Botany: An Introduction to Plant Biology - Mauseth, James D. 2017
Genetics and Evolution
Seed Plants II: Angiosperms

Chapter Opener Image: The poppy flower (Argemone) shows two important ways in which angiosperms differ from other seed plants. First, the pollen-producing organs (anthers, yellow) and the ovule-producing ones (carpels, red) occur together in most flowers whereas they always occur as separate cones in gymnosperms. Second, carpels are “closed,” which means they have no opening that allows pollen to just fall into the carpels and onto the ovules; instead the pollen must germinate and send a pollen tube through the stigma and style, carrying the two sperm cells downward to the ovule and egg. This results in a prolonged period of contact during which the stigma and style might reject the pollen tube if it is not the proper type. In many angiosperm species, the carpels exert considerable control over which pollen tubes are allowed to grow and which are not.
OUTLINE
✵ Concepts
✵ Changing Concepts About Early Angiosperms
✵ Classification of Flowering Plants
✵ Basal Angiosperms
✵ Monocots
- Commelinoid Monocots
✵ Eudicots
- Basal Eudicots
- Rosid Clade
- Asterid Clade
Box 23-1 Plants and People: Maintaining Genetic Diversity
Box 23-2 Plants and People: Vegetarianism: Alternatives to Eating Meat
LEARNING OBJECTIVES
After reading this chapter, students will be able to:
✵ List several derived characters of angiosperms.
✵ Explain the changes that occurred in the transition from gymnosperms to angiosperms.
✵ Compare monocots and eudicots.
✵ Name the several groups that are extant descendants of the basal angiosperm clades.
✵ Describe the features of monocots.
✵ Give 10 familiar examples of monocots.
✵ Identify three distinctive features of eudicots.
✵ Discuss the evolution of eudicots into basal eudicots, rosids, and asterids.
 Did You Know?
Did You Know?
✵ There are more species of flowering plants than all other groups of plants combined.
✵ Some flowers, such as those of cattails, maples, and oaks, are so tiny and inconspicuous you might not realize they are flowers.
✵ The largest flower in the world, Rafflesia arnoldii, is about 1 m in diameter and is produced by a parasitic plant.
✵ All angiosperms produce seeds, but orchids produce very tiny, underdeveloped dust-like seeds that are almost microscopic; the embryo is just a few dozen cells or less.
![]() Concepts
Concepts
Of all the clades that have arisen since embryophytes became distinct from charophyte green algae, the flowering plant clade—the angiosperms—contains the greatest number living species—257,000. These are all classified together in a single division, the Magnoliophyta (often called the Angiospermophyta). It is common to refer to angiosperms as the most advanced group of plants, or the most derived; occasionally, they are called the peak of plant evolution, but these statements should be examined carefully. Certainly the group as a whole has a greater number of derived features than any other group, and especially important is the large number of derived characters that are unique to angiosperms and that distinguish them from other plant clades. The group is named for having flowers—a significant synapomorphy that at once unites all angiosperms and separates them from all other clades. Other important derived characters are numerous methods of mutually beneficial interactions with animals that result in the plants being pollinated or having their seeds distributed while the animals received nectar or pollen or some other food. At the same time, angiosperms and animals have become more adept at fighting each other: As angiosperms evolved to provide reward for pollinators, natural selection also was giving them numerous new types of antiherbivore compounds, ranging from outright poisons (poison ivy, poison sumac, and deadly nightshade) to milder chemicals that deter feeding (chili peppers, mustard oil, and onion oils). Wood of almost all angiosperms is more complex than that of conifers: It contains vessels, which provide a low-friction pathway of water conduction, and it contains axial wood parenchyma (parenchyma cells mixed among the vessels and tracheids), which makes wood a living tissue that protects itself from wood-boring insects and fungi and can influence the distribution of water as it is pulled upward through the plant.
Do these features, however, make angiosperms the most advanced of all groups? Keep in mind that the simple spore-based reproduction of ferns has allowed them to reproduce successfully, spread into many habitats, and diversify into numerous sizes and shapes. Conifers also are extremely successful; one clade in particular, Pinaceae, is a relative newcomer, having arisen within the conifer clade after the angiosperm clade had already made quite a few advancements. Pinaceae is diversifying and speciating rapidly, producing not only many new species but also new genera.
The point is that we must not think about evolution as a linear progression forward and upward, with each new group leaving the previous ones behind. Instead, think in terms of real plants and populations. Some population of a species encounters an isolation barrier that causes it to diversify until at some point some of its members cannot reproduce sexually with others, and at that point, two very similar but distinct species exist. Both are free to continue evolving. After the early embryophytes became distinct from charophytes, they diverged into two clades, the zosterophyllophyte/lycophyll clade and the rhyniophyte/euphyll clade: Both were extremely successful, undergoing large numbers of modifications and diversifying greatly. After the early ferns originated, they too continued to diversify, with most variation occurring in habit, leaf size and shape, type of sporangium, type of indusium, and type of sporophyll. None evolved to have wood, flowers, or fruits, but they are not more primitive than the angiosperms; instead, different features have evolved in the two groups.
The earliest fossils clearly recognizable as parts of flowering plants are preserved pollen grains in rocks more than 130 million years old. We have not found any fossilized leaves or flowers from that time, and thus, we believe the earliest flowering plants must have lived in hilly or drier areas, areas where few sites were suitable for fossilizing plant parts. Large calm lakes with fine-grained sediments are best for fossilizing plants, and these are rare in dry areas or rocky hillsides. The first angiosperms probably were small trees or shrubs because plants of this type survive well in drier regions and because the most likely ancestors—other seed plants such as seed ferns or gymnosperms—were woody trees. As discussed earlier, the term “gymnosperm” may refer to an unnatural group; it is being used in this chapter to mean “all the seed plants except angiosperms.”
The evolutionary transformation of a gymnosperm into a flowering plant was not a simple process, and it involved many alterations. The most obvious was the conversion of gymnospermous sporophylls into stamens and carpels, resulting in the formation of flowers. Because many gymnosperms have rather flat, leaflike sporophylls, arranged in spirals around an axis, and microsporophylls never occur with megasporophylls except in cycadeoids, those flowering plants that have similar reproductive parts are believed to be the most relictual. For example, in a group known as the basal angiosperms, such as Amborella, Nymphaeales (water lilies), Austrobaileyales, and Magnoliales (Magnolia) have flat stamens without distinct filament and anther portions, arranged in spirals, and their sporogenous tissues (microspore mother cells) form relatively large, prominent internal masses (FIGURE 23-1).
In most angiosperm carpels, the edges of sporophyll primordia crowd against each other and grow shut, sometimes leaving a visible suture, sometimes closing so completely that no sign of a seam remains. This is called a closed carpel, and it develops into a fruit that encloses the embryos as they develop into seeds (“angio” means “clothed”). Carpels in the basal angiosperms are leaflike, resembling young leaves whose blades have not yet opened. Instead of a stigma and style, the ovary edges have rows of secretory hairs that secrete a thick liquid that both seals the seam and functions as a stigmatic surface (FIGURE 23-2). Whereas ovules in gymnosperms are exposed on the megasporophyll surface, the folded nature of the megasporophyll in flowering plants offers ovules some protection, and probably much more important is the fact that the pollen tube (microgametophyte) must now interact with the stigmatic area in addition to the nucellus, allowing more opportunity for the sporophyte to “test” the pollen. Weak pollen or that of the wrong species can be inhibited from germinating, and the sporophyte permits only healthy pollen of the proper species to pass through the stigma and reach its ovules (FIGURE 23-3). Fewer ovules are wasted by exposure to improper or unhealthy pollen.
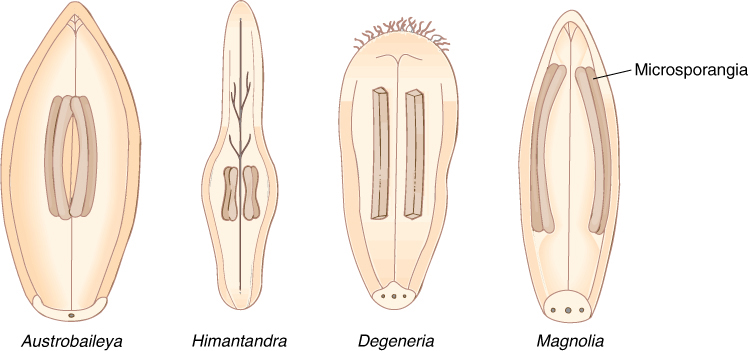
FIGURE 23-1 Several groups of angiosperms have stamens with what are believed to be relictual features; these features have not undergone much evolutionary change, so they are still similar to the ancestral features. The stamens (microsporophylls) of these four living angiosperms are much fleshier, flatter, and more leaflike than most other stamens of living species. Also, the sporogenous tissue is rather massive, similar to that of cycads and cycadeoids. Austrobaileya, Himantandra, and Degeneria are unfamiliar to most students; they are plants of South Pacific islands.
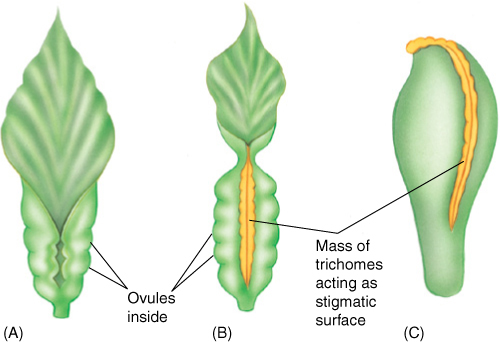
FIGURE 23-2 Carpels are megasporophylls; therefore, we expect the most relictual living carpels to be rather leaflike, somewhat resembling megasporophylls of cycads. Several living angiosperms do have carpels resembling young leaves that have failed to open (A); the two halves of the blade are pressed together, a rather large amount of vascular tissue is present, and a stigma and style are absent. Hairs along the blade margin act like a stigma. If such a carpel produced ovules only at the base and if the upper part elongated, it would begin to resemble a more derived, modern carpel (C).
In the transition from their ancestral group to being angiosperms, fertilization evolved such that the second sperm cell of the pollen tube fuses with the polar nuclei of the megagametophyte, producing the endosperm nucleus. This process of double fertilization is universal in flowering plants. Within the vegetative body, the major transitions were the evolution of vessel elements and sieve tubes. Leaves became broader and developed reticulate venation, and they became more polymorphic, adaptable to a variety of functions, not just photosynthesis.
Structural modifications are most easily seen and studied, but probably a more fundamental transition was the acquisition of developmental plasticity, the capacity to survive mutations that alter growth and development. Conifers lack developmental plasticity; for example, most people never have trouble recognizing an unfamiliar plant as being a conifer when they see one because all conifers greatly resemble each other. They are all large or giant trees with one or a few main stems and needle- or scale-shaped leaves. They are always green or brown, never brightly colored except for the red seeds of yew and the gray-blue berry-like cones of junipers. This uniformity is even more striking when one considers that gymnosperms dominated Earth for hundreds of millions of years and could have diversified into almost any habitat. On the other hand, angiosperms have tremendous developmental plasticity: They have diversified so greatly that not only hundreds of thousands of species, but also thousands of types, now exist (FIGURE 23-4). They range from gigantic (Eucalyptus, oaks, and elms) to tiny (mustards, certain daisies; Figure 23-4A); woody to herbaceous; perennial to biennial to annual; temperate to tropical; desert habitats to mesic to rain forest to aquatic; autotrophic to partially parasitic to fully parasitic; and epiphytic to subterranean (Figure 23-4B) to endophytic (living inside another plant, such as Rafflesia and Tristerix).
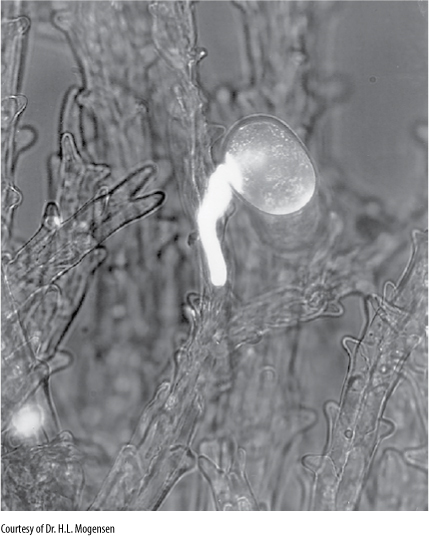
FIGURE 23-3 In many angiosperms, complex compatibility interactions occur between either the pollen and stigma or the pollen tube and stigma/style. In some cases, if the stigma and style detect an improper pollen tube, they deposit callose around it and inhibit its growth. The callose block is visible here as a white sheath (×60).

FIGURE 23-4 (A) This dwarf daisy (Monoptilon) is by no means the smallest flowering plant. Many species of mustard, often the first flowers of spring, are also tiny. (B) Ombrophytum subterraneum is a parasite on roots of surrounding plants. The plant consists of an irregular, lumpy “tuber” and two inflorescences with tiny pink flowers. The root and stem base of the host are visible. (C) Many angiosperms, such as this Hydrilla, have adapted to living submerged in water, where they need little sclerenchyma and have no transpiration. (D) All the fruits and vegetables in this market were harvested from angiosperms. Almost all the plants we eat are angiosperms, and, in addition, most of our domesticated animals eat angiosperms such as hay, oats, and corn.
The evolutionary changes involved in the conversion of a gymnospermous line into an angiosperm line did not occur instantaneously, nor did they occur in all species. Double fertilization was probably the first transformation because it is universally present. It also occurs in some gnetophytes, and we are not certain whether it is a shared ancestral feature (a symplesiomorphy) indicating that flowering plants and gnetophytes belong to the same clade or whether it is independent, convergent evolution (homoplasy) in two unrelated groups. The ability to produce bisexual flowers must also have been one of the first transformations because all flowering plant clades—including basal angiosperms—have megasporophylls located above microsporophylls on a single axis. Although many angiosperms species produce either staminate or carpellate flowers, we are certain that in every case this has been a loss of parts from a bisexual ancestral flower. Vessel elements possibly evolved next because only one or two basal species lack them (Amborella, Drimys). This has been a conundrum for many years: Quite a few plants were considered to be basal angiosperms on the basis of having wood without vessels; it was thought that these were primitively vesselless (that they lacked vessels because their ancestors lacked them). Many other features, however, were inconsistent with the idea of these being a natural, early group, and the current hypothesis is that perhaps only Amborella is primitively vesselless and that the others arose after vessels had originated but then these groups lost them. If so, they are secondarily vesselless, and their tracheid-based, gymnosperm-like wood is a derived feature that looks like a primitive one and misled us for years. Sieve tubes may have originated next; several species still have sieve cells in their phloem. All gymnosperms and seed ferns are or were woody plants and so are most of the basal angiosperms and eudicots; furthermore, wood in basal angiosperms has many gymnosperm-like features; therefore, we are confident that ancestral flowering plants were woody perennials. Many extant angiosperms are herbs, as are water lilies and peppers in the basal angiosperms; however, we are certain that this has resulted from the loss of the vascular cambium in these lines, and we are certain this has occurred repeatedly in many clades. The annual habit—the ability to germinate, grow, reproduce, and then die all within a single year—is a uniquely angiosperm feature; no other group of seed plants can do this. Being an annual requires that plants produce flowers shortly after germination, during a period when most gymnosperm plants would still be producing a seedling body with juvenile features. Seedlings and adults often differ strongly in their body characters, and this ability of plants only a few months old to flower may be an important factor in angiosperms’ developmental plasticity: Just because a plant has flowered does not mean that it has to die. After seedlings developed the capacity to flower, they could continue to live and grow large, retaining seedling features rather than developing adult ones.
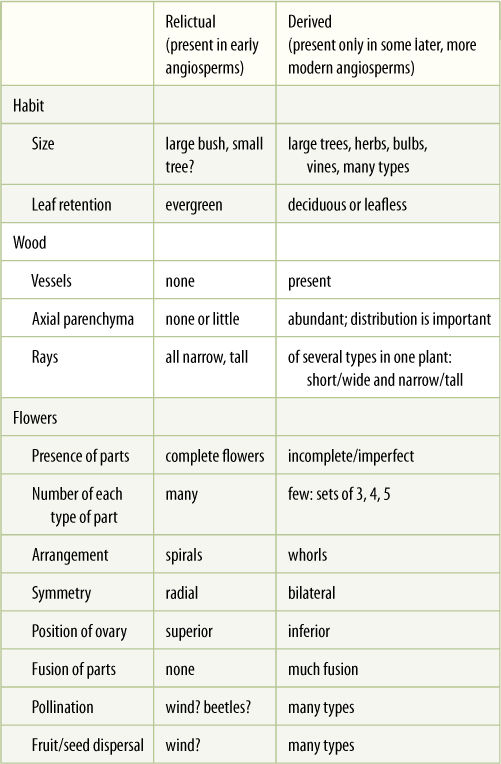
Other derived features are the fusion of the carpels into a single structure (a pistil), fusion of petals into one structure (sympetally), and floral zygomorphy, that is, flowers that are bilaterally symmetrical, not radially symmetrical (TABLE 23-1). Even more specialized characters such as succulence, parasitism, epiphytism, bulbs, corms, tubers, tendrils, and insect-trapping leaves originated only much later, after angiosperms had diversified into many clades; consequently, each of these innovations is present in only a few families or species. It should not be thought that the flowering plants are now “finished,” that all their evolution has already happened. Some groups do seem to be changing very little now, but others, especially grasses, composites, bromeliads, and orchids, are still changing and evolving so rapidly that it is difficult to keep up with them. Similarly, we have no reason to expect that the flowering plants will always be the most derived group; lycophytes dominated Earth for millions of years, only to be displaced by gymnospermous seed plants, which in turn are now being overshadowed by flowering plants.
![]() Changing Concepts About Early Angiosperms
Changing Concepts About Early Angiosperms
Concepts about the nature of early angiosperms have changed as our knowledge of existing and fossil plants has become more complete. In the last century, wind-pollinated trees (alders, elms, oaks, and plane trees) were grouped together in a “subclass Hamamelidae” and were considered the most relictual living flowering plants (FIGURE 23-5A). This seemed reasonable because these species tend to be large trees with dense wood, their flowers are small and simple, usually without sepals and petals, and many gymnosperms have similar features. After hamamelid wood anatomy was studied, however, it became obvious that they could not be relictual because their wood contains vessels, fibers, and abundant parenchyma, features not found in gymnosperms. Furthermore, hamamelid flowers are simple through specialization and reduction: Wind-pollinated flowers do not need to attract pollinators, and thus, they do not need to be large or colorful—they do not need petals or sepals. Recent DNA studies indicate that this syndrome of large, wind-pollinated trees is a derived condition within angiosperms, and furthermore, it has evolved several times in various clades: This is an example of homoplasy, of convergent evolution, and the hamamelid taxa are now separated from each other and placed among the clades we believe to be their true relatives.
TABLE 23-1 Relictual and Derived Features in Division Magnoliophyta
Approximately 100 years ago, C. E. Bessey developed the hypothesis of the ranalean flower, in which a Magnolia-type flower was thought to be relictual (FIGURE 23-5B). Such a flower is generalized; that is, it has all parts (sepals, petals, stamens, and carpels), and these are arranged spirally. Also, carpels occur in a superior position, above the other parts. It is easy to postulate the evolution of all of the various existing flower types from a ranalean, generalized ancestor. For example, monocot flowers often have three stamens and three carpels, whereas eudicot flowers usually have four or five of each; either condition could arise by a simple reduction of the large number present in a ranalean type flower. Also, the staminate or carpellate imperfect flowers of dioecious species could also arise easily by loss of parts.

FIGURE 23-5 (A) Catkins of pecan (Carya illinoensis); the axis bears many flowers, all with stamens but not carpels. Other flowers on the same plant would be carpellate and give rise to pecan fruits after being pollinated. (B) A flower of Magnolia is considered the typical ranalean flower. All parts are rather massive and are not specialized for one particular type of pollinator. Very few flowers have so many carpels, almost always fewer than 10; in contrast, seed cones of conifers and cycads have large numbers of megasporophylls. Each carpel secretes a drop of liquid that catches pollen grains, much as conifer megasporangia do.
Most botanists had long ago concluded that angiosperms are monophyletic. Features as complex as double fertilization, flowers, and developmental plasticity probably did not evolve more than once. Almost all recent DNA studies also indicate monophyly of angiosperms.
Currently, many paleobotanists and taxonomists believe that the transition from gymnosperm to angiosperm occurred during the Jurassic and Lower Cretaceous Periods of the Mesozoic Era. The earliest leaf fossils definitely considered to be those of angiosperms are from the Lower Cretaceous Period, approximately 130 million years ago. They represent both dicot and monocot leaves, for example Magnoliaephyllum, Ficophyllum (fig-like leaves), Vitiphyllum (grape-like leaves), and Plantaginopsis (similar to Plantago) (FIGURE 23-6). These fragments are rare in such old rocks, constituting only approximately 2% of all fossils present. In rocks about 50 million years more recent, from the Upper Cretaceous Period, many more types of angiosperm leaf fossils are present, outnumbering the leaf fossils of gymnosperms and ferns. These Upper Cretaceous leaf fossils originally were assigned to extant genera such as Populus (poplar), Quercus (oak), and Magnolia on the basis of overall shape and size, but careful examinations of fine venation, cuticle, and stomata often show that these fossils are not truly part of modern genera.
The original misidentification of these leaves contributed to the erroneous hypothesis that angiosperms must have originated in the Jurassic Period, become abundant enough to leave a few fossils in the Lower Cretaceous Period, but then suddenly, in the short span of 30 to 50 million years, rapidly diversified into most of the major extant families and even many of the modern genera. After this supposed burst, genera such as Populus, Quercus, and Magnolia would have had to remain relatively unchanged for tens of million years, up to the present. Such a burst of diversification followed by extreme stability did not seem plausible. With the discovery that the Upper Cretaceous and Tertiary leaf fossils only resemble leaves of modern genera but are not identical to them in subtle features, a much more gradual evolution can be assumed to have occurred. The early angiosperms were well established by the Lower and Mid Cretaceous Periods, but probably no modern genus was yet in existence, only precursors that greatly resembled modern genera. These would have evolved gradually into true poplars, oaks, magnolias, and so on.
The oldest wood that seems to be from an angiosperm comes from the Aptian Epoch (125 million years ago) of Japan; unequivocal angiosperm wood occurred approximately 120 million years ago. Flowers and fruits appear for the first time in the Lower Cretaceous Period, which must mean that their forerunners evolved in the Jurassic Period (FIGURE 23-7), about the same time that dinosaurs were flourishing (see the timeline on the inside cover on the back of the book). In the last several years, China has explored for fossils much more vigorously, and rich new fossil beds are yielding many new specimens of early angiosperms. Often the fossils consist only of abscised carpels or carpels attached to a receptacle. Unfortunately, sepals, petals, and stamens often abscise even while a flower is still on a plant, and as a result, they are rarely present with the carpels. Perianth parts and stamens of modern plants also tend to be delicate and to decay quickly rather than fossilize; the same may have been true of the early flowers. Pollen that could have come from either relictual dicots or monocots is found in Lower Cretaceous strata.

FIGURE 23-6 A leaf of Magnoliaephyllum showing strong similarities to the leaves of living magnolias; however, fine details visible by microscopy show that this is not identical to a true Magnolia leaf.
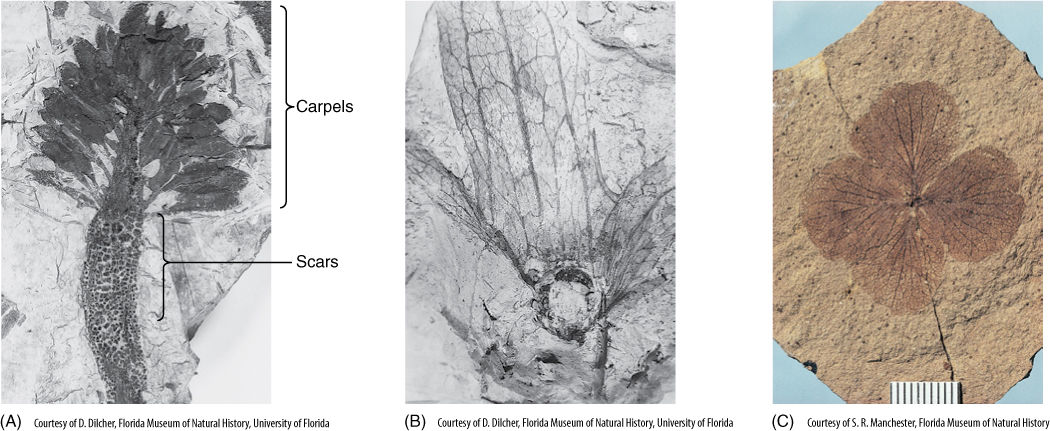
FIGURE 23-7 (A) Part of a flower from the Cretaceous Period; numerous spirally arranged carpels were still attached at the time it died and started to fossilize. The spiral arrangement is similar to that found in flowers of Magnolia and water lily. Below the carpels are spirally arranged scars where structures abscised; we do not know if they were sepals, petals, or stamens. (B) Part of a fossil fruit, Paraoreomunnea puryearensis, approximately 60 million years old. It was a winged fruit similar to those produced today by members of Juglandaceae, the family of walnuts and hickories. (C) Fossil of a Hydrangea flower, 44 million years old. This flower is much more modern and derived than that of (A), having fewer parts.
Much attention is now being given to gymnosperms of the Jurassic and Triassic Periods, and the focus is centering on cycadophytes and glossopterids (FIGURE 23-8). As more is learned about this group of gymnosperms, we realize how much the various groups had begun to develop angiosperm-like features.
As you read the following discussion about flowering plants, it will be helpful to study a timeline chart on the evolution of true plants.
![]() Classification of Flowering Plants
Classification of Flowering Plants
Magnoliophyta is such a large group with so many families, genera, and species that it is rare for an individual taxonomist to attempt to study and classify the entire group. In the 1980s and 1990s, the most widely used monograph of the entire division was An Integrated System of Classification of Flowering Plants by Dr. Arthur Cronquist of the New York Botanical Garden. Since then, cladistic studies involving DNA, biochemistry, and anatomy often proposed phylogenies that agreed well with those of Dr. Cronquist for certain clades but disagreed for others. The areas where the two systems were inconsistent became the topics of more intense scrutiny of all characters, with subsequent adjustments in our concepts of phylogeny and classification. Currently, two widely used reference books are Plant Systematics, A Phylogenetic Approach (3rd edition) by W. S. Judd et al., and Plant Systematics by Michael Simpson. Their approach is the basis for the treatment used here. Also of special importance is the Angiosperm Phylogeny Group (APG), which is a semi-informal collaboration of many systematists; their classifications can be found on the website for the Missouri Botanical Garden, www.mobot.org/MOBOT/research/APweb/.

FIGURE 23-8 A reconstruction of a Glossopteris, a possible relative to the ancestors of flowering plants. The leaves had a prominent midrib and reticulate venation. The leaves were deciduous, and the wood had annual rings, indicating that it lived in a temperate climate.
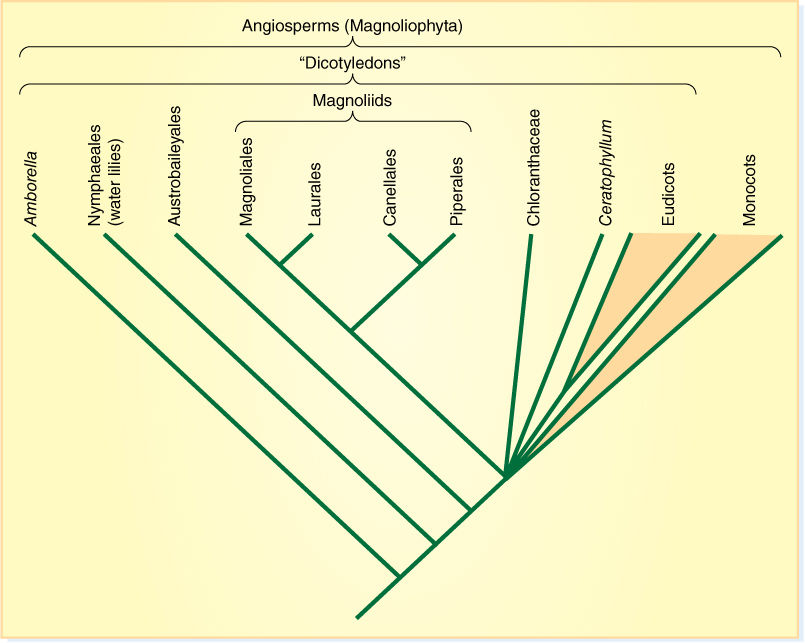
FIGURE 23-9 One recently proposed cladogram of angiosperms. Basal angiosperms are Amborella, Nymphaeales, Austrobaileyales, and magnoliids. The origin of Chloranthaceae, Ceratophyllum, monocots, and eudicots is an unresolved polychotomy.
Soon after their origin, flowering plants began to follow two distinct lines of evolution, and currently, almost all angiosperms are classified as monocots or eudicots (FIGURE 23-9). No single character always distinguishes a monocot from a eudicot, and some species would fool most botanists. In general, monocots, as their name implies, have only one cotyledon on each embryo, and other typical characters are the following: Their leaves usually have parallel veins because the leaves are elongate and strap shaped (grasses, lilies, and irises); vascular bundles are distributed throughout the stem, not restricted in one ring, and monocots never have ordinary secondary growth and wood (a few have anomalous secondary growth). Flowers of monocots have their parts arranged in groups or multiples of three: three sepals, three petals, three stamens, and three carpels (FIGURE 23-10). Eudicots are much more diverse and include a greater number of families, genera, and species. Eudicots have two cotyledons and reticulate venation in the leaves; vascular bundles occur in only one ring in the stem. They can be woody, herbaceous, or succulent or have any of many highly modified forms. Flower parts occur in sets of five most often and sometimes in sets of four, but they only rarely occur in sets of three.
A surprising result of cladistic studies is that the monocot/eudicot divergence did not occur right away, but instead, the early angiosperms diverged into several clades now called the basal angiosperms. These are not newly discovered species; it is just that we had classified them as monocots or dicots before, but many different DNA sequences indicate they had become reproductively separate from the other angiosperms very early. Of the basal angiosperms, Amborella and Austrobaileyales greatly resemble eudicots; however, they cannot be classified with the eudicot clade because the common ancestor that includes both them and the eudicots also includes monocots, and we cannot construct a group that includes some descendants of a common ancestor but excludes others. Similarly, Nymphaeales (water lilies) have many features of monocots, but classifying them as monocots would create a polyphyletic group.

FIGURE 23-10 Lilies and their relatives are excellent for demonstrating the typical monocot flower: They have three of each type of appendage and all are large enough to be seen easily. The three carpels are fused together, but a transverse section would show three chambers. One petal and two stamens were removed to reveal the ovary.
Plants and People
BOX 23-1 Maintaining Genetic Diversity
The flowering plants are particularly adept at evolving rapidly; in the short time they have existed, they have diversified greatly. The result is that hundreds of thousands of species exist, each differing from the others in features such as flowers, fruits, vegetative structure, and metabolism. Within this panoply of diverse plants, we and our ancestors have discovered a wealth of foods, medicines, and soul-soothing flowers.
Now, at the beginning of the 21st century, we have become aware of the horrific ecological destruction occurring on Earth, with vast areas of vegetation being cut or burned or flooded with artificial lakes behind hydroelectric dams. We have also begun to realize that we are causing the extinction of hundreds of species every year and, in the process, their genetic information is being lost. On a philosophical basis, we can debate whether one species, Homo sapiens, has the right to destroy so many other species, but on a more pragmatic basis, we should consider whether we are unwittingly destroying potential sources of new foods and medicines. Consider just two examples: A relatively unknown species, quinoa (Chenopodium quinoa), can be grown in relatively dry regions with poor soil, yet it produces an abundance of nutritious, protein-rich seeds. And Pantadiplandra brazzeana produces a small protein that is 2,000 times sweeter than sugar; this could possibly be used as a completely natural, safe, nutritious, low-calorie sweetener, decreasing the amount of sugar or artificial sweeteners that we consume.
How can botanists preserve the genetic resources of the plants, animals, and microbes that coexist with us? Several types of efforts are needed, and two particularly important ones are habitat preservation and germ plasm banks (germ plasm refers to the cells that produce gametes). Germ plasm banks were started several years ago as rather simple seed storage facilities, where bags of many types of seeds were maintained under cool, dry conditions. After several years, while most seeds were still healthy, they would be germinated, grown into mature plants, and then their fresh, healthy seeds would be collected and stored. After several years, the process would be repeated.
This is still done for many species, but now the seeds can be kept for extremely long periods, held at very low temperatures or even in liquid nitrogen. Furthermore, it is now possible to maintain some seeds as tissue or cell cultures, either grown slowly in culture tubes or kept as frozen cells. In the foreseeable future, it may be that some plants, or at least some of their genes, will be kept simply as DNA molecules in solution or in cloning vectors.
Currently, there are over 100 germ plasm banks throughout the world, preserving at least 3 million types of plant material. Examples are the Svalbard Global Seed Vault, built in an abandoned coal mine in the permafrost of northern Norway, and the National Seed Storage Laboratory (NSSL) located at Colorado State University and run by the U.S. Department of Agriculture. The NSSL was established in 1958 and has more than 232,000 seed types. At present, its size is being quadrupled, and its capacity will be more than 1 million seed samples. The Millennium Seed Bank Project at Kew Gardens in England reached its goal of safeguarding 25% of all the world’s plant species in October 2020.
The purpose of germ plasm banks is not merely to store and maintain plant genetic diversity but also to make it available. Botanists can request samples and use them for research and plant breeding programs. If some are found to have useful traits, they can be grown as crops. Of course, it is also hoped that in many cases we can use these plants to help restore or rebuild ecosystems that we have already damaged.
![]() Basal Angiosperms
Basal Angiosperms
The basal angiosperms contain the living descendants of several groups that originated while angiosperms were still a young clade. Their ancestors became reproductively isolated from the other early flowering plants before distinctive angiosperm traits had originated. It would be easy to assume that the descendants of the three earliest diverging groups would all display a large number of primitive traits and few derived ones: They “should” look primitive, and they “should” look like taxa that had just barely become distinct from some gymnosperm sister group. All organisms evolve, however, and so did the descendants of the basal angiosperms. The three groups of extant descendants of these clades—Amborellaceae, Nymphaeaceae, and Austrobaileyales—have not remained static evolutionarily and have not preserved all ancestral features intact. One group should be familiar to you: Nymphaeaceae are the water lilies (FIGURE 23-11). They are small, soft-bodied herbs with vascular bundles scattered like those in monocots, and they completely lack any wood; in most, their stems must be submerged underwater, and exposure to ordinary conditions on land would kill them. Their large colorful flowers are pollinated by animals. They have almost nothing in common with either gymnosperms or seed ferns. Amborella trichopoda (there is only one species in this clade) is very different, being small trees in forests of New Caledonia. Their wood contains tracheids but no vessels and little parenchyma. This species is dioecious, some plants having staminate flowers, others having carpellate flowers (and staminodes that indicate these evolved from bisexual ancestors). Both flowers are small with 5 to 11 tepals (not differentiated into sepals and petals). Staminate flowers have numerous stamens, and carpellate flowers have 5 or 6 carpels whose edges do not seal tightly but instead have interlocking, secretory hairs that act as a stigma. Austrobaileyales contains woody trees with bisexual flowers, with stamens and carpels being similar to those of Amborella.

FIGURE 23-11 (A) Flowers of water lilies have numerous sepals, stamens, and so on, with all parts arranged in spirals like the leaves of a rosette plant. This is believed to be a relictual feature similar to a gymnosperm cone. Monocot flowers would be derived from an ancestral flower like this by mutations that result in only three or six of each type of floral appendage. (B) Amborella.
Because water lilies, Amborella, and Austrobaileyales differ in so many features, they cannot all resemble the earliest angiosperms. Instead, one or the other—or all three—have undergone considerable evolutionary change since their clades originated. Water lilies have little in common with either gymnosperms or seed ferns; almost certainly many of their features have changed considerably. Amborella and Austrobaileyales appear to have changed little, but it is important to realize that they have not remained static. We cannot assume that every feature of these woody trees is like that of early angiosperms; instead, we assume that features present in them and gymnosperms are relictual features, plesiomorphies.
Ancestors of the magnoliid clade diverged from early angiosperms a bit later than Amborella and Austrobaileyales, but this conclusion is based mostly on DNA evidence—morphologically, the magnoliids do not differ greatly from these two earlier clades (TABLE 23-2). For example, family Magnoliaceae contains trees with wood similar to that of gymnosperms in that it lacks vessels, fibers, and axial parenchyma; Magnolia flowers have numerous stamens and carpels arranged in spirals (see Figure 23-5B). Furthermore, carpels are not fused together into a pistil as occurs in almost all monocots and eudicots. Another important feature is that their pollen grains have only a single germination pore: They are uniaperturate, as are all other basal angiosperms and monocots. Eudicots have three germination pores (FIGURE 23-12). Other magnoliids are laurels and avocado (Laurales) and peppers and peperomias (Piperales).
TABLE 23-2 Representative Families of the Magnoliid Clade

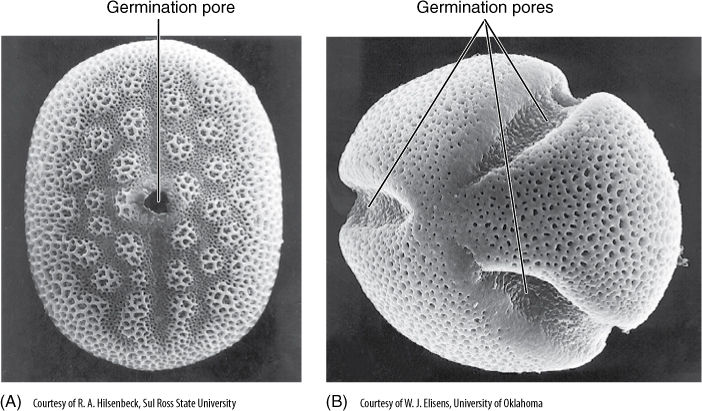
FIGURE 23-12 (A) All early fossil pollen has only one aperture, but almost all living eudicots with many derived features have three apertures; therefore, uniaperturate pollen in living angiosperms, such as this Siphonoglossa, is considered a relictual feature (×300). (B) Pollen of Lophospermum has three germination pores; the pores are located in grooves, something that does not occur in any early fossil pollen (×300).
![]() Monocots
Monocots
Currently, monocots are widely believed to have arisen from early angiosperms approximately 80 to 100 million, perhaps even 120 million years ago. Because all monocots lack ordinary secondary growth and wood, their ancestors were probably herbs with either no vascular cambium or little cambial activity. The gynoecia of many monocots are composed of several carpels (usually three) that are either free of each other or at most only slightly fused together (Figure 23-10). The perianth usually consists of three outer and three inner members; they often look so similar that rather than using the terms sepals and petals, the perianth members are just called tepals. Monocot tepals rarely fuse to each other to form a tube; instead, they remain free. These features must have been present in the group from which the monocots evolved. Water lilies (Nymphaeales) have similar features. An early set of angiosperms may have resembled Nymphaeales and may have given rise to both the water lilies and the monocots.
One theory about the parallel venation of monocot leaves postulates that the ancestors had broad leaves and lived as aquatic plants. The leaves evolved to a more reduced, simple type without a blade, a form more adaptive for a submerged leaf (FIGURES 23-13A and B). Some of these plants moved into drier habitats where their leaves were not submerged and a broad or long lamina would be advantageous. Mutations that resulted in a basal meristem were selected, resulting in strap-shaped leaves (FIGURE 23-13C). In the evolutionary lines of “broadleaf” monocots such as palms, philodendrons, and dieffenbachias, a marginal type of meristem evolved; it was located at the end of the residual leaf, resulting in the formation of what appears to be a petiole and lamina (FIGURE 23-13D and FIGURE 23-14).
Early monocots diverged into a series of clades whose extant members are classified into approximately ten orders (FIGURE 23-15).
Alismatales
Alismatales contains many aquatic herbs such as Sagittaria (arrowhead; FIGURE 23-16) and many aquarium plants such as Hydrocharis, Najas, and Hydrilla (TABLE 23-3; see Figure 23-4C). These plants are most often found in swamps and marshes, partly or entirely submerged. Although many species retain a large number of plesiomorphic features, others have become highly modified in response to the unusual selection pressures associated with an aquatic habitat. Plants of Hydrocharitaceae, Najadaceae, Posidoniaceae, Ruppiaceae, and Zosteraceae grow completely submerged in seawater and are known as “sea grasses.” They have no transpiration, and thus, mutations that result in the loss of stomata are not selectively disadvantageous. Air chambers make the plants buoyant; therefore, mutations that prevent the formation of lignified fibers are selectively advantageous. The plants do not waste carbon and energy by producing fibers that are not needed for support. Such plants tend to be thin and delicate, having very little sclerenchyma and almost no xylem. Tissues have been lost or simplified; the ancestors of Alismatales were more complex and massive and had more sclerenchyma. Members of Alismataceae have a part of their bodies emergent above the water’s surface, and some are entirely emergent whenever their marsh habitat dries temporarily. These plants are less highly modified, often having large leaves, considerable amounts of fiber, and a thick cuticle on the leaf epidermis. It is necessary for a species to be adapted to the most adverse conditions that it frequently encounters during one plant’s lifetime.

FIGURE 23-13 (A) Monocots are believed to have originated as semiaquatic plants that inhabited swamps and marshes. (B) The leaf may have consisted only of a leaf base and petiole, the entire blade having been lost evolutionarily. (C) A “pseudolamina” evolving from a petiole. (D) A “broad leaf” evolving from a petiole.

FIGURE 23-14 Some monocots, such as this banana (Musa), have broad leaves, but their lamina is not homologous to that of a dicot leaf. In this monocot broad leaf, all vascular bundles run parallel to each other from the midrib to the margin, whereas in a dicot lamina, they diverge and branch into a reticulate pattern.
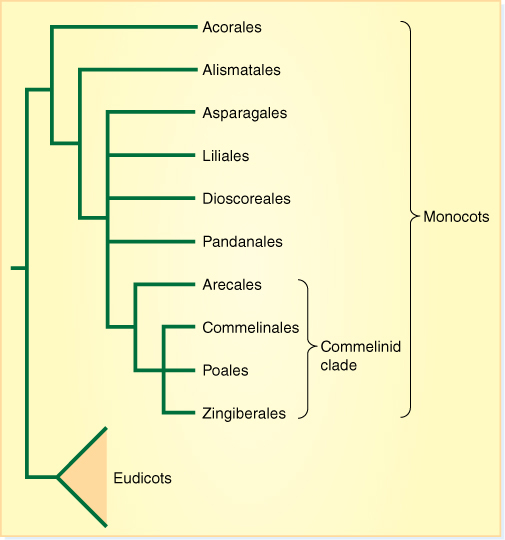
FIGURE 23-15 One recently proposed cladogram of monocots. There are many unresolved polychotomies.

FIGURE 23-16 Sagittaria, a member of Alismatales, is dioecious; plants have either staminate (A) or carpellate (B) flowers.
TABLE 23-3 Representative Families of Alismatales
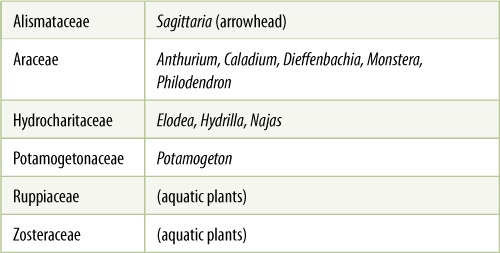

FIGURE 23-17 (A) This Anthurium (Araceae) is an aroid, one of the monocots that have a eudicot type of broad leaf; compare its reticulate venation with the parallel venation of banana (Figure 23-14). Many members of this family are adapted to conditions of low light and high humidity; most are members of tropical or subtropical habitats. (B) Araceae have a characteristic inflorescence with a sheath-like bract (a spathe) surrounding a thick column (a spadix). Tiny staminate flowers are embedded in the top of the spadix, carpellate ones in the base (hidden by the spathe here). Peltandra virginica.
In the most relictual members of Alismatales, flowers are large and showy, with three sepals and three petals, but in others, they are highly modified, especially in those species in which the flowers do not emerge from the water and pollination occurs below the surface. Such flowers often completely lack a perianth.
Family Araceae (aroids) contains numerous familiar houseplants: Philodendron (250 species), Anthurium (500 species), and Arisaema (Jack-in-the-pulpit; 100 species). Dieffenbachia (dumb cane) and 1,800 other species also belong here (FIGURE 23-17). This family is characterized by the evolution of a distinctive inflorescence: tiny flowers, either unisexual or perfect, embedded in a thick stalk, the spadix. Staminate flowers are located near the top of the spadix, carpellate flowers near the base. The spadix is surrounded by a large bract, the spathe.
Liliales
In the past, Liliales was defined broadly as a large group with many highly derived families; it was basically defined as the “petaloid” monocots—those with large, colorful flowers. Recent DNA evidence indicates that Liliales is a smaller clade with approximately 11 families and 1,300 species. Many of its previous families are now grouped in Asparagales. Important features in Liliales are the presence of spots or lines on the petals and of rather ordinary nectaries formed at the bases of tepals or stamens; flowers in Asparagales typically have septal nectaries (discussed below). One of the largest families, Liliaceae, contains so many ornamental plants, mostly bulbs, that most people are familiar with them and think of them as the “typical” monocots (Figure 23-10). Liliaceae contains Lilium (lilies), Tulipa (tulips), Calochortus (mariposa lily), and Fritillaria. Colchicaceae contains Gloriosa (flame lily) and Colchicum, the source of the alkaloid colchicine that prevents microtubule polymerization.
Perhaps the most unusual member of Liliales is Smilacaceae, with the single genus Smilax (catbrier). Despite being closely related to lilies, these are tough, fibrous vines with leaves that have a petiole and broad lamina with reticulate venation—not lilylike at all. At first glance, it is easy to assume they are dicots.
Asparagales
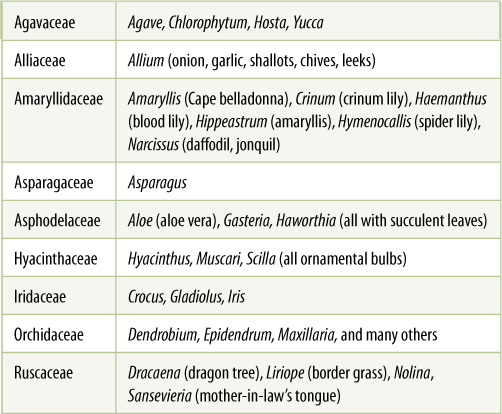
This is a large clade with many families, species, and types of biology. By examining this clade, we get a sense of evolution as diversification (TABLE 23-4). There are several morphologic and DNA synapomorphies that unite this group. Most form nectaries in an unusual way: Their carpels fuse side by side starting at their bases, the fused sides being called septa. They do not fuse all the way up to the style, however, and the open areas secrete nectar; these are septal nectaries. Beyond this, the group is extremely diverse in morphology (FIGURE 23-18), ranging from small, delicate bulbs like Hyacinth, chives, and onion (Allium) to vining epiphytes such as many orchids. Bulbs, rhizomes, and corms are common. Agaves and yuccas have giant, fibrous perennial leaves. Iris has flattened sword-shaped leaves, and many members such as Narcissus (daffodils) have delicate leaves that live only during spring and summer. Several members of Agavaceae (Yucca brevifolia, Joshua tree) and Ruscaceae (Dracaena, dragon tree) have anomalous cambia and grow to be large, highly branched trees. Asparagales has diversified biochemically as well, with different groups having distinctive chemical compounds, such as the sulfur compounds in onion and garlic or particular alkaloids in Amaryllidaceae. Flowers are large, colorful, and showy for the most part, and in Orchidaceae, numerous types of insect pollination have evolved, many accompanied by distinctive aromas.
TABLE 23-4 Representative Families of Asparagales
The Orchidaceae is the largest and most diverse family of Asparagales. The most familiar, ornamental ones are epiphytic, but many are terrestrial and one is a subterranean parasite. Orchid flowers are highly modified from the ancestral conditions of angiosperms, being zygomorphic (bilaterally symmetrical) with complex shapes, colors, and fragrances that attract specific pollinators. Orchids are unusual in producing hundreds or thousands of tiny seeds in each fruit. The seeds are dust-like and so undeveloped at “germination” that they must form a symbiosis with certain fungi in order to survive long enough to form roots and leaves.

FIGURE 23-18 Members of Asparagales. (A) Iris flowers appear to have many unfamiliar parts, but the uppermost petaloid parts are actually stamens; the long dark regions are sporogenous tissues, the anthers (some cultivated irises are sterile and do not have sporogenous tissue). In many angiosperms, structures that appear to be petals are really sterile stamens. (B) Like bamboos and palms, yuccas and agaves prove that herbs do not have to be soft and small. The leaves are long and narrowly triangular with parallel venation, and they may be thick at the base. They are arranged as a rosette around a short (agaves) or long (yuccas) stem. Most have a needle-sharp point and razor-like hooked spines along the edges of the leaf.
Dioscoreales
This small order has only one family, Dioscoreaceae, and is mentioned because it has a familiar, important food crop and unusual morphology. Yams are starchy “tubers” produced by several species of Dioscorea and are a major source of carbohydrates for many people of tropical areas. Also, dioscoreas have petiolate, broad leaves with reticulate venation and are easily mistaken for dicots.
Commelinoid Monocots
The following four orders of monocots are known as the commelinoid monocots (Figure 23-15) because they differ from the others in several unusual synapomorphies: They have unique types of epicuticular wax. Their walls have unusual types of hemicelluloses and ultraviolet-fluorescent compounds. Their pollen contains starch, as does their endosperm. Multiple studies of rbcL, atpB, and other DNA regions support this as a distinct clade.
Arecales
This order contains familiar plants, the palms, in family Arecaceae (an old name for the family is Palmae; FIGURE 23-19). There are about 3,500 species, all of which are easily recognizable by their solitary trunk (only a few species have branched trunks), which varies from 1.0 m wide in some and only 1.0 cm in others; all have scattered vascular bundles, which is typical of monocots. Leaves of palms always occur only near the shoot apex, never distributed along the length of the stem. In a few species, trunks are prostrate (palmetto palm, Sabal) or vines (Daemonorops). All species have simple leaves that, after fully expanded, are torn by wind into either a pinnate pattern (feather palms) or palmate one (fan palms). Coconuts (Cocos) and dates (Phoenix) are two types of palm fruit, but many others exist. Palm flowers are seldom seen because they are usually tiny, approximately 5 mm across, and are formed only high up in the tree.

FIGURE 23-19 (A) Palm flowers usually occur in large inflorescences at the top of a plant. (B) Although palm flowers are small, they have typical monocot organization with their parts in sets of three. Hyphaene coriacea.
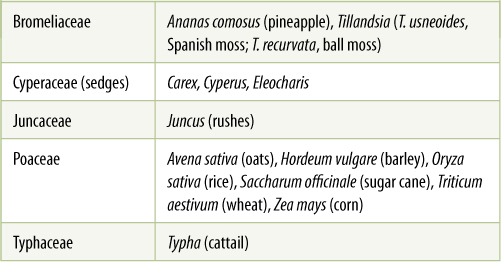
Poales
This order contains the grass family Poaceae as well as several other familiar groups such as cattails (Typha), bromeliads, and sedges (TABLE 23-5).
Poaceae contains about 8,000 species and are much more than just the plants in the lawn (FIGURE 23-20). They also include most foods, such as wheat (Triticum), barley (Hordeum), oats (Avena), rye (Secale), corn (Zea), rice (Oryza), and sugar cane (Saccharum), as well as a major building material of the tropics, bamboo (the subfamily Bambusoideae). Humans began farming grass crops as early as 10,000 years ago, and approximately 50% of all calories consumed by people come from grass seeds. Also, our main sources of meat—cattle, pigs, and sheep—are raised on grassland and fed corn. Grasses are abundant in flat, open, dry regions in the central areas of all continents, such as the rangelands of the United States and Africa, the steppes of Russia and Ukraine, and the pampas of Argentina; this accounts for one quarter of all vegetated land on Earth. Some grasses grow as tight clumps, and others spread by rhizomes or runners; either way, their shoot apical meristems tend to be located near the ground, protected from fire and grazing animals. As a result, they survive both better than most other types of plants. Widespread grasslands are a new ecologic development, however, arising less than 25 million years ago. During the Miocene epoch, extensive forests disappeared and were replaced by grasslands, making possible an increase in grazing animals and perhaps even in ancestral hominids switching from a life in trees to one on the ground in African rangelands.
TABLE 23-5 Representative Families of Poales
All grasses are wind pollinated, so sepals and petals are of little importance and are reduced to bristle-like structures. The three carpels are fused together, but in most grasses, the three stigmas remain separate. An “ear” of corn is an inflorescence, and each “silk” is a compound style and stigma. Within the fused ovary of grasses there is just one ovule; once fertilized, it matures into a seed whose seed coat fuses firmly to the developing fruit wall. The objects we usually call “seeds” of corn, wheat, and oats are actually single-seeded dry fruits known as caryopses (singular, caryopsis). Nearly the entire bulk of the seed is endosperm; the embryo is small but well developed.

FIGURE 23-20 Grass flowers are very reduced, simplified, and wind pollinated. The usual two small, dry scales (called a lemma and a palea) are thought to be remnants of an ancestral bract and two fused tepals; the third tepal does not form. Three anthers with long filaments and three carpels fused together with long feathery styles and stigmas are usually present. Grass flowers occur grouped together in complex, compact inflorescences, and inflorescence characters are important for identifying grasses.
Closely related to grasses are sedges (Cyperaceae with 4,000 species; FIGURE 23-21) and rushes (Juncaceae). All three families contain plants of wet, marshy areas, with tiny, reduced flowers and small dry fruits. Sedges and rushes are often confused with grasses, but “sedges have edges, rushes are round, and grasses are hollow from top to ground.” Sedge stems are triangular in cross-section. Those of rushes are round and solid, and those of grasses are round but hollow. This simple rhyme is mostly true, but there are exceptions.
Other members of Poales are cattails (Typhaceae) and Bromeliaceae (bromeliads). Cattails grow in ponds and marshy areas. Each plant spreads by thick, horizontal subterranean rhizomes, with some axillary buds growing out as more rhizomes, and other axillary buds growing upward and producing the familiar green leaves: What appears to be a cattail plant is really just a single branch. Their characteristic reproductive structures consist of a tip with many staminate flowers and thousands of carpellate flowers, although by the time we see them, the staminate flowers have usually withered and the carpellate flowers have matured into tiny fruits (Figure 23-21B).
Bromeliaceae contains some of the most beautiful tropical epiphytes, their large, brightly colored inflorescences being easily visible even in thick jungle vegetation (FIGURE 23-22). Epiphytic species extend as far northward as the subtropics, Spanish moss and ball moss (Tillandsia) occurring from Florida to Texas. Other species are terrestrial and usually xerophytic, such as Puya of Chile and Peru and pineapples (Ananas). These xerophytic terrestrial forms are believed to be relictual, and their capacity to withstand drought may have made it easier for bromeliads to adapt to life as epiphytes. Because bromeliads occur in only the Americas, it is assumed that they evolved after South America separated from Africa approximately 80 million years ago. Had they evolved earlier, numerous species should occur in the African rain forests and coastal deserts.
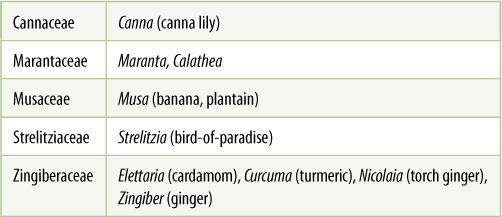
Zingiberales
This order contains some of the most familiar of all houseplants: Maranta, Calathea, canna lilies (Canna), and gingers (Zingiber, Hedychium), as well as some that are best known in the warmer southern states—banana (Musa) and bird-of-paradise (Strelitzia) (TABLE 23-6). Members of Zingiberales differ from most other monocots in that they tend to have large, showy flowers pollinated by insects, birds, or bats. Furthermore, many of their flowers have derived features; adjacent sepals are often fused to each other, forming a tube, and the same is often true of the petals; many flowers are bilaterally symmetrical. The gynoecium typically consists of three carpels that have fused almost completely. Very often the gynoecium is inferior, located below the sepals and petals, a condition that is always considered quite derived, the result of considerable evolutionary modification.

FIGURE 23-21 (A) Most sedges grow in wet, marshy areas. (B) Cattail, Typha, has hundreds of staminate flowers above and thousands of carpellate flowers below. As a cattail breaks apart, the tiny one-seeded fruits blow away; seedlings are miniscule, like short threads.
The largest family, Zingiberaceae, has approximately 1,000 species. Almost all are tropical, and most are soft, nonleathery herbs. The leaves are broad and have a petiole (Figures 23-13 and 23-14). During the evolution of the Zingiberales, mutations that resulted in the petiole/leaf becoming flattened and broadened were selectively advantageous: The plants grow in the heavy shade of jungle understory where light interception is important.

FIGURE 23-22 (A) Many epiphytic bromeliads catch rainwater with a tank made by their overlapping leaves; they absorb water through their leaf bases rather than through their roots. Their flowers develop underwater. (B) Spanish moss, Tillandsia usneoides, is a bromeliad, not a moss. It grows attached to tree trunks as an epiphyte, not a parasite. This species grows in the southeastern United States, where humidity is high and rainfall is frequent. (C) Many bromeliads have brilliantly colored flowers and bracts, which may make the flowers more visible in the dim light of rain forests.
TABLE 23-6 Representative Families of Zingiberales
![]() Eudicots
Eudicots
The eudicots constitute a much larger group than the monocots and are divided into numerous clades, some of which have diversified into many families, others having fewer members (FIGURE 23-23). Virtually every type of organ, tissue, and metabolism has several or many variations, so they are more difficult to characterize than are the monocots. One especially distinctive feature is that their pollen grains have either three germination pores (tricolpate) or have some condition derived from the tricolpate mechanism (Figure 23-12). Furthermore, their flower parts are arranged in whorls, not spirals, and their stamens usually have a well-defined filament (stalk) and anther; they are not flat or leaflike at all. These characters are greatly modified from those of sporophylls found in other vascular plants. Current cladograms of eudicots have a large number of orders, too many to be covered here. Those that have special importance and that contain plants that you might know are presented below.
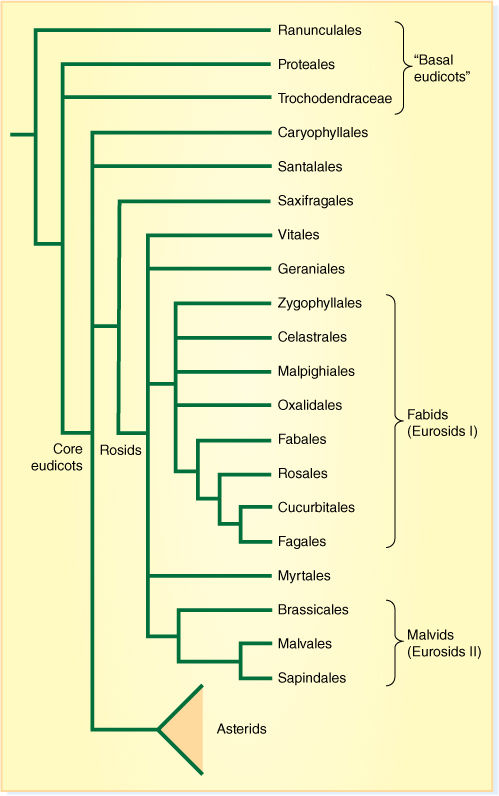
FIGURE 23-23 One recently proposed cladogram of eudicots. There are several unresolved polychotomies
Basal Eudicots
In older classifications in which all flowering plants were assigned to either Monocots or Dicots, many families did not seem to fit well in either group. Several, such as water lilies, are now assigned to the basal angiosperms; Ranunculales and several others are believed to be clades that diverged at early stages in eudicot evolution.
Ranunculaceae is interesting because its flowers have so little fusion of parts: Each flower usually has many stamens and carpels, all of which remain separate of the others. Stamens may remain free or may fuse together into a tube or even fuse to the carpels. Familiar members of Ranunculaceae that have numerous, free parts are buttercups, windflower (Anemone), and Clematis (FIGURE 23-24A).
The poppy family, Papaveraceae (FIGURE 23-24B), is well-known for its numerous ornamental species such as Argemone (prickly poppy), Eschscholzia (California poppy), and Papaver (poppies). Papaver somniferum is opium poppy, the source of a milky latex harvested for opium; morphine, a strong analgesic (pain killer) that depresses the cerebral cortex, is extracted from opium, as is codeine. Morphologically, poppies are unusual in being either herbs or shrubs with a very soft wood.
Several hamamelid families are now believed to be basal eudicots rather than basal angiosperms. These are large trees with reduced wind-pollinated flowers, usually in dangling inflorescences containing many staminate flowers and just a few carpellate flowers. A familiar example is Platanaceae, sycamores (also called plane trees, Platanus): These large trees have small, inconspicuous clusters of flowers in the spring that mature into spherical clusters of hundreds of tiny dry fruits.
Caryophyllales
DNA-based studies have combined a large number of families into a group called Caryophyllales (TABLE 23-7). Most of these families had long been known to be closely related and were also known as Caryophyllales; in new treatments, the old group is referred to as “core Caryophyllales.” Examples of the core group are cacti (Cactaceae), iceplant (Aizoaceae), portulaca (Portulacaceae), bougainvillea and four-o’clocks (Nyctaginaceae), spinach, beets, and Russian thistle (Amaranthaceae), and carnations and chickweed (Caryophyllaceae) (FIGURE 23-25). The core Caryophyllales share many derived characters, but one is especially important. Whereas other flowering plants have anthocyanin pigments in their flowers, almost all Caryophyllales instead produce a group of water-soluble pigments called betalains (FIGURE 23-26). The ancestral group may have lacked petals in their flowers, a condition common in many Caryophyllales; although many appear to have petals, we believe that these are modified sepals in some families and modified stamens in others.

FIGURE 23-24 (A) Carpels in Ranunculales remain free of each other at maturity; they do not fuse during development. Consequently, each flower produces many separate fruits. Clematis. (B) Poppies (Papaveraceae) have numerous free stamens but fused carpels. They have no styles, and thus, the stigmatic surface is the top of the ovary.
Another unifying character of Caryophyllales is that endosperm develops only a little and then fails to continue growing. Instead, nucellus cells proliferate and form a nutritive tissue called perisperm, which surrounds the developing embryo (FIGURE 23-27). Perisperm is usually absorbed almost completely by the time the seed is mature.
A third feature that unites this order and distinguishes it from all other taxa is the nature of sieve tube plastids. In Caryophyllales, phloem plastids contain deposits of fibrous protein located as a ring just interior to the plastid membrane (FIGURE 23-28). In other angiosperms, plastids contain either starch or crystalline protein with a different structure.
TABLE 23-7 Representative Families of Caryophyllales
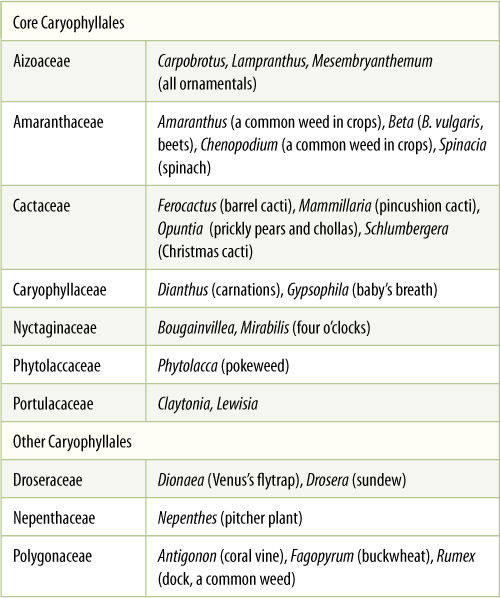

FIGURE 23-25 Caryophyllales: (A) Echinocereus coccineus (Cactaceae) and (B) Carpobrotus (Aizoaceae). The brilliant colors in the petals of both are due to betalain pigments.
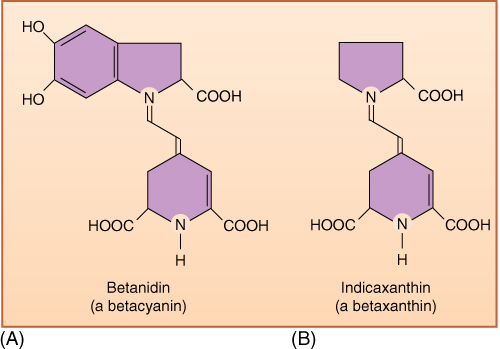
FIGURE 23-26 Core Caryophyllales are characterized by the presence of betalain pigments in flowers and fruits. There are two basic types: (A) betacyanins (red to violet), and (B) betaxanthins (yellow).
The Caryophyllales are postulated to have arisen from ancestors similar to Ranunculaceae. Most members of Caryophyllales are herbaceous, with either no wood, very little wood, or unusual, anomalous wood, so the ancestral group is suspected to have been herbaceous or shrubby, having lost the capacity for extended, massive secondary growth. The time of origin of Caryophyllales is approximately 70 to 80 million years ago, at an important time of continental drift. At that time, all land in the southern hemisphere was joined together in a giant continent now called Gondwana. Caryophyllales originated in southern Gondwana, and its earliest families spread across that continent. Then continental drift caused Gondwana to break up, first with South America moving westward and later with Australia, India, and Antarctica moving eastward. Families that had already come into existence were carried along with the moving land masses and are now present on most or all the southern continents (except for Antarctica, whose southward drift doomed all the plants it carried). Families that originated after the breakup of Gondwana arose in individual continents and are now mostly confined to those. For example, cacti arose in South America and later spread to Central and North America, but its seeds could not cross either the Atlantic or Pacific Oceans; thus, cacti do not occur naturally on any other Gondwanan continent. Similarly, Didieriaceae arose in east Africa and Madagascar and occur nowhere else.
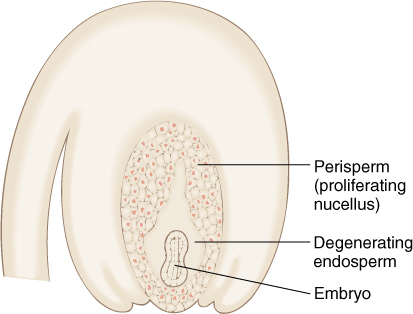
FIGURE 23-27 In core Caryophyllales, endosperm develops for only a short while or not at all and is not sufficient to nourish the embryo. Instead, the nucellus (megasporangium) cells proliferate and act like endosperm, forming a tissue called perisperm.
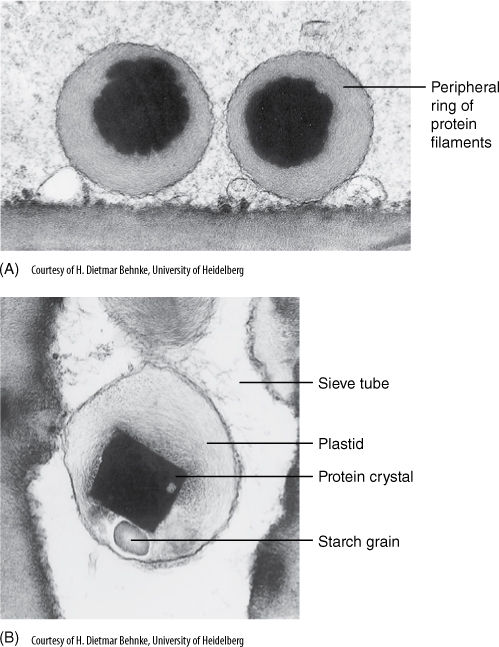
FIGURE 23-28 Plastids in sieve tube members are simple, but they may accumulate particles of starch or protein or both. The nature of the accumulated material is highly specific with regard to the family, and phloem plastid analysis is important in studying the evolution of flowering plants. Sieve tube plastids of Monococcus (A) lack a central cubic protein crystal, but those of Macarthuria have one (B). Both are members of Caryophyllales, and as a result, both have a peripheral ring of protein filaments (both × 40,000).
Several of the other families that are now placed in the expanded Caryophyllales are the buckwheats (Polygonaceae), pokeweeds (Plumbaginaceae), and two groups of carnivorous plants, Droseraceae (sundews and Venus’s flytrap) and Nepenthaceae (pitcher plants).
Santalales
This is a small order of highly modified plants, most of which are parasitic. The sandalwood family (Santalaceae) contains the large tree Santalum from which sandalwood incense is obtained. It appears to be an ordinary tree, but its roots make fine, almost imperceptible connections to roots of surrounding plants and parasitizes them. Also in this family are common mistletoes, Viscum and Phoradendron, which are used as decorations at Christmas (FIGURE 23-29). These mistletoes have chlorophyll and are photosynthetic; therefore, they are just hemiparasitic, but some members are holoparasitic, having no chlorophyll at all. Viscum minimum lives entirely within its host plant, Euphorbia. Loranthaceae is a large family with about 900 species, all of which are parasitic, but none of which occurs in temperate North America.

FIGURE 23-29 Mistletoe (Phoradendron tomentosum) is a hemiparasite; it carries out some photosynthesis. Its haustorial root system penetrates the host tree branch at the top of the photograph.
Rosid Clade
The remaining eudicots are members of two very large, very diverse clades, the rosids and asterids (see Figure 23-23). The rosid clade (named for the rose order Rosales) consists of many families that, taken as a whole, are so diverse with respect to vegetative body, flowers, chemistry, and ecology that it is difficult to see they are all related; however, some share enough characters with others to indicate a relationship, and those of the second group share different features with a third group and so on until a larger picture of phylogenetic relationships can be distinguished. DNA analysis has added a great deal of information to identifying patterns of relationships.
The rosids consist of several small orders and two large groups. One of the small orders has few members but great economic significance. Vitales contains Vitaceae, the grape family. Vitis is the genus of grapes that give us juice, raisins, and table grapes. Vitis vinifera is the wine grape, and despite the enormous variety of wines, virtually all are made from varieties of this one species. The other small order, Geraniales, contains the geranium family, Geraniaceae.
The two large clades of rosids are the fabids (also called eurosids I) and the malvids (eurosids II). These two clades together contain more than 100 families, and it is difficult to give any universal characters (TABLE 23-8). As a group, rosids are more derived than basal tricolpates from which they arose and have derived characters that differ from those of Caryophyllales. Rosids are especially interesting in that none of them has any of the highly relictual features found in many basal angiosperms.
TABLE 23-8 Representative Families of the Rosid Clade
An important character in rosids is the presence of pinnately compound leaves. This is believed to have been the ancestral condition. Although some living species have simple leaves, these apparently have arisen from compound leaves, perhaps by suppression of all leaflets except one. Whereas simple leaves are an early, ancestral condition for the angiosperms as a whole, they are a later derived condition for rosids.
Although this clade is named for the order Rosales, roses should not be considered typical; they are just one group of many (FIGURE 23-30). Because the rosids consist of 14 large orders with over 50,000 species, only a few can be mentioned here. Five of the orders contain almost 75 percent of the species: Fabales (legumes), 19,000 species; Myrtales (Eucalyptus and evening primrose), 9,000; Malpighiales (poinsettia), 1,600; Rosales (roses, elms, marijuana), 6,300; and Sapindales (maples, horse chestnuts, creosote bush, and the species whose resins are valued as frankincense—Boswellia—and myrrh—Commiphora), 5,800. Members of this subclass include roses, of course, and legumes (peas, beans, peanuts, Mimosa, redbud, and clover; Fabaceae = Leguminosae); Fuchsia, evening primrose (Onagraceae); dogwood (Cornaceae); the spurges that look like cacti and often have an extremely poisonous milky latex (Euphorbiaceae); maples (Aceraceae); and dill, celery, carrot, parsley, and hemlock (Apiaceae = Umbelliferae).
The rose family is important not only in an evolutionary sense but also economically. Rosaceae contains numerous ornamental genera, including Rosa (roses), Crategus (hawthorn), Spiraea, Cotoneaster, Pyracantha, Photinia, Potentilla, Chaenomeles (flowering quince), and Sorbus (mountain ash). The family also provides most of the fruits that can be grown in temperate climates: Malus (apple), Pyrus (almond, apricot, cherry, nectarine, peach, plum, and prune), Eriobotrya (loquat), Fragaria (strawberry), and Rubus (blackberry, loganberry, and raspberry).
Fabaceae (the legume family: beans, peas, lentils, and peanuts) is also a large family having bodies that vary from herbaceous annuals to shrubs, vines, and long-lived trees. It is important economically as a source of many foods, drugs, dyes, and woods, and many of its species are the dominant plants in arid areas and deserts. Many species have root nodules with symbiotic associations with nitrogen-fixing bacteria, and thus, they can grow in poor soils yet still produce protein-rich seeds. Legumes are a critically important source of proteins for people in arid areas.
Asterid Clade
The most derived large clade of eudicots is the asterid clade (TABLE 23-9), which contains plants such as sunflower, periwinkle, petunia, and morning glory. Asterids, being a sister clade of rosids, originated perhaps as recently as 60 million years ago, and even its most basal members were much more highly derived than plants in the basal angiosperms. The majority of asterids can be easily distinguished from other angiosperms on the basis of three features: (1) They have sympetalous flowers (their petals are fused together into a tube); (2) they always have just a few stamens, not more than the number of petal lobes; and (3) stamens alternate with petals (FIGURE 23-31). Asterids exploit very specialized pollinators that recognize complex floral patterns, and such plants could not evolve before derived, sophisticated insects appeared.
Many chemical differences exist between this group and all others. It lacks many specialized chemicals found in other clades: It has no betalains (present in core Caryophyllales), benzyl-isoquinoline alkaloids (magnoliids), ellagic acid (FIGURE 23-32), or proanthocyanins (rosids). Instead, many asterids have iridoid compounds, which occur only rarely outside this group. Certain families produce numerous, very potent chemicals that deter animals or kill them outright. Apiaceae (celery, dill, and fennel) have secretory canals containing oils and resins, triterpenoid saponins, coumarins, falcarinone polyacetylenes, monoterpenes, and sesquiterpenes. The poison Socrates was given to kill himself came from Conicum (hemlock) in the Apiaceae. It is not important for you to learn this list or understand what these chemicals are, but rather to see that in addition to new morphological, anatomical, and reproductive characters originating evolutionarily, new defensive chemicals also arise.
Currently, asterids have the greatest number of species (approximately 60,000), but they are grouped into two small orders (Cornales, Ericales) and two groups of orders, lamiids (also called euasterids I) and campanulids (euasterids II) (FIGURE 23-33). One family (sunflowers, daisies; Asteraceae [old name is Compositae]) contains fully one third of all the species and is the largest family of eudicots. Examples of asterids are milkweeds (Asclepiadaceae); potato, tomato, red peppers, eggplant, tobacco, deadly nightshade, and petunia (Solanaceae); morning glory (Convolvulaceae); thyme, mints, and lavender (Lamiaceae); and Asteraceae with sunflowers, dandelions, lettuce, Chrysanthemum, ragweed, and thistle (FIGURE 23-34).
Many asterids are extremely important medicinally. Apocynaceae (oleander family) contains periwinkle, Vinca, from which are extracted vinblastine and vincristine, two of our most potent anticancer drugs. Rubiaceae contains, in addition to coffee (Coffea), Cinchona, from which we derive the antimalaria drug quinine. Another rubiaceous genus, Cephaelis, provides us with ipecac, an emetic used to induce vomiting in cases of oral poisoning. Heart disease is treated with cardiac glycosides extracted from Digitalis (Scrophulariaceae, the snapdragon family); these compounds make heart muscle beat more slowly and strongly, increasing output of blood from the heart and improving circulation.
The order Lamiales (families of olive, Penstemon and snapdragon, gesneriads, acanths, and trumpet-creeper) is quite derived in its floral characters; the flowers tend to be bilaterally symmetrical and of sizes and shapes that permit only certain insects to enter and effect pollination. The floral bilateral symmetry further forces the insect to enter in one particular orientation. Pollinators capable of doing this tend to be sophisticated and able to recognize different flowers easily. They learn which flowers provide the greatest rewards and then search for other flowers of identical shape, color pattern, and fragrance, thus providing efficient pollination for the plant species.
The giant family Asteraceae (asters, daisies, and sunflowers) contains 1,100 genera and 20,000 species distributed worldwide in almost all habitats except dense, dark forests. They range from important food plants (Lactuca—lettuce) to ornamentals to weeds. The characteristic daisy/sunflower type of inflorescence makes them instantly recognizable. Members of Asteraceae have a wide range of unique chemical defenses against herbivores: sesquiterpene lactones, monoterpenes, terpenoids, and latex canals that contain polyacetylene resins (FIGURE 23-35). The presence of these chemicals makes composites extremely resistant to animals that eat plants or lay eggs in them; it also causes them to be irritating to human skin, resulting in numerous cases of contact dermatitis. The Asteraceae is a young family, perhaps no more than 36 million years old, which probably originated in the Oligocene Epoch of the Tertiary Period.

FIGURE 23-30 Rosids. (A) Hibiscus. (B) St. John’s wort (Hypericum spathulatum). (C) Immature gourd fruit (note the withered petals). (D) Theobroma cacao, the source of chocolate. Flowers and fruits are borne on the trunk and large branches, not on twigs. The seeds are harvested, fermented, roasted, and ground into cocoa powder. (E) Lupine (Lupinus latifolius). (F) Mexican plum (Prunus mexicana). (G) Strawberry (Fragaria).
TABLE 23-9 Representative Families of the Asterid Clade

FIGURE 23-31 (A) In asterid flowers, such as Digitalis purpurea, all petals fuse to each other and form a tube; only certain pollinators can fit into the tube far enough to obtain nectar—and inadvertently carry out pollination. (B) Flowers of potato (Solanum tuberosum) also have fused petals, but they are so open that we can see that stamens alternate with petals.
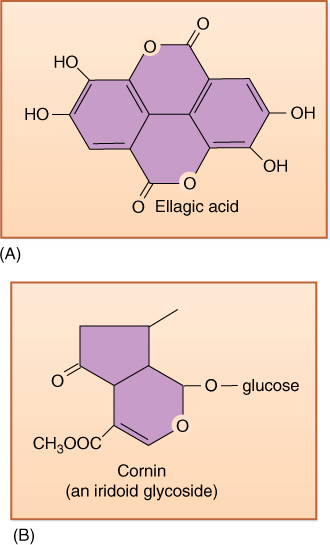
FIGURE 23-32 (A) Ellagic acid is common in several eudicot clades but absent from one of the most derived, the asterids. Its presence deters many insects from eating the plants, but by approximately 50 million years ago, many types of insects had evolved a tolerance to it. (B) Many asterids produce iridoid compounds such as this cornin. They are a relatively new class of chemical defense compounds, and thus, few insects tolerate them; iridoid compounds may be partly responsible for the success of the asterids by keeping herbivory to a minimum.
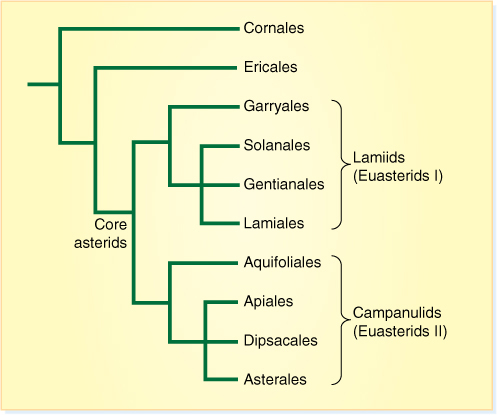
FIGURE 23-33 One recently proposed cladogram of asterids.

FIGURE 23-34 Asterids. (A) Potato (Solanum tuberosum). (B) Milkweed (Asclepias viridis). (C) Goatsbeard (Tragopogon in Asteraceae). The “flowers” of asters are really inflorescences, groups of many true, small flowers, called florets. Each outer “petal” is a ray floret (each with one greatly enlarged petal), and each central flower is a disk floret. The dark cylinders are anthers that have fused around styles. The innermost disk florets have not yet opened. (D) Flowers of Cavendishia (Ericaceae) show the derived feature of sympetally—petals fused together as one unit, except at their tips. (E) In Hydrangea, what appear to be large petals are really bracts located below the tiny flowers.
Plants and People
BOX 23-2 Vegetarianism: Alternatives to Eating Meat
Our ancestors evolved to be omnivores. As a result, we modern humans can eat seeds, leaves, and fleshy roots because our flat molars allow us to grind and mash them before we swallow them, and our small intestine is so lengthy it ensures that plant matter remains inside us long enough to be digested. Similarly, we can eat meat because our incisor and canine teeth allow us to rip meat from bones (think of barbequed ribs, fried chicken, and pork chops) and our digestive system produces plenty of protein-digesting enzymes.
However, just because we have the ability to eat meat does not mean we must eat it. A large number of people have adopted some form of vegetarianism to reduce the amount of meat in their diet or eliminate it altogether. There are several types of vegetarian diet:
✵ Ovo-lacto vegetarians (often just called vegetarians) eat a diet based almost exclusively on plant products such as fruits, vegetables, greens, and seeds (in the form of foods like bread, pasta, and oatmeal) with some eggs and dairy products (milk, cheese, ice cream, and yogurt) (FIGURE B23-2). The concept here is that animals do not have to be killed to obtain eggs and milk. For some vegetarians the only acceptable animal-based food is eggs, for others only milk-based foods, and for others even fish and seafood is acceptable.
✵ Vegans avoid all foods obtained from animals, including meat, eggs, and all dairy products. Strict vegans also do not wear clothing made from animals, such as leather belts, shoes, and coats or anything made with fur or turtle shell.
✵ A fruitarian diet consists of only fruits, nuts, and seeds, items that can be gathered without harming a plant. Fruitarians will not eat roots such as carrots, beets, and cassava because those can be obtained only by killing the plants.
✵ Flexitarians have a flexible approach to meat and other animal products in their diet; they eat mostly plant foods and merely reduce the amount of meat they eat. For example, flexitarians might set the goal of having meat-free lunches or designating several days of the week as being meat-free. The advantage of a flexitarian approach is that a person can succumb to the occasional hamburger or steak without feeling they have failed and might as well give up.
There are several reasons for reducing or eliminating animal products in our diet and our daily lives. Many people do so for ethical reasons: They do not want to be responsible for animal suffering or death. There is a range of empathy here. Red meat is muscle tissue that can only be obtained by killing cattle. Cattle are mammals like we are, and they definitely feel pain and suffering. Obtaining milk from cows, goats, or other mammals (only mammals produce milk) does not involve killing them, so there is little or no suffering in many dairy operations. Gathering eggs from chickens or other birds also does not involve harming them, although some poultry farms keep the animals crowded together in poor conditions and clip their beaks so they won’t peck each other. Eating fish and seafood involves killing the animals, but many people feel little emotional connection to fish, shrimp, and clams.

FIGURE B23-2 This is an ovo-lacto vegetarian meal because it contains eggs and cheese in addition to plant-based foods. A dessert of pie and ice cream fits into such a diet.
When choosing your diet for ethical reasons, there are various issues to consider. Raising cattle in the United States and elsewhere is often cruel. Most cattle are raised on factory farms, crowded into large, dirty feedlots where the animals must stand in their own feces; they are given feed that includes antibiotics, growth supplements, and even the ground up debris from cattle that were slaughtered earlier. When it is time to kill them, the cattle are poked with prods that give them an electrical shock, forcing them into a line into the bright, noisy slaughterhouse (cattle are terrified by bright lights, noise, and having people moving around them, especially above them). Laws in many countries prohibit the slaughter of any animal that is not healthy enough to walk into the slaughterhouse, but that law is sometimes violated. PETA (People for the Ethical Treatment of Animals; www.peta.org) has developed a set of guidelines for treating cattle more humanely, including giving them uncrowded pastures for grazing and safe food, and when they are to be killed the slaughter house should be quiet, dimly lighted, and with as few people as possible. The animals are still killed, of course, but at least their life and death were not miserable. For many people this humane treatment reduces their ethical objections and they will eat meat occasionally. To be certain the meat you eat has come from humanely treated animals, it is best to shop at reputable health food stores or organic food stores; don’t be afraid to ask about the treatment of the animals.
Veal is especially objectionable because it is the flesh of calves that are killed while still so young they have never eaten anything other than their mother’s milk. Often, the calves are kept in cages so small they cannot move because movement would cause their muscles to develop, making the meat less tender. The calves also may be kept in the dark to keep them quiet. It is difficult to justify treating animals like this just to have a particular kind of dinner.
Eggs and milk do not require the slaughter of animals, and therefore many people do not believe it is unethical to consume these products. Here too it is best if the chickens and cattle are treated humanely and are allowed access to open pastures or pens with fresh air and water. People with the strictest ethical standards try to avoid all animal products because they believe it is unfair for us to breed and maintain animals just for our own use. It is difficult, however, to eliminate the use of all animal products: Sugar is whitened by filtering it through the ash of animal bones, most cheese is made with rennet from calves’ stomachs, clear juices (apple and grape) and alcoholic beverages are clarified with gelatin from animal hides and bones, and so on.
Other people avoid animal products for economic and ecological reasons. When we feed animals corn, soybeans, oats, and other feeds that we ourselves could eat, we are eating at a high trophic level. The plants are primary producers, the livestock animals are primary consumers, and we are secondary consumers. Typically, it takes 10 pounds of food at one trophic level to make 1 pound of food at the next level. Consequently, we need to feed cattle or chickens 10 pounds of oats just to get 1 pound of eggs or milk or meat. The other 9 pounds are lost as carbon dioxide and methane (both greenhouse gasses), urine, feces, and waste. If we ate the plants ourselves, we would need to farm only one tenth as much land as we use now, producing much less sewage. It is estimated that as much as 70% of all the wheat, corn, and other grains we cultivate are used to feed livestock. Also, huge areas of tropical rainforest have been and are being cut down and converted into pasture for cattle just to produce hamburger. Reducing the amount of animal products we eat is an important aspect of a philosophy of living more simply, of trying to minimize the impact each of us has on our environment.
An important consideration for anyone considering a vegetarian diet is whether it is healthful. The answer is easiest for a flexitarian diet: Virtually all studies show that diets low in meat and high in fruit and vegetables are much more healthful than ones with more meat. And it is now well established that even a strict vegan diet provides all the nutrients we need for a healthy life. We need to be especially careful to obtain adequate amounts of vitamin B12, which is not found in any plant but which can be obtained as a supplement or by consuming yeast products. Many people believe that athletes and bodybuilders need a high protein intake and must have meat, but that has been shown repeatedly to not be true. Most of us Americans consume much more protein than we need, and the plants of a vegetarian or vegan diet provide adequate amounts for anyone. We must be careful to obtain adequate amounts of all eight essential amino acids. All plants, animals, fungi, and other organisms construct their proteins from the same 20 amino acids, and we humans can synthesize 12 of them within our own bodies, but our metabolism cannot make 8 of them. Dairy and egg foods supply adequate amounts, and so do the proteins of soy, amaranth, quinoa, and several other seed plants. The proteins of many food plants such as wheat, rice, beans, chickpeas, and others have adequate amounts of 18 or 19 amino acids but only sparse amounts of 1 or 2 others. We can avoid having a deficiency by eating what are known as complementary foods, foods in which one member has plenty of an amino acid that the other member lacks, and vice versa. For example, a combination of brown rice and beans, beans and corn, tofu (soy) and rice, or one of hummus and wheat pita bread provides adequate amounts of all 20 amino acids. Think about a typical Mexican meal that includes beans and corn tortillas; these two plant foods together provide all essential amino acids. The same is true of a typical American lunch: A peanut butter sandwich (peanuts and wheat) provides complementary proteins.
The take-home message here is that we should eat less meat and animal products. Both our bodies and our environment will be healthier.
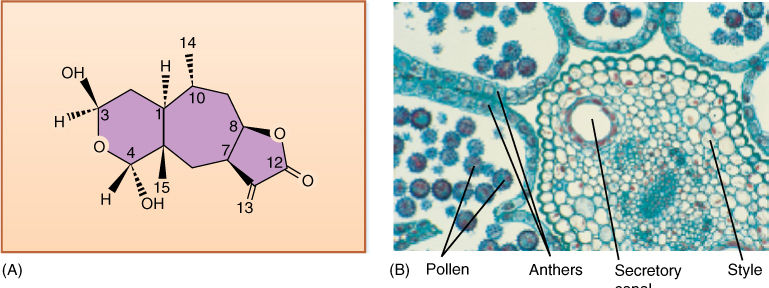
FIGURE 23-35 (A) Hymenoxon is a sesquiterpene lactone, a member of the chemical arsenal of asterids. This particular compound occurs in bitterweed (Hymenoxys odorata) of the southwestern United States; the plant is eaten by sheep only during droughts when no other plants are available. Sesquiterpene lactone causes hemorrhaging of all internal organs of the sheep. (B) Many members of Asteraceae contain secretory canals lined with cells that produce a variety of toxic compounds. This canal is in the style of sunflower (Helianthus).
 At the Next Level
At the Next Level
1. Focus on a monocot group. The angiosperms are such a large, diverse group that it is difficult to become familiar with all of them unless you spend your entire life studying them. Fortunately, you do not have to know them all. But I do recommend selecting a particular group and studying it at the next level. I recommend the Alismatales (Table 23-3) because these are unusual: Most have become adapted to living in water and many could be mistaken for mosses or algae until you look closely. You already know a great deal about typical angiosperms; the Alismatales (especially Hydrilla, Najas, and Ruppia) will surprise you at how extreme angiosperm modifications can be yet still be understandable.
2. Focus on a eudicot group. For this I recommend the rose family, Rosaceae. You already know many of its species (roses, apples, cherries, strawberries), so read even more about these: You know apple fruits but are you familiar with the tree? You know strawberry fruits but what about the plant? Would you ever guess that an herbaceous strawberry plant is related to a woody apple tree? This family has evolved by natural selection to have many, many growth forms, and we have applied artificial selection to alter the flowers, fruits, and vegetative bodies even further.
3. Wheat, rice, corn: Grasses keep us alive. The grass family Poaceae has hundreds of species, and many have been domesticated as foods. Wheat, rice, and corn are familiar examples of plants we rely on; almost every person on Earth eats one of these grasses every day. There are many other grasses that have been domesticated as well. Research the species we cultivate and learn about the benefits and problems of each. Why do we cultivate more wheat than rye (and does this vary with weather conditions)? We probably drink more barley than wheat; why?
SUMMARY
1. All flowering plants, members of division Magnoliophyta, have closed carpels, long pollen tubes, double fertilization, and, with very few exceptions, sieve tubes and vessels.
2. Division Magnoliophyta is almost universally believed to be monophyletic; all of its evolutionary lines can be traced back to a single common ancestor. The earliest fossils of leaves and wood date to approximately 125 to 130 million years ago, and fossil pollen from earlier periods has been found.
3. The earliest angiosperms may have been weedy shrubs that grew in areas not conducive to fossilization.
4. Early characteristics of angiosperms are believed to be flowers with many sepals, petals, stamens, and carpels arranged in spirals; uniaperturate pollen; radially symmetrical flowers; insect pollination; and wood without vessels. Later, tracheid-like vessels with scalariform perforation plates arose. Early angiosperms are believed to have been perennial; none was annual or herbaceous.
5. Some major evolutionary modifications that have occurred in many groups are reduction in the number of each flower organ; imperfect flowers; bilateral symmetry of flowers; nectaries and fragrances that attract only specific pollinators; wind pollination; various seed or fruit distribution mechanisms; biennial or annual herbaceous habit; wood with shorter, wider vessel members; larger amounts of wood parenchyma; and numerous types of antipredator compounds—perhaps two or three generations of these.
6. Division Magnoliophyta contains basal angiosperms, monocots, and eudicots.
7. Monocots can generally be recognized by their long, strap-shaped leaves; flower parts in threes; parallel leaf venation; numerous vascular bundles in the stem, not arranged in a ring; and no ordinary secondary growth.
8. Eudicots can generally be recognized by broad, not strap-shaped leaves; reticulate leaf venation; flower parts in fives or fours, but not threes; pollen tricolpate or tricolpate-derived; vascular bundles in the stem arranged in one ring; and woody growth or an annual or biennial herbaceous body.
9. In the classification presented here, monocots contain several orders and the commelinoid clade, and eudicots contain basal eudicots, rosids (with fabids and malvids), and asterids (with lamiids and campanulids).
IMPORTANT TERMS
angiosperm carpels
anthocyanin pigments
asterid clade
basal angiosperms
betalain pigments
campanulid order
closed carpel
commelinoid monocots
Division Magnoliophyta
double fertilization
eudicots
fabids
generalized flower
iridoid compounds
lamiid order
malvids
monocots
perisperm
pistil
primitively vesselless
ranalean flower
rosid clade
secondarily vesselless
septal nectaries
sympetally
tricolpate (pollen)
uniaperturate (pollen)
zygomorphy
REVIEW QUESTIONS
1. The earliest fossils clearly recognizable as parts of flowering plants are _____________________ __________________ in rocks more than ______________ million years old.
2. What transformations must have occurred if gymnosperms really were the ancestors of angiosperms? Which might have occurred earlier (are present in all relictual angiosperms) and which later (are absent from some of the most relictual angiosperms)?
3. If someone were to examine the stamens of most flowering plants, they would never automatically think of them as sporophylls because they have very little leaflike structure, but look at the four stamens in Figure 23-1. These are from flowering plants. What leaflike characters do they have? Are they flat and at least somewhat resemble the blade of a leaf? Do they have vascular bundles that branch, at least somewhat resembling the reticulate venation of a dicot leaf? Most dicot leaves have three vascular bundles entering their petioles. Which two of these stamens also have this leaflike character? Which two have only one vascular bundle?
4. During fertilization in flowering plants, one sperm cell nucleus fuses with the egg nucleus, producing the diploid zygote. The other sperm nucleus fuses with the ___________________________________, producing the triploid ________________________, which develops as temporary tissue that nourishes the embryo. This process is called ___________________ fertilization. What tissue nourishes the embryo in gymnosperm seeds?
5. Why were many modern genera of dicots initially thought to have come suddenly into existence about 130 million years ago? Why do we now think that those leaf fossils do not really belong to the modern genera they were named for?
6. What is meant by a closed carpel as opposed to an open cone scale?
7. Describe what the early angiosperms may have been like with regard to flowers, body, ecology, wood, and pollen. For each of the characters, what living angiosperms still have those characters? In what ways have those characters changed in certain lines of evolution (Hint: See Table 23-1)?
8. Concepts about the nature of the early angiosperms have changed (and may change again in the future). In the last century, members of the subclass Hamamelidae were considered the most relictual of the living flowering plants. What kind of pollination do these plants have—insect pollination, wind pollination, or bird pollination? Give the common names of four members of this group. Although these plants are woody trees like many conifers, their wood has three characters that wood of conifers does not have, so we no longer think that hamamelids are the most relictual flowering plants. What are the three characters?
9. About 100 years ago, C. E. Bessey developed the hypothesis of the “ranalean” flower. What are some of the characters of this type of flower? Is the ranalean flower wind pollinated or insect pollinated?
10. Do most botanists believe that the flowering plants are monophyletic, or do they believe that they are actually an unnatural group consisting of several lines of evolution that do not share a common ancestor?
11. Describe Amborella and water lilies. On the basis of morphology, would you conclude that they are closely related? What evidence is used to support the hypothesis that they are ancient lineages from basal angiosperms?
12. All plant families have a name that ends in “-aceae,” and some families also have old names, still commonly used, without this ending. The palms are a family with two names. What is the new name that ends in “-aceae,” and what is the older name? What kind of a compound leaf do the feather palms have? The fan palms? Why are palm flowers so seldom seen by most people?
13. Philodendron is a plant you may know. They and their close relatives are characterized by a distinctive inflorescence. It has tiny flowers embedded in a thick stalk called a __________________, and it is surrounded by a large bract called a _____________________.
14. What is the name of the grass family? About how many species does it contain? Name seven genera (and give the common names) of grasses that are used for food. What kind of pollination do grasses have—animal or wind?
15. Zingiberales contains some of the most familiar of all houseplants. Name several of these.
16. Liliaceae contains many familiar plants. Name six. Which closely related family contains the irises? Which contains yams?
17. The Orchidaceae is the largest and most diverse family of monocots. What is the most common orchid habit (growth form)—being an epiphyte or being terrestrial? Are orchid flowers bilaterally symmetrical or radially symmetrical? Describe orchid seeds.
18. Certain eudicot clades contain plants with many relictual features. Name and describe several.
19. Core Caryophyllales contains many familiar plants. Can you list four members of this group? The group is unusual in having _______________________________ pigments rather than anthocyanins in their flowers. Another feature of this group is that endosperm develops little or not at all, and instead, the seeds have ______________________, which develops through proliferation of cells of the ____________________.
20. Give the name of the families of each of the following:
a. Camellias
b. Chocolate
c. Cotton
d. Violets
e. Pumpkins and watermelons
f. Cranberries and blueberries
21. One important character in rosids is the presence of ___________________ ___________________ leaves. The subclass is named for the rose family, Rosaceae, and this one family has many genera that produce fruits that we like to eat. Name the genera of the following fruits, all in the rose family:
a. Apples
b. Almonds, apricots, cherries, peaches, and plums
c. Strawberries
d. Blackberries and raspberries
22. Textbooks often use cacti and euphorbias (spurges) to illustrate convergent evolution because many euphorbias have globular, succulent bodies that strongly resemble those of cacti, yet the two families are not closely related. What are the names of the euphorbia family and of the cactus family? Which major clade is each in?
23. Name the families that contain the following species:
a. Sunflowers, dandelions, and daisies
b. Milkweeds
c. Potato, tomato, and tobacco
d. Mints and lavender
e. Coffea (coffee)
24. Think about the Coniferophyta and the Magnoliophyta of this chapter. Which has more families, genera, and species? Which has more diversity of body types? Which has species in the greatest number of habitats (deserts, forests, rain forests, underwater, grasslands, marshes, epiphytic habitats, and so on). Which group originated earlier in evolution, and which originated later? The older group has had more time to diversify. Is it the more diverse of the two?
Design Credits: Hummingbird: © Tongho58/Moment/Getty; Green Plant Cells: © ShutterStock, Inc./Nataliya Hora; Purple Tulip: © ShutterStock, Inc./Marie C Fields; Dandelion: © ShutterStock, Inc./Danielkreissl; Poppy: © ShutterStock, Inc./Saruri; Plant Icon: © ShutterStock, Inc./Vector; Digging Man Icon: © ShutterStock, Inc./Z-Art