Botany: An Introduction to Plant Biology - Mauseth, James D. 2017
Preface
Overview of Plant Life

Chapter Opener Image: The life of each of these plants has much in common with our own lives. Each started from a fertilized egg, grew into an adult by obtaining energy and nutrients, as well as defending itself from diseases and pests. Each inherited its genes from its parents and each will make efforts to have sexual reproduction. Like us, these plants have gradually evolved such that they now differ from their ancient ancestors. Also like us, each plant interacts with many other organisms, some of which help it, some of which harm it; no plant lives in isolation from all other organisms.
OUTLINE
✵ Concepts
✵ Overview of Plant Structure
✵ Overview of Plant Metabolism
✵ Overview of Information in Plants
✵ Overview of Plant Diversity and Evolution
✵ Overview of Plant Ecology
Box 2-1 Alternatives: Familiar Plants and Some Confusing Look-Alikes
Box 2-2 Alternatives: Plants Without Photosynthesis
Box 2-3 Plants and People: Toxic Compounds
Box 2-4 Botany and Beyond: Noah’s Flood and Population Biology
LEARNING OBJECTIVES
After reading this chapter, students will be able to:
✵ Describe the basic internal and external organization of plants.
✵ Compare plant and human metabolism.
✵ List three sources from which plants receive information.
✵ Define and give examples of haploid and diploid plants.
✵ Explain the concept of clades.
✵ Describe the interconnected dynamic between photosynthetic organisms and organisms that respire.
 Did You Know?
Did You Know?
✵ Plants, as distinct from algae, have existed for more than 420 million years.
✵ Photosynthesis captures energy in sunlight and is almost the only source of energy for all life on Earth.
✵ There are almost 297,000 species of living plants and about 1,260,000 species of animals.
✵ Plants are adapted to live everywhere on land in extraordinarily diverse conditions except where it is severely cold (under permanent snow and ice) or dry (parts of the Sahara Desert).
✵ Plants defend themselves from herbivores with spines and poisons but simultaneously provide food for animals that pollinate them.
![]() Concepts
Concepts
Anyone reading this book is probably familiar with animal life. We each know quite a bit about our own body, our organs, our metabolism, and the way in which we inherited DNA from our parents. We know that all humans are members of one extremely diverse species, Homo sapiens, a species in which individuals differ from each other in almost every possible character such as height, athletic ability, intelligence, health, talent, interests, goals, ambitions, spirituality, and more. We know that we had ancient ancestors, hominids who preceded the first Homo sapiens, and we had ancient relatives such as Homo neanderthalensis (Neanderthal Man) and Homo habilis. These other members of Homo are extinct now, but several distant relatives such as chimpanzees, orangutans, and gorillas are part of our evolutionary family. Most of us are probably not too familiar with our own ecology except that we are social animals (we live in cities and do things in groups), we consume a large part of Earth’s resources, and produce harmful pollution.
Beyond our knowledge of our own biology, most of us are also familiar with the biology of other animals such as dogs, cats, mosquitoes, bees, whales, and so on. Even if this is the first biology book you have ever read, you are almost certainly a knowledgeable biologist already.
Your knowledge of animal biology is an excellent foundation for learning about plant biology. Plant life is based on all the same fundamental principles that underlie the lives of animals, fungi, and all other living beings. Of course, plants do many things differently, but let your knowledge of your own biology guide your study of plant biology. And always keep the two fundamental questions in mind: What are the alternatives? What are the consequences of each alternative?
![]() Overview of Plant Structure
Overview of Plant Structure
Think about the many types of plants you know. Trees, garden flowers, grass, bulbs, cacti, and vines for example. Tremendous diversity is present, but if we examine each of these, we find they all share the same pattern of body organization. With only a few exceptions, the body of every plant is organized in the following simple way.
Almost every plant consists of one or several stems, each of which has leaves attached at regions called nodes (FIGURE 2-1; TABLE 2-1). Just above the attachment point is an axillary bud, some of which develop into branches whereas others develop into flowers, but many axillary buds remain dormant and never do anything. Stems are typically slender, less than 5 mm in diameter, but wide ones occur in cacti and other desert plants that use them to store water (FIGURE 2-2). Some stems are extremely short, as in carrots and cabbage, but vines and bamboo typically have extremely long stems.
Leaves too are diverse. They range in size from the gigantic leaves of palms to ones that are merely large, as in Philodendron, down to ones that are small, such as those of rosemary (FIGURES 2-3, 2-4). Although cacti are often said to be leafless, all have tiny foliage leaves that are almost microscopically small. Leaves vary greatly in shape and texture and in being evergreen or deciduous, but the important thing is that almost all stems bear leaves; they are a part of the fundamental pattern of plant bodies. Hornworts and some liverworts are plants that never produce any kind of leaf at all (see Chapter 20 for more details).

FIGURE 2-1 Shoot tip of cottonwood (Populus deltoides) in late autumn. It has a central stem with 15 leaves attached (only the petioles are visible in this close-up view); each petiole attaches to the stem at a node. Five nodes each contain one prominent axillary bud, each covered with protective bud scales. The very tip of the shoot has a terminal bud. Bud scales protect the delicate cells at the center of each bud; after surviving the winter, these five axillary buds will grow and produce many small flowers. In springtime, the terminal bud will grow as a continuation of the shoot, producing more leaves, nodes, and internodes, making the branch longer.
Another aspect of the fundamental organization of plants is that almost all plants have roots (FIGURE 2-5). In almost all plants, roots are confined to one end of the stem, but it is also common to see roots emerge along a stem, such as in ivy that climbs on fences and trees, attached to them by numerous groups of roots (FIGURE 2-6). Underground stems like those of irises and ginger also produce roots along their length rather than at just one end (FIGURE 2-7). Roots are less varied than stems or leaves, and most roots are usually easy to recognize as roots.
TABLE 2-1 The Organs that Compose Most Plants
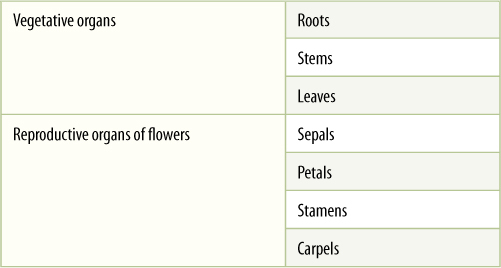

FIGURE 2-2 Like many plants adapted to deserts, golden barrel cactus (Echinocactus grusonii) stores water in very broad stems composed of thousands of water-filled cells. Spines prevent animals from eating the cactus to obtain water. Because the stem is so broad, it is exceptionally heavy, so succulent plants typically branch much less than non-succulent plants: This plant has only one stem, somewhat like the single stem of a palm tree. The spines are highly modified bud scales (compare with the bud scales of cottonwood in Fig. 2-1), so each spine cluster is an axillary bud. The leaf that makes the axil for each spine cluster is microscopic and cannot be seen here.
Roots, stems, and leaves are the vegetative organs of a plant. By that, we mean that they are responsible for obtaining energy and materials for growth and survival, but they do not carry out sexual reproduction. When a plant is ready to reproduce, reproductive organs develop. The reproductive organs of most ferns are barely distinguishable from vegetative organs: They are basically just foliage leaves that produce sporangia on their underside (FIGURES 2-8). Without turning a leaf over to see if it has sporangia and spores, one would never guess that it is not just an ordinary leaf. Reproductive structures are a bit more complex in the seed plants. Conifers make seed cones and pollen cones, angiosperms make flowers (FIGURES 2-9, 2-10). Only flowers will be discussed here in this overview; details about conifers and ferns are presented in later chapters.

FIGURE 2-3 Like many palms, Bismarkia nobilis has gigantic leaves. Each petiole is more than 1 m long, and the giant leaf blades are both long and wide. An individual palm usually has many fewer leaves than do other trees, but because each palm leaf is so large, palms have as much total photosynthetic surface area as do many other plants. For some unknown reason, many people refer to the leaves of palms and ferns as “fronds” even though they are true leaves.

FIGURE 2-4 This is a very common weed in most gardens, and you may have already spent some time pulling it up. It is in the genus Chamaesyce, and plants tend to grow flat against the ground, often with a circular form only a few centimeters in diameter. These leaves are tiny, only about 1 mm across. The plant is covered in flowers, plants of Chamaesyce bloom even while only a few weeks old; each flower here is emerging from an axillary bud.
Flowers appear to be so diverse it seems as if there must be thousands of types. Just think of how diverse roses, irises, lilies, snapdragons, and petunias are. But here too there is only one basic organizational plan, and once you know it, you can easily understand almost any flower. Each flower is a reproductive shoot; that is, each flower has a short stem with leaf-like structures attached to it. Sepals protect the rest of the flower as it develops, and petals attract pollinators after the sepals spread apart allowing a flower to open. Above the petals are stamens that produce pollen, followed by carpels that each contain one or more ovules (FIGURE 2-11). Stamens and carpels are essential because each pollen grain contains two sperm cells and each ovule contains an egg cell. Insects, birds, and wind carry pollen from stamens to carpels, usually from the stamens of one plant to the carpels of a different plant of the same species. After a sperm cell fertilizes an egg cell, an embryo and the surrounding tissues grow to be a seed while at the same time some carpel tissues develop into a fruit. In angiosperms, all seeds develop inside a fruit—that is one of the defining characters of being an angiosperm (FIGURE 2-12).

FIGURE 2-5 These onions are whole plants, with roots at one end, the shoot at the other. Onion leaves are a bit unusual because they are part of a shoot called a bulb: The purple portion of each leaf (the leaf base) is swollen and stores water and nutrients, the green upper portion carries out photosynthesis.
Alternatives
BOX 2-1 Familiar Plants and Some Confusing Look-Alikes
Most of us have no trouble recognizing a very large number of plants, at least while they are in bloom or if certain characteristic features are present. Apples, oranges, peaches, and pears are fruits we all recognize, and if they are still on their trees, we have no trouble recognizing the whole plant as well (FIGURE B2-1A). Strawberries are very familiar, but you might be initially uncertain if you saw them growing on the soil surface, supported by thin, delicate stems (such stems are runners). Daisies (often called asters) are also usually easy to recognize because even though there are hundreds of species, most all look like the one in FIGURE B2-1B. The same is true for morning glories (FIGURE B2-1C), lilies, water lilies, and many others (FIGURE B2-1D).
In contrast, several groups of plants can cause confusion. Many people believe any plant that is succulent and spiny is a cactus (FIGURE B2-1E), whether they are looking at an Agave (FIGURE B2-1F), a Yucca (FIGURE B2-1G), an Aloe (FIGURE B2-1H), or one of the succulent spurges (“spurge” may be an unfamiliar name and refers to members of the genus Euphorbia [FIGURE B2-1I]). These are all succulent, most are spiny, and most live in deserts, but agaves and yuccas are much more closely related to lilies and irises than to cacti (their long, thick leaves are extreme versions of lily leaves). Furthermore, although many botanists also mistakenly call spurges cacti, the spurges are in a completely different family and are mostly native to Africa, whereas cacti are native to the Americas. I heard a person guess that a giant Agave americana must be an Aloe vera: He made a really good guess because the plants do look alike, even though that agave is hundreds of times larger than an aloe.
Does it really matter if people confuse these plants? Not really, because they are correct that the plant is a large, succulent spiny desert plant. If it is important to have the correct name, they can ask at a garden center or a botany department or perform a Google image search.
Other plants that are often confused with each other are roses and camellias (the flowers are quite similar; FIGURE B2-1J) and palm trees, cycads, and ferns (some have similar shapes and leaves; in fact, in areas where cycads are common, people often use cycad leaves instead of palm leaves on Palm Sunday; FIGURE B2-1K). People often describe most aquatic plants as “moss,” but true mosses almost never grow in water: Long stringy ones that float are usually algae; those with stems, leaves, and flowers are flowering plants; and a few very small water plants are ferns. Many of us (even professional botanists) use the word “moss” for any very tiny plant whether it is a moss or just a small plant that resembles a moss (FIGURE B2-1L and FIGURE B2-1M).
“Grass” is used correctly almost all the time but applied to the wrong groups sometimes. Lawn grasses are grass (except for clover), as are wheat, rye, barley, rice, corn, and even giant bamboo (no, it is not a tree; it is a giant, very tough grass). If you see a grass-like plant growing in standing water, however, it is almost certainly a sedge and not a grass (FIGURE B2-1N). Sedges are distinguished from grasses by feeling the stems and remembering this mnemonic: “Sedges have edges, but grasses are round from tip to ground.” Using the wrong name is not a terrible problem; few of us know the names of very many plants, and as we discuss plants with our friends, using exactly the right name is not often necessary. It is like using the word “bug” for spiders, insects, and various other tiny animals.
There is a similar situation with the words we use for plant parts. You will often hear the word “frond” when people talk about the leaves of palms or ferns. “Frond” is just fine, but it is not a precisely defined term; it is just that for some reason, some people use it instead of the word “leaf” for palms and ferns. Similarly, “stalk” is not a precisely defined, technical term, and neither are “rind,” “peel,” “skin,” and several others. Just keep in mind that some precise botanical terms such as “stem” and “leaf” are the same as ordinary English words, and some like “epidermis” are identical to the terms used by zoologists, even though a plant epidermis is completely different from an animal epidermis. If you have already studied a lot of zoology, be very careful as you come across botanical words that are the same as zoological or medical words: The meanings are probably very different.
Plants have common names and scientific names, and a great deal of effort is made to ensure that each species of plant or animal or any other organism has only one scientific name. For some plants the name of the plant’s genus is the same as its common name; agaves are in the genus Agave, and citruses are in the genus Citrus. When we write the common name, we do not capitalize or italicize it, but we do if we are referring to the scientific name of the genus: oranges, lemons, and limes are all citruses in the genus Citrus. Lemons are Citrus limon, oranges are Citrus sinensis, and limes are Citrus latifolia. It is natural to think of “limon” as the species name of lemons, but that is not correct; the species name is Citrus limon. The scientific name of all species always has two words: The first is the genus name and the second is the species epithet; therefore, the species name is the genus + species epithet. When I mentioned agaves above I used the common name agave, the genus name Agave, and the scientific name of the giant agave, Agave americana.

FIGURE B2-1 (A) An orange tree. (B) Daisies. (C) Morning glories. (D) Oxblood lilies. (E) All the tall or globe-shaped plants are cacti; those with thick gray, yellow, or green leaves are agaves. (F) Agave americana. (G) Yucca torreyi. (H) Aloes. I) A spurge (Euphorbia, not a cactus). (J) Camellia flower. (K) A cycad, often called a “sago palm.” (L) Selaginella peruviana (despite the species epithet this plant grows in Texas, not Peru; not a moss). (M) A liverwort (Frullania inflata, not a moss). (N) A sedge, not a grass.

FIGURE 2-6 This is an old vine of poison ivy (Toxicodendron radicans), climbing up a tree trunk, attached to it by clusters of adventitious roots. These roots mostly just attach to the tree’s bark, and the poison ivy also has ordinary roots at the base of the vine that absorb water and nutrients. Even though this part of the poison ivy plant is so old that it has shed its leaves, it is still extremely poisonous and should not be handled.

FIGURE 2-7 This a rhizome of ginger (Zingiber officinale) that was purchased at a grocery store, planted for a few weeks, and then dug up for examination. By planting the rhizome, we activated it and stems grew upward; new adventitious roots spread in all directions but would have soon turned downward. All new parts are white because plants cannot synthesize chlorophyll while in the dark. The new adventitious roots emerge from the new shoots: As each shoot grows upward toward the light, it makes its own particular supply of roots.

FIGURE 2-8 (A) This is the upper surface of a leaf of sword fern (Polystichum munitum), and it carries out photosynthesis. But in many ferns, some leaves also produce spores on their undersides, and it is necessary to turn the leaf over to be certain. (B) This is the lower surface of a sword fern leaf, and it is involved in reproduction. Each of the round structures is called a sorus (plural: sori; pronounced SOAR eye), a group of dozens of minute structures (sporangia) that each produce spores. Only some of the leaves of sword ferns have sporangia, other leaves would have nothing special when turned over.
If flowers have this fundamental organization of sepals, petals, stamens, and carpels, how can there be so many types of flowers? And especially so many flowers in which this organization is not easy to see? One reason is that in many flowers, one or two of these organs are extremely underdeveloped or even absent completely. If a flower is pollinated by birds or insects, then having brightly colored petals helps a flower to be noticed. But if pollen is carried by wind, then it is a waste of resources to produce petals: Wind-pollinated flowers often lack petals completely (FIGURES 2-13, 2-14). Similarly, certain plants produce clusters of tiny flowers and rather than making sepals for each flower, they just make one set of large bracts that protects the entire cluster; these flowers lack sepals. Stamens and carpels too can be missing from the flowers of certain species.

FIGURE 2-9 This is a seed cone of a pine tree (Pinus). It was pollinated many months ago, and now the cone is growing larger as are the many seeds inside it. Tough, hard cells and sticky resin protect the seeds as they grow. When the seeds are mature, the cone will expand enough to cause the scales to separate and allow the seeds to fall out.
In addition, flowers differ from each other because in some, the various organs are fused to each other whereas in other flowers, all organs remain separate and distinct. It is common to see all the sepals of a flower fused to each other such that they form a tube, and petals often fuse to each other (FIGURE 2-15). Such fusions almost never occur among leaves. We could go on listing flower modifications, but the important thing is that each and every type of flower, no matter how exotic, is just a modification of one basic plan.
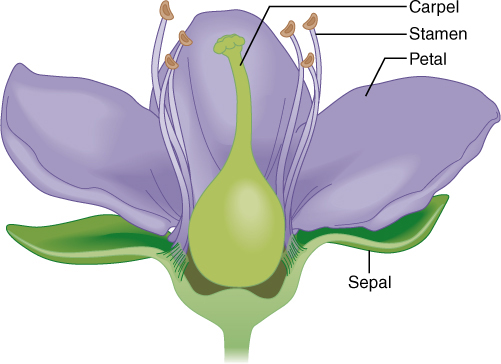
FIGURE 2-10 This is a simplified, diagrammatic representation of the four basic organs of flowers.
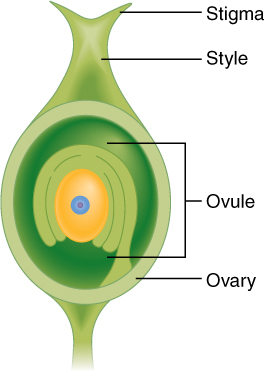
FIGURE 2-11 Carpels consist of three parts: a stigma, a style, and an ovary. Within the ovary is one or several ovules, each of which contains an egg cell (only one ovule is shown here). If the carpel is pollinated and the egg is fertilized, the fertilized egg develops into an embryo, the rest of the ovule develops into a seed, and the ovary develops into a fruit.
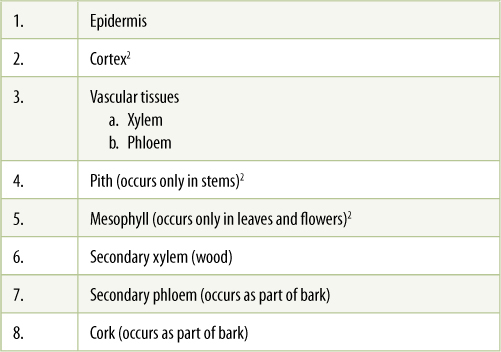
The internal organization of plants is even simpler and more uniform than the external form (TABLE 2-2). While young, every part of a plant—every root, stem, leaf, petal, and all the rest—consists of an external epidermis, internal ground tissues, and vascular tissues (FIGURE 2-16). Similar to our own epidermis, the epidermis of a plant protects the body from bacteria and fungi, and it inhibits water from evaporating out of the body. “Ground tissue” is a somewhat vague term that refers to inner tissues in general. The ground tissue in roots is cortex; in stems it is both pith and cortex; and in leaves and flowers it is referred to as mesophyll. Ground tissues of stems are easy to see in potato tubers: Each tuber is a short, wide stem that consists of very broad pith and a moderately wide cortex. Leaves of Aloe have a thick succulent ground tissue, the watery mesophyll.

FIGURE 2-12 A seed pod of coral bean (Erythrina herbacea) is a fruit similar to the pod of a bean or pea: When it is ripe, it opens by twisting, releasing the mature seeds. This is a dry fruit, in contrast to an apple or tomato, which are fleshy fruits.

FIGURE 2-13 These are three clusters of ash (Fraxinus) flowers; ash is wind pollinated, so there is no need for large colorful petals to attract animal pollinators. Each of these three clusters consists almost entirely of anthers, the part of a stamen that produces pollen.

FIGURE 2-14 We often never notice the flowers of wind-pollinated trees because they are very small, lack petals, and are located so high in trees. But after they release their pollen, the flowers are abscised from the trees and fall. Shown here are long, stringlike clusters of the staminate flowers of pecan (Carya illinoensis); although we often do not notice them while still in the trees, they are hard to miss when on sidewalks, cars, and roofs.

FIGURE 2-15 In this flower of Datura wrightii, the five green sepals have fused to each other, making a tube, and the five white petals have fused to each other, making a second tube. The tip of each petal remains separate and recognizable. The five stamens do not fuse to each other and are distinct. The five carpels have fused to each other, so only a single style and stigma are present.
Plants have two distinctly different vascular tissues. Xylem (pronounced ZY lem) transports water and minerals upward from roots to all aerial parts, especially leaves and flowers (FIGURE 2-17). The air around a plant absorbs most of that water but some of the water and all the minerals remain inside the plant for its own use. Phloem (pronounced FLOW em) carries sugars and various organic compounds as well as minerals and water throughout the plant. For example, leaves produce sugar by photosynthesis, and phloem carries most of that sugar out of the leaf into the stem; some is transported downward where it nourishes roots while some is transported upward to flowers and fruits. Xylem and phloem almost always occur together in a small number of vascular bundles.
TABLE 2-2 The Tissues that Compose Most Plants1
1 Reproductive structures such as flowers and cones have additional tissues; these are discussed in Chapters 9 and 22.
2 People often speak of cortex, pith, and mesophyll as tissues but technically each is a region of an organ, and each might be composed of various tissues. For example, leaf mesophyll is the region between the upper epidermis and the lower epidermis, and it consists of both photosynthetic tissue (the green cells) and vascular tissue (the leaf veins).
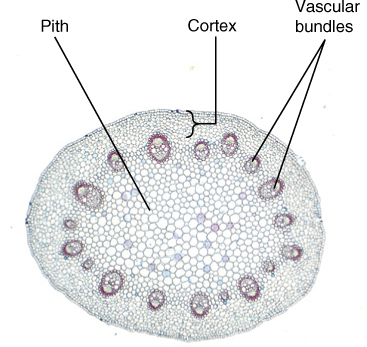
FIGURE 2-16 This cross-section of a buttercup (Ranunculus) shows an arrangement of tissues that occurs in almost all seed plant stems: epidermis, cortex, a ring of vascular bundles, and pith in the center.
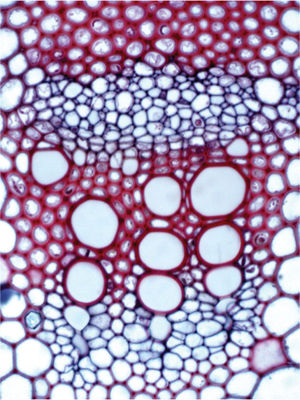
FIGURE 2-17 This is a light microscope view of a vascular bundle in a stem of sunflower (Helianthus annuus). The eight large round open cells at the bottom are xylem vessels that carry water upward from the root to leaves and flowers. Just above these are small, angular phloem cells that carry sugar. The upper mass of red cells are fiber cells that make a cap along the outer edge of the phloem, not only giving the stem strength to stand upright, but also to protect the phloem from insects that might try to eat into it or suck the sap through a long proboscis, as aphids do.
It can be confusing to compare the vascular systems of plants and animals without thinking about each carefully. We humans pump blood out of our heart into a single wide aorta, which then branches into many large arteries, each of which in turn branches into narrower arteries that finally branch into arterioles and capillaries. No plant ever puts all its water, sugars, and minerals into just one large vascular bundle. Even the most slender stem will have 10 to 20 vascular bundles; this provides plants with safety through redundancy. If an insect bites into a stem and breaks one or two vascular bundles, the stem will still have many others that are intact and functional (FIGURE 2-18).
The description of plant structure to this point refers to young plants as well as to young growth at the ends of branches and roots. This is called the primary plant body, and all the tissues and organs are produced by cells located at the very tips of stems and roots. Herbs are plants that have only this primary body; examples include lilies, lettuce, lawn grass, and ferns. Herbs are usually small plants that live only a few months before they reproduce and die. Woody plants are defined as those that produce a secondary body inside their primary body. This is easier to understand with less technical words: Woody plants are those that produce wood and bark (FIGURE 2-19). In an herb, all cells mature into epidermis, ground tissue, or vascular tissue; once these are formed, no immature cells are left. But in plants that will become woody, there is a sheet of cells that runs through all the vascular bundles; the cells within this sheet do not mature. Instead they divide and produce new cells; this region is the vascular cambium. Cells produced to the inner side of the vascular cambium become wood (technically called secondary xylem); those on the outer side become secondary phloem (which is part of the bark). All vascular cambia share the same basic organization, but can vary greatly in the speed with which they produce cells and in the amount of cells they produce. The vascular cambia of giant conifers and hardwood trees produce massive amounts of wood and bark, whereas those of roses and other shrubs produce much less. Woody plants also have cork cambia, sets of dividing cells that contribute waterproof, damage-resistant cork cells to the bark.

FIGURE 2-18 This is a cross-section of a small wisteria vine, and the many dark holes are vessels in the xylem. Each is the open end of a long tube that runs deeply into the stem, and the other part of each tube extended far into the part of the stem that was cut away. Even in this narrow young vine, water is carried by hundreds of vessels and if one or two become damaged, the rest can still function. Each year, the vine would add even more vessels as it made more wood along the edge of this wood, just interior to the bark.

FIGURE 2-19 It is easy to distinguish woody plants from herbs here: The trees are woody plants, the grasses are herbs. Even though grasses may live for many years, they never form secondary xylem or phloem (they never make wood or bark), whereas almost all trees begin to add secondary xylem and phloem to their bodies even while they are only a few months old.
A word of caution before we leave this overview of plant structure. Although almost all plants are organized as described here, many plant parts have evolved such that they carry out unusual functions and have shapes that fool even professional botanists. The spines of cacti are modified bud scales of axillary buds even though they have almost no features typical of a bud scale (FIGURE 2-20). The beautiful white or yellow flowers of daisies are actually entire groups of tiny flowers rather than being one single flower as it would appear (see Figure B2-1B). And palm trees, despite being big, hard, and long-lived, are herbs: They never make even a single cell of wood nor do they have a vascular cambium. Plant structure is one of the easiest aspects of biology to study because plants do not run or fly away; we can walk up to them and look at them closely. Most structures are easy to see with the naked eye, even more are visible with an inexpensive hand lens.

FIGURE 2-20 This is Pereskia grandifolia, one of the few cacti that still produce large, easily recognizable foliage leaves. The spines that emerge from its leaf axils are modified bud scales: There is an axillary bud there as we would expect, but the bud scales develop as spines, rather than as the typical scales as in cottonwood (Figure 2-1). All plants produce at least two types of leaves: foliage leaves and bud scales.
![]() Overview of Plant Metabolism
Overview of Plant Metabolism
All organisms—plants, animals, fungi, and all the rest—must take in energy and raw materials from their environment. This generalization has no exceptions whatsoever. We animals take in both energy and raw materials as the organic materials we call food. Plants do not eat, of course; instead they take in resources through several separate processes. Energy is provided by sunlight and is captured by the process of photosynthesis. The bulk of a plant’s body is composed of organic molecules that can all be traced back to the reaction of carbon dioxide and water during photosynthesis. Plants obtain the needed water through their roots, and carbon dioxide is absorbed from air through the epidermis. Minerals are a fourth raw material plants need, and these too are absorbed by roots.
Photosynthesis is the central metabolism by which plants acquire energy. For the vast majority of plants and algae, there is absolutely no alternative at all. The exceptions are parasitic plants that absorb organic material from a host plant. Plants are green because they contain the pigment chlorophyll, a molecule that both captures the energy of light and then passes that energy to other chemicals, transforming them. Through several metabolic steps, the captured light energy forces carbon dioxide to react with water, which yields oxygen and carbohydrate. The oxygen diffuses out of leaves as a waste product; the carbohydrate is used as the starting point for all metabolic pathways (FIGURE 2-21).
Photosynthesis only occurs when light, chlorophyll, and carbon dioxide are present. It does not occur at night, in dark caves, deep in the oceans, nor does it occur after a plant has shed its leaves in autumn. Photosynthesis also does not occur in organs such as roots, bark, and flowers that do not have chlorophyll (FIGURE 2-22). Consequently the light energy captured by photosynthesis must be moved from leaves out to other organs that do not photosynthesize, and this transport is carried out by phloem. Also energy must be stored from daytime to night and from summer to winter. Most plants store energy by polymerizing glucose molecules together into starch, which is more compact and stable than sugar. A few plants, such as peanuts, sunflower seeds, and avocadoes, convert glucose to fats and store those as lipid droplets within their cells, but plants never have adipose tissue like ours.
Alternatives
BOX 2-2 Plants Without Photosynthesis
Are ordinary stems and foliage leaves really adaptations that improve photosynthesis? Or do plants have plant-like structure just because that is the way things are? It is easy to take really common things for granted and not even ask ourselves if there are alternatives. We can create many models and consider theoretical aspects, but we always want to verify our theories with real data, with empirical evidence. That is a principle of the scientific method. To see just how much of ordinary plant structure is the result of natural selection that improves photosynthesis, we need plants that do not photosynthesize, that are heterotrophic, and that have been heterotrophic for so long that their bodies have had many millions of years to evolve without being restricted by the need to photosynthesize. The plants we need are parasitic plants.
Approximately 4,000 species of parasitic plants occur in about 18 families (TABLE B2-1), and the ability for one plant to attack another has arisen evolutionarily about 12 times. Because there has been a dozen different starting points and because some groups had more time than others to become adapted to their hosts, parasitic plants differ widely from each other in structure, physiology, and so on. Two fundamental types of parasitic plants occur.
Hemiparasites have chlorophyll and produce all, or at least part, of their own glucose; they merely obtain water, minerals, and perhaps some organic compounds from their hosts. Hemiparasites still have leaves with stomata and rather ordinary stems. Their roots, however, penetrate the epidermis or bark of host plants rather than soil.
Holoparasitic plants have neither chlorophyll nor photosynthesis: All of their ATP is produced by aerobic respiration of glucose obtained from the host plant, and they probably need little reducing power. In all cases, after the ancestral parasitic plants became holoparasitic and lost any need for photosynthesis, mutations that curtail leaf development became beneficial: Most holoparasites have either small, scale-like leaves or none at all. Dodder (Cuscuta) is one of the few holoparasites that has any ordinary plant-like features left in its body (FIGURE B2-2A). It has long, slender stolon-like shoots with tiny leaves; it twines around host plants, inserting haustoria (modified roots) into them and then growing onward. Its plant-like features of nodes and internodes seem to be necessary for it to spread from branch to branch of a host or from one plant to another.
TABLE B2-1 Families that Contain Parasitic Plants

Most holoparasites are subterranean. Their seedling roots invade host roots, then the parasite’s embryo develops into a rhizome-like shoot with small, tough, protective, scale-like leaves. Being underground, it is invisible to herbivores, and temperatures and humidity are more stable, less variable than above ground. Many photosynthetic plants also have rhizomes, but many of their axillary buds grow upward as aerial, chlorophyllous branches; that never occurs in holoparasites. A few of their axillary buds grow above the soil surface as inflorescences, displaying flowers to pollinators (FIGURE B2-2B).
In other holoparasite species, development is even less like that of ordinary plants: The subterranean “rhizome” has no leaves at all, no axillary buds, nodes, or internodes. The growing apex consists of a mass of meristematic cells but has neither leaf primordia nor a root cap. Internally, the “rhizome’s” tissues are not typical of either root or shoot, having neither a single ring of bundles typical of a stem nor a central mass of xylem as ordinary roots do. Even the surface is just a rough, irregular proliferation of cells rather than a smooth sheet of epidermis. The term “runner” is often used to indicate that it is not an ordinary plant organ.
Several holoparasitic plants live entirely within the body of their hosts. After their seedling root penetrates the host, the rest of the embryo dies. The “roots” grow deeper into the host, penetrating its cortex, phloem, xylem, and pith, and they make up 100% of the parasite’s body. The parasite has no shoot at all. The term “roots” is in quotation marks because these structures have no features typical of roots. Technically this is an endophyte, a plant that lives within another. The endophyte of the mistletoe Tristerix aphyllus, which grows inside the cactus Trichocereus chilensis, consists only of a branching web of uniseriate filaments of parenchyma cells; after some months (?) or years (?), the filaments become multiseriate as their cells undergo longitudinal divisions (FIGURE B2-2C). At some point, a bit of phloem develops and an occasional, isolated vessel element—the entire plant body has no roots, stems, leaves, epidermis, cortex, pith, and only a few cells of xylem and phloem. Detailed studies are needed, but it appears that every surface cell of the endophyte body absorbs water, minerals, and organic molecules from the host’s tissues. With the entire body being absorptive, there is little need for conduction. When stimulated to flower, parenchyma cells just below the host epidermis proliferate into a nodule of callus, an adventitious inflorescence apical meristem forms, and then it breaks its way through the host epidermis. Because it has an ordinary apical meristem, the inflorescence has epidermis, cortex, a ring of vascular bundles, and pith (but no chlorophyll). Endophytes of Viscum minimum, Pilostyles thurberi, and several Rafflesia species are similar (FIGURE B2-2D).
After a green photosynthetic photoautotroph has produced glucose, it then synthesizes all other organic compounds needed for its structure and metabolism (see Figure 2-21). It may be that some holoparasites do the same thing; they may merely obtain glucose from their hosts and then use various types of respiration and anabolic pathways to synthesize all other organic molecules. It would not be surprising, however, to find some species of parasitic plants that have become dependent on their hosts for at least some amino acids, lipids, or vitamins. Despite being so modified, the endophytic bodies of holoparasites are healthy, thriving, and well adapted to respiration-based metabolism in a unique environment.
Holoparasitic plants show us that many features of ordinary plants must be very expensive and risky: After a plant has an alternative to photosynthesis, ordinary plant features are no longer adaptive, and mutations that prevent their development are advantageous. On the other hand, for plants that do photosynthesize, plant-like bodies are worth the trouble.

FIGURE B2-2A The slender orange stems are Cuscuta, a holoparasite that still has leaves and stems, but its internodes are extremely long and slender and its leaves are tiny. It is producing adventitious roots that develop as haustoria, penetrating to the vascular tissues of the host, Justicia.

FIGURE B2-2B Prosopanche americana is holoparasitic, and most of its highly modified body remains underground; it has no need for photosynthesis, obtaining all its water and nutrients from the roots of its host. This flower is also highly modified: The black and white structure is a massive set of fused stamens.
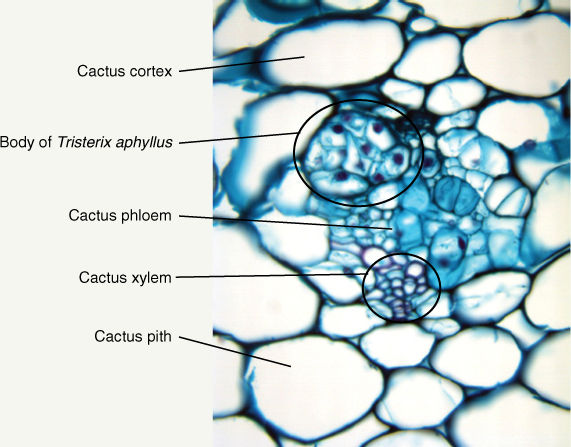
FIGURE B2-2C The small cluster of cells with large red-stained nuclei are part of the body of Tristerix aphyllus, a holoparasite whose vegetative body grows entirely within the body of its host, in this case, the cactus Trichocereus chilensis. The body of T. aphyllus consists of just parenchyma and lacks leaves, stems, epidermis, and virtually all other features of an angiosperm body.

FIGURE B2-2D The dark blobs do not belong to the visible shoot; instead, they are the flowers of a holoparasitic plant, Pilostyles thurberi, growing completely inside its host, the shrub Dalea. Holoparasites typically must emerge from underground or from inside their hosts when they flower.
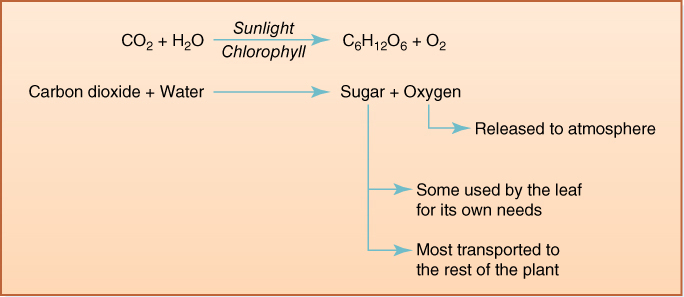
FIGURE 2-21 Simple diagram representing the raw materials and products of photosynthesis; carbohydrates produced by photosynthesis are subsequently converted to many other compounds.
Respiration is another type of energy metabolism and it occurs in all organisms, with no exceptions whatsoever. As just mentioned, glucose is used to store energy and to move it from organ to organ. But a problem with glucose is that it stores too much energy for certain needs. Cells force many metabolic pathways to run by adding a small bit of energy to two or three steps of the reaction. A molecule called ATP (adenosine triphosphate) provides just the right amount of energy for most of the steps (FIGURE 2-23). Glucose has about 36 times as much energy as is carried by ATP. Respiration breaks down glucose and transfers its energy to ATP. During photosynthesis, light energy is captured as carbon dioxide combines with water, producing oxygen and glucose. Respiration is more or less the reverse: Oxygen and glucose react, producing carbon dioxide and water with the energy being transferred to ATP rather than being given off as light.
Plants also need minerals such as nitrogen, phosphorus, potassium, iron, and others. Plants use these in much the same way we do; phosphate is needed for ATP; nitrogen becomes part of amino acids and nucleic acids; iron is attached to molecules that transfer electrons, and so on. Also like us, plants are not especially effective at recycling minerals; we lose minerals as we sweat, urinate, and defecate. Plants lose minerals has they shed their leaves, withered flowers, and fruit. They pack their seeds with minerals that the seedlings will need, and all those minerals are provided by the parent plant. The amount of minerals a plant needs is surprisingly small; the ash that remains after a fire contains all the minerals that had been present in the wood.

FIGURE 2-22 Only the leaves of these palms (Washingtonia filifera) are green and photosynthetic, they are the only parts of the trees that produce sugar. All other parts, such as the long stems and the underground root systems, have no chlorophyll and must obtain all their energy in the form of sugars transported downward from the leaves by phloem.
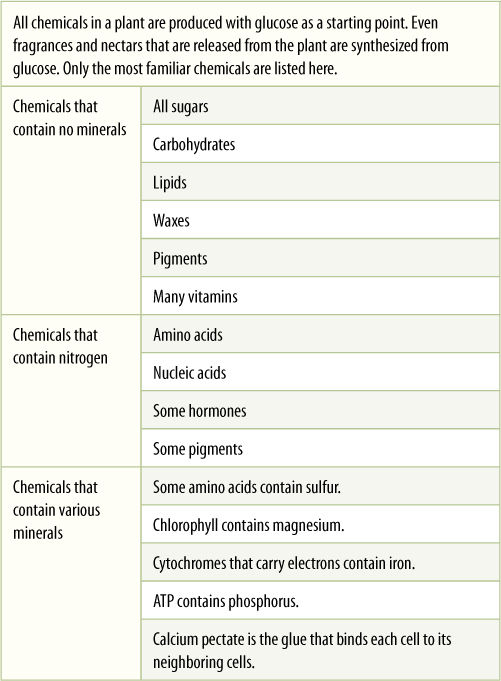
Plants are master chemists. Starting with just glucose, minerals, and water, they synthesize every single molecule they need for all aspects of their life (TABLE 2-3). They synthesize all amino acids, nucleic acid, and fatty acids; they make all the pigments of flowers and fruits; they make waxes, oils, resins, and fragrances. And poisons—many plants defend themselves from pests with an amazing array of poisons. Some are bitter or irritating, others kill animals quickly; some of the most toxic chemicals interfere with cells in the brain and nervous system. Although we humans need a complex diet that includes several amino acids, fatty acids, and vitamins that we cannot make for ourselves, that is never true of plants. Their balanced diet is sunlight, carbon dioxide, water, and minerals.
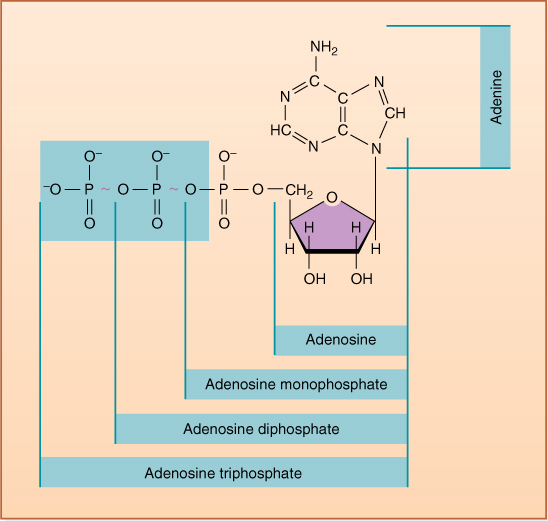
FIGURE 2-23 Adenosine triphosphate. Breaking off the last phosphate to produce adenosine diphosphate (ADP) results in a more stable set of electron orbitals, and energy is given off. The same is true of removing the second phosphate, but not the third.
Plants and People
BOX 2-3 Toxic Plants
Consider two facts: (1) Plants live among herbivores, and (2) the world is full of uneaten plants. What is the cause? Many plants produce toxic compounds: Poison ivy, death camas, poison hemlock, and deadly nightshade are familiar examples, but the full list of toxic plants is very long. Many plants we think of as being nonpoisonous are only harmless because we stop eating things that are bitter or that burn our mouths: Few people would eat enough chili peppers to get a lethal dose. The following are some toxins in plants we live with. Fascinating and well-illustrated accounts of many are given in Handbook of Poisonous and Injurious Plants by Lewis S. Nelson, Richard D. Shih, and Michael J. Balick (2007, Springer).

FIGURE B2-3 Each leaf of poison ivy has three leaflets and the defensive compound urushiol.
Anticholinergic (antimuscarinic) poisons. Many nerve cells have receptors that detect the presence of a small molecule called acetylcholine, which causes a reaction in the nerve cells when it binds to the receptors. Plants such as Atropa (deadly nightshade) and Datura (jimsonweed) produce chemicals that bind to one of these receptors (called the muscarinic receptor), preventing acetylcholine from stimulating proper nerve transmission.
Nicotine-like alkaloids. Alkaloids are small molecules that contain nitrogen and somewhat resemble amino acids. Alkaloids of Conium (poison hemlock), Nicotiana (tobacco), and Sophora (mescal bean) block another acetylcholine receptor (this one called the nicotinic subtype). Symptoms occur in the parasympathetic and sympathetic parts of the nervous system, the brain, and the junctions where nerves attach to muscles.
Convulsant poisons. The small tree Strychnos produces strychnine, which causes motor nerve cells to be hyperexcitable and results in convulsions. Several other plants produce convulsants.
Capsaicin. Chili peppers such as habaneros, jalapeños, and tabascos in the genus Capsicum contain capsaicin, which induces some of our sensory nerve cells to release a chemical called substance P, which specifically stimulates other nerve cells that normally respond when we are being burned. Artificial selection by humans has resulted in chili peppers that range from having almost no capsaicin (bell peppers) to some that are dangerous (cayenne, chiltepin, and Thai). Capsaicin is hydrophobic, and thus, drinking water will not cool your mouth; however, the fat in cheese, butter, and milk will absorb it and give some relief.
Cardioactive compounds. Digitoxin and digoxin in Digitalis (foxglove) inhibit a set of proteins that use the power of ATP to pump sodium and potassium across a membrane. When this pump is blocked in heart cells, it leads to an increase in the concentration of calcium within the cells. Although foxglove leaves are injurious if eaten improperly, these chemicals are used medicinally to slow and strengthen heartbeat.
Cyanogenic glycosides. These are nontoxic in the plant, but enzymes in our digestive system cleave them into two parts: a sugar and cyanide. Cyanide inhibits the final step in aerobic respiration, preventing generation of ATP. These toxins are found in some of our most common fruits—Malus (apples) and Prunus (“stone fruits” such as cherries, peaches, apricots)—but in parts that we do not eat such as apple seeds or peach pits. Poisoning most often occurs when apricot seeds are used as components of herbal medicines.
Mitotic inhibitors. Microtubules create the spindle that pulls chromosomes to opposite ends of the cell during nuclear division. The alkaloid colchicine in autumn crocus, Colchicum autumnale, prevents the assembly of microtubules. Cells exposed to this poison cannot form a spindle, and division fails. We humans have many tissues with rapidly dividing cells, such as the lining of our digestive tract, bone marrow cells that produce red blood cells, and seminiferous tubules in men. Catharanthus roseus (also called Vinca rosea, periwinkle) produces the alkaloids vinblastine and vincristine that also block cell division; these are used in chemotherapy to stop cancer cells from dividing.
Toxalbumins. These proteins bind to ribosomes and prevent them from translating messenger RNA into protein. Ricinus (castor bean) produces the toxalbumin ricin, one of the most toxic substances known. Jatropha species (coral bean) has a higher concentration of a related toxalbumin, and a single seed can cause severe poisoning.
Urushiol. About half of all people are allergic to urushiol, an oily substance in species of Toxicodendron (poison ivy, poison oak, and poison sumac). All parts of these plants can trigger an allergic reaction.
Remember that these plants do not have toxic compounds in order to protect themselves, or with the purpose of keeping herbivores away. It is just that toxic or injurious plants are more likely to be left uneaten compared with nontoxic plants and therefore are more likely to produce more seeds and more progeny.
TABLE 2-3 Chemicals that Plants Produce Starting with Glucose
![]() Overview of Information in Plants
Overview of Information in Plants
A universal feature of all known life on Earth is that DNA (deoxyribonucleic acid) is used to store information. Just as with energy and matter, organisms take in information, store it, move it from place to place, transform it, use it, and then pass it onto their offspring when they reproduce. The greatest source of information for any individual is its parents: the DNA contained in the sperm cell and egg cell that initiated the life of the individual. In plants, DNA is stored in the nucleus of each cell, as well as in plastids and mitochondria, which are also present in all living plant cells. When a cell needs to use some of the information, it transforms the needed information by making messenger RNA (ribonucleic acid), and then it transfers that information to the area of the cell where it is needed. Most of the information is further transformed as the messenger RNA guides the production of specific proteins. Smaller pieces of RNA, often called microRNA, act by binding to various components of the cell, activating some, inhibiting others, and in general fine-tuning the activities of many genes. The DNA of each species is unique; for example, cherries (Prunus avium) and peaches (Prunus persica) have almost identical DNA, but the information in several genes in one differs just enough from that of the other so that cherries are small, red, and shiny whereas peaches are large, yellow, and fuzzy, and each has distinct flavors and aromas.
A plant, just like any other organism, also uses information from the environment. Environmental information must be perceived then transferred to where it is needed, and finally the plant must respond appropriately. While the plant is still nothing more than a dormant seed, the environmental information it responds to is often very simple: It is warm enough to germinate. As temperatures climb from winter to springtime, at some point they are warm enough and many seeds then respond by becoming active, respiring more rapidly, and germinating. Seeds of other species remain dormant because they are dry, not cold; they need to detect when their surroundings have become wet enough to support a growing seedling.
The environment also supplies a large amount of more complex information, in many forms (FIGURE 2-24). Sunlight provides the energy for photosynthesis, of course, and it also indicates the direction up (the direction of brighter light) from down (the direction of less light and more shade). In all parts of the world except near the equator, sunlight provides information about seasons: Days are shorter in winter, longer in summer. By detecting this, a plant measures the season and responds by flowering and fruiting at the correct time and by becoming dormant in autumn. Roots are not exposed to sunlight, but they detect gravity and thus know which direction is down. Deeply buried bulbs also detect gravity and respond in two opposite ways: Roots follow gravity downward, whereas shoots grow upward (FIGURE 2-25). We are now learning that some plants are much more perceptive than we ever suspected. Tendrils bend toward anything that gives them even the slightest touch, leaves detect the growth of a single fungus spore on their surface, and some orchids detect whether a pollinator has merely landed on one of their flowers or if it has actually taken the pollen.

FIGURE 2-24 All plants here obtain energy from their environment, but think of the two trees. They both obtain information about which way is up (the opposite direction of the force of gravity pulling on all parts). For the grasses in the pasture, up is in the direction of the brightest light, but for the tree on the right (closer to the shed), the brightest light is away from the larger tree that is shading it, so “bright” and “up” are not the same thing for it. These trees also need to detect the approach of winter and abscise their leaves while they can, and they probably detect the presence of many insects and fungi as well.

FIGURE 2-25 When these bulbs became active and started growing, all parts were so deeply buried in soil that they could not detect any sunlight. But both new roots and new leaves detect gravity, with roots growing downward, leaves growing upward.
Various parts of a plant itself create information that informs the rest of the plant of their activities. Healthy leaves produce hormones that are carried to the roots: A large amount of hormone indicates the leaves are active and need water. The roots respond by growing and absorbing water and nutrients. In autumn, as leaves prepare to be shed, they stop producing the hormone, so the amount of the hormone decreases, indicating the leaves no longer need water. Roots respond by slowing their growth and conserving their energy. After a flower is pollinated, the developing fruit sends signals indicating that it needs nutrients for itself and the embryonic seeds inside. This communication among the parts of a plant is necessary to keep the plant integrated and all parts developing in coordination. It is especially remarkable considering the long distances involved between the thousands of leaves, flowers, branches, and roots of a large tree.
![]() Overview of Plant Diversity and Evolution
Overview of Plant Diversity and Evolution
All the thousands of species of plants of the world are not just a hodgepodge of randomly associated features. Instead there are several large, recognizable groups such as mosses, ferns, conifers, and angiosperms. Within each group, we are likely to find certain features and unlikely to find others. For example, mosses never have xylem or phloem and they never have seeds either. As a second example, ferns always have vascular tissue but they never reproduce with seeds. Several aspects of the groups in TABLE 2-4 are not quite accurate, but the groups and terms presented there have been used for more than 100 years, and you will still encounter them often in books about gardening and roadside flowers. An overview of them will be given here. Definitions of important terms are grouped together in TABLE 2-5.
Mosses, liverworts, and hornworts are often grouped together and called bryophytes (pronounced BRY oh fights) (FIGURES 2-26 to 2-29). They are similar to each other because they not only lack vascular tissues and seeds, but also because plants in all three groups are haploid. Mosses, liverworts, and hornworts are unique among plants because they live their lives with only one set of chromosomes, not two. Their bodies are very simple, not much more complex than the algae that were the ancestors of plants.
TABLE 2-4 Overview of the Main Clades of Organisms
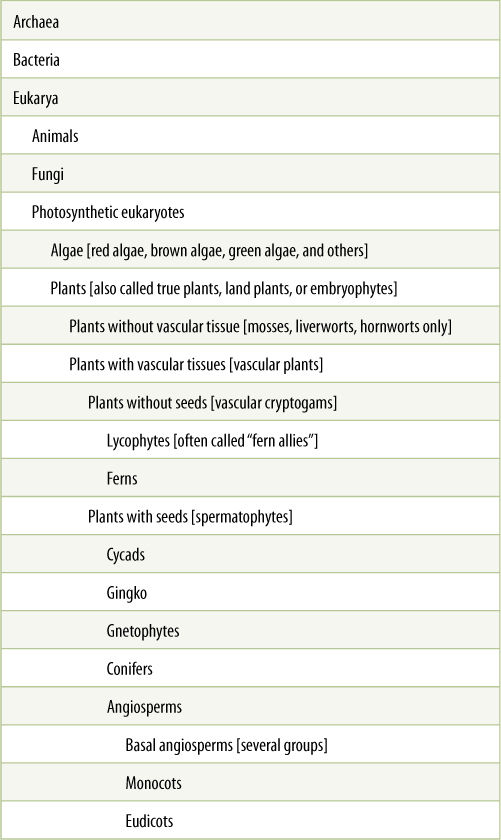
TABLE 2-5 Terms Used for Plant Groups

* Names in formal classifications are capitalized but informal names are not. The informal names of some of the groups here are: angiosperms, conifers, gnetophytes, and mosses. Their formal names are: Angiospermophyta, Coniferophyta, Gnetales (some people use Gnetophyta), and Musci (an old name for mosses) or Bryophyta (a newer name for mosses).
All other groups of plants have vascular tissue and are diploid. Many vascular plants resemble bryophytes in being unable to form seeds; they reproduce using spores and are typically called the spore-bearing plants. These are often referred to as ferns and “the fern allies” (FIGURES 2-30 to 2-32).
An old way of thinking about plants is that ferns and “fern allies” had advanced above the bryophytes as they evolved to have vascular tissue, but then their evolution “stagnated” while other vascular plants continued to advance evolutionarily to the point where they could make seeds as well as vascular tissues. The more advanced seed plants then divided into two groups, those with naked seeds (the gymnosperms) and those with “clothed seeds” (the angiosperms, often called the flowering plants). Here, “clothed seeds” refers to seeds formed inside a fruit in contrast to naked seeds that are produced in a cone. The term “naked” refers to the time when a pine cone is extremely young; at that point, its scales are far enough apart that you can look in and see the individual naked ovules. The carpels of angiosperms are always sealed, and the ovules are never visible without cutting the young fruit open. There are other differences between gymnosperms and angiosperms that make us think of angiosperms as being more advanced: Angiosperms have more complex wood and leaves, as well as several modifications that improve photosynthesis. Angiosperms also cooperate with animals that pollinate their flowers and distribute their seeds whereas conifers never rely on animals for pollination, and only a few cooperate with animals for seed dispersal.

FIGURE 2-26 Mosses cover the rocks in this stream in the Olympic National Park. Mosses have no xylem or phloem and cannot conduct water very well, so they are typically very small plants. Although we associate mosses with damp places, many are adapted to deserts, being inactive except on rare days when it there is rain, fog, or dew.

FIGURE 2-27 These are plants of the leafy liverwort Bazzania trilobata. As with mosses, the bodies of leafy liverworts consist of a stem and leaves, but never any roots, xylem, or phloem. It is usually necessary to have a hand lens or magnifying glass to distinguish between a leafy liverwort and a moss.

FIGURE 2-28 This is a rather common thalloid liverwort (Conocephalum conicum) that is easy to recognize: Its bodies never have any hint of leaves or stems, they are flat and about 1 cm wide, and they occasionally fork into two growing points. Their upper surface has a distinctive diamond-like pattern that resembles reptile skin.

FIGURE 2-29 Hornworts such as this Phaeoceros laevis are widespread, but they only thrive when it has been rainy for awhile; they grow in shady places, sometimes on bare soil, sometimes mixed among grasses. Each long projecting cylinder (about 1 cm long) is the “horn” and is actually the diploid portion of the plant (technically, a sporophyte): Each horn is the same phase of the life cycle as is a tree, bush, shrub, or any other familiar plant. The small flat patches of tissue in contact with the soil are the gametophytes, the haploid part of each plant: It is the part that is similar to mosses, leafy liverworts, and thalloid liverworts. The life cycle of plants is much more complex than that of animals, but is fascinating.
We can arrange the different groups in a linear sequence that seems to reflect plant evolution. The earliest true plants were more advanced than algae because they had simple tissues and organs but little else. This would be the moss level of evolution. Other plants advanced to the level of having vascular tissues, others continued evolving to the point of having seeds. Then one group evolved more than all the rest, to the point of having flowers and fruits. Some would even refer to angiosperms as the “pinnacle” of plant evolution. In this method, emphasis is placed on the level to which a group has evolved. The technical word for level is grade; we are grouping plants into grades, with the lowest grade being nonvascular plants, the highest grade being angiosperms. We still encounter this concept of grade when people refer to the “lower plants” and “higher plants.” This grouping by grade is very handy if you are not too worried about accuracy, but we now have a newer method of grouping plants, one based on groups that consist of all the species that evolved from a common ancestor. Such a group is a clade.
Fortunately, grouping plants into clades gives us almost the same result as grouping them into grades (FIGURE 2-33). We need to abandon only a few of the old terms. For example, we no longer use the term “gymnosperm.” We suspect—but are not at all certain—that seeds evolved only once. If so, then all seed plants alive today descended from that original species of seed plant, and we say it was the common ancestor to all seed plants. Some of its descendants evolved into cycads, others became ginkgos, another group evolved into conifers, and still another group became angiosperms. There are two important points here. The first is that a clade consists of an ancestor and all its descendants, so the group “seed plants” is acceptable—it is a clade. But the word “gymnosperms” refers to all seed plants except angiosperms; it includes the common ancestor of seed plants but only some, not all descendants, so “gymnosperms” is not a clade. We prefer to not to use the term “gymnosperm” if we care about being accurate.

FIGURE 2-30 (A) This Selaginella lepidophylla is used as an example of a “fern ally” because of its interesting biology: It is a resurrection plant. It is a diploid vascular plant with xylem and phloem, and it grows in harsh deserts with long dry periods. This photograph shows the plant after autumn rains, when the plant is hydrated, healthy, and active. Compare with Figure 2-30B. (B) This is also Selaginella lepidophylla but after a long dry period; the plant has curled up, minimizing the amount of surface exposed to dry air and UV light. Although it looks dead, it is not only alive but very healthy, merely dormant, somewhat like a dry seed that is alive. After a rain, these plants would uncurl and look like those in Figure 2-30A. If forcibly uncurled now, the inner parts would be bright green.
The second important point is that the ancestors of any clade of living organism all lived in the past, and for clades such as angiosperms or conifers or seed plants, the ancestors lived in the very distant past. Consequently, we can never be completely certain which ancient, extinct group was the common ancestor of any modern clade; instead we have to study the living organisms and try to infer the features the common ancestor had. Using angiosperms as an example, all modern as well as fossil angiosperms have vascular tissues; they produce flowers and they make seeds, they never make anything similar to a pine cone, and they are never haploid like bryophytes. From this we assume that the common ancestor of all flowering plants was a diploid vascular seed plant that made flowers but not cones. At present, we are able to analyze the sequence of nucleotides in the genes of organisms (we say that “we sequence their genes”) and at present we have sequenced all the genes (the entire genome) of many species. This provides an abundance of new information for constructing and testing our theories and models.

FIGURE 2-31 These plants of Equisetum telmateia are diploid vascular plants that are now considered to be part of the ferns. These are descendants of an ancient group: Fossils of plants that have similar features can be found in rocks as much as 150 million years old. Equisetums are called horsetails or scouring rushes, and are distinctive in having a very coarse surface (due to having silica in their cell walls); they can be found almost everywhere, along streams and along railroad tracks in moist areas.

FIGURE 2-32 This is a single plant of sword fern (Polystichum munitum); it has a short vertical stem that supports the many leaves, and below ground are many fibrous roots. Ferns are diploid and have xylem and phloem, but their spores grow into tiny haploid plants that resemble the bases of hornworts (Figure 2-29). This large sword fern pictured here is the equivalent of a hornwort horn (both are sporophytes). In rainy areas of the Pacific Coast of the United States, sword ferns are so abundant they resemble meadows of grass; indeed, millions of years ago before grasses evolved, most “grasslands” were actually huge patches of ferns. Ferns and dinosaurs were abundant at the same time, long before grasslands existed—the herbivorous dinosaurs ate ferns and fern allies.
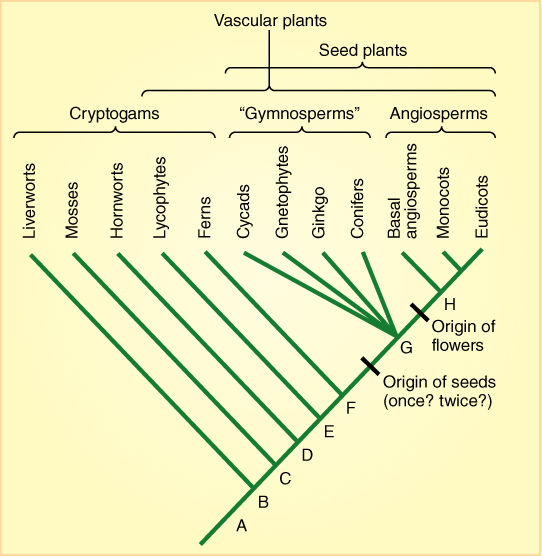
FIGURE 2-33 This cladogram is a hypothesis about the ways in which modern plants are related to each other. At the very base (marked by A) would be the ancestral plants, not much different from algae. Many years later, at B, some plants began to evolve into liverworts while the rest of the plants continued evolving other features. At C, some plants began to evolve into mosses, and so on. At some point between D and E, vascular tissue evolved in the main group of plants, and all plants that descended from these early vascular plants still have vascular tissue as well. That means that the xylem in ferns resembles the xylem in flowering plants not by accident but because both evolved from the same ancestral xylem (to be really careful: The genes that control the development of xylem in ferns and the genes that control the development of xylem in flowering plants resemble each other because both sets of modern genes evolved from the same set of ancestral genes). The lines that show the ancestry of cycads, gnetophytes, Ginkgo, and conifers all start from the same point (G): That means we do not yet know which diverged earliest, and we do not know which group is most closely related to which other group. The line for the ancestry of angiosperms diverges a bit later (H), but it could be that this is incorrect and we should draw it coming from G, the same point as the cycads and others.
The old concept of grades assumed that cycads, ginkgos, and conifers were evolving more slowly than angiosperms and, given enough time, they would finally at long last evolve up to the level of having flowers. Similarly, maybe one day in the future, ferns would evolve to the point of having seeds, and so on. It is like moving from grade to grade in school: Everyone starts out in the first grade and then all move along toward graduation, some moving faster than others, some stopping completely. This is the second problem with grouping plants into grades: It assumes that all plants have the same goal, that having vascular tissue and seeds and flowers is the optimum way a plant can be adapted to its environment. We no longer believe this.
We now realize that there are multiple ways for an organism to be adapted to its environment. This is reflected in the questions “What are the alternatives?” and “What are the consequences?” Consider mosses and angiosperms, two groups at the opposite end of the old grade system. Mosses are tiny and simple whereas most angiosperms are large and complex. But mosses are very well-adapted to their environment: They thrive in low light, on brick walls or bare stone; they need only enough sunlight, water, and mineral nutrients to make a few spores and then they can reproduce. In the moss clade, evolution by natural selection has found solutions to survival that work well for small, simple plants. Having vascular tissue and seeds and so on would not make a moss any more adapted than it is without them. This is a crucial point I will emphasize many times: There are many, diverse ways to be adapted, numerous ways of thriving. There is not one single structure or physiology that will be optimum for every organism. No organism is a pinnacle of evolution.
Botanists are working harder than ever trying to figure out how plants are related, which clades are valid. The following are the ones you should know as you continue into this book. Plants are a clade: True plants originated from green algae only once. The technical name for plants, to distinguish them from algae, is embryophytes, but this is used in only the most technical writing. It had been suspected that mosses, liverworts, and hornworts might have had a completely separate origin, perhaps they did not descend from the same ancestor that produced all the other plants. But the sequences of nucleotides in the genes of bryophytes are so similar to those of all other plants that we are now confident they too are true plants. The terms “vascular cryptogams” and “fern allies” are handy but not accurate. Early diploid plants diverged into two clades, both with vascular tissue and without seeds. One clade, which we call the lycophyte clade (pronounced LIE co fight), was very successful initially and evolved to be large trees with many complex features, but almost all became extinct. At present, the surviving descendants are Lycopodium, Selaginella, and a few other genera; these are the “fern allies” you will see in wildflower books (see Figure 2-30). The second clade was a bit slower at the start, and for millions of years was overshadowed by the abundant members of the lycophyte clade. Many of the early members became extinct, but this clade produced all the ferns and seed plants. The term most often used now is the euphyllophyte clade (pronounced you FILL oh fight). The ferns make up a subclade of the euphyllophytes, and they have been very successful without having seeds (see Figures 2-31, 2-32). Ferns thrive in all sorts of habitats, not only shady, damp forests, but also hot, dry deserts. They never produce wood, so they are all herbs, even the very tall ferns called tree ferns. Some ferns have evolved to have sophisticated relationships with ants, relationships that are mutually beneficial.
The ferns make up a well-defined subclade within the euphyllophyte clade, but we are not certain how to group the remaining vascular plants. Many are known only as fossils of isolated leaves or wood or reproductive organs; it is very rare to ever find a fossil of an entire plant with all parts still attached to each other. Some especially intriguing fossils look like fern leaves with seeds on them, so they were named “seed ferns.” But no seed fern is alive today, and no living fern ever has anything that even slightly resembles a seed. We believe that the name “seed fern” was a poor choice, but by now it is very commonly used. The many fossils of seed ferns are too incomplete for us to know if there was only one group of early seed plants or several. If there was only one group of seed ferns, then probably seeds evolved just once. If so, all seed plants make up one clade, the seed plants (more technically, spermatophytes). But if seeds evolved two or three times, then “seed plants” is not a clade, not a closely related group that all descended from the same common ancestor. If seeds evolved several times, “seed plants” would then be the name of a grade and could only be used informally.
Other extinct seed plants lived after seed ferns. Many look like intermediates in the evolution of modern seed plants from seed ferns, but the picture is not yet clear. Many people are taking a cautious approach now, tentatively considering all the seed plants as being related. They assume that seed plants evolved only once, and then diverged into several major groups, most of which have some members that are still alive today. According to this theory (which is the one followed in this book), the early seed plants gave rise to these clades: cycads, gnetophytes, Ginkgo, conifers, and angiosperms.
Until recently, it was thought that all members of the angiosperm clade were either monocots or dicots (FIGURE 2-34). We used to think that after the first flowering plants originated, the group immediately diverged into two distinct clades, monocots and dicots. But new evidence indicates that split did not occur immediately. Early flowering plants evolved and diverged into several groups before the monocots originated and became so distinctive. The term “dicot” had referred to all the angiosperms except monocots, but like the term “gymnosperm,” this is unacceptable because it does not include all the descendants of the common ancestor of the group (it leaves out monocots). At present, we use the terms basal angiosperms for those angiosperms that diverged early; monocots for the clade of lilies, palms, agaves, orchids, and their relatives; and eudicots (pronounced you DIE cots) for all the rest of the angiosperms. You will still see the term “dicot” used in many books, and it most often means “basal angiosperms plus eudicots.”
Although the terms “dicot,” “gymnosperm,” and “fern allies” are being abandoned, it is good for you to know why they were invented and how they were being used. Even though you will still see them occasionally, it is best if you avoid using them.

FIGURE 2-34 (A) These avalanche lilies (Erythronium montanum) are shown as representative monocots, but the monocots are a large, diverse group. Many have large showy flowers, although flowers of grasses and sedges are small. Most monocots have parallel venation in their leaves, and many die back to a persistent subterranean bulb or rhizome. (B) These Joshua trees (Yucca brevifolia) are shown as another representative type of monocot. Many people think of monocots as small, delicate plants like lilies, irises, crocuses, and so on, but many, such as Joshua trees, become very large and tough; other strong monocots are palms, agaves, yuccas, bamboos, and many bromeliads. (C) This lupine, Lupinus latifolius, is shown as a representative eudicot, but eudicots are a gigantic and diverse group. Of the plants you see every day, unless a plant is obviously a monocot, conifer, or fern, then it is probably a eudicot. Eudicots can be trees, shrubs, herbs, vines, parasites, and succulents, as well as many other types of plants.
Botany and Beyond
BOX 2-4 Noah’s Flood and Population Biology
The biblical story of Noah’s flood in Genesis provided a powerful stimulus for developing the sciences of population biology, biogeography, and evolution. Various biblical scholars had studied Old Testament texts, the Jewish Torah, and archaeological evidence, trying to establish dates such as the birth of Jesus (1 ad, which is 2,017 years ago now) and the construction of Solomon’s temple (832 BC). Although there is no archaeological evidence, various Bible scholars concluded that the flood of Noah occurred in 2348 BC. Noah’s flood was of special interest to early scientists because at that point according to the Bible, all life on Earth had been destroyed by God except for the creatures in the Ark. Scientists formulated the question “Do plants and animals reproduce rapidly enough to repopulate a completely denuded Earth in the time since Noah?” Thus a scientist posing this question in 1700 ad, for example, could assume that Earth had been repopulated in 2,348 + 1,700 = 4,048 years.
First consider the Bible passages that the scientists relied on. Genesis 7: 2—3 states that God told Noah to take seven pairs of every kind of “clean animal” (those that could be used for sacrifice; Genesis 8: 20); only one pair of every kind of “unclean animal” (which could not be used for sacrifice); and also seven pairs of every kind of bird. Genesis 7: 20 states that the flood completely covered the tops of all mountains; that means that there was no dry place anywhere that animals could use as a refuge and survive. Although we are all familiar with the rain lasting 40 days and 40 nights, the Bible states that it took a long time for the waters to drain away after the rain stopped. For the early scientists the critical passages are Genesis 8: 4 and 5 that state that the Ark was floating for 7 months before it settled on Mount Ararat, and it was 10 months before the tops of other mountains were visible and Noah opened the doors. Especially important was the explicit statement that God’s plan succeeded: Genesis 7: 23 states that everything outside the Ark had been killed.
In the book of Genesis, God had set up the perfect experiment in population biology. All animals had been killed (there is no mention of Noah needing to collect seeds to save the plants), and Earth had to be completely repopulated with all types of animals and birds starting with just one male and one female of each of the unclean animals and just six males and six females of each of the clean animals (Genesis states that Noah sacrificed one male and one female of each type of clean animal after the doors of the Ark were opened).
Could just 12 cattle (clean) breed rapidly enough to produce all the herds and wild cattle on Earth in 1700 ad? Could just two pigs (unclean) be the source of all swine now present? The first step in answering this was to determine the fecundity of animals: How old do they have to be before they produce their first young? How many young do they produce in each litter? How many survive long enough to breed? And how many sets of offspring do they produce in their lifetime? Fortunately it was about this time that wealthy landowners began keeping track of these sorts of numbers for cattle, poultry, fish, fowl, and even the deer in their private forests. The answer for every type of animal was the same: Yes, all animals produce young rapidly enough to repopulate Earth in much less than 4,000 years.
The early scientists then began to refine their thoughts and reformulate the question. At present we believe in a “balance of nature:” Plants capture carbon and energy through photosynthesis, then the plants are consumed by herbivorous animals or by pathogenic fungi and bacteria. The herbivores are eaten by carnivores. If there are too many herbivores, they damage the plants too much, and some herbivores starve, which in turn affects carnivores. An outbreak of a pathogenic fungus that damages plants will cause various animals to starve, and so on. The balance between all these factors changes from year to year, so the relative numbers of organisms also change yearly. But prior to the analysis of Noah’s flood, people believed that a different kind of “balance of nature” was maintained by God: God had created the various carnivores, herbivores, and plants such that plants would prosper just enough to supply the herbivores with exactly the right amount of food, and likewise the number of herbivores would be perfectly correct for the numbers of carnivores in the world. The balance would be perfect, never changing, and with no waste or shortage unless God specifically causes a drought or pestilence.
As early scientists studied animal fecundity, they realized that the numbers did not fit together to make a divinely balanced system. All organisms are too fertile. If the previous interpretation of Genesis were correct, God had created plants and animals with the wrong properties to fit into a system that would be perfectly balanced and unchanging to the end of time. They realized that the story of Genesis as well as their own research pointed not to a stable, harmonious, garden-like Earth but rather to one of change and competition, of some organisms increasing at the expense of others. Population biology was born and has thrived ever since.
![]() Overview of Plant Ecology
Overview of Plant Ecology
Plants live in a complex, dynamic, and changing world. Earth’s complexity is easy to see. Some areas are mountainous with cold temperatures and thin, rocky soil but other areas are low, warm, and moist. Volcanoes release lava and ash that are initially sterile but that gradually change into rich acidic soil. In contrast, limestone breaks down to form alkaline soil. In addition, some soils are so rich in certain minerals they are toxic and plants cannot grow on them, whereas other soils have a balance of mineral nutrients that encourages luxuriant plant growth. On mountains and in valleys, the slopes that face the equator receive more sunlight and are much warmer than the sides that face away.
Earth is dynamic—energetic—for many reasons. As Earth rotates on its axis, a particular area faces the hot sun during the day but faces the extreme cold of outer space at night. Throughout a year, seasons change from spring to summer and so on, each characterized by the amount of energy available from sunlight. Earth’s atmosphere and its oceans are two gigantic fluids that circulate, moving energy from place to place. We experience the circulation as cold fronts of frigid polar air that move southward, pushing warm air aside, or as storms and hurricanes that carry water and energy in such concentrated forms they destroy buildings and push over trees.
Earth’s hot interior is another source of energy that affects life on the planet’s surface. Erupting volcanoes catch our attention, but Earth’s interior heat also powers continental drift (technically known as plate tectonics): Hot, molten rock rises along the center line of each ocean then spreads outward as newly-formed sea floor. This pushes the old sea floor to the side, which in turn pushes continents from place to place. North and South America formerly were widely separated by thousands of miles, but the continental drift of each changed direction and now they are moving toward each other. Central America is the result of this collision, which is still occurring: North and South America first made contact with each other less than 2 million years ago. As they continue to push into each other, they crumple Earth’s crust, pushing Central America upward and causing numerous volcanic eruptions throughout the region. As the two continents finally came into direct contact, many animals and plants could migrate from one to the other. For example, armadillos have been slowly migrating from South America into North America, and have only recently arrived in the southern United States. Unfortunately, sabre-tooth tigers migrated south, and as they spread into South America, they killed off many large animals, driving many species to extinction. Cacti originated in South America and were confined to just that continent until continental drift finally brought North and South America together; cacti could migrate north into Mexico and the deserts of the United States as animals carried their seeds (FIGURE 2-35). The creosote bush that gives all the deserts of the southwestern United States their distinctive aroma also came from South America (FIGURE 2-36).
Earth is a changing world. For comparison, think about our moon: It is complex but not dynamic. The craters we see today are the same as they were thousands or millions of years ago, with the only new activity to ever occur is an occasional meteor strike. But Earth is never the same, not even from one year to the next. Mountains are lifted by volcanism or continental collision, and they are worn down by erosion. Flooding rivers carry silt that settles into rich deltas and marshlands filled with life. Fires drastically alter the numbers of plants and animals in forests and prairies that burn. As continents drift, their changing positions affect the circulation of the oceans and thus the circulation of heat and moisture. During the time North and South America were widely separated, the Pacific and Atlantic Oceans were in contact and water circulated around each continent. But once the continents collided and formed Central America, oceanic circulation was altered. Water was forced to flow northward and southward, carrying warm water toward the poles and making those areas warmer than they had been before. Plants and animals located in the far northern parts of North America and Europe experienced climatic warming.

FIGURE 2-35 This cactus, Stetsonia coryne is at home in Argentina; it is one of many South American cacti. For almost any cactus you would encounter north of the Panama Canal, there are similar but unrelated species in South America. There were no cacti in North America until continental drift brought North and South America close enough together for seeds to be carried northward.

FIGURE 2-36 Creosote bush (Larrea tridentata) is at home in the deserts of North America but its closest relative, Larrea divaricata and two other species of Larrea, live in South America. The plants in South America resemble those in North America so closely that we believe that at some very recent time, a seed or two of L. divaricata was carried northward the entire distance and then thrived in the new habitat. Once in North America, the plants gradually evolved just enough that we consider them to be a new species, L. tridentata, distinct from the ancestral L. divaricata.
Much earlier than that, just before plants evolved from green algae, all continents had drifted together into one gigantic continent called Pangaea (pronounced pan GEE uh). This was so enormous that its center was a desert: Storms blowing in from the oceans would rain themselves out before they could reach the center of the supercontinent. But because all land was connected, the newly evolved plants could spread easily from place to place such that by the time Pangaea broke apart, each new continent carried some plants, and no continent was left without any plants. The first splitting of Pangaea ran from east to west, creating two gigantic continents, Laurasia (North America, Europe, Russia, and China in the north) and Gondwana (Africa, South America, India, Australia, and Antarctica in the south). Imagine if Pangaea had broken apart before plants evolved from algae. Plants would have originated on one but not the other. On one continent, plants would have spread and diversified, and would have evolved into many clades. But the other continent would have remained barren until at some point millions of years later, some spores or seeds would have somehow blown or drifted to it (FIGURE 2-37).
A crucial cause of Earth being a changing world is life itself. Living organisms take up energy and nutrients from soil, water, and air and then release their waste products back. Plants take so much carbon dioxide out of the air that they have changed the atmosphere. And because carbon dioxide is a greenhouse gas, the changing levels of carbon dioxide affect the temperature of Earth’s surface—the biosphere—the place where life occurs. Photosynthesis produces oxygen as a waste product that is released into the air. Before the evolution of photosynthesis, Earth had no free oxygen at all; there was no aerobic respiration nor were there any animals. The photosynthetic production of oxygen is carried out by cyanobacteria, algae, and plants; without these organisms, animals and fungi could not exist. Similarly, as animals and fungi respire their food, they give off carbon dioxide, which of course is valuable for plants. Thus photosynthetic organisms have changed Earth such that it is a better habitat for organisms that respire, and vice versa, organisms that respire are a source of the carbon dioxide needed by photosynthetic organisms (FIGURE 2-38).

FIGURE 2-37 For most of us, it is difficult to imagine what Earth was like before the evolution of true plants: There was no life on land at all. For billions of years, life existed as bacteria and archaeans in the oceans; later there were fish and algae, but nothing at all on land until at the earliest, about 450 million years ago. Most of the surface was rock, and if any soil particles formed, they would have mostly been washed into the oceans because there were no plant roots to hold them in place. The area shown here is in Death Valley National Park, which does have plant and animal life, but it is widely scattered and confined to areas with at least occasional water.
Plants, animals, and all other organisms interact in more immediate, direct ways as well. Interactions may involve one organism benefiting by harming the other (this is predation), by both organisms helping each other (mutualism), by one organism benefitting without harming the other (commensalism), and by one organism harming another with no benefit to itself (amensalism) (TABLE 2-6). Herbivores and omnivores are examples of predation. They benefit by eating plants and this of course harms the plants, so plants need to defend themselves from animals. There is no one single optimal defense; instead many alternative types of defense have evolved, such as having spines or being poisonous (FIGURE 2-39). Pathogenic fungi and bacteria also prey on plants.

FIGURE 2-38 (A) The grass and herbs here capture energy and minerals, converting them into the organic molecules of the plants’ bodies. Herbivores such as deer eat the plants, using the organic compounds as their own food. The respiration of the animals returns carbon dioxide back to the air; the animal dung also is broken down by fungi, releasing carbon dioxide. Thus there is a cycling of carbon dioxide, minerals, oxygen, and other components. (B) In addition to large mammals such as deer, insects such as this caterpillar also consume immense amounts of plant material and are an important component of the carbon, nitrogen, and mineral cycles.
TABLE 2-6 Direct Interactions among Organisms

Animals often harm plants without benefiting from the damage they do; this is an amensal relationship. For example they trample plants as they walk across them or when they lie down. Here too it is beneficial for plants to somehow defend themselves.
The presence of trees often creates a shady, cool area in which smaller plants, such as mosses and ferns, can live. The shade-loving plants benefit without affecting the trees; this is a commensal relationship (FIGURE 2-40).
In mutualistic relationships, both organisms benefit. When animals pollinate flowers, both the plant and the animal benefit: The animal obtains nectar or pollen as food, and plants have their pollen grains carried from one flower to others, improving cross pollination and sexual reproduction. The evolution of fleshy, nutritious fruits is similar: Animals benefit by eating the fruits whereas plants benefit from having their seeds disbursed, often in a pile of “organic fertilizer” (FIGURE 2-41). These cases of mutualism differ fundamentally from those of predation and amensalism. In predation for example, the organisms are battling against each other. When an animal eats a plant, some surviving seedling of that plant might have a defensive mutation that might make it more spiny or poisonous than its parent was. If so, that seedling is more likely to survive and, over many generations, spininess or toxicity may gradually increase in that species of plant. But if so, then it will be helpful to the animals if they have mutations that allow them to overcome the plant’s new defenses. This battle between plants and animals has continued for the hundreds of millions of years that both have existed.

FIGURE 2-39 This pasture is being overgrown by prickly poppy (Argemone albiflora) because cattle and deer will not eat it even if starving. The leaves and stems are covered with spines; it contains a toxic milky juice as well. If there were no herbivores, the grasses might grow well enough to compete against the prickly poppy and drive it out, but with cattle eating the grass, the prickly poppies have a distinct advantage and might eventually take over the entire pasture.

FIGURE 2-40 Two trees have died and fallen across this stream, which keeps their wood moist enough for moss plants to grow on them. The mosses are benefitting from the presence of these dead trees but the trees are deriving no benefit at all from the mosses. Similarly, surrounding trees that are still alive provide shade that is beneficial to the mosses, but the mosses provide no benefit to the living trees either.

FIGURE 2-41 This pile of Texas lignum-vitae (Guaiacum angustifolium) seeds has passed through an animal, probably a javalina, so all the fruit pulp has been digested but the seeds are intact. Five have already germinated. The javalina probably ate the fruits far from this area and then defecated here, thus spreading the seeds away from the parent plant. Both the plant and the animal benefit in this mutualistic relationship.
But a completely new type of evolution began to occur as animal-pollinated flowers evolved. Because plants benefit when an animal carries its pollen to another flower of the same species, any mutation in the plant that helps the pollinator will be beneficial to both. Similarly any mutation in the animal that makes it a better pollinator helps both itself and the plant. This is called co-evolution and it causes certain plants and animals to become well-adapted to each other (FIGURE 2-42). Keep in mind that at the same time that the plant’s flowers are producing nectar that benefits a pollinator, the plant’s leaves may be producing poisons that keep herbivores away.
Plants interact with each other. Of course, plants never eat each other; instead plants compete with each other for water and minerals in the soil, for sunlight, and for the attention of pollinators and seed dispersors. In an oak forest, for example, trees may grow so close together that the roots of any one tree touch those of the surrounding trees and there is no room for a new oak seedling. Because all the trees are the same species of oak, they probably are almost identical genetically and each is about as good as any other at absorbing water and nutrients. Oaks are wind pollinated so they are not competing for pollinators. But as mentioned before, there are many ways of being adapted so there might be small wildflowers growing between the oaks, wildflowers that can tolerate growing in shade and whose small root systems will fit into any spaces between the roots of the oaks. Habitats are complex, so there may be rocky cliffs that are too dry for the oak trees but are just fine for junipers. In this forest, the oak trees compete with each other for water, minerals, and light; they make the habitat shady enough for wildflowers, and the junipers are using a resource (rocky cliffs) that oaks cannot.

FIGURE 2-42 Orchids have extremely complex flowers as the result of co-evolution between each species of orchid and one particular species of pollinator. The benefit of this to the plant is that a pollinator carries the orchid’s pollen only to other flowers of the same species; none is wasted by being accidentally carried to a different species of plant. On the other hand, if the pollinators die due to a disease or bad weather, the orchid then has no alternative means of transferring its pollen. This Phalaenopsis is popular in cultivation.
Because all organisms evolve, a set of organisms such as a forest never stays the same. At the same time, the physical environment also is changing, due both to geological processes and the effects of organisms. The biosphere is complex, dynamic, and changing. The plants, animals, geology, and climate that were present in the past were different than they are now, and those of the future will also be different. But for studying and understanding life, the process, principles, and concepts are always the same.
 At the Next Level
At the Next Level
1. Alternatives and consequences. A theme of this book is to ask “What are the alternatives?” and “What are the consequences of various alternatives?” Parasitic plants are discussed at several points because they often represent extremely unusual alternatives to the biology of non-parasitic plants. I recommend the book Parasitic Flowering Plants by Henning S. Heide-Jørgensen (2008, Brill Academic Publishers) because it provides many interesting examples of highly modified plants and is well-illustrated. Even as a beginning botanist, you will understand most of it and benefit from knowing how extreme certain alternative can be. You might find it difficult to believe that some of these are actually plants.
2. Evolution by natural selection. Charles Darwin and Alfred Russel Wallace discovered evolution by natural selection in the 1850s, and both evolution and natural selection are dominant themes in this textbook. Darwin wrote The Origin of Species in 1859 and it was the crucial introduction of basic principles to the world. However I recommend at this point that you read Darwin’s Voyage of the Beagle (1989, Penguin Books), which is his account of his trip and his collecting. It is written as a narrative and is wonderfully easy to read. Darwin wrote it shortly after he returned from his voyage, many years before he had figured out the principles of evolution by natural selection. As you read Voyage of the Beagle, you will have a better understanding of evolution than Darwin did when he was writing it. In certain parts of the book, it is fascinating to see how Darwin puts the clues together correctly and comes to key understandings using the scientific method. But in other parts, it is excruciating to read as he misses the clues and misses the point. Voyage of the Beagle is an excellent example of how the scientific method is used, how Darwin’s thought processes and reasoning finally allowed him to see the principle that is the basis of all aspects of biology.
SUMMARY
1. Almost all plants consist of stems, leaves, and roots. Flowers and cones are modified stems and leaves.
2. One of the defining characters of angiosperms (flowering plants) is that their seeds always develop inside a fruit.
3. The internal organization of plants is very uniform and simple; virtually all stems have epidermis, cortex, vascular tissues (xylem and phloem), and pith; roots lack pith; leaves consist of epidermis, mesophyll, and vascular tissues.
4. All plants have a primary body but only woody plants have a secondary body, which consists of wood and bark.
5. All organisms must take in energy and raw materials from their environment; they also give off waste products to the environment.
6. All known organisms store heritable information in DNA.
7. Plants detect and respond to information from the environment as well as from various organs within the plant itself.
8. A grade is a level of evolution, and it is an artificial group because various unrelated organisms might evolve to a particular level independently. A clade is a natural group because it contains a common ancestor and all the descendants of that ancestor.
9. There are multiple ways for an organism to be adapted to its environment; there is no single optimal solution to any biological problem.
10. The biosphere is constantly changing due to physical and geological forces, as well as to the presence of life itself.
11. Organisms interact with each other, often with animals and fungi consuming plants and plants needing to defend themselves. But some interactions, such as pollination, are mutually beneficial to both plants and animals. Plants also compete with each other for various resources.
IMPORTANT TERMS
angiosperm
basal angiosperm
biosphere
clade
common ancestor
cormophyte
cryptogam
dicot
embryophyte
eudicot
euphyllophyte clade
fern
Gondwana
grade
gymnosperm
herb
Laurasia
lycophyte clade
monocot
nonvascular plant
Pangaea
phloem
photosynthesis
plant
reproductive organ
respiration
seed plant
spermatophyte
spore-bearing plant
thallophyte
thallus
tracheophyte
vascular plant
vegetative organ
woody plant
xylem
REVIEW QUESTIONS
1. Name the three vegetative organs of a plant.
2. How do the reproductive organs of ferns differ from foliage leaves?
3. Name the four basic organs of a flower. Describe situations in which flowers might not need one or two of these organs.
4. Name the two vascular tissues found in vascular plants. What does each transport?
5. Are there any types of organisms that do not need to take in energy and raw materials from their environment? Discuss the source of energy for plants as opposed to animals; describe some of the raw materials that are needed by plants.
6. ______________________ is the central metabolism by which plants acquire energy; it occurs only when ______________________, ____________________, and ___________________ are present.
7. Each plant receives a great deal of information from its parents by means of the DNA in which two cells? Describe the types of information that plants receive from the environment. Why is it that environmental information cannot be stored in DNA?
8. The activity of the roots of a plant must be coordinated with the activity of leaves. Describe how this occurs. What type of information molecule is used?
9. Which three groups of plants are referred to as bryophytes? They differ from all other plants in two main features; name the features.
10. Which group of plants does the term “angiosperm” refer to? What are the three main subgroups of angiosperms? Do angiosperms have vascular tissue? Do they have seeds? In conifers, seeds develop inside cones, but in angiosperms, seeds always develop inside of _____________.
11. Evolution by natural selection causes organisms to become more adapted to their environment. Is the environment uniform across all of Earth? Does the environment change with time? Considering this, do you think that there is only one single optimum way for a plant to be adapted? If not, what is the alternative?
12. Have North and South America always been connected by Central America? Once the two continents joined, plants and animals could move from one to the other; animals can walk, fly or swim, but how does a plant species spread from one area to another? South America has thousands of plants adapted to the Amazon Rainforest; do you think it is likely that any of those plants will be able to migrate to North America across the deserts of Mexico, Texas, and Arizona?
13. The concept of biosphere was touched on only briefly in this chapter, but you have many resources in this book and online to explore this concept. What are some of the geological and biological components of the biosphere? We humans are burning fossil fuels and adding carbon dioxide to the atmosphere; how does this affect the biosphere?
14. Plants and animals usually battle each other; describe why and how this occurs. Once animal-pollinated flowers evolved, a new type of plant—animal interaction was possible; describe it. How is it possible for a mutation in a plant to be beneficial for both the plant and for an animal?
15. Earth is constantly changing, and so are all organisms as they evolve. But despite so much change, it is possible to study and understand life because several things are always constant; what are those things?
Design Credits: Hummingbird: © Tongho58/Moment/Getty; Green plant cells: © ShutterStock, Inc./Nataliya Hora; Purple tulip: © ShutterStock, Inc./Marie C Fields; Dandelion: © ShutterStock, Inc./danielkreissl; Poppy: © ShutterStock, Inc./Saruri; Plant icon: © ShutterStock, Inc./Vector; Digging man icon: © ShutterStock, Inc./Z-art