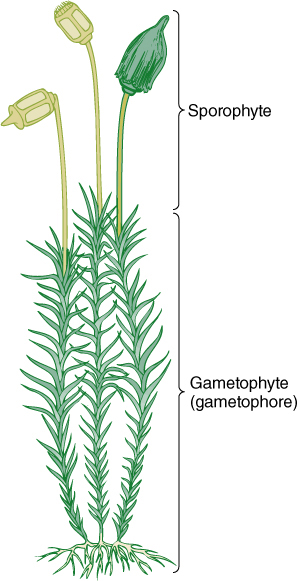
FIGURE 20-10 The individual shoots of a moss superficially resemble those of a flowering plant, having stems, leaves, nodes and internodes, and even buds. None of these structures is homologous to the equivalent organ in flowering plants, but the same set of names is used for both groups. Dark green, haploid tissue; light green, diploid tissue.
Moss stems are always slender and have little tissue differentiation (FIGURE 20-13). The surface layer is at most only slightly different from the underlying layers and is not called epidermis. The stem tissues, all called cortex, may be uniform in all parts, or the outer cells may be slightly narrower with walls that are somewhat thickened. Inner cells are larger, more parenchymatous, and chlorophyllous. In a few species of mosses, stems have hairs, but stomata do not occur.
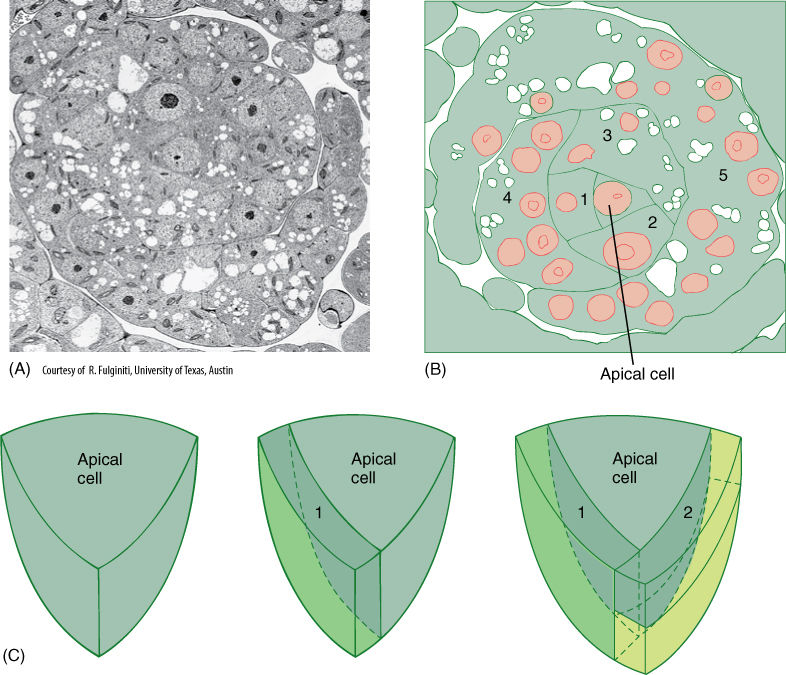
FIGURE 20-11 (A) and (B) Cross-section of the shoot apex of the moss Physcomitrium pyriforme: The apical cell is in the center, and its most recent daughter cell is to the left (1). The next older cell (2) has subdivided, as have successively older derivatives (3, 4, and 5) (× 2,400). (C) The apical cell is shaped like an inverted pyramid.
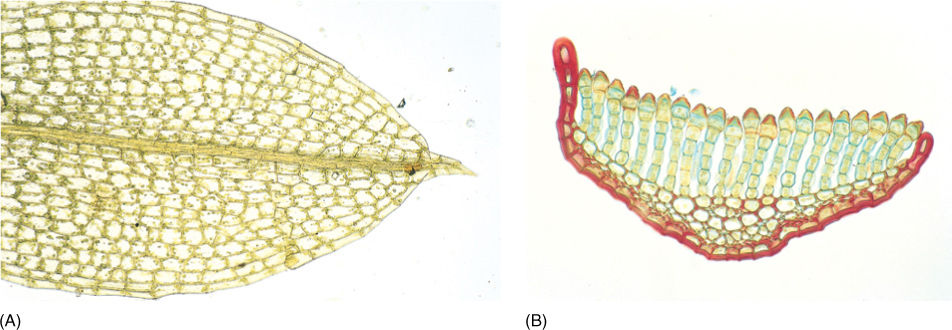
FIGURE 20-12 (A) Many moss leaves are only one cell thick, but the leaf “midrib” is thicker and is true parenchyma. (B) In the family Polytrichaceae, leaves bear long sheets of cells on their upper surface; this greatly increases the volume of photosynthetic tissue but does not decrease the high surface-to-volume ratio as it would if this were a solid tissue (×200).
Water Transport
In some mosses, primarily the family Polytrichaceae, the innermost cortex is composed of cells called hydroids that conduct water and dissolved minerals. They are elongated cells that lose their cytoplasm when mature (FIGURE 20-14); their end walls are partially digested, but they are not removed entirely. Each hydroid is aligned with those above and below it. Species that have hydroids typically also have leptoids, cells that resemble sieve cells (Figure 20-14B). They are elongate, have relatively prominent interconnections with adjacent cells, and lack nuclei at maturity, although they do retain their cytoplasm. Adjacent parenchyma cells are unusually cytoplasmic and rich in enzymes, just as are companion cells.
The majority of mosses lack hydroids and leptoids; water is conducted along the exterior of their stems by capillary action. Leaves and stems are so small they form spaces narrow enough to act as capillary channels and transport water. In some species, such as Funaria, leaves curl and become more closely appressed to the stem as they dry; when rain or dew occurs, the dried plant has even more capillary spaces than in the hydrated condition when the leaves are spread away from the stem. In mosses that lack leptoids, sugar is moved simply between parenchyma cells by slow transport.
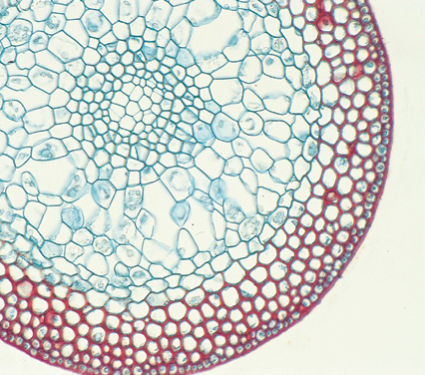
FIGURE 20-13 Although most moss stems do not have vascular tissues, they do support the shoot, and most have a layer or two of thick-walled cells (stained red here). Sugars and minerals must be transported from leaves to the shoot apex, gametangia, and sporophytes; therefore, living parenchyma cells are necessary (×40).
At the base of the stem are rhizoids, small, multicellular trichome-like structures that penetrate the surface of the substrate. Rhizoids only anchor the stem; they do not absorb either water or minerals. They lack chloroplasts and have reddish walls.
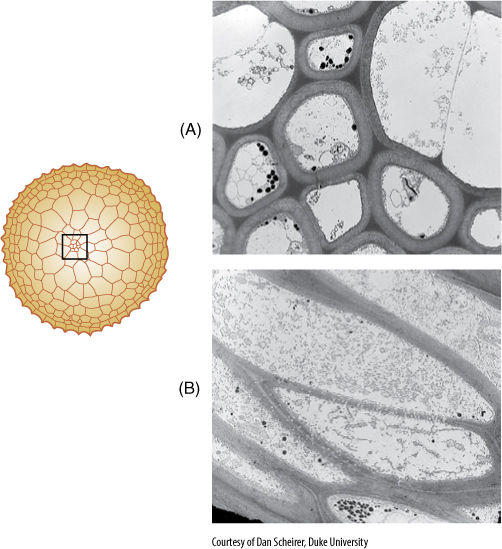
FIGURE 20-14 (A) Hydroids lose their cytoplasm at maturity, just as do tracheids and vessel elements. Hydroids, however, do not have specialized secondary walls. (B) Leptoids are analogous to sieve cells, phloem cells that are simpler than sieve tube members. All pores are small, and the cells remain alive at maturity.
Development
Growth of the gametophore begins when a spore germinates and sends out a long, slender chlorophyllous cell. This cell undergoes mitosis and produces a branched system of similar cells; the entire network is a protonema (plural, protonemata) (FIGURE 20-15 and FIGURE 20-16). A protonema superficially resembles a filamentous green alga but can be distinguished by its numerous small chloroplasts in each cell. Algal cells have only one or two large chloroplasts. Eventually, nodules of small cytoplasmic cells form on the protonema, organize an apical cell, and then grow upright as a stem with leaves—the gametophore. Protonemata are perennial and can grow extensively, producing many buds. The filamentous cells usually break when mosses are collected, and thus, a tuft of gametophores may appear to be independent plants when in fact they have all arisen from a single protonema.
Reproduction
The gametophore at some point produces gametangia. All mosses are oogamous; that is, every species has small biflagellate sperm cells and large nonmotile egg cells. Sperms are produced in microgametangia called antheridia, which consist of a short stalk, an outermost layer of sterile cells, and an inner mass of cells that differentiate into sperm cells (see Figure 20-2). Eggs occur in megagametangia called archegonia: Each is shaped like a vase with a long neck (see Figure 20-3). The neck is hollow at maturity, and the single egg is located at the base.
Antheridia and archegonia occur on the same gametophore in bisexual species (Funaria, Pottia), whereas other species have both male and female gametophores (Barbula, Polytrichum, Rhacomitrium). Depending on the species, gametangia may occur mixed with leaves along the gametophore stem or may be clustered at the stem tip, surrounded by a special cup-shaped whorl of leaves (FIGURE 20-17). Raindrops fall directly into the cup, splashing sperms out and carrying them as far as 50 cm. Without this mechanism, sperms must swim to archegonia (FIGURE 20-18).

FIGURE 20-15 A moss spore has germinated and grown into these filaments of cells, which make up a protonema; later some cells will divide into apical meristems that will grow into gametophores. Although this looks like a green alga, algae do not have so many chloroplasts in each cell (×250).
When sperm cells are mature, the antheridium breaks open and liberates sperm cells either by contracting the sterile outer cells or by accumulating liquid below the sperms and pushing them out. Secretion of sucrose from the archegonia guides sperms as they swim toward the archegonia and then down the neck to the egg, where one sperm cell effects fertilization.
The Sporophyte Generation
In all embryophytes, the megagamete and subsequently the zygote are retained by the gametophyte; however, unlike those of mosses, megagametophytes of vascular plants are small and offer little nourishment or protection to the embryonic sporophytes. Megagametophytes of flowering plants typically have only six cells other than the egg, and both synergids and antipodals often degenerate just after the egg is fertilized (undergoes syngamy). All nutrition for the zygote is supplied by the grandparent generation, the sporophyte, by way of endosperm formation. In contrast, moss gametophytes are both large and photosynthetic, and they support the sporophyte throughout its entire life. The moss sporophyte is never an independent, free-living plant (Figure 20-16).
The zygote of a moss undergoes a transverse division, and the basal cell, located at the bottom of the archegonium, develops into a small, bulbous tissue called the foot. The foot is the interface with the gametophore, from which it absorbs sugars, minerals, and water. Its cells are transfer cells in many species. The upper cell grows by cell division and expansion into a simple apical sporangium called the capsule, consisting of an outer layer of sterile cells and an inner column of sterile cells (the columella). A ring of sporogenous cells undergoes meiosis, producing haploid spores (FIGURE 20-19). Between the foot and the sporangium is a narrow stalk, the seta (plural, setae). All moss sporophytes have this basic, simple structure; none is ever branched or has leaves, bracts, or buds of any kind.
Although morphologically simple, the sporophyte is relatively complex structurally. It has a true epidermis with stomata at least on the base of the sporangium. Dehiscence of the sporangium is more elaborate than the opening of the gametangia: The apex of the sporangium differentiates as a caplike lid, the operculum, which separates from the rest of the sporangium as cells are torn apart. Cell breakage is elaborate and precise, resulting in one or two rows of beautiful, exquisitely complex teeth, called peristome teeth (Figure 20-19). The teeth respond to humidity, bending outward and opening the sporangium when the air is dry and bending inward and trapping the spores when the air is humid. Spores are released when they are light, dry, non-sticky, and easily carried by air currents. The apex of the sporangium in many species is covered by a calyptra, a layer of cells derived from the neck of the archegonium. As the embryo begins to grow, neck cells also proliferate. They keep pace with sporophyte growth at first but later grow more slowly and are torn away from the gametophore.
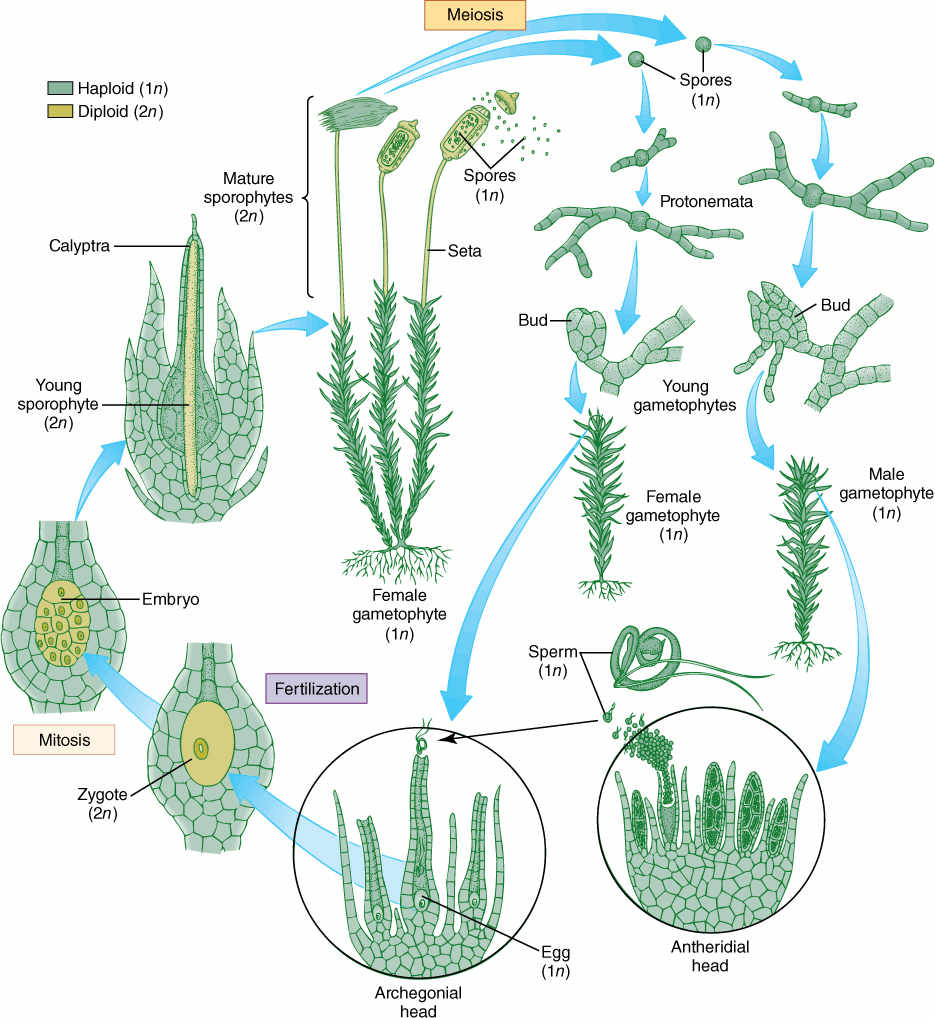
FIGURE 20-16 Life cycle of a moss. Details are given in the text.
Virtually all mosses are homosporous: All spores are the same size and appear to be identical. A few species of Macromitrium and Schlotheimia produce two types of spores in each capsule. The larger spores develop into large gametophytes with archegonia, and the smaller spores grow into dwarf males that live epiphytically on the females (Figure 20-17B).
Metabolism and Ecology
The small size and lack of conducting tissues are two critical factors in the metabolism and ecology of mosses. Vascular plants tend to be large and their bulk protects them from short-term fluctuations in air humidity and moisture availability. Should the air become dry for several hours, most seed plant leaves and stems would not die, even if they were not receiving water from the roots by means of xylem. Very large plants, such as trees and succulents, can withstand dry conditions for weeks, months, or even years before losing a fatal amount of water. Leaves on moss gametophores, however, have only a thin, incomplete cuticle; if exposed to dry air for even a few minutes, the plants dry out. Without vascular tissue, stems and leaves can become desiccated, even while the rhizoids are in contact with moist soil or tree bark.
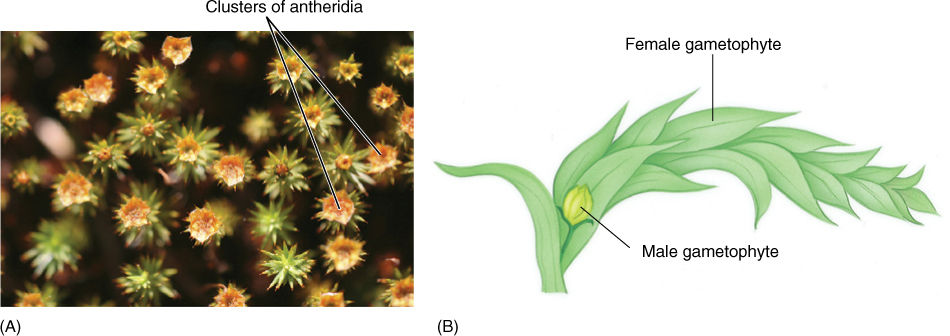
FIGURE 20-17 (A) In this moss, antheridia are clustered together at the gametophore apex; in other species, they might be located along the stem. (B) The phenomenon of dwarf males is common in many plants, animals, and algae. Each sperm cell is much smaller and less expensive to produce than each egg, but usually, many sperm cells are not carried to eggs; thus, a large number of sperm cells must be produced. If sperm delivery can be ensured, the number produced can be decreased, and even a small, dwarf male can produce adequate numbers. Delivery can be assured if the male gametophyte grows on the female as an epiphyte.
Several mechanisms compensate for the inability of mosses to retain water. Many species grow in permanently moist microhabitats such as rain forests, cloud forests, and the spray zones near waterfalls. Even habitats that are not constantly humid may have microhabitats that are continually moist: The foot of a rock cliff is usually damp because dew and mist concentrate there as runoff from the cliff. Shallow depressions in rock or soil retain moisture and are protected from drying winds. These sites are very small, but so are the mosses that live within them.
Other mosses are tolerant of desiccation. Drying does not damage them as it does most vascular plants and algae. Like lichens, many mosses can lose much of their water rather rapidly without dying or even being injured. As long as about 30% of their weight is water, they remain dormant but alive. If rain falls or dew forms, water is absorbed rapidly, and within a few minutes, respiration and photosynthesis are occurring at normal levels. In mild temperate regions, mosses may remain moist and metabolically active except for certain times during summer months. In deserts and dry range lands, on the other hand, mosses may be turgid primarily during winter. From spring to autumn, the mosses are dry and inactive every day, perhaps receiving enough dew on certain nights to be metabolically active for a few hours in the early morning.
Desiccated mosses are remarkably resistant to high or low temperature and to intense ultraviolet (UV) light. A common moss, Tortula ruralis, can be frozen safely in liquid nitrogen (—196°C) when dry and can also withstand brief periods at +100°C. Resistance to UV light is important for species that grow in full sun on the surface of rocks or at high altitudes on mountains, where the atmosphere is too thin to block UV radiation.
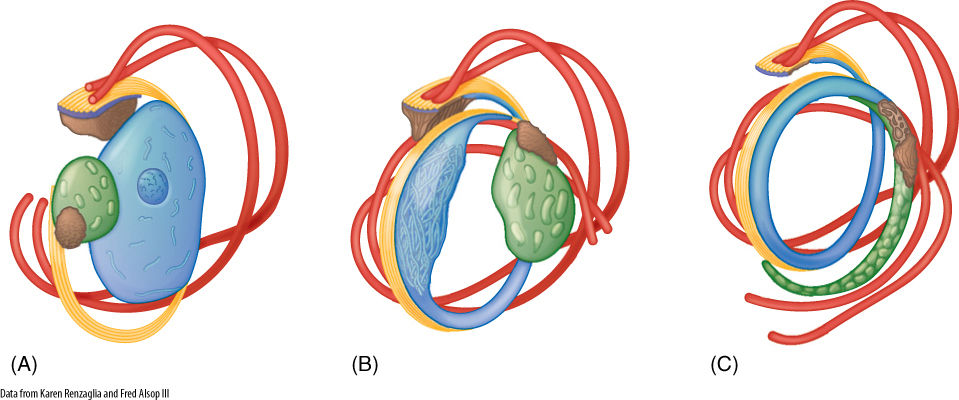
FIGURE 20-18 (A) The organelles of a young sperm cell have rather typical shapes (blue, nucleus; green, chloroplast; red, flagella and basal bodies; brown, mitochondrion; yellow, microtubules that anchor the flagella). (B and C) During differentiation, the organelles become modified, resulting in an elaborate spiral-shaped sperm cell at maturity (C). Cytosol and cell membrane are not shown.
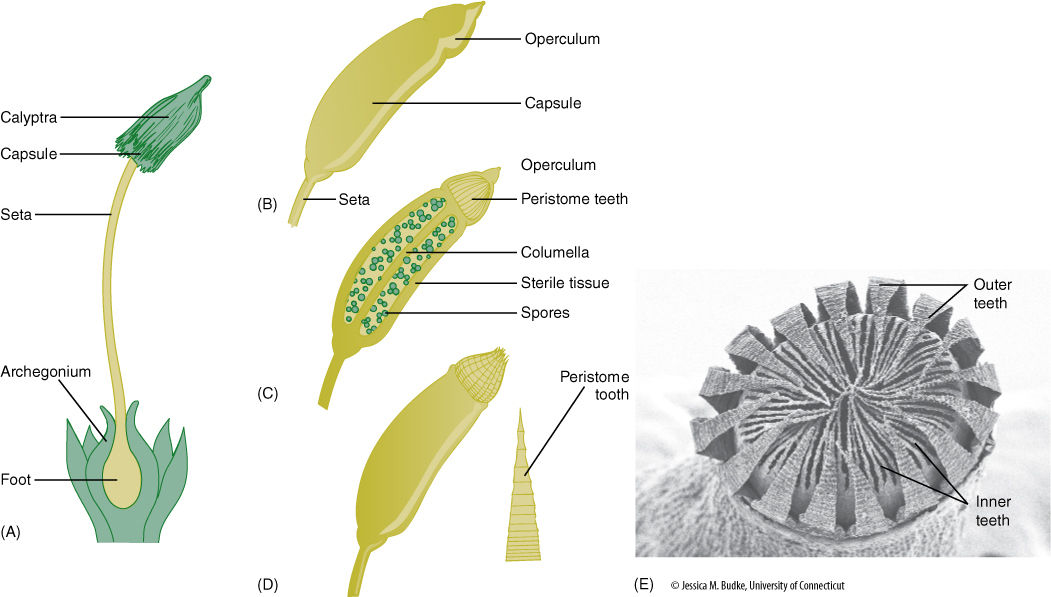
FIGURE 20-19 Aspects of a moss sporophyte. (A) The foot is embedded in the remnants of the archegonium. (B) External view of the capsule (sporangium) with the operculum still in place covering the teeth. (C) Median section view showing the columella and spores. (D) External view after the operculum has fallen off. Spores can be shed when the peristome teeth bend back. (E) Scanning electron micrograph view of peristome teeth.
Many moss species thrive at low temperatures near or even below 0°C. When larger vascular plants die or abscise their leaves in winter, much more sunlight reaches the soil surface or tree trunks where mosses are growing. Snow or winter rains provide abundant moisture. Mosses that can carry out metabolism at cold winter temperatures can take advantage of the plentiful light and moisture.
Certain mosses can grow on hard, impervious surfaces because they have no roots that must penetrate the substrate. Consequently, mosses can be found on bare rock (Andreaea, Hedwigia), brick or mortar (Tortula muralis), and bark as well as soil. At the other extreme, Sphagnum mosses live on the surface of quiet water at the edges of lakes and ponds (FIGURE 20-20). Their leaves and stems are buoyant, and thick mats build up.
In almost all environments, mosses are important in the later establishment of other species, particularly vascular plants. Like lichens, they can colonize a bare surface and dissolve the rock with acids that leak from their cells. Leaves and stems of gametophores catch and hold dust particles, creating small pockets of soil in which spores and seeds of vascular plants can become established. A similar process occurs even on the ground; in forests or grasslands, the moss layer on the soil may be thick enough to act as a moist, airy seed bed. It may hold water better than the soil and thus improve the microhabitat for seedlings.

FIGURE 20-20 Sphagnum moss grows at the edges of streams or ponds where there is little water movement. The plants become large and tangled, creating a mat that is solid enough to support the weight of an animal. These are known as quaking bogs because they quake and undulate with every step. Given enough time, seeds of vascular plants germinate, and leaves and debris blow onto the bog and decompose, slowly converting it to a rich soil.
![]() Division Hepatophyta: Liverworts
Division Hepatophyta: Liverworts
Like mosses, liverworts are small plants that have an alternation of heteromorphic generations (TABLE 20-5). The smallest individuals of Cephaloziella are only 150 by 2,000 mm. Few species have plants that ever become large, the maximum being approximately 5 by 20 cm in Monoclea. Some species are leafy and greatly resemble mosses (FIGURE 20-21A); others form small, solid, ribbon-like gametophytes (see Figure 20-8 and FIGURE 20-21B). The sporophyte is even less conspicuous than in mosses and is also completely dependent on the gametophyte.
The Gametophyte Generation
Hepatic gametophytes are divided into two basic groups: leafy liverworts (orders Jungermanniales and Haplomitriales) and thallose liverworts (Figures 20-21 and FIGURE 20-22). In both, the gametophyte phase is initiated when spores germinate and establish a small, temporary protonematal phase. Liverwort protonemata are never as extensive or ramified or as long lived as those of mosses. Instead, after only a few cells are produced, an apical cell is established and growth of the gametophore begins.
The gametophore of leafy liverworts greatly resembles that of a moss—thin leaves on a slender stem; however, liverwort leaves typically have two rounded lobes with no midrib and no conducting tissue, whereas moss leaves are pointed and usually have a midrib. Liverwort leaves are arranged in three clearly defined rows, the leaves of two rows being much larger than those of the third. In prostrate liverworts, in which the shoot grows flat against the substrate, the underside row of leaves is reduced and may even be completely suppressed or replaced by rhizoids.
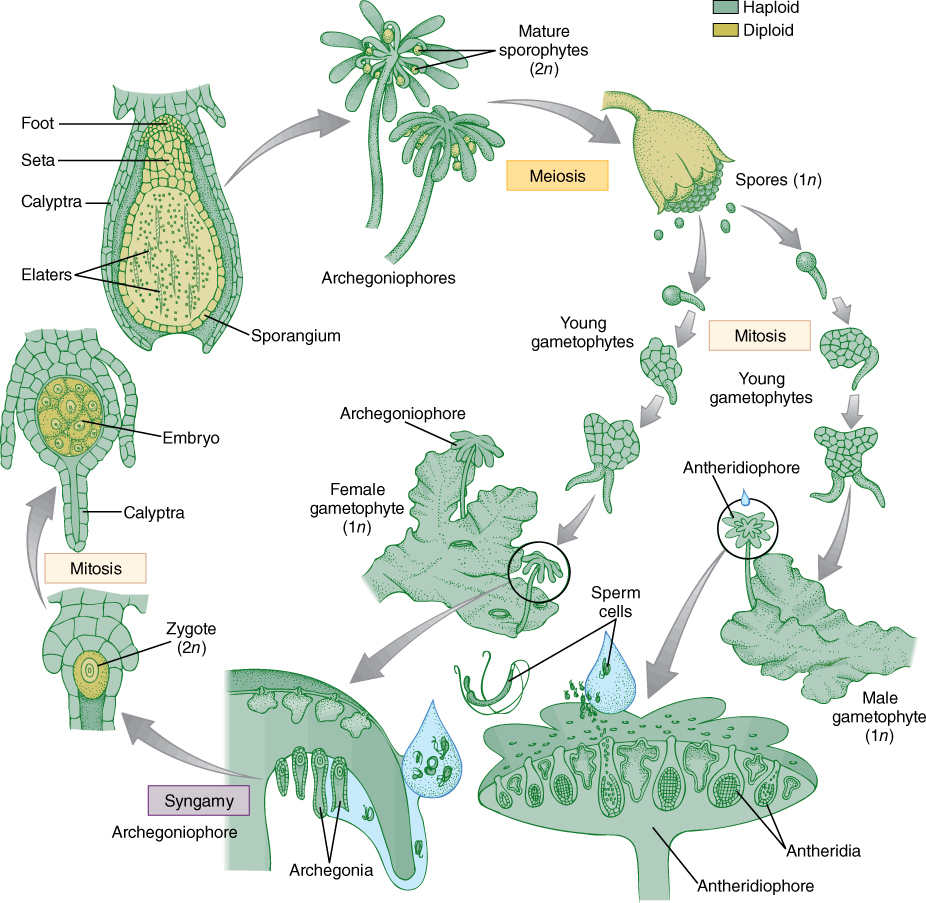
TABLE 20-5 Classification of Division Hepatophyta

FIGURE 20-21 (A) Leafy liverworts such as this Lophocolea can be very easily confused with mosses. Their distinguishing features (not visible at this low magnification) are their typically lobed leaves, which grow from two apical points, and their basal pouch. Moss leaves are never like this. (B) Thallose liverworts have rather thick bodies and are not easily confused with mosses. This is a moist, active Oxymitra incrassata; notice its papery scales. White dots are air pores; clefts contain gametangia. When young, the gametophyte had one apical meristem and one cleft (upper left), and then the apex divided dichotomously so two apices and two clefts are now present. Each can divide again. (C) In dry seasons, plants of Oxymitra desiccate and fold inward, bringing the scales over their body, protecting themselves. They survive months of severe heat and drought.
FIGURE 20-22 Life cycle of Marchantia. Details are given in text.
The gametophore stem grows by an apical cell with three, four, or five sides, one uppermost and the others embedded in the stem. Stem tissue is just simple parenchyma; no thick-walled cells or conducting cells occur in the majority of species. Virtually all liverworts have some or many cells that contain characteristic oil bodies.
Thallose liverworts show less resemblance to mosses. They are not leafy at all but rather flat and ribbon like or heart shaped and bilaterally symmetrical. The body is sometimes referred to as a thallus (plural, thalli), a body without roots, stems, and leaves. Currently, “body” is much more commonly used than “thallus.” Bodies of thallose liverworts are stratified and tend to be much thicker than those of leafy liverworts and mosses. The side next to the substrate bears unicellular rhizoids, and many cells contain large oil drops. Cells in the side away from the substrate have no oil but are rich in chlorophyll. The cells are loosely arranged as an aerenchyma with large air chambers that open to the exterior by means of large air pores (FIGURE 20-23). Air pores are not stomata—they have no guard cells and cannot be closed.
Some species are even simpler. Sphaerocarpos texanus is a small, thin ribbon a few cells thick at the center, but the rest of the body is only one cell thick. It has simple rhizoids on the bottom but no internal air chambers and no scales or other types of vegetative differentiation. These plants are basically as simple as the sea lettuce alga, Ulva, or the charophyte, Coleochaete. The liverworts Pallavicinia and Pellia are also this simple.
Liverwort gametophores may be either bisexual, producing both antheridia and archegonia, or unisexual, depending on the species. Leafy liverworts may bear their gametangia either mixed with regular leaves or positioned on specialized side branches and surrounded by modified leaves. In thallose species, antheridia and archegonia may be grouped together and surrounded by a tube of chlorophyllous cells. Marchantia is probably the most familiar thallose liverwort because it grows easily on moist soil in greenhouses. Its gametangial production is particularly elaborate. Male gametophores of Marchantia produce an umbrella-shaped outgrowth called an antheridiophore (FIGURE 20-24). It has a stalk several millimeters tall, and dozens of antheridia grow from its upper surface, each surrounded by a rim of sterile cells. Archegoniophores also are stalked, but their apex is a set of radiating fingers that project outward and droop downward; the underside has numerous archegonia (FIGURE 20-25). Archegoniophores and antheridiophores occur on separate plants in Marchantia; antheridia and archegonia are not produced by the same gametophyte.
If sperm cells are carried to the archegoniophore by raindrop splashing, they swim into the archegonium neck and fertilize the egg. The zygote is retained and grows into a small sporophyte (FIGURE 20-26). Surrounding gametophore tissue expands with it temporarily, forming a protective sheath, and the archegonial neck expands into a calyptra. In Marchantia, sporophytes are so small that each archegoniophore can support many of them.
The Sporophyte Generation
Little variability exists in the sporophytes of most liverworts, and their basic morphology is like that of mosses. In fundamental structure, however, the two groups are quite distinct. Most liverwort sporophytes have a foot, seta, and calyptra-covered sporangium, but the seta is extremely delicate, composed of clear, thin-walled cells that collapse quickly. Many members of Marchantiales have no seta at all. The sporangium is globose and while young is bright green and chlorophyllous, but it has no stomata. The outer layer of sterile cells is much thinner than in moss sporangia, often only one cell thick. The liverwort sporangium lacks a columella, the central mass of sterile cells found in mosses; dehiscence occurs by means of four longitudinal slits, not by an operculum, and there are no peristome teeth. Within the sporangium, some cells do not undergo meiosis but rather differentiate into elaters—single, elongate cells with spring-shaped walls. When the sporangium opens, the elaters uncoil, pushing the spores out (FIGURE 20-27). All liverworts are homosporous.
Strikingly simple sporophytes are produced in Riccia and Ricciocarpus. The zygote grows into a spherical mass within the archegonium of the gametophore body. No foot or seta is formed; instead, the inner cells of this mass undergo meiosis and produce spores. As the surrounding gametophore and sporophyte tissues age, die, and decay, the spores are finally liberated.
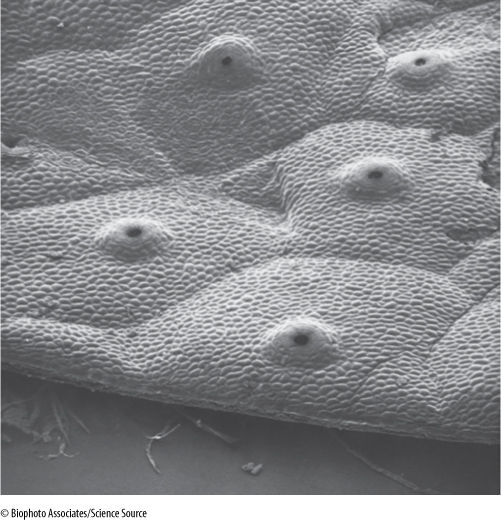
FIGURE 20-23 Although gametophytes of thallose liverworts may become very thick (a few millimeters), they are never solid; rather, they have numerous chambers, and both carbon dioxide and water vapor can diffuse through them easily (×425).
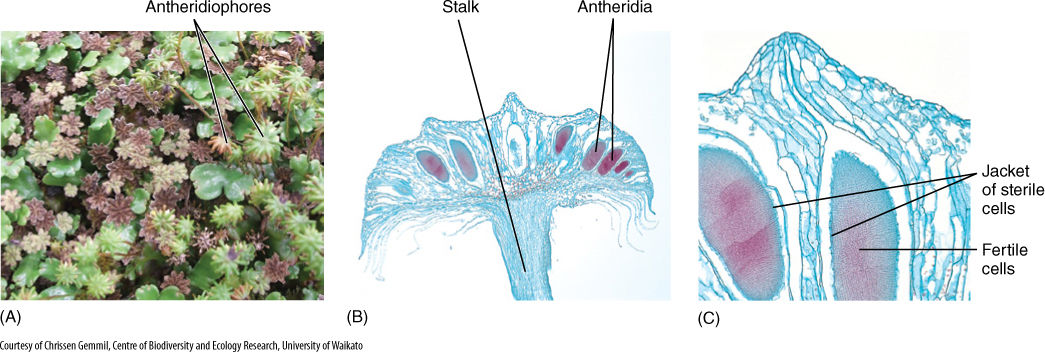
FIGURE 20-24 (A) These are antheridiophores of Marchantia; as their name indicates, these structures bear (-phore) antheridia. These are gametophytic tissues, and the antheridia are hidden within the upper surface (four archegoniophores are also present). (B) Low-magnification view of antheridiophore with antheridia (× 8). (C) Like antheridia in mosses and all other embryophytes, those in liverworts consist of a jacket of sterile cells surrounding fertile cells that differentiate into sperms (× 32).
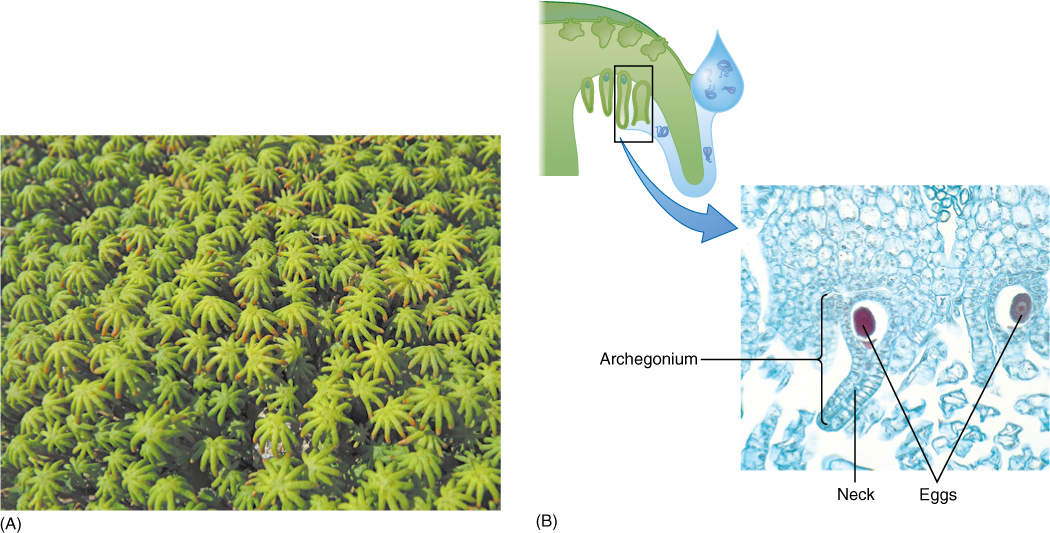
FIGURE 20-25 (A) Archegoniophores of Marchantia are easily recognizable because they have finger-like segments radiating from the stalk. Archegonia are located on the underside (× 3). (B) Each archegonium consists of a long, tubular neck and a slightly swollen base, all only one cell thick. Each archegonium contains only one egg (× 160).
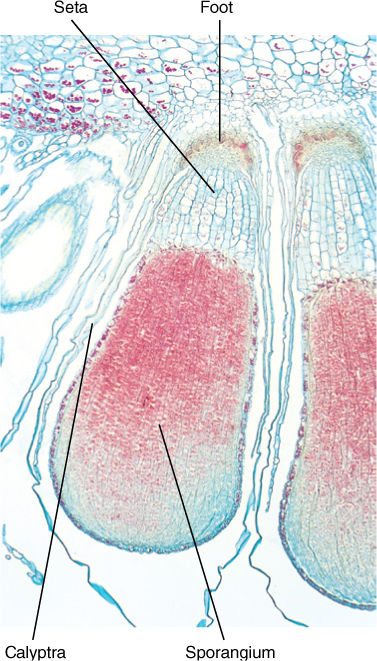
FIGURE 20-26 Liverwort sporophytes consist of foot, seta, and capsule (sporangium), but no elaborate set of teeth as in mosses. Instead, the apex breaks into several segments, all of which curl back and release the spores (× 20).
![]() Division Anthocerotophyta: Hornworts
Division Anthocerotophyta: Hornworts
Hornworts are a group of small, inconspicuous thalloid plants that grow on moist soil, hidden by grasses and other herbs (see Figures 20-6, 20-9, and 20-28 and TABLE 20-6). They rarely inhabit tree trunks or bare rock; therefore, one does not often encounter them unless looking for them specifically. Roadside cuts and stable eroded soil in shade are good sites for hornworts. Hornworts number approximately 100 to 150 named species in five or six genera, with Anthoceros and Phaeoceros being common examples. Many species names are probably synonyms—that is, several names applied to the same species because many hornworts have such variable growth forms that individuals of the same species can differ greatly under varied conditions.
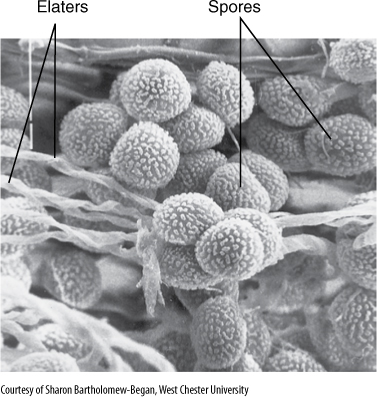
FIGURE 20-27 Spores and elaters of the liverwort Haplometrium hookeri.
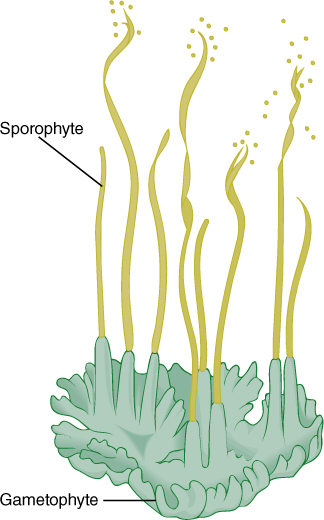
FIGURE 20-28 The “horns” of a hornwort are sporophytes that grow continuously from a basal meristem. The lower part of the sporophyte is surrounded by gametophyte tissue.
Hornworts superficially resemble thalloid liverworts, the gametophores of many species being ribbon shaped and thin, without a distinct stem or leaves; however, hornworts never contain oil bodies, whereas liverworts almost always do. As in all other embryophytes, an alternation of heteromorphic generations occurs, with the sporophyte depending on the larger, photosynthetically active gametophyte, but hornworts are quite distinct from all other embryophytes, including the other nonvascular plants. One of the most striking features is the presence of a single large chloroplast in each cell as opposed to the numerous small plastids present in all other nonalgal plants (FIGURE 20-29). The single chloroplast per cell is characteristic of many algae and is surprising in hornworts. Furthermore, hornwort chloroplasts have a pyrenoid, an algal feature absent in all other embryophytes. Hornwort chloroplasts probably represent the condition of the ancestors of all embryophytes. Unfortunately, we do not know what the chloroplasts of the earliest plants were like because they are not preserved in any known fossil.
TABLE 20-6 Classification of Division Anthocerotophyta

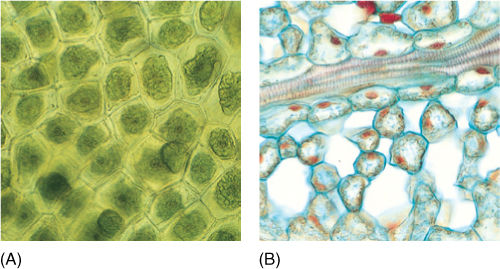
FIGURE 20-29 (A) Hornwort cells are unique among embryophytes: Each has just one plastid. Outside of hornworts, this feature is known only in algae. Although it may seem trivial, it must represent a significantly distinct cell/plastid relationship. The plastid’s reproduction must be carefully coordinated with that of the cell, and a special mechanism is necessary to ensure that both progeny cells receive one each (× 60). (B) Cells of privet leaf with many chloroplasts each (× 60).
The Gametophyte Generation
As few as three or four protonema cells are produced before the gametophore phase is established in most species. Gametophores are always thin, at least along the edges. Only in the center do they become more than four or five cells thick. They may be shaped like a ribbon or a heart, or they may grow outward irregularly, forming a disk. The upper surface in many is smooth, but in others thin, chlorophyllous lamellae grow upward. The gametophyte is parenchymatous, rather succulent but brittle. It does not tolerate drying; gametophores of hornworts typically live for less than 1 year in temperate climates. They act as winter annuals, appearing in the cool, moist autumn months, growing during winter, producing sporophytes in the spring, and dying before summer. In some species, they form oil-rich “tubers” as inner cells fill with oil and outer cells die; one thallus can produce several tubers.
Internally, hornwort gametophytes have numerous chambers. Young plants have mucilage chambers formed as cells break down and their contents are altered chemically into mucilage. These become invaded by Nostoc cyanobacteria. All hornworts form this symbiosis and presumably benefit by receiving nitrogen compounds from the Nostoc.
Gametangium development in hornworts is distinctive (FIGURE 20-30). Unlike all other embryophytes, the antheridial initials are not surface cells on the hornwort gametophyte. Instead, a special mucilage chamber forms near the upper surface; then cells lining the chamber grow into it and become antheridia. The chamber expands as the antheridia grow, and finally, its roof breaks; only then are antheridia exposed to the environment. As sperm cells mature, sterile outer cells of antheridia transform their chloroplasts into chromoplasts, becoming orange or yellow.
Archegonia are formed from superficial cells, but archegonia do not completely surround the egg as do the flask-shaped archegonia of other embryophytes. Rather, the egg lies below a short neck and neck canal but is surrounded by vegetative thallus cells. Even the neck cells are not particularly distinct but rather are difficult to distinguish from ordinary thallus parenchyma cells.
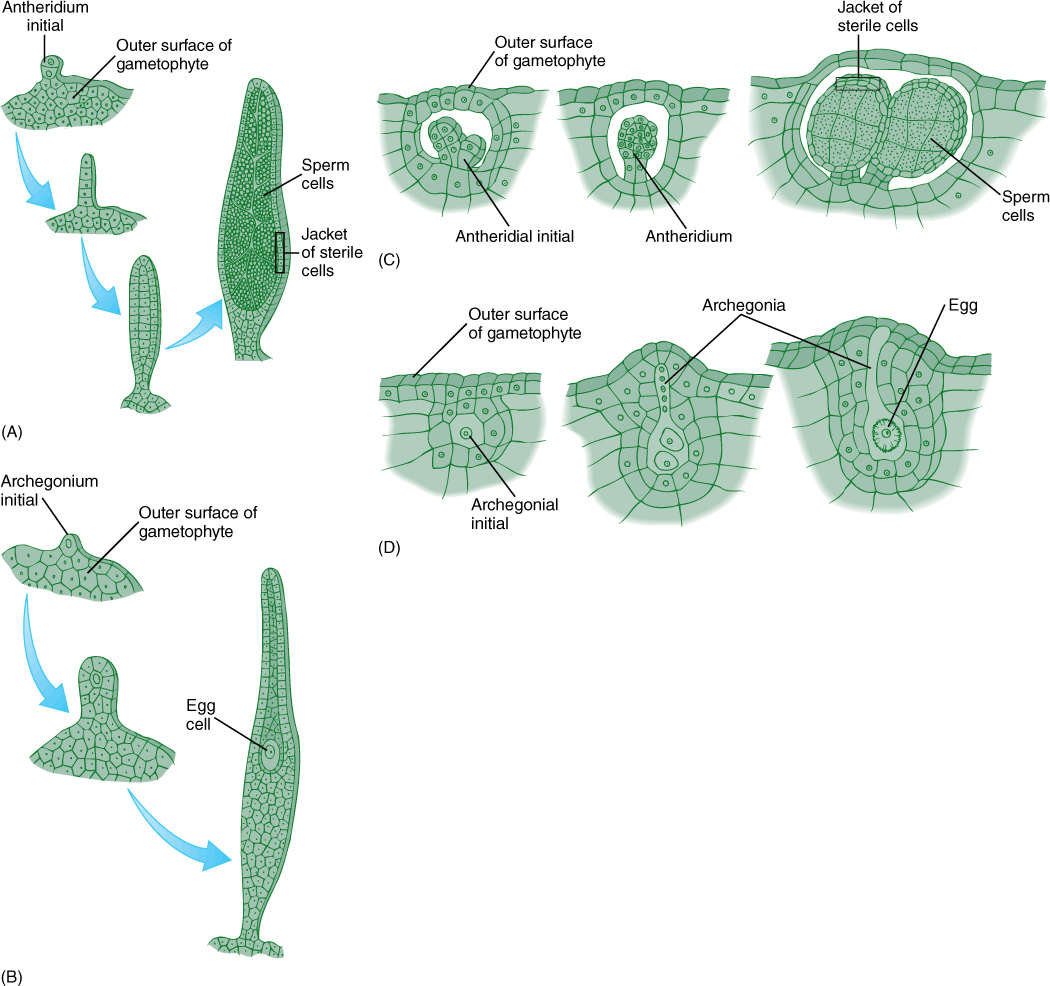
FIGURE 20-30 (A) and (B) In mosses, the initial cell in the formation of either an antheridium or an archegonium lies on the surface of the gametophyte. Then cell divisions change this single cell into a mass that has sterile jacket cells and reproductive central cells. This pattern occurs in all mosses, liverworts, and vascular plants. The gametophytes of flowering plants are so reduced that the pattern is not obvious. (C) and (D) The gametangia of hornworts are unique; the antheridia (C) do not form on the true surface of the gametophyte, and the egg (D) is not surrounded by discrete archegonial cells.
After fertilization occurs, the zygote divides longitudinally; in mosses and liverworts, it divides transversely.
The Sporophyte Generation
Similarities between sporophytes of hornworts and those of mosses or liverworts are not easy to find. Hornworts have a foot embedded in gametophore tissue, but there is no seta or discrete sporangium (see Figure 20-28, FIGURE 20-31, and FIGURE 20-32). Instead, just above the foot is a meristem that continuously produces new sporangium tissues. As the newly formed cells are pushed upward, they grow, differentiate, mature, and die. They are simultaneously being replaced by more cells from the basal meristem. Consequently, the sporangium is a long, horn-like cylinder, typically 1 or 2 cm long in Anthoceros and Phaeoceros, but up to 12 cm in some species. At the tip, the sporangium is mature and open as a result of dehiscence along two linear apertures. The outer layer of sterile cells is thick, up to six cells deep, and chlorophyllous. It has stomata in Anthoceros and Phaeoceros but lacks them in Notothylos, Dendroceros, and Megaceros. Spores are green, golden yellow, brown, or black and in some species are multicellular when ready to be released. Hornworts have a columella as in mosses, but unlike the elaters of liverworts, those of hornworts (often called pseudoelaters) are multicellular and do not have spirally thickened walls. The basal meristem is active over a long period, depending on moisture availability and temperature, and large numbers of spores can be produced by each sporophyte. Several attempts have been made to remove the sporophyte surgically from the gametophyte and grow it in laboratory conditions, but even though it is chlorophyllous, it dies. It is not known whether death is caused by a lack of minerals, insufficient photosynthesis, or absence of growth factors, vitamins, or hormones from the gametophyte.
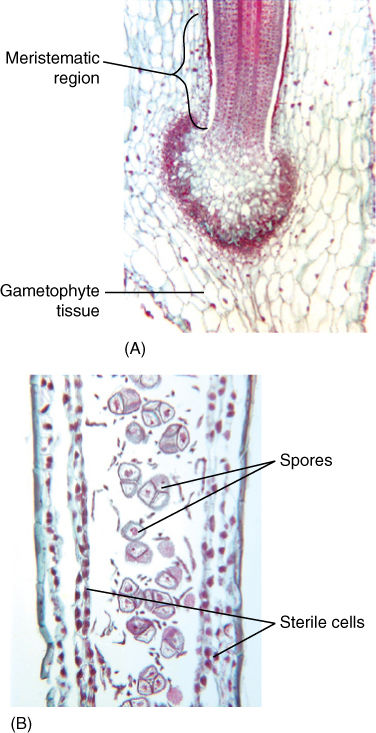
FIGURE 20-31 (A) The base of the sporophyte resembles a foot embedded in the gametophyte, and recently transfer cells have been discovered; therefore, active nutrient transport into the sporophyte must be occurring. Just above the foot is a meristematic region. (B) At a higher level, equivalent to Figure 20-32C, spores are mature.
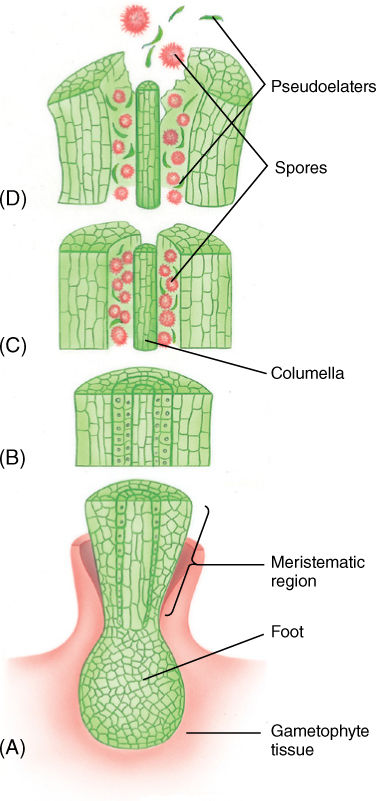
FIGURE 20-32 A longitudinal section through the sporophyte. At the base is a meristematic region (A); higher, above the basal meristem, sporocytes (spore mother cells) undergo meiosis (B). At higher levels the spores become mature (C) and are then released (D). This type of continuous meiosis is unknown in plants other than hornworts.
 At the Next Level
At the Next Level
1. Gametophytes as complex plants. Every angiosperm has two very simple gametophytes: the male gametophyte (pollen) and the female gametophyte (the embryo sac, having eight nuclei and seven cells, living inside an ovule). The gametophytes of bryophytes are much more complex, having leaves, stems, and rhizoids (but never having roots). Almost all plant biology is focused on sporophytes (trees, shrubs, etc.), but every plant without exception has a gametophyte stage. For independent study at the next level, I recommend reading about the gametophytes of every major group of plants, and the best place to start is in the following chapters of this book, plus Chapter 9 Flowers and Reproduction. After that, check online or in books on plant morphology. It is extremely difficult to study most gametophytes in person because, other than those of ferns, most gametophytes are small, live only briefly, and are rare at best. If you have a local expert than can help you find some in nature, that would be a great experience.
2. Bryophytes as wildflowers. I hope you pay some attention to the plants on campus or near your home; perhaps you know the names of some trees and cultivated landscaping plants, maybe some wildflowers. People often notice mosses but have a difficult time learning their names: Many wildflower identification books list only two or three mosses and often with photographs in which important details cannot be seen clearly. But there are several excellent guides to bryophytes written for beginners. I recommend the following:
1. California Mosses by Bill Malcolm, Nancy Malcolm, Jim Shevock, Dan Norris (2009, Micro-Optics Press)
2. A Trailside Guide to Mosses and Liverworts of the Cherokee National Forest by Paul Davison and Mark J. Pistrang (2008, Blurb)
3. Common Mosses of the Northeast and Appalachians (Princeton Field Guides) by Karl B. McKnight, Joseph R. Rohrer, Kirsten McKnight Ward, and Warren J. Perdrizet (2013, Princeton University Press).
Even if you do not live in the areas specifically named by these guides, most mosses and liverworts are so widespread that these books are valid for many states.
3. Asexual reproduction. Bryophytes excel at asexual reproduction. In many species, almost any bit or piece of gametophyte tissue that happens to break off and then lands in a suitable site will grow into a new plant. In addition, many bryophytes produce small clusters of cells called gemmae (singular: gemma; pronounced GEM ee and GEM uh) that are easily dislodged by rain, wind, or animals and then grow into new plants that are clones of the original. Other species produce branches whose leaves fall off and grow, or the branches themselves fragment and grow. In all cases, gemmae or fragments are so tiny and lightweight that they can be blown hundreds of miles, allowing the plant to be dispersed over a wide area. In general, bryophyte species are much more widespread than seed plant species. Gemmae and asexual reproduction are significant alternatives to seeds and sexual reproduction.
SUMMARY
1. True plants first originated at least 450 million years ago, having evolved from green algal ancestors. Necessary modifications would have included methods of water conservation: low surface-to-volume ratio, compact body, cuticle, and coordination of reproduction with presence of water.
2. The presence of plants on land changed the land as a habitat; with crowding, competition for light would have become important and an upright habit selectively advantageous.
3. The evolution of vascular tissue had profound consequences: Relatively strong, vertical stems could be supported. Large plant bodies could be integrated, and parts could differentiate and specialize.
4. Nonvascular plants are classified here in three divisions, emphasizing their differences and possible separate evolutionary origins: Bryophyta (mosses), Hepatophyta (liverworts), and Anthocerotophyta (hornworts).
5. All three groups of nonvascular plants are embryophytes. They have gametangia and sporangia in which only the internal cells differentiate into gametes or spores; a jacket of sterile cells is always present.
6. In all three groups of vascular plants, the life cycle consists of an alternation of heteromorphic generations. The gametophyte is the larger and more persistent and photosynthetically active phase; the sporophyte depends almost entirely on the gametophyte for carbon, energy, and minerals.
7. The setae, stems, and leaf midribs of some mosses have cells that facilitate long-distance conduction: Leptoids transport sugars, and hydroids transport water. It is not known whether they are homologous or merely analogous to xylem and phloem.
8. Archegonia and antheridia may be borne on the same gametophore or on distinct, unisexual gametophores.
9. Neither the egg nor the zygote is released. The new sporophyte develops initially within the archegonium. When fully mature, it has a foot and sporangium; many have a seta.
10. A moss spore grows into a filamentous protonema that produces buds that develop into thick parenchymatous gametophores with stems, leaves, and rhizoids.
11. Liverworts may be either thallose or leafy. Liverworts contain oil droplets, have bilobed leaves, and have no thick leaf midrib, no columella in the sporangium, and no operculum or peristome teeth.
12. Hornwort sporophytes differ from those of all other plants: They have a foot and a basal meristem, and thus, sporogenous tissue is formed continuously.
13. Hornwort antheridia do not arise from surface cells, and the egg is not completely surrounded by distinct archegonial tissue.
IMPORTANT TERMS
air pores
antheridiophore
antheridium (plural: antheridia)
archegoniophores
archegonium (plural: archegonia)
calyptra
capsule
elaters
foot
gametophores
hornworts
liverworts
mosses
operculum
peristome teeth
protonema
rhizoids
seta
thallus
REVIEW QUESTIONS
1. Some vascular plants produce seeds; others do not. Vascular plants that do not produce seeds are known as ____________________, whereas vascular plants that do produce seeds are known as ____________________. Are there any plants that produce seeds but which do not have vascular tissue?
2. What are some of the modifications necessary if an alga is to become evolutionarily adapted to living on land? Is a single modification sufficient, or are several necessary?
3. Why would it be necessary for an evolutionary line to develop stomata and guard cells before it developed an extremely impervious cuticle? Why must vascular tissues precede the evolution of roots and active apical meristems?
4. The following organisms are often called mosses, but they are not actually closely related to mosses at all. What groups of plants do they actually belong to?
a. Spanish moss
b. Club mosses
c. Slimy, bright green “mosses” of ponds and slow-moving streams
d. “Reindeer moss”
5. What are the three groups of nonvascular plants? How would you determine whether an unknown specimen is a vascular plant?
6. The nonvascular plants of this chapter are believed to be true plants, just as ferns, conifers, and flowering plants are true plants; however, there are two tissues that the nonvascular plants do not have. Which two tissues?
7. If the leptoids of mosses were found to contain a protein whose gene had the same nucleotide sequence as the gene that codes for P-protein, would that be significant evidence for either the homology or analogy of leptoids and phloem?
8. You will see sporophytes only if you examine mosses closely. They look like green or brown “_____________” standing up on the green gametophyte, but sporophytes are (circle one: present almost all the time, only present at certain times of the year).
9. Do mosses have an alternation of isomorphic or heteromorphic generations? That is, can you easily tell a moss gametophyte from a moss sporophyte? When we look at leafy green moss plants, what are we seeing—the gametophyte or the sporophyte? In a flowering plant species, would the equivalent stage be the plant or the pollen grains and megagametophytes?
10. The leafy, green moss plants that are so familiar are gametophytes, haploid plants. This is very different from flowering plants and other seed plants. Does a leafy green moss plant grow from a spore or from a fertilized egg? Does the moss plant have both a paternal parent and a maternal parent?
11. Draw a single moss plant, similar to the one in Figure 20-10. Be certain to show the gametophyte and the sporophyte. Now draw one without the sporophyte, showing only the gametophyte. The sporophytes usually have only a very brief life, and after they shed their spores, the gametophytes let them die.
12. Draw and label the life cycle of a moss; be certain to show gametangia and sporangia. Which parts are haploid and which are diploid? Where and when does meiosis occur? Plasmogamy? Karyogamy?
13. In the majority of mosses, which lack hydroids and leptoids, water is conducted along the _____________ of the plant by _____________ action.
14. The leafy, green moss plants, being gametophytes, have gametangia, structures that produce gametes. What is the name of the gametangium that produces sperm cells? The gametangium that produces egg cells? Can one single moss gametophyte bear both of these? Do some species have plants that produce only sperm cells? Other plants that produce only egg cells?
15. The sporophyte of a moss usually has a stalk called a ____________ and a simple apical sporangium called a ____________.
16. Many people often think of mosses as plants adapted to rainy areas, areas that are usually wet. Are any mosses adapted to deserts? Can some mosses lose much of their water—the way a seed does before being planted—and still survive?
17. The liverwort Marchantia is one of the largest and most common. There is a good chance that you will find it if you search carefully in moist places (you may have to search many places). Is it a leafy liverwort or a thallose one? Describe its surface texture (see Figure 20-8). If you are very lucky, you may find it with archegoniophores and antheridiophores. What are these structures, and how would you recognize them if you saw them (what do they look like)?
18. Unlike Marchantia, some liverworts are as simple as a true plant can possibly get. Describe the body of Sphaerocarpos texanus. If you were shown one of these plants, how would you be certain it was not an alga (Hint: It would be almost impossible; there is only one way, not mentioned in the text. Algae tend to have only one chloroplast per cell; true plants—including S. texanus—always have many, except in the hornworts)? The point of this question is to have you think about how little difference there is between some algae and some true plants.
19. What are some of the ways in which liverworts differ from mosses? How do hornworts differ from both? Do the three have similar life cycles?
20. What are the “horns” of hornworts? What do they produce? They have a meristem. Where is it located?
21. An important consideration in the evolution of any organism is gene flow. What are some of the mechanisms by which genes move through the habitat in nonvascular plants? In a dense, cool forest, how strong are wind currents? Could they carry spores very far? What would you guess might be the maximum distance sperms can swim? How far can a raindrop splash a sperm or a spore?
Design Credits: Hummingbird: © Tongho58/Moment/Getty; Green plant cells: © ShutterStock, Inc./Nataliya Hora; Purple tulip: © ShutterStock, Inc./Marie C Fields; Dandelion: © ShutterStock, Inc./danielkreissl; Poppy: © ShutterStock, Inc./Saruri; Plant icon: © ShutterStock, Inc./Vector; Digging man icon: © ShutterStock, Inc./Z-art