Botany: An Introduction to Plant Biology - Mauseth, James D. 2017
Ecology
Populations and Ecosystems

Chapter Opener Image: All organisms are sensitive to the conditions in their environment. The area shown here is at such a high elevation in Rocky Mountain National Park that trees are right at their limit of survival. The higher elevations on the upper right side of the image are so cold and stressful that trees cannot survive at all, whereas nearby on the lower left, conditions are sufficiently mild that trees achieve almost their full height. The concept of “stressful” must be evaluated for each species: Every part of this area is too stressful for any tropical plant, but every part is suitable for the grasses that grow abundantly between the trees here. And most of the area in the foreground is too hot for the dwarf alpine plants that thrive at even higher altitudes on the mountaintops in the background.
OUTLINE
✵ Concepts
✵ Plants in Relationship to Their Habitats
- Abiotic Components of the Habitat
- Biotic Components of the Habitat
✵ The Structure of Populations
- Geographic Distribution
- Age Distribution: Demography
r- and K- Selection
✵ The Structure of Ecosystems
- Physiognomic Structure
- Temporal Structure
- Species Composition
- Trophic Levels
Box 25-1 Plants and People: Niches in the Jet Age
LEARNING OBJECTIVES
After reading this chapter, students will be able to:
✵ Compare an organism living in isolation to an organism living in a population.
✵ Define abiotic and biotic habitat components.
✵ Provide examples of how each of the following abiotic components affects organisms: climate, soil, latitude and altitude, and disturbance.
✵ Provide examples of how each of the following biotic components affects organisms: the plant itself, other plant species, and organisms other than plants.
✵ Describe various interactions between organisms such as competition, mutualism, and commensalism.
✵ Recall two of the plant competition theories.
✵ Define a limiting factor and describe how it affects a plant.
✵ Explain generation time, intrinsic rate of natural increase, and carrying capacity, and describe how they affect population growth.
✵ List four common ecosystem structures.
✵ Describe the flow of energy and carbon through the various trophic levels of an ecosystem.
 Did You Know?
Did You Know?
✵ All the members of a species that live close together constitute a population.
✵ Whereas an individual has certain characters (it is short or tall, resistant to disease or not), a population has all the characters of all its members.
✵ The members of populations harm each other (they compete for resources) and help each other (together they provide enough nectar and pollen to support a population of pollinators).
✵ Just as human populations have an age structure, some members of plant populations are very young, some are old, many are middle-aged.
![]() Concepts
Concepts
Ecology is the study of organisms in relationship to all aspects of their surroundings. Throughout this book, emphasis has been placed on the importance of analyzing structure, metabolism, and diversity in terms of adaptation and fitness. Mutations result in new alleles that alter the phenotype; then natural selection eliminates those that are less well adapted but retains those that increase fitness. Most factors have been discussed individually—for example, the effect of trichomes in deterring insects from chewing leaves and the ability of carotenoids to protect chlorophyll from excess sunlight. But most factors that affect a plant’s health and survival are present together in the habitat, acting on the plant simultaneously. We must try to understand the plant in relation to its entire habitat, to all components of its surroundings.
An individual plant never exists in isolation in a habitat; instead, there are other individuals of the same species and together they constitute a population (FIGURE 25-1). Individuals of plant populations often do not interact strongly, but the biology of the population is more than just the sum of all the biologies of individuals. In a species that consists of either dioecious or self-sterile individuals, a population can carry out successful sexual reproduction, but an individual cannot. If pollinated by animals, a single individual usually cannot produce enough nectar or pollen to keep even one pollinator alive, but a population of plants can sustain a population of pollinators. On the negative side, a population may be dense enough that spores from a pathogenic fungus are ensured of landing on at least one susceptible individual and then spreading to others, whereas if an individual could exist in isolation far removed from others, it might be safe from pathogens, predators, and natural catastrophes such as fires (FIGURE 25-2).
A population also does not exist in isolation, but rather coexists with numerous populations of other plant species as well as populations of animals, fungi, protists, and prokaryotes. All of the populations together constitute a community, which when considered along with the physical, nonliving environment is an ecosystem. These contribute additional levels of interactions and complexity and make it more difficult to be precisely certain of the effects of factors on individuals. The presence of trichomes may deter leaf-eating insects, but those insects might have frightened away more damaging egg-laying insects. Besides, leaf-eating insects might have had a beneficial role by dropping nutrient-rich fecal pellets to the soil where they might have encouraged growth of mycorrhizal fungi. The structure, metabolism, and diversity of plants cannot be fully understood without understanding the ecosystem.

FIGURE 25-1 The numerous individuals of one species constitute a population. They depend on each other for gametes during sexual reproduction and also support a healthy population of pollinators and seed dispersers. Because there are several species, the photograph shows a community of several populations. When the soil and atmospheric components are added, an ecosystem results.

FIGURE 25-2 It is more economical to cut trees growing in dense populations. Identical trees in sparse populations will survive.
Populations are rarely stable; change is much more common. The two photos in (FIGURE 25-3) show dry grassland in West Texas that experienced a wildfire, the Rock House Fire. It started on April 9, 2011, west of the small town of Marfa, and strong winds spread it rapidly northward for almost a month before it could be contained. There had been good rains the previous year, so grasses had grown large and thick and provided abundant fuel for the fire (Figure 25-3A). Consequently, the fire was unusually hot and burned many trees, cacti, agaves, yuccas, and other native plants (Figure 25-3B). If the previous year had been dry, there would have been less grass and the fire would have either been cooler, or it might not have been able to spread and many more plants would have survived. If the native mammals—buffalo—were still here, they would have eaten the grass, removing fuel. The buffalo, however, was hunted almost to extinction in the mid 1800s so that trains could run without hitting the animal on the tracks. Many plants survived the fire; most of the yuccas (Figure 25-3B) are alive and should recover in 2012, if there are adequate rains. The ecology of an area is a complex interplay of many factors, some of which change from year to year.
![]() Plants in Relationship to Their Habitats
Plants in Relationship to Their Habitats
The habitat is the set of conditions in which an organism completes its life cycle. For migratory animals, the winter area, summer area, and migration routes are all habitat components. No plant is migratory, but portions of plants are; spores, pollen, fruits, seeds, and vegetative propagules move through the habitat.

FIGURE 25-3 Grassland in West Texas before (A) and after (B) the Rock House Fire in 2011.
How much of the surroundings should be considered part of the habitat is debated. Many factors do not appear to affect certain plants at all. Small herbs on the forest floor do not seem to influence large trees; their presence or absence has little effect on the mineral nutrition of the trees or on their pollination (FIGURE 25-4). An experiment might remove all of the small annuals and then examine whether the trees are affected; however, it would be difficult to measure the growth of entire trees, especially their roots, and it might take years to see any effect on the trees’ metabolism. The small herbs may be important; perhaps they harbor spiders that catch insects that would otherwise kill the trees’ seedlings. If so, then removing the herbs might cause an increase in the insect population and decreased survival of tree seedlings; with fewer seedlings of this species, it might be possible for the seedlings of a different species to survive better. After many years, the forest composition would be changed as a result of the removal of the herbs.
On the other hand, we do know that many components impact others directly. Pollinators are critically important aspects of the habitat for the plant species they pollinate, and any disease organisms or predators that prey on those pollinators are also important to the plant. Aspects of the habitat that definitely affect a plant constitute its operational habitat, whereas all components, whether with known effect or not, are its habitat.

FIGURE 25-4 The redwoods here completely dominate the habitat, each tree containing many times the bulk and volume of any single herb. They provide filtered light and protection from wind for the understory plants, which are incapable of tolerating full sunlight. Do the herbs affect the trees in any way? Yes, the redwood seeds germinate only after a low, cool fire is fueled by the understory herbs and shrubs.
For example, consider the redwood forests in California. It was discovered that there were no redwood seedlings in the national parks, and redwood seeds were not germinating.
After a recent forest fire, redwood seeds sprouted and grew vigorously. The fire not only stimulated the seeds to germinate, but by burning the understory plants, released minerals that increased soil fertility. We now realize that shrubs and herbs are vital to the success of the giant redwoods: They fuel quick, cool fires that are too low to damage the large redwoods but are necessary for seed germination. The policy of preventing forest fires—a natural factor of this ecosystem—was harming the redwoods.
Habitat components are of two types, abiotic and biotic. Abiotic components are nonliving and are physical phenomena: climate, soil, latitude, altitude, and disturbances such as fires, floods, and avalanches. Biotic components are living factors: the plant itself, other plant species, and species of animals, fungi, protists, and prokaryotes.
Abiotic Components of the Habitat
Climate
Climate is critically important to all organisms; most species are restricted to certain regions primarily because they cannot live in climatic conditions outside those regions. Climate itself has many components—temperature, rainfall, relative humidity, and winds being just a few.
The average temperature of a habitat is not as important as its extremes: the lowest winter temperature and highest summer temperature. Many species of bromeliads, aroids, and orchids are restricted to the tropics because those are frost-free habitats. Rain forests along the west coast of California, Oregon, and Washington receive adequate rain, but freezing winters prevent them from being suitable habitats for most tropical species. On the other hand, many temperate trees must have a winter dormancy period accompanied by weeks of subfreezing temperature in order to be vernalized and bloom. If cultivated in areas with warm winters, the plants grow and survive well but do not reproduce. The highest or lowest temperatures during a plant’s lifetime are important. If a tree species must be 10 years old before it can flower but killing temperatures occur every 8 or 9 years, the species cannot survive there.
The growing season of an area is often determined by the date of the last severe, killing frost in the spring and the first killing frost of autumn. The length of the growing season must be adequate for sufficient photosynthesis, growth, development, and reproduction; if not, a reproductive population cannot survive even if it can tolerate the temperature extremes of winter and summer.
Moisture occurs as rain or snow or as hail that supplies water but also damages leaves, buds, flowers, and animals. Habitats range from extremely dry (deserts) through progressively moister all the way to marshes, lakes, and rivers that are virtually all water. Just as with temperature, the total amount of precipitation may not be as important as seasonal extremes or the timing of precipitation. A constant drizzle that occurs almost year round supports certain types of ecosystems, whereas the same amount of rain, distributed as just winter snowfall and occasional summer thunderstorms, results in a different type of ecosystem.
Numerous metabolic processes respond proportionally to abiotic factors. Once there is sufficient moisture for marginal survival, increased amounts of water produce increased growth and reproduction. There is usually an upper limit: With too much water, roots drown for lack of soil oxygen (FIGURE 25-5A). Between the low and high extremes is the tolerance range of the organism. Ranges vary greatly from species to species (FIGURE 25-5B). Some are extremely broad. Most temperate plants across the northern United States and Canada, especially in the midwest, tolerate summer highs of over 100°F (37.8°C) and winter lows below —40°F (—40°C). Plants of southern Florida, Puerto Rico, and Hawaii tend to have much narrower tolerance ranges, being killed by both cool temperatures and hot ones.
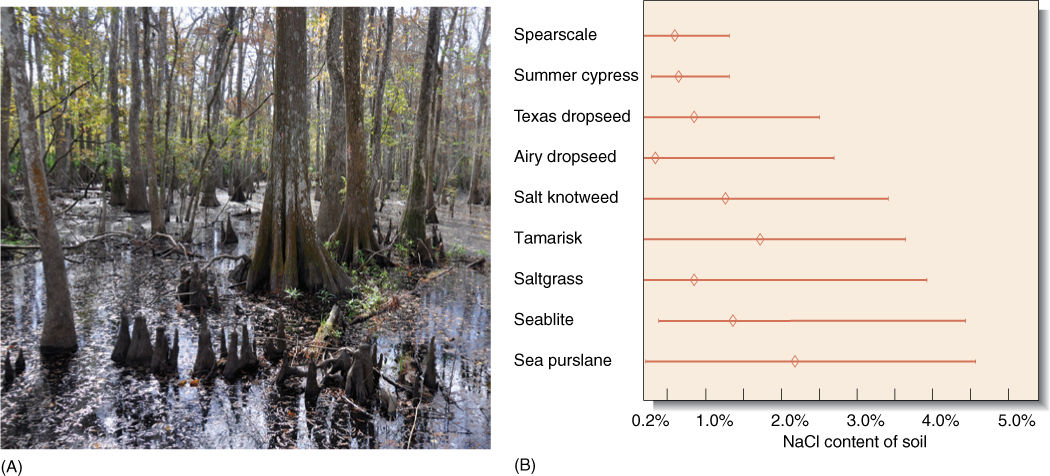
FIGURE 25-5 (A) Swampy bayous of southern Louisiana contain so much water that most plants cannot grow; their roots drown. Bald cypress (Taxodium) is well adapted, however; its roots form “knees” filled with aerenchyma that allow atmospheric oxygen to diffuse into the submerged roots. (B) Plants vary in their ability to tolerate salty soil. These species occur around salt flats in Oklahoma and Kansas; sea purslane and seablite survive even in the presence of strong salt concentrations, whereas others tolerate less. By being tolerant of soil salt, these plants can occupy habitats on the edges of salt flats, which are free of other plants. Not shown are the many species that have virtually no salt tolerance. Diamond marks indicate the optimal concentrations.
Soil Factors
Soils are formed by breakdown of rock. Initially, the resulting soil is thin and virtually identical to the parent rock in its chemical composition; consequently, young soils are variable in the amounts of macronutrients and micronutrients they have available. Because nitrogen is not a significant component of any type of rock, all young soils are deficient in it.
The first plants that invade a new soil, called pioneers, must tolerate severe conditions. The soil is sandy, with relatively large particle size, and most minerals are still locked in the rock matrix. The soil has little water-holding capacity, and the first plants have no neighbors to help moderate wind, provide transpired humidity, or otherwise temper the environment (FIGURE 25-6). Pioneer plants often are associated with nitrogen-fixing prokaryotes; many lichens contain cyanobacteria, and some angiosperms have root nodules. As pioneers live on the soil, they change it significantly; carbon dioxide from root respiration produces carbonic acid and accelerates chemical weathering. Dead plant parts such as leaves, fruits, roots, and bark become substrates for soil organisms, and their decay contributes humus, greatly increasing the soil’s water-holding capacity. Roots may penetrate to the bedrock, entering larger cracks and then expanding and breaking the rock physically.

FIGURE 25-6 This “soil”—a recent lava flow—consists mostly of rather large rock fragments with little water-holding capacity and very few dissolved essential elements. The plants growing here, pioneers, not only tolerate these conditions but actually change them. The acids they release as they decompose greatly accelerate chemical weathering and soil formation.
After many years, a thick soil may result that has a distinct soil profile with three layers or horizons (FIGURE 25-7). The A horizon is uppermost and is sometimes called the zone of leaching; it consists of litter and debris, and as this breaks down, rainwater washes nutrients from it downward into the next layer, the B horizon, or zone of deposition. The B horizon is the area where materials from the A horizon accumulate. It is rich in nutrients and contains both humus and clay. Below is the C horizon, composed mostly of parent rock and rock fragments.
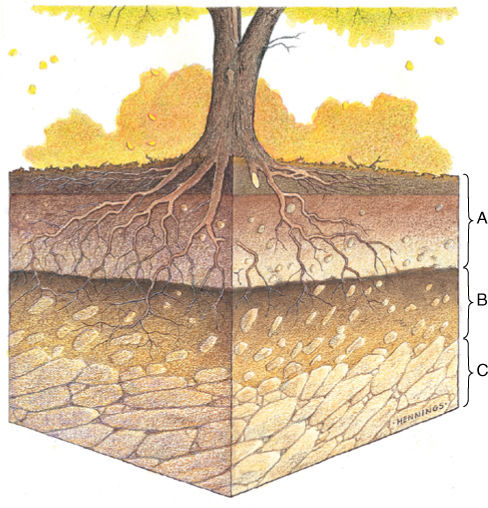
FIGURE 25-7 Most soils show three horizontal layers, or horizons. The relative thicknesses of the A horizon (zone of leaching) and B horizon (zone of deposition) depend on many factors, the abundance of vegetation and humus being especially important.
Whereas young soils differ because of the chemical nature of their parental rock, older, more mature soils are less diverse. As the rock weathers, essential elements are absorbed by roots and become trapped in the plant body; nonessential elements are leached away. As the plants or their parts die, they decay slowly, releasing the essential elements that re-enter the soil. There they are taken up again. As a result, essential elements cycle repeatedly, alternating between organisms and the A and B horizons, whereas other elements are gradually washed downward into the water table and are removed by underground water flow.
Latitude and Altitude
Latitude contributes many factors to the abiotic environment. At the equator, all days are 12 hours long; no seasonal variation occurs, and plants cannot measure season by photoperiod. At progressively higher latitudes to either the north or south, summer days become progressively longer, as do winter nights (FIGURE 25-8). Above the Arctic and Antarctic Circles, mid-summer days are 24 hours long, as are mid-winter nights. At intermediate and higher latitudes, day length is an excellent indicator of season, and some species are sensitive to photoperiod.
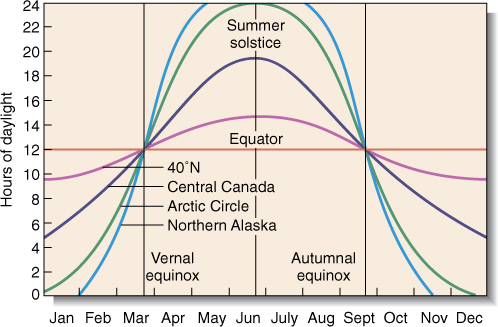
FIGURE 25-8 The line for day length at 40 degrees north, which corresponds to the center of the United States, shows that the longest day (summer solstice, about June 21) is just less than 15 hours long, whereas the shortest day (winter solstice, about December 21) is only about 10 hours long. Closer to the equator, the difference between the longest and shortest days is less, and near the equator the difference is too little for plants and animals to be able to use it as a seasonal indicator.
The amount of light energy that strikes a given area of Earth’s surface each year also varies with latitude; in equatorial regions, the sun is always nearly overhead, and each square meter of surface receives a maximum amount of radiation (FIGURE 25-9). At higher latitudes, the sun is only rarely overhead, usually only near midsummer. At other times, when the sun is low, light strikes the Earth obliquely, and less energy is received per square meter; even at noon on winter days the sun is low in the sky and days are not bright. At high latitudes, temperatures fluctuate greatly on both a daily and seasonal basis. Soil formation is slow in the cold latitudes, and often, what soil has formed is blown away by strong winds. In much of Alaska and Canada, soil was scraped away during the ice ages 18,000 years ago.
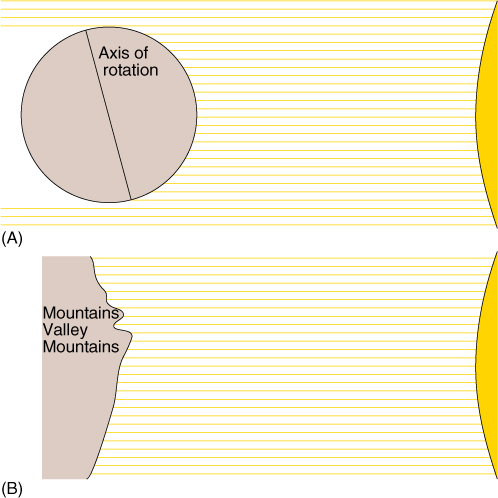
FIGURE 25-9 (A) During winter in the Northern Hemisphere, the North Pole is pointed away from the sun, and the amount of energy received per square meter is at a minimum. The South Pole points toward the sun, and the Southern Hemisphere receives more direct light, resulting in a large amount of energy per square meter.
Regions of high altitudes on mountain tops are similar to those at high latitude. There are high winds and poor soil; much or all the year is cold, and the growing season is short. Water may be present mostly as snow and ice, and thus, physiological drought is frequent. The areas may have varying day lengths, depending on their latitude. An additional stress present in high-altitude habitats is intense ultraviolet light. High altitudes are above much of Earth’s atmosphere and thus are not fully shielded by ozone, oxygen, carbon dioxide, and water vapor.
Disturbance
Disturbances are phenomena such as fires, landslides, snow avalanches, and floods; they produce a significant, often radical change in an ecosystem quickly (FIGURE 25-10). Disturbances affect the biotic factors directly, often completely eliminating many or all individuals from an area and also altering the soil, but they have little or no impact on other abiotic factors such as climate, latitude, or altitude. The elimination of large numbers of individuals by a disturbance alters species relationships in the ecosystem. Man-made disturbances have been caused by insecticides, herbicides, hunting, and habitat destruction (FIGURE 25-11).
Fire is a natural, common component of many dry ecosystems. With little moisture, fallen leaves and twigs decay so slowly that a thick layer of debris builds up. Living plants tend to have waxy cuticles and water-proofing resins that make them especially flammable. Lightning storms often occur without rainfall, starting fires that burn rapidly and cause great destruction. Many plants and animals of such ecosystems have become fire resistant as a result of natural selection caused by frequent fires. The bark of certain species of pine trees is so thick that a rapidly moving, moderate fire does no damage to the vascular cambium and other living tissues; the lower portions of the trunk have no branches because of self-pruning, so flames cannot reach high enough to ignite needles; only herbs and small shrubs are burned. Furthermore, seed cones of lodgepole pine and jack pine open only after being exposed to the heat of fire; this adaptation results in the release of seeds after many competing plants and predatory animals have been killed and the forest floor is open and sunny. Also, the soil is enriched by minerals in the ash, making it an ideal site for pine seedlings. However, if fires do not occur frequently enough, usually because firefighters put them out, understory shrubs and small trees grow tall and large; so much brush and dead wood accumulate that when fire does occur it is extremely hot. As it burns to the top of shrubs and understory trees, flames may reach the lowest branches of the pines, igniting the crowns (FIGURE 25-12). Once this occurs, the fire can spread rapidly through the canopy of trees, killing them.

FIGURE 25-10 (A) Fires kill not only plants but also pollinators, herbivores, and pathogenic fungi and bacteria. Fire also releases minerals back to the soil and decreases shading. A quick, cool fire does not damage rhizomes, tubers, seeds, or trees with thick bark, but an intense, hot fire can kill most organisms, sterilizing the soil. (B) Within a few months after a moderate fire, mosses, grasses, and wildflowers have begun regrowth. (C) and (D) The eruption of Mt. St. Helens radically altered the surrounding ecosystems, but there too plants, animals, and other organisms are recovering.

FIGURE 25-11 Humans destroy habitats on a massive scale; hydroelectric dams permanently flood valleys upstream and eliminate the flooding that had been a natural part of the downstream habitat. Spawning grounds for fish are also destroyed. A few small dams are now being removed in an attempt to restore river ecology.
Six months later the positions are reversed. (B) The variability of energy received per square meter of surface is affected by topography. Away from the equator, the side of a mountain or valley that faces the equator also faces the sun and receives more energy. The sides that face the poles are cooler. Very often, the two sides of a valley running east to west have different vegetation.
Many grasses have adaptations that permit them to benefit from fires. Many prairie grasses in the midwest and the saw grasses of the Florida everglades grow in dense clumps with their shoot tips and leaf primordia at or below ground level, protected from fire by soil and the living bases of leaves. Leaf tips may be dead and dry, and when fire occurs, the dead portions burn, releasing their minerals, but the bases of the plants are unharmed.
Annuals and short-lived plants do not survive fires, but their seeds, buried underground, do. Bulbs, rhizomes, tubers, or corms easily survive small fires.
Biotic Components of the Habitat
The Plant Itself
An individual itself, just by being in a habitat, modifies the habitat and is a part of it. Habitat modification may be beneficial, detrimental, or neutral to the continued success of that species in its own habitat. In beech/oak forests of the northern United States, the trees modify the habitat by producing a dense canopy that results in a heavily shaded forest floor (FIGURE 25-13A). With such low light levels, few seedlings grow well, but two that do are those of oaks and beeches. As a result, mature trees create a habitat that suits their seedlings and aids their own successful reproduction.

FIGURE 25-12 (A) If fire occurs frequently, understory shrubs are burned back before they become tall, and there is always a large space between them and the lowest branches of the dominant trees. Fire cannot get high enough to ignite the tree canopy. (B) If fire occurs infrequently, understory shrubs become tall, reaching the lowest branches of the dominant trees. Fire then burns upward from the shrubs into the canopy, igniting highly flammable needles, twigs, and cones. Even if the trees are not killed outright, their shoot meristems are destroyed, and no further growth is possible. (C) and (D) In most deserts, native grasses and herbs are so sparse that fire cannot occur, but introduced exotic herbs and grasses occasionally form a population dense enough to support fire. This has occurred in Arizona, putting cacti and yuccas at risk.
Pine forests are more open, but still the forest floor is shaded by mature pines. Pine seedlings, however, need full sunlight and do not grow well below the canopy of older pines. Seedlings of other species flourish in these conditions and crowd out the few pine seedlings that may occur. Pine trees modify their habitat adversely for their continued success; only disturbances can create the open habitats needed for pine seedlings (FIGURE 25-13B).
As glaciers retreat, they leave behind moraines—great mounds of rubble, sand, and boulders. The soil is poor, with no humus and few available nutrients, but pioneer species such as alder, Dryas (in the rose family), willow, and fireweed colonize recently exposed moraines (FIGURE 25-14). They survive in the open conditions and tolerate low levels of nutrients; alders and Dryas have root nodules containing symbiotic nitrogen-fixing bacteria that supply nitrogen. Within a few years, decay of their leaves and bark has enriched the soil sufficiently that Sitka spruce and western hemlock become established. After this, however, the spruce/hemlock forest creates too much shade and eliminates the pioneers that have altered the habitat to their own detriment by enriching it.
Other Plant Species
When several individuals, of either just one or several species, occur together, the possibility for interaction is created. If the interaction is basically beneficial for both organisms, it is described as mutualism, but if it is disadvantageous, it is competition. Competition is a situation in which two populations do not grow as well together as they do separately because they use the same limited supply of resources. Many plants are believed to compete with others for light, soil nutrients, water, and the attention of pollinators and seed dispersers, among other things. If a single plant were allowed to grow by itself, in many cases, it might grow more rapidly, become larger, and produce many more gametes than it would if other plants were nearby. Roots of other plants might grow among its roots and remove water and nutrients. The competitors might grow taller than it does and intercept sunlight by putting their leaves above its leaves. Their flowers, even if they did not produce more nectar, might still distract its pollinators such that its pollen would be carried to the stigmas of the wrong species and its own stigmas would receive foreign pollen.

FIGURE 25-13 (A) In this forest, the two dominant species, beeches and oaks, alter the habitat so that it is suitable for their own seedlings. (B) Although much more open and sunny than a beech/oak forest, a pine forest is still too shaded for pine seedlings. Pines seem to alter the habitat adversely for their own long-term survival, but the needles they drop are highly flammable and are the main cause of frequent fires that kill oak seedlings but not pine seedlings.

FIGURE 25-14 As a glacier retreats, the rubble and sand left behind is extremely poor soil, and of course, the climate is usually harsh; however, several species of pioneers can grow here, and their activity enriches the soil, permitting invasion by less hardy species.
The role of competition has been extensively debated. One theory postulates that the result of competition is competitive exclusion: Whichever species is less adapted is excluded from the ecosystem by superior competitors. The species that get sunlight and other resources win; those that do not lose and are eliminated. If this is true, then very little competition occurs in a typical ecosystem; each species is assumed to be adapted to a particular set of conditions, a niche, that no other species is adapted to use as efficiently. For example, some species are adapted to full sunlight and others to partial shade (FIGURE 25-15). In an ecosystem, the former must be a canopy tree and the latter an understory species. This theory of little competition predicts that if certain species are removed, the others do not benefit from the unused resources because they are not adapted for them. This is sometimes found to be the case.
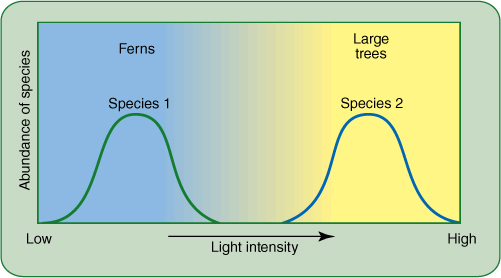
FIGURE 25-15 Both ferns and large trees need light, but many fern species cannot tolerate high light intensity and must have partial shading. Large trees often must have intense light; they do poorly if shaded. Growing together, they do not compete for sunlight; each uses a portion of the resource the other does not use. If either were removed, the other would not grow better.
The concept of niche is difficult to define exactly; basically, it refers to the set of aspects of the habitat that directly affect a species. For example, a particular marsh species occupies a particular semiaquatic niche defined by a range of soil moisture or flooding, a range of seasonal rainfall, a range of temperature, a paucity of root grazing caused by lack of swimming herbivores, and the presence of appropriate pollinators. Another species of marsh plant may grow in the same general geographical area, even the same marsh, but occupy a different niche because it may have a different pollinator or may grow in areas of the marsh that are slightly more acidic than the microhabitats occupied by the first species. As long as even one factor differs, they occupy different niches.
As an alternative to the theory of competitive exclusion, a second theory postulates that species overlap in their tolerance ranges (FIGURE 25-16A), and when grown together, each has exclusive use of the portion of the range not used by the other. In the overlap zone where the habitat is suitable for both species, usage is determined by competition: The one that is more adapted occupies the overlap zone exclusively. The weaker species occupies only part of its potential niche. The two can coexist, but if either is removed, the other then has its full range of resources and grows better. Examples of this type of competition have been found to occur.
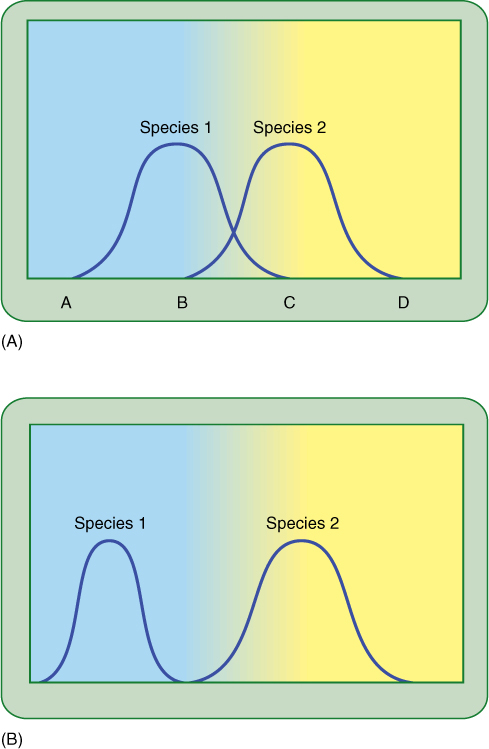
FIGURE 25-16 (A) Two species compete if they can use the same resource. Here, species 1 would have exclusive use of the resource under conditions A to B, whereas species 2 would have exclusive use between C and D. With conditions between B and C, they would compete. If the resource were water availability, species 1 might grow in drier areas and species 2 in wet areas, and both could grow in moist areas. (B) Over time, mutations that improve the ability of a species to compete may be selected. In this case, species 1 evolves such that it is specialized in dry habitats and now cannot live in moist conditions. This may increase its total fitness because moist environments have more insects and fungi that require expensive defense mechanisms, mechanisms that it no longer needs.
A long-term result of competition should be species modification by natural selection. If two species compete with partial tolerance-range overlap, then mutations are beneficial if they allow each to use more efficiently its exclusive portion of the range. Through time, the species diverge until competition is reduced (FIGURE 25-16B).
The geographic ranges of most populations are extensive enough that they contain a diversity of biotic and abiotic factors: Each may include hills, valleys, and plains; rocky soil and rich soil; and grasslands and open woodlands. Because of this diversity, a population may be competing for sunlight in one part of its geographic range but competing for water in another and for other factors in still another. Ecosystem diversity causes different subpopulations to specialize for certain features, especially if gene flow is not rapid and thorough. This could be the beginning of divergent speciation, of course, but long before the various subpopulations could be called subspecies, they would be considered ecotypes, each specialized in response to particular ecosystem factors at its locality. The various ecotypes resemble each other so strongly that they clearly belong to the same species but have enough differences for ecologists and taxonomists to suspect that separate ecotypes exist. Experiments are necessary to prove that natural selection is in fact altering the genome because phenotypic differences can result simply from the growing conditions themselves. For example, beans grown in rich soil with adequate water are strikingly different from those grown in poor soil with little water and other stresses. To test whether ecotypes really exist, transplant experiments are performed: Plants from each site are transplanted to the alternate site, and plants from both sites may be grown together in a common garden at an intermediate site. If the transplanted individuals take on the phenotype of the naturally occurring plants at that site, then genetic factors were not important, just stresses; however, if the transplanted individuals retain their original phenotype or if they die, then genetic divergence had begun.
Also, success in competition depends on factors other than the ones involved in the competition. Species A may grow rapidly and shade species B when water is abundant, but species B may be more drought tolerant and can outgrow and outcompete species A when water is scarce. Water availability may vary over the geographic range such that A dominates in wet areas and B in dry areas. Also, because rainfall varies from year to year, species A may be more common some years and species B more abundant in other years.
Animal ecology includes a variety of negative interactions in addition to competition; predation—the killing of prey—and parasitism are two examples. Insectivorous plants do trap and kill insects, but no plant kills and digests other plants in a predator—prey relationship. Parasitic plants are not uncommon. Mistletoes are familiar examples, and hundreds of species of parasitic plants in at least 12 families are known.
Numerous animals, fungi, protists, and prokaryotes are saprotrophs, living on dead organisms; vultures, maggots, wood-rotting fungi, and decay bacteria are examples. No plant species of any type is known to attack and digest dead plant or animal material; roots do absorb the minerals, nitrates, and sulfates released from decomposing humus, but the roots do not participate in the enzymatic attack.
Organisms Other Than Plants
Animals, fungi, and prokaryotes are obviously important biotic aspects of a plant’s habitat. Interrelationships between plants and these other organisms can be either beneficial or detrimental for one or both partners.
Plants and animals have many relationships that are examples of mutualism, in which both species benefit. Most instances of pollination are mutualistic—the animal receives nectar or a portion of the pollen and the plant benefits from pollen transfer. Seed dispersal by fruit-eating animals (frugivores) also benefits both species as long as the animal does not chew the seeds and digest the embryos.
A famous example of mutualism is the association of certain ants and acacias in tropical Central America. Acacias are small trees that have enlarged, hollow thorns at their leaf base. The thorns are used as ready-made, stout, waterproof homes by ants (FIGURE 25-17). Because of the large number of thorns on a single plant, the ants benefit from having abundant housing, but in addition, acacias produce nectar, and their leaflet tips are modified into golden yellow food bodies (called Beltian bodies) that are filled with glycogen. The ants receive both housing and food. The plant benefits because this species of ant is aggressive; they patrol the plant and attack any animal that touches it, even large mammals. If the leaves of another plant come close to an Acacia, the ants destroy the leaf, keeping the Acacia unshaded. As a result, the plant is free of pathogenic fungi and insects, and it grows in full sunlight. This mutualism is obligate; the plant cannot survive in nature without the ants, and the ants are not found away from the plant. A similar mutualism occurs between ants and Hydnophytum in Southeast Asia.
Commensal relationships, in which one species benefits and the other is unaffected, are also common between plants and animals. When birds build nests in trees, the birds benefit, and the tree is (usually) unharmed. When sticky fruits or seeds, such as cocklebur, stick to an animal’s fur or feathers and then are dispersed, the plant benefits and the animal is unharmed. One-sided negative relationships also occur: Animals trample and kill small plants without being affected. Competition, in which both parties are adversely affected, probably is not common between plants and animals.
Predation is a relationship in which one species benefits and the other is harmed (FIGURE 25-18); the species that benefits seeks out the other and uses it specifically for food or some other form of resource. Animals that eat plants are herbivores and the process is herbivory, but it is often more precisely delimited as browsing (eating twigs and leaves of shrubs—deer, giraffes) or grazing (eating herbs—sheep, cattle). Insects also lay eggs in plants, and then their larvae feed on plant tissues as they tunnel through them. Aphids and spittlebugs suck sap rather than chew tissues. Insects and birds both harvest leaves and twigs for nest construction.
Many interrelationships between plants and fungi or bacteria are harmful to the plant, but the fungi and bacteria are described as being pathogenic rather than predatory. Either may cause mild disease or be so virulent that they kill the plants quickly. A large percentage of the microbes are saprotrophs, living on dead plant tissues such as leaves, logs, fallen fruit, and sloughed bark. This benefits the fungi and bacteria without harming the living plants, and it can actually help them. This process speeds up the release of mineral nutrients, especially nitrogen compounds, enriching the soil. A case of plants attacking and parasitizing fungi may be known. Orchid seeds are tiny and lack chlorophyll; they remain moribund until invaded by soil fungi, then they turn green and grow well. It had been assumed that the seedling received nutrients and perhaps growth factors from the fungus and in return provided it with carbohydrate; however, tests have not revealed any benefit to the fungus, just to the plant.

FIGURE 25-17 (A) Ant Acacia plant. (B) These thorns on an ant Acacia are swollen and hollow and serve as excellent nests for ants. They form as part of the Acacia’s own normal development; the presence of ants does not induce their formation. (C) The leaf tips of ant acacias develop as Beltian bodies (yellow), food bodies rich in glycogen. As long as an Acacia is occupied by ants, all other vegetation is kept cleared away, but if the ants are killed with insecticide, the acacias are overgrown quickly.
Plants and People
BOX 25-1 Niches in the Jet Age
A niche is defined as a set of conditions in which a particular species can thrive, outcompeting other organisms. Cacti occupy desert niches in North and South America, and water hyacinths proliferate in the waterways of the Amazon rain forest, filling the niche consisting of calm water surfaces in warm areas. But the desert niches occur in many geographical regions other than the western hemisphere, and tropical rivers flow quietly all around the world. Why do cacti and water hyacinths grow only in restricted areas? Basically, it is simply because they have not been able to spread to the other areas because their natural range—the site where they originated evolutionarily—is surrounded by vast regions of inhospitable barriers. Under natural conditions, cacti could establish themselves in the desert niches of Africa, the Middle East, China, and Australia only if migratory birds happened to carry seeds from one continent to another. Such long-distance dispersal is extremely rare.
Perhaps we should say that it was extremely rare. Humans have now far surpassed migratory birds as agents of long-distance dispersal. Planes and ships connect all parts of the world, causing an interchange of plant and animal species on an unprecedented scale. Some of the transport is accidental as seeds are caught in clothing of tourists or are attached to hides or other material being shipped. Zebra mussels came to the United States in the bilge water of ships, and insects are carried in fruit and produce and even in the stagnant water trapped in old tires being imported for recycling. Of course, even microbes and viruses are transported over long distances, as is the case with HIV, the virus that causes AIDS. Other transport is intentional, as living plants or animals are imported specifically for cultivation as crops or ornamentals. Carp were imported into our rivers as a food source; killer bees were brought to Argentina from Africa for research and then escaped from a broken laboratory cage. The vine kudzu was planted in the southern states as a ground cover to control erosion along road cuts and on canal banks, and water hyacinth apparently was introduced into our waterways simply because it is pretty.
All of these organisms, and many more, share a common feature—they have thrived in the new habitats in which we have placed them. All have found conditions that permit their rapid growth and reproduction. Because we did not bring along their natural predators or pathogens, they tend to be free of the organisms that could limit their expansion. They have all undergone population explosions and are proliferating rapidly. Zebra mussels are clogging waterways through the American northeast and are spreading down the Mississippi River; killer bees have spread throughout South America, Mexico, and Texas and into New Mexico in just 10 years. In such cases, it turns out that these organisms are more highly adapted to the conditions here than are our native species. Consequently, the exotic, introduced species outcompete the indigenous species. Carp have crowded out many types of American fish, and kudzu covers thousands of square kilometers of forests, killing the trees by shading them and cutting off their sunlight (FIGURE B25-1).
No insecticides or herbicides are specific enough to control only the introduced species, and trying to eliminate plants, insects, and fish by hand is almost impossible. Most control efforts center on searching the home habitat to find predators or disease organisms and then introducing those into the new location, but because the original problem was caused by the introduction of an exotic organism, there is, of course, considerable reluctance to bring in another exotic organism. An ideal solution would be to find a pathogen that preys only on the problem species and on no other. Ideally, then, when the original exotic species is eliminated, the second will also die out for lack of food. Caution must be used because the pathogen may be able to attack native species that are related to the exotic species, or a mutation may occur in the pathogen that allows it to attack native species as well.
Already, the transport of species to new habitats where they can proliferate and outcompete native species has been a serious problem. It can only become worse as our jet-age global movement brings about increased travel and transport of material.

FIGURE B25-1 Kudzu (Pueraria lobata), which was introduced from Japan, has no pathogens or pests here in the United States. It grows rapidly, covering rocks, trees, telephone poles, and even buildings. Trees covered by kudzu die from a lack of sunlight.

FIGURE 25-18 Buffalo and plants are in a predator—prey relationship in which the buffalo benefits but the plants do not. The buffalo are herbivores, more specifically they are grazers, and they not only eat the plants but also trample them with their hooves and destroy them when they wallow to rid themselves of fleas and ticks.
Examples of plant—fungus mutualism are well known and recently have been shown to be of much greater importance than ever before suspected, as in the case of mycorrhizae. The mycorrhizal fungus transports phosphate into the plant and receives carbohydrate; both benefit. Many plant species grow only poorly in nature if soil fungi are killed with a fungicide, and it has been postulated that one reason the plains of the American midwest lack trees is because the type of mycorrhizae beneficial to trees cannot compete well with the mycorrhizae beneficial to grasses. A complex relationship has been discovered with Indian pipe (an achlorophyllous parasitic plant) and a mycorrhizal fungus: Indian pipe parasitizes the fungus, drawing nutrients from it, and the fungus in turn obtains carbohydrates from its other mycorrhizal partners that are chlorophyllous and photosynthetic. Indian pipe basically parasitizes other plants, using the mycorrhizal fungus as a bridge.
The nonplant organisms add a great deal of complexity to a plant’s habitat, and numerous types of interrelationships are possible. Only those relationships involving plants have been mentioned here, but the animals are competing with each other, as are the fungi and prokaryotes. Also, interactions occur between animals, fungi, protists, and prokaryotes. The ecosystem is extremely complex, and it is virtually impossible to predict how disruption of one part might affect other parts. Although the operational habitat may be simple, the real habitat contains so many factors linked to so many other factors that we must be careful in our treatment of ecosystems.
![]() The Structure of Populations
The Structure of Populations
Populations can be thought of as having many types of structure; their distribution through the habitat is an important one, as is the age structure of the individuals.
Geographic Distribution
Boundaries of the Geographic Range
The ability of a plant species to spread throughout a geographic area is a result of its adaptations to the abiotic and biotic components of that area. Although most habitat components act on the plant simultaneously and most should be considered important, at any given time and locality, one factor alone determines the health of the plant. This factor, whatever it may be, is the limiting factor. As described for photosynthesis, at a medium level of carbon dioxide, increasing the amount of light causes an increase in photosynthesis, whereas increasing the concentration of carbon dioxide does not; light is the limiting factor. As light intensity is increased, however, a point is reached at which brighter light does not cause more rapid photosynthesis. Then, an increase in the level of carbon dioxide does result in greater photosynthesis, and carbon dioxide becomes the limiting factor.
The concept of limiting factors applies to all aspects of a plant’s interaction with its habitat. In areas of high rainfall, more water probably does not result in better plant growth, but plants growing in the shady part of that ecosystem might benefit from more light, whereas those growing in sunny spots might grow faster if more nitrogen compounds were available in the soil. Still other plants might not respond to extra light or nitrogen but would benefit from decreased herbivory. If each plant received extra amounts of the factor that had been limiting it, its growth would increase until some other factor became limiting. If the shaded plant received extra light, its growth rate would increase to the point where nitrogen perhaps became limiting. If a plant is placed in an optimal environment and given adequate amounts of nutrients, light, water, and freedom from pathogens, growth and reproduction increase greatly, but not infinitely; at some point, the plant’s innate capacity becomes the limiting factor. Crops on irrigated farms with weed control and pesticides are an example (FIGURE 25-19).
Any factor of the ecosystem can act as a limiting factor. Water is important to many species; most cannot live in desert regions because of lack of water, and most cannot live in marshes because of excess water. Extreme temperature inhibits plant growth in many regions; even if given adequate water, some plants cannot conduct it as rapidly as it would be transpired at high temperatures. For other species, high temperatures apparently cause enzyme systems to lose synchronization, and metabolism does not function correctly. A lack of warmth in winter is a limiting factor that keeps many species restricted to the tropics; temperate rain forests have adequate moisture and even richer soil than do the natural habitats of most tropical species, but the temperate rain forest habitat has freezing winter temperatures.

FIGURE 25-19 On modern American farms, plants are given optimal amounts of fertilizer and water; pesticides and insecticides keep pathogens under control, and a variety of weed-control measures eliminate competition from other species. If the seeds are not planted too close together, each plant has adequate room and grows as rapidly as possible. It is limited only by its own innate capacity for growth.
Biotic factors are also critical; many desert plants grow much more rapidly if given more water than occurs in their habitat, but their ranges do not extend into moist regions: They cannot compete well against the plants that are already there and well-adapted to moist conditions. Another important consideration is that plant species that rely on animals for pollination or seed dispersal cannot reproduce where their animal partners do not exist; the geographic range of these plants may be set by the limiting factors of the animals.
Soil factors often produce abrupt boundaries for the geographic ranges of populations. Both mineral composition and soil texture are important (FIGURE 25-20). Soils derived from limestone, sandstone, or serpentine often have characteristic species growing on them. Beaches with loose, porous, sandy soils have species distinct from those on nearby soils that are more compact and contain more humus. The limiting factor for a particular species may be the same factor over its entire geographic range, but often it varies from area to area.
Local Geographic Distribution
In addition to large-scale geographic distribution of the entire population, small-scale, local distribution of individuals with respect to each other is also important. Individuals have one of three types of local distribution: random, clumped, or uniform. The term random distribution is used whenever there is no obvious, identifiable pattern to the position of individuals (FIGURE 25-21A). A random pattern has no predictive value; knowing the position of one plant does not let you estimate the position of another plant. In most habitats, many individuals seem to be distributed at random, but that may simply be due to the presence of many small-scale patterns that are difficult to detect or one large pattern that is too complex to see.
Clumped distributions are those in which the spacing between plants is either small or large, but rarely average (FIGURE 25-21B). This can result from many factors. Seeds of a plant often fall near the plant, not at uniform or random distances from it. If a bird or other animal eats many fruits and seeds, it will probably “deposit” them all together in a lump of organic fertilizer.

FIGURE 25-20 (A) The distribution of these plants is easy to understand. Most of this granite outcrop is so smooth that all seeds are washed off by rain—nothing can grow on it. Where it has cracked, however, soil accumulates, seeds germinate, and plants thrive. (B) Soil in the lowest part of this valley floor is too wet for the trees, and rocky soil on the slopes is too dry.
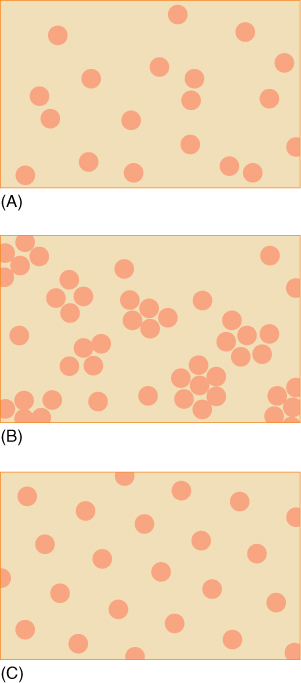
FIGURE 25-21 Types of distribution on a small scale. (A) This appears to be a random distribution, but examination of a larger area might have revealed a pattern. (B) Clumped distribution. (C) Uniform distribution.
Uniform distributions are the types that occur in orchards and tree plantations; all individuals are evenly spaced from their neighbors (FIGURE 25-21C). In natural populations, uniform distributions are not extremely common; those that do occur are thought to result from intraspecies competition. The roots of one individual may establish a zone that prevents the germination or growth of others. Zones can also be established, at least theoretically, by the release from the plant of chemicals that inhibit other plants. Such chemicals are called allelochemics and the inhibition is allelopathy. One example may be the purple salvias of California (Salvia leucophylla); they grow in a relatively uniform spacing with a zone of bare soil surrounding each shrub (FIGURE 25-22). Several chemicals, particularly terpenes, are given off from these plants and have been shown to inhibit growth of plants in the laboratory; it is known also that these do accumulate in the soil near Salvia. It is not certain that these actually are allelochemics, however, because some experiments have shown that rabbits, mice, and birds that live in the salvia shrubs are mostly responsible for the bare patches; if a wire cage is placed over a bare patch to exclude animals, plants do grow there. Presumably the bare patches are narrow because it is risky for the animals to venture too far from the cover of the salvia. It may be possible that terpenes are allelochemics under certain circumstances but that typically animal herbivory prevents other plants from growing near salvia.

FIGURE 25-22 Each bush of Salvia leucophylla is surrounded by a zone devoid of vegetation. Allelochemics given off by the salvias penetrate the top layers of soil and inhibit seed germination. If a seedling can get its root through the top zone, the plant may grow well, but herbivory by rabbits and mice that live in the protection of the salvias then becomes a problem.
Age Distribution: Demography
The manner in which a population responds to various factors in its habitat is affected partly by its age distribution, its demography—the relative proportions of young, middle-aged, and old individuals. Analysis of age distribution has been applied mostly to animal populations and may be difficult to apply to plants, but the fundamental aspects are important and easy to understand. Imagine a species in which a pair of individuals produces four offspring by the time they die; the four offspring also double their numbers, and thus, there are eight after they die. Future generations would contain 16, 32, 64, 128 individuals, and so on. This population is undergoing an exponential rate of increase (FIGURE 25-23A), and there are always greater numbers of young individuals than old ones. This is important in determining whether most members are very young and highly susceptible, moderately young and vigorous, older and well established, or very old and senescent.
Two factors affect the possible rate of population growth: generation time and intrinsic rate of natural increase. Generation time, the length of time from the birth of one individual until the birth of its first offspring, affects the rapidity of population growth: Annuals have a generation time of 1 year or less and can increase rapidly, whereas most conifers and angiosperm trees must be several years old before they produce their first seeds. For easier comparisons, we often measure population increase in terms of generations, not years.
The second factor, intrinsic rate of natural increase or biotic potential, is the number of offspring produced by an individual that actually live long enough to reproduce under ideal conditions. Even with optimal conditions, a large percentage of seeds do not germinate, and many seedlings die before they are old enough to reproduce; therefore, the biotic potential does not equal the number of seeds produced. For many species, biotic potential is a large number, represented in population equations as r. Plants that produce a large number of healthy, viable seeds over their lifetimes have a large r, a large biotic potential, and their populations can potentially increase greatly each generation. A species that produces fewer seeds than another species can reproduce faster than the second if it has a very short generation time, however. Mustard plants are small, live for only 1 year, and produce just a few seeds each, whereas oaks are large trees, each of which produces thousands of seeds in its lifetime. A mustard population, however, can grow more rapidly than can an oak population.
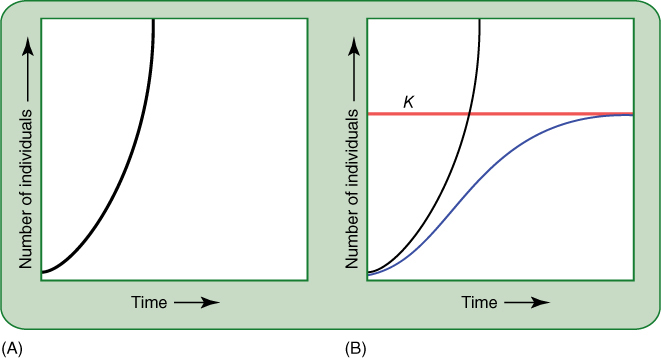
FIGURE 25-23 (A) Curve representing the number of individuals in a population that is growing exponentially. For this to occur indefinitely, growth must be controlled only by the organism’s innate capacity for growth and the number of individuals present. In real situations, habitat limitations cause growth to be somewhat slower than the theoretical maximum. (B) Graph showing the more realistic situation in which the carrying capacity, K, is included. Instead of increasing infinitely (black curve), the growth rate (purple curve) decreases as the population size approaches the carrying capacity of the habitat. When population size equals carrying capacity, population increase stops; the death rate equals the birth (germination) rate.
The biotic potential is measured under ideal conditions, but such conditions do not often occur in nature. Furthermore, after they do occur, even in a laboratory experiment, the very existence of the plants finally disrupts those ideal conditions. After the population becomes large, the plants must compete for water, nutrients, and space. The number of individuals in each population that can live in a particular ecosystem is limited; that number is the carrying capacity and it is symbolized by K. Theoretically, a population increases until the number of its individuals (N) becomes close to K; at that time, crowding and competition result in poorer growth, lower reproduction, and decreased chances that seedlings will be in suitable sites (FIGURE 25-23B). Birth rate (germination) decreases and death rate increases. These factors continue as the number of individuals continues to approach K; when N and K are equal, population growth stops because death rate equals birth rate.
Many factors cause death rate to increase and birth rate to decrease as population size approaches carrying capacity (N approaches K). A large, densely crowded population is an ideal target for herbivores; most of the progeny of one pair of insects are likely to find suitable plants wherever they go in a dense plant population, so most survive. The large plant population is ideal for the insects; therefore, the insects’ population growth rate can increase toward their own biotic potential. The same is true of pathogenic fungi and bacteria. Even without considering predators or pathogens, large populations of plants may alter the environment physically, making it less ideal, as mentioned for the shading of pine seedlings by mature pine trees. A habitat filled with herbivores or pathogens or shade has a lower carrying capacity than the original habitat had.
On the basis of theory alone, we would expect that after a species invades a new habitat, it would undergo exponential growth as in Figure 25-23B, first increasing rapidly, later more slowly, and finally remaining stable with numbers that neither increase nor decrease. In reality, many factors prevent real populations from acting like ideal ones. As the population approaches K, it may not slow sufficiently and may overshoot the carrying capacity; there would be too many individuals temporarily, followed by a die-off that drops the number far below the carrying capacity (FIGURE 25-24). With other species, as N approaches K, an explosive increase in the populations of many types of pests may occur, and the numbers of aphids, mites, caterpillars, and fungi may increase. The plant’s pests have overshot their own K, and they kill so many plants that the plant population falls far below its K, perhaps almost to zero. From there, it may then increase again, undergoing major cycles. Alternatively, as with pines, as the population increases, they change the environment to one that favors other species; their own reproduction falls to zero, and as the adults die, the entire population is lost.
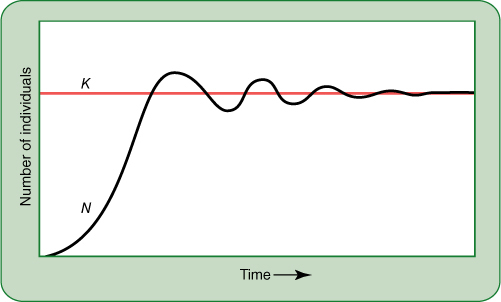
FIGURE 25-24 If a population increases slowly toward the carrying capacity of its environment, it may slow at K, as shown in Figure 25-23B. But often growth is too rapid and a large number of extra individuals temporarily survive; they then die off because of limited resources and the population falls below the carrying capacity.
r- and K-Selection
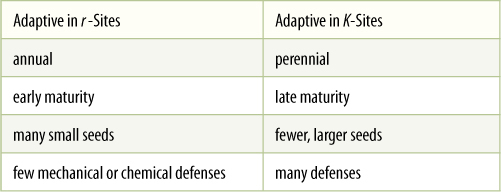
As a population increases, theoretically it goes through a young phase in which numbers of individuals are low and resources are plentiful. Population growth is limited by the species’ own biotic potential, r. Later, conditions are crowded, resources are scarcer, and population growth is governed by the carrying capacity of the ecosystem, K. The lifetime of any single individual is typically much shorter than the time required for a population to pass through this full development. Therefore, which is more advantageous to the species, to become adapted to r conditions or to K conditions? The two are very different and require distinct, often mutually exclusive adaptations.
r-Selection
A disturbance usually produces r conditions. A fire or flood destroys many individuals in the area, and resources are plentiful for the few that remain, whether they are seeds, survivors, or immigrants carried in by wind, animals, or the flood itself. Pioneers that produce many seeds quickly have an advantage in that most seeds find suitable sites; if seeds form too slowly, the sites may be filled by seeds of other invaders (FIGURE 25-25). Because population density is low, the spread of predators and pathogens is slow, and thus, the threat from them is not great; having antiherbivore and antifungal defenses is not so important because neither of those two agents is the limiting factor. Actually, the biotic potential is the limiting factor, and as a result, mutations that increase r are selectively advantageous (TABLE 25-1). Plants grow quickly, have few defensive compounds or structures, flower quickly, and produce many small seeds. Because most disturbances are impossible to predict and are widely scattered, seeds also must be adapted for widespread dispersal. r-selected species typically are annuals or small shrubby perennials because the disturbed habitat gradually changes back into a crowded one that is no longer suitable for the pioneer r species. As more species of plants, animals, and fungi re-establish themselves in the area, the r-selected species are at a disadvantage; they have few defenses against predators and are too short to compete for sunlight. Only another disturbance can save them at this site; usually their population numbers fall to zero or close to it. The species itself survives because many seeds have emigrated to other sites, at least a few of which are appropriately disturbed areas (FIGURE 25-26).
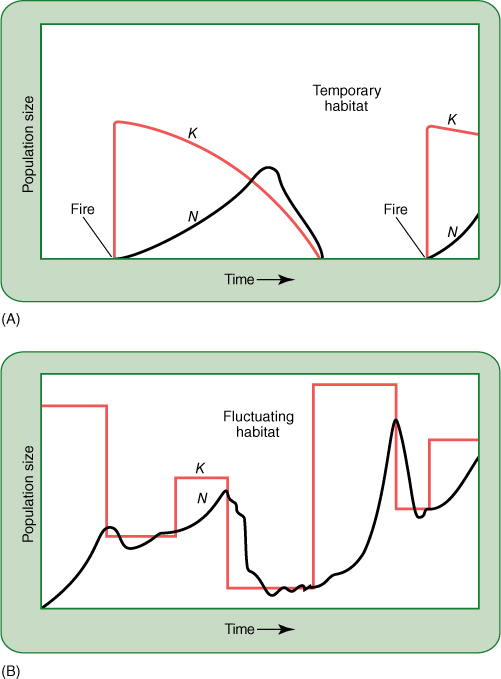
FIGURE 25-25 (A) Most habitats have no carrying capacity (K = 0) for species that are not a natural component of the habitat’s ecosystem, but a disturbance such as a fire, flood, landslide, or disease outbreak may suddenly make it suitable for new species. Clearing an area for construction suddenly increases its carrying capacity for many weeds. They had been present in very low numbers because they compete poorly with the normal vegetation present before construction. The carrying capacity for the weed species soon begins to fall as other plants invade and the natural vegetation returns to areas not covered with asphalt or houses. (B) A site’s carrying capacity may vary less drastically as the populations of competitors, pests, and pathogens rise and fall in their own cycles. If most of the herbivores are suddenly killed by a bacterial disease outbreak, the carrying capacity for the plants suddenly increases. This lasts only until new herbivores move in or some other pest responds to the increased number of uneaten plants.
TABLE 25-1 Selectively Advantageous Traits in r-Selected and K-Selected Species
Some types of disturbances are predictable: killing temperatures in temperate winters and lethal hot/dry conditions in desert summers. For these environments, r-selected species are ideally suited. In spring, the habitat becomes suitable and may be almost devoid of plants. Small annuals grow and reproduce quickly, and population growth is extremely rapid until the habitat becomes disturbed by winter or summer climate.

FIGURE 25-26 Most plants of disturbed habitats, such as these coastal dune plants, are r-selected species: Their wind-blown seeds land, germinate, and grow quickly, and then produce many more seeds. They can grow so rapidly and reproduce so abundantly because almost no energy or mineral resources are spent on antiherbivore defenses or drought adaptations. Because of periodic storms, freshly exposed, highly disturbed sand dunes are always ready for invasion.
Some types of disturbances—especially those caused by us humans—are not predictable. We disturb areas by mining, farming, building cities, damming rivers, and so on. Some of these disturbances eliminate all natural organisms completely, others alter the carrying capacity of habitats. For example, we consume large amounts of fresh water, drawing it from rivers and lakes. Some of the water is returned as treated or untreated waste water, but much of it is used for farming and never returns to the rivers. Consequently, most rivers now do not bring as much fresh water to the oceans as they did formerly. The United States and Mexico draw so much water from the Colorado River and the Rio Grande River that in many years no fresh water reaches the oceans at all: The mouths of the rivers are completely dry. This reduced input of fresh water reduces the carrying capacity of coastal marshes and shore communities (FIGURE 25-27).

FIGURE 25-27 A dam was recently constructed so that water could be used for irrigation. The lack of fresh water entering the ocean here now has changed the habitat and caused many of the mangrove trees to die.
K-Selection
Conditions in a crowded habitat, where a population is close to its carrying capacity, select for phenotypes very different from those that are beneficial in a disturbed habitat (Table 25-1). In a disturbed region, virtually every spot is a suitable site for seed germination and growth, but in a K habitat, almost every possible site is filled. After an individual dies, its site may become occupied by the seeds or rhizomes of a different species. It is advantageous to live for a long time, holding on to a site. To survive as a long-lived perennial is difficult, however, and large amounts of carbon and energy must be diverted into antipredator defenses. These resources are then not available for growth or reproduction, both of which are much slower than in an r-selected species. Many long-lived conifers such as redwoods, Douglas firs, and bristle-cone pines are good examples of K-selected species. K-selected species also face intense competition from other plant species, and therefore, adaptations that increase the ability to use scarce resources are beneficial. Examples of such adaptation may be the capacity to use low amounts of light or soil strata that are poor in nutrients.
Species that are r-selected can occur next to ones that are K-selected. Avalanches in dense forests open up small sites suitable for r-selected species. The same is true for hurricanes, fires, and floods. The floor of a deciduous forest is temporarily an r site during springtime, between the time when temperatures become warm enough for germination and growth and the time when the canopy trees put out their new leaves. For several weeks, the forest floor is sunny, warm, rich in nutrients, and temporarily uncrowded. Small r-selected plants that grow and reproduce quickly can complete their life cycles before the larger trees come out of dormancy and block the sunlight.
![]() The Structure of Ecosystems
The Structure of Ecosystems
Many concepts can be considered in the structure of an ecosystem. Different botanists think about ecosystems in ways that are influenced by their own interests. Four of the most commonly mentioned structures are the physiognomic structure, temporal structure, species diversity, and trophic levels.
Physiognomic Structure
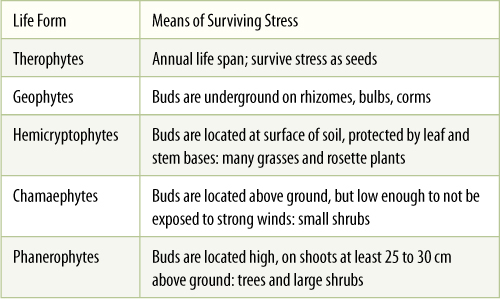
The physical size and shape of the organisms and their distribution in relation to each other and to the physical environment constitute the physiognomic structure. Trees, shrubs, and herbs are the three most useful categories, but in addition, a system of life forms was defined by C. Raunkiaer in 1934 (TABLE 25-2). The criterion for classification was the means by which the plant survives stressful seasons, such as by placing buds below ground (geophytes: bulbs, rhizomes) or winterizing aerial buds (phanerophytes: trees, vines). Regions of the world that have similar climatic conditions have similar physiognomic structures unless the soil is particularly unusual (FIGURE 25-28).
TABLE 25-2 Life Forms of Raunkiaer
An almost infinite number of combinations of life forms, vertical structures, and characteristics such as abundance of broadleaf plants, conifers, and sclerophyllous plants are possible (FIGURE 25-29). An almost infinite number of types of ecosystems might be expected but, actually, only a few basic types exist. Although differences exist between various types of forests or grasslands or marshes, each category is easily recognized as a common type of ecosystem.
Temporal Structure
The changes that an ecosystem undergoes with time constitute its temporal structure; the time span can be as short as a day or can encompass seasons or decades. For animals, a daily cycle can be especially obvious, with some animals active at night (nocturnal) and others during the day (diurnal). Many plants also have daily rhythms of flower opening and closing.
Plants change dramatically with the season, as do other organisms. Spring is typically a time of renewed activity, with production of flowers and new leaves. This is not simultaneous for all species; often the understory plants become active earliest, benefiting from the open canopy. Wind-pollinated flowers are usually produced before leaves expand and block the wind. Leafing out and flowering must be coordinated not only with the end of low-temperature stress conditions but also with the habits of pollinators and herbivores. It does a plant no good to produce flowers when temperatures are still too low for its pollinators. Through late spring and early summer, various species flower at distinct times, controlled by plant maturity, photoperiod, or adequate rainfall. In most ecosystems, there is no time during the summer when nothing is in bloom. The continual presence of some species in flower provides nectar and pollen for insects throughout the season.
Species that bloom early in spring probably form fruits and mature seeds during summer; species that flower later release their seeds in late summer or autumn. After seeds are dispersed, some remain dormant until the following spring. Others germinate and grow into a low rosette that survives the winter, even growing slightly on warm winter days. When spring arrives, the seedling is already well rooted and can begin to grow quickly while the seeds of competitors are just starting to germinate.
Late summer and autumn bring changes that depend on the ecosystem; in the northern United States, herbs die while shrubs and trees develop resting buds. The entire plant enters light dormancy; then the first cool days initiate deep dormancy. Leaves and fruits are abscised, removing the last sources of food for most animals. In the southern and southwestern United States, cooler autumn weather is often more welcome than the first warm days of spring because the summer is so much more severe than the winter. Most gardening is done in autumn rather than spring, and fall wildflowers are abundant and dramatic. The growing season extends at least to December for shrubs and many herbs, and the small rosette plants may never become truly dormant.

FIGURE 25-28 (A) This desert scrub vegetation near Tucson, Arizona, with small trees, large bushes, and saguaro cactus, is characteristic of much of the southwestern United States and northern Mexico, where summers are hot with occasional rainstorms and winters are cool and moist but freezes are not severe. (B) The northern part of the central valley in Chile, just north of Santiago, has climatic conditions similar to those in the southwestern United States, and the vegetation of the two regions resembles each other in habit (life forms), distribution, and other features. Only by looking carefully does one notice that the species are very different.
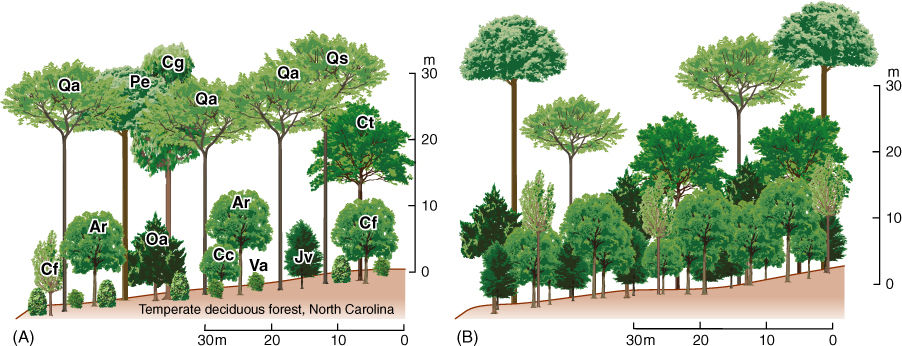
FIGURE 25-29 The presence of trees, shrubs, and herbs (phanerophytes, chamaephytes, and therophytes) gives an ecosystem vertical structure. The trees form the canopy. The shrubs, short trees, and saplings of tall trees form a middle level understory or subcanopy. The herbs and seedlings are the ground-level plants. Vines can occur in all three levels. (A) A temperate forest. Ar, Acer rubrum (red maple); Cc, Cercis canadensis (redbud); Cf, Cornus florida (dogwood); Cg, Carya glabra (pignut hickory); Ct, Carya tomentosa (mockernut hickory); Jv, Juniperus virginiana (red cedar); Oa, Oxydendrum arboreum (sourwood); Pe, Pinus echinata (shortleaf pine); Qa, Quercus alba (white oak); Qs, Quercus stellata (post oak); and Va, Viburnum affine (arrow-wood). (B) A tropical rain forest has a much more complex structure.
In tropical ecosystems, winter and summer do not exist, but an alternation of dry and wet seasons governs ecosystem change. Coastal marsh, wetland, and reef ecosystems may be strongly affected by seasonality; rainy seasons dilute the salt water, whereas in dry seasons rivers deliver less freshwater and mineral-rich silt.
Over long periods—many years—most ecosystems undergo gradual, often dramatic changes. This process of succession is discussed in the next chapter.
Species Composition
Species composition refers to the number and diversity of species that coexist in an ecosystem, and it depends on whether the climate is mild or stressful, the soil is rich or poor, and the species’ tolerance ranges are broad or narrow (FIGURES 25-30 and 25-31). Stressful climates with poor soils support a low number of species because so few species are adapted to such conditions (FIGURE 25-32). On the other hand, mild climates and rich soils support an abundance of species because most plants have tolerance ranges that include such climatic and soil conditions (FIGURE 25-33). Competition is intense, but apparently natural selection has resulted in habitat partitioning, with each species occupying a narrow portion of the various resource gradients (Figure 25-31). The presence of a large number of species actually creates more niches that can be filled by new species; the presence of trees makes it possible for epiphytes and parasites to occur in the ecosystem.
Trophic Levels
Trophic levels are basically feeding levels. Each ecosystem contains some members, autotrophs, that bring energy into the system. Photosynthesis is by far the dominant method, accounting for virtually all energy input. Green vascular plants are most important, but algae and cyanobacteria carry out about one third of all photosynthesis worldwide, and lichens and nonvascular plants are important in cold, high latitudes and altitudes. Chemosynthetic bacteria bring chemical energy into ecosystems, and although this is minor now, it was the only method before photosynthesis evolved.
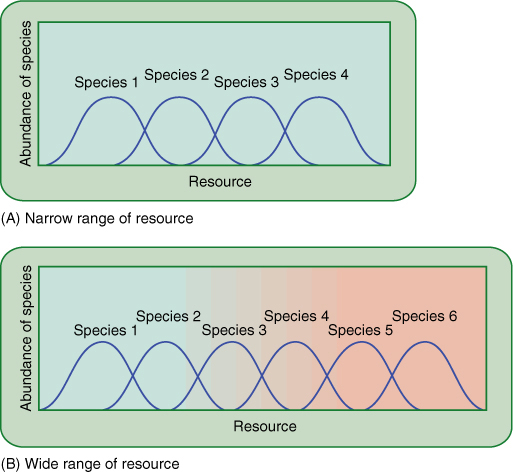
FIGURE 25-30 In the ecosystem represented by (A), the range of the resource available is narrow compared with that of the ecosystem in (B); in general, it is able to support fewer species. For example, the resource may be water, with the ecosystem in (B) having a variety of areas that range from dry to moist to lakes or streams, whereas the ecosystem in (A) is just marshy or just desert.
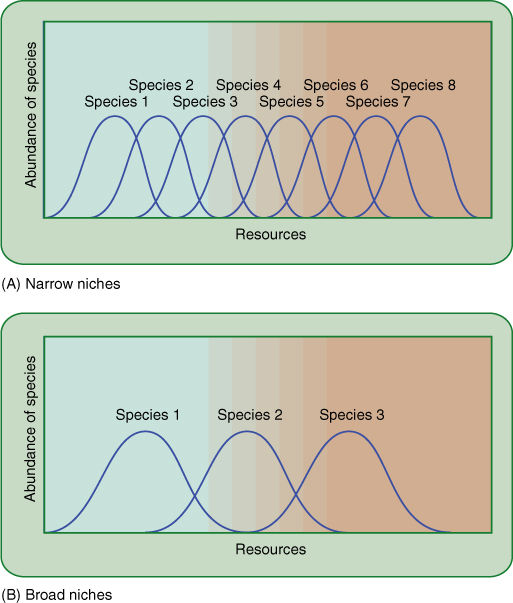
FIGURE 25-31 Two ecosystems with similar ranges of resources can differ in the number of species they contain. One may be occupied by a large number of very specialized species, each adapted to only a narrow range of the resource (A), whereas the other may be occupied by a few species of generalists that grow well under a variety of conditions and exclude most competitors (B).
Autotrophs are known as primary producers for obvious reasons, and they are the first step of any food web. They are the energy and nutrient supply (food) for the herbivores, which constitute the primary consumers (sometimes called secondary producers). Herbivores are preyed on by carnivores, the secondary consumers. Omnivores exist at both trophic levels. Decomposers such as fungi and bacteria break down the remains of all types of organisms, even those of other decomposers (FIGURE 25-34).
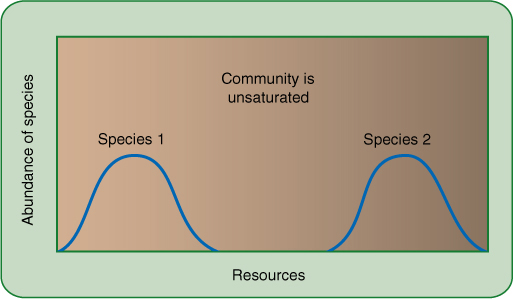
FIGURE 25-32 Adapting to certain harsh conditions is difficult, and only a few species may have succeeded. Consequently, harsh habitats may have many unused resources.

FIGURE 25-33 This climate is nonstressful, the soil is rich, and no toxins are present. Many species live here, and many more could if protected from competition. Species diversity here is high.
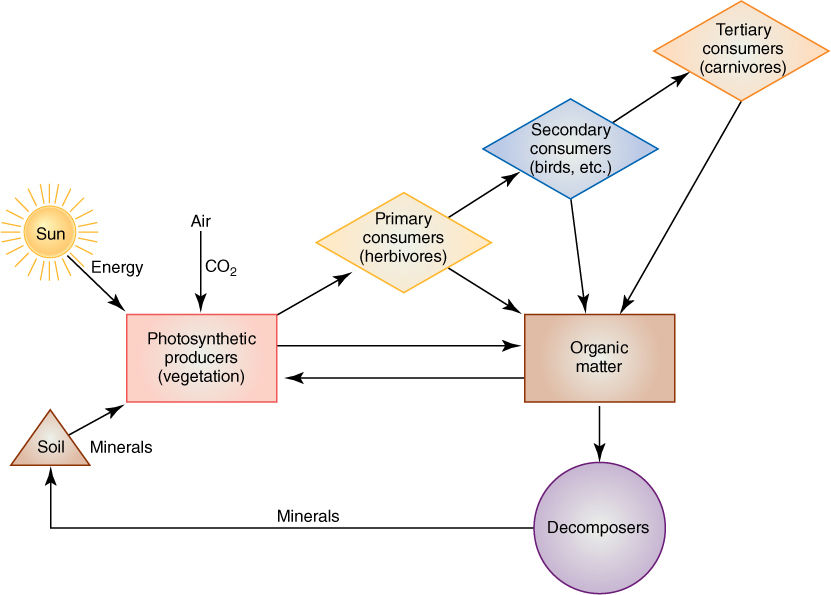
FIGURE 25-34 Energy and materials flow through the ecosystem as a network rather than in a straight line. Each level contributes to the decomposers (they even decompose other decomposers), and omnivores eat both plants and other animals. The primary producers never draw energy from the other stages, but many depend on animals and decomposers to return essential elements to the soil. At each step, respiration returns most of the carbon back to the air, and most of the energy is liberated as heat.
As plants photosynthesize, energy and carbon compounds enter the ecosystem. As plants are eaten, their energy and carbon compounds move to the herbivore trophic level, then to the carnivore trophic level, and finally on to the decomposers. This is referred to as the energy flow and the carbon flow of the ecosystem. At each step, much of the food is used in respiration, resulting in the production of ATP, heat, carbon dioxide, and water. The carbon dioxide is released back to the atmosphere, where it can be used in photosynthesis again; the energy temporarily contributes to warming the planet, but it is ultimately radiated into space. The portion of the food that is not respired is available for growth and reproduction. As a very rough approximation, approximately 90% of an animal’s food is respired, and 10% is retained as growth or gametes; therefore, when herbivores eat 1,000 kg of grass, 900 kg are respired, and 100 kg become herbivore tissues. Similarly, a carnivore that eats the 100 kg of herbivore would only gain 10 kg and after dying would support just 1 kg of decomposers.
Along with the flow of energy and carbon, minerals flow through the ecosystem (FIGURE 25-35). Plants absorb minerals from soil and incorporate them into their bodies as amino acids, ATP, coenzymes, and so on, and these are consumed by herbivores. Many of these compounds are broken down by animals, and the minerals are lost as waste. After organisms die, the minerals that remain in their bodies are released by decomposers and become part of the soil again. In addition, minerals can be completely lost from an ecosystem as they are carried away by rainwater and streams, especially if erosion is occurring or as cities dump mineral-rich human waste into rivers. Such minerals are carried to the oceans and become part of marine ecosystems. Many algae, protists, and marine animals have bodies with heavy, mineral-rich shells and bones; as those organisms die, their bodies sink to the depths of the ocean and are unavailable to living organisms of the land or ocean surface.
The movement of energy and biomass from one trophic level to the next is often represented as a pyramid of energy or a pyramid of biomass (FIGURE 25-36). Such pyramids are useful for illustrating these principles in textbooks, but in reality, it is extremely difficult to measure all of the biomass of the primary producers of even a simple ecosystem. It is almost impossible to measure it accurately in a forest ecosystem, and it is impossible to measure the biomass or energy content of the fungi and bacteria that are acting as decomposers. Pyramids of numbers are more easily obtained; often there are fewer individuals in each higher trophic level, but of course, millions of aphids can live on one tree.

FIGURE 25-35 (A) The movement of minerals is easy to understand with sea birds and small oceanic islands. Many small oceanic islands are composed of rock that breaks down into a soil that is poor in nitrates and phosphates. But sea birds eat fish, then roost on the islands where they defecate, depositing mineral-rich guano. This fertilizer not only allows plants to flourish on the islands, but the minerals are eventually washed back into the ocean, enriching the growth of algae and fish, completing the cycle. (B) The ground on this island is white because of the thick layer of guano. During rainy seasons, many species of plants grow among the cacti.
Pyramids of biomass and numbers have become important as a means of illustrating the impact of the human introduction of herbicides, pesticides, and toxic wastes. Although some of these are biodegradable, many are not, and they persist unchanged and toxic in the environment. Imagine a toxic herbicide applied at the low rate of one part per million—1 g per 1,000 kg of pasture grass. Herbivores will eat the 1,000 kg of plants, and the pyramid of biomass shows that they will respire 900 kg away; however, the 1 g of herbicide is not oxidized or excreted. Instead, it accumulates in the animals, now at a concentration of 1 gm per 100 kg. This is 10 times stronger than the application rate. As carnivores eat the herbivores, the concentration rises to 1 g per 10 kg, now 100 times stronger. Human metabolism is no different; these chemicals move through the food web to us as well. Toxic materials are not distributed uniformly within the bodies of animals and humans but become concentrated in our livers and kidneys as well as in fatty tissues such as our brain and spinal cord.
The role of decomposers in the ecosystem is vitally important; decomposers are so ubiquitous as well as microscopic that we take them for granted. If there were no decomposers in nature, plants could grow only as rapidly as weathering breaks down rock; as soon as minerals were released, they would be absorbed and locked into a plant and would then move through the food web. When the organisms died, the minerals would remain locked in the nondecomposing body. Sterilized (canned) food does not decay even though moist and kept at room temperature because no decomposers are present. Only by the physical and chemical weathering of rock and dead bodies could plant growth continue, but with decomposers present, minerals are recycled rapidly; decomposers release the minerals to the environment through their membranes, and their own bodies are so small and delicate that after death they are broken down immediately by other decomposers or by inorganic chemical weathering.
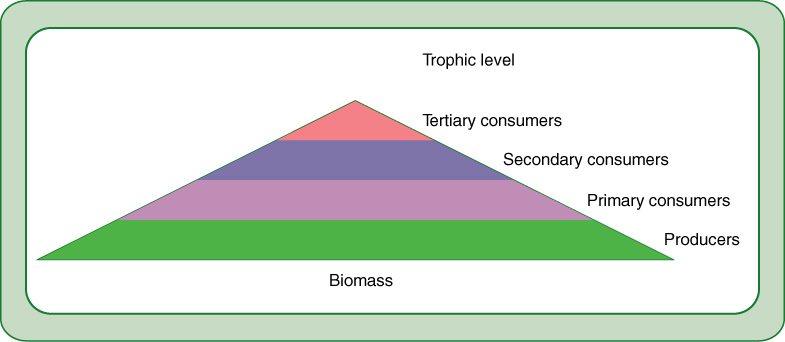
FIGURE 25-36 A pyramid such as this can represent the number of organisms at each level, the weight of all the individuals at each level, the respiration at each level, and so on. Such analysis can be difficult to do, but these relationships are extremely important. If a national park is established to preserve a species of wolf or mountain lion, it must be known how extensive its food web is and how large of an area must be set aside to ensure an adequate number of primary consumers.
 At the Next Level
At the Next Level
1. Generation time, lifespan, and demography. Some plants reproduce while they are only a few months old; others must live for decades before they reproduce. Some plants live for just a few months, others for thousands of years. Before you do any research, think about the ways these two characters might be related. Obviously a plant that lives only a few months cannot also be one that does not reproduce until it is decades old. But can long-lived plants reproduce early or do you think they have to wait? Look up data on generation time and lifespans for a variety of plants and see if you can construct your own mathematical models of what their population structure might be like.
2. A plant’s intrinsic rate of natural increase (biotic potential). A plant’s intrinsic rate of natural increase, symbolized as r in the text, is crucial in many ways. When we collect plants or animals from nature, such as valuable wood like ebony and mahogany, or seafood like fish and shrimp, it is important to avoid overharvesting. The biotic potential of a species gives us some information about its capacity to replace lost individuals. Similarly, as we try to improve crop plants, we need to know which factors limit their growth rate or the rate at which they produce seeds, fruit, or whatever it is we need from them. Biotic potential can be measured accurately in greenhouses and experimental farms, and we can also get a rough estimate from observing plants in nature. Do plants that grow rapidly and reproduce early, such as mustards, have a higher biotic potential than those that grow more slowly and reproduce later?
3. The life forms of Raunkiaer. The life forms in Table 25-2 are based on the ways in which plants survive stressful seasons (buds underground or buds located high, and so on). Many plant ecologists use these life forms as information about how ecosystems function. For example, geophytes such as bulbs and rhizomes are common where fires are frequent whereas hemicryptophytes such as grasses are abundant where animals eat taller plants with exposed buds. You should be able to find considerable information about life forms and ecology.
SUMMARY
1. Ecology is the study of organisms in relationship to all aspects of their surroundings. Each plant is part of a population of individuals of the same species, and these co-exist with other species, forming a community. When climate and physical surroundings are also considered, the whole is an ecosystem.
2. The biology of a population is not simply the sum of all the individuals because plants interact positively and negatively with each other.
3. Habitat components may be biotic or abiotic. Abiotic factors include climate, soil, latitude, altitude, and disturbances. Biotic factors include all living organisms and the results of their activities.
4. Disturbances are disruptions of portions of an ecosystem; although they may cause great destruction, they may also be necessary for maintaining the ecosystem such that there is not a succession, a long-term change in species present.
5. By existing in an ecosystem, a plant affects the ecosystem; the plant may produce changes that are detrimental for the survival of itself or its progeny, or it may alter the ecosystem such that it is more suitable for its progeny.
6. The amount of competition varies from species to species and from one ecosystem to another. In general, where climates are mild and soils rich, species diversity is great, and competition is most important in excluding species from the ecosystem. In harsher conditions, fewer species exist, and the conditions themselves limit the number of species that exist successfully.
7. With a large geographic range, a species may face different types of competition or stress in different parts of its range.
8. Plants interact with animals, fungi, protists, and prokaryotes in numerous ways, but most interactions basically can be divided into mutualistic (beneficial to both) or predatory (detrimental to one, beneficial to the other).
9. Populations have at least two basic types of structure: (1) their geographic range, which is set by limiting factors, and their local distribution (random, clumped, or uniform), and (2) the distribution of the ages of all the individuals (demography).
10. The biotic potential of a species (r) is a measure of its reproductive success: the mean number of offspring per parent that survive to reproductive maturity. An r-selected species is one that is adapted to sites temporarily free of competitors and rich in resources.
11. The carrying capacity (K) of an ecosystem is a measure of the number of individuals of a particular species that can be sustained by the ecosystem. A K-selected species is one that is adapted to stable, relatively unchanging sites where competition may be very important.
IMPORTANT TERMS
biotic potential
carrying capacity
commensal relationships
community
competition
demography
ecosystem
ecotypes
habitat
herbivory
K-selected species
life forms
limiting factor
mutualism
niche
pathogenic
pioneers
population
predation
primary consumers
primary producers
r-selected species
tolerance range
trophic levels
REVIEW QUESTIONS
1. Ecology is the study of organisms in relation to ______________________ aspects of their surroundings.
2. Define each of the following: population, community, habitat, and ecosystem. Why is the biology of a population different from the biology of an individual?
3. How much of a plant’s environment should be considered its habitat? Would the habitat of a wind-pollinated, wind-dispersed species be simpler than one with animal-mediated pollination and seed dispersal? Why?
4. Consider the redwood forests of California. We know that the shade of the big trees affects the shrubs and herbs, but do the small plants affect the trees? What about the seeds of the redwood trees? What role did fire play? If a new disease were to kill off all the shrubs and herbs but not directly harm the trees, would the trees ultimately be impacted by the disease?
5. Habitat components are of two types, abiotic and biotic. List several components of each type.
6. Why are climate extremes often more important than average climatic conditions? What is the “growing season” of a plant, and what are some conditions that affect its length?
7. How are soils formed? Through the breakdown of what material? Why are new soils not suitable for most plant species?
8. What is a pioneer species? Are they more likely to be r- or K-selected? Why?
9. Why do parts of Alaska and Canada have such thin soil today? What happened to the soil that used to be there? Approximately how many years ago did this region lose its soil?
10. Fire is a critically important, natural disturbance in many ecosystems. Describe some plant adaptations that allow plants to survive fires.
11. If we could eliminate all other organisms so that a certain region contained only the soil and the plant itself, would this be a stable habitat? Does the plant itself change its habitat? Explain.
12. When several individuals occur together, the possibility for interaction is created. If the interaction is basically beneficial for both organisms, it is called _________________, but if it is disadvantageous, it is ________________. Do the two organisms have to be the same species or different species, or does it matter?
13. What is competition between plants? What is competitive exclusion? How does competition affect natural selection and evolution?
14. What is a niche? Is it a physical, tangible space or object? If a giant redwood grows side by side with a small fern, are they in the same niche? If not, how does the niche of one differ from that of the other?
15. What is a transplant experiment? What is a common garden? What is the point of using these two types of experiment?
16. When birds build nests in trees, the birds benefit, but the trees are usually unharmed. What kind of a relationship is this? Give two more examples of relationships like this.
17. Although most habitat components act on a plant simultaneously, at any given time and locality, one factor alone determines the health of the plant. This is called the_____________________factor.
18. Plants often grow in clumped distributions. Give one cause of this.
19. What is the term that describes the length of time from the birth of one individual until the birth of its first offspring? Why do we not use this term for the time between the individual’s birth and its own death?
20. Describe biotic potential. Describe carrying capacity. How do these two factors affect population growth and the ultimate stable size of a population?
21. Consider the population growth curve of Figure 25-23B. Would a young population (left side of curve) have a greater proportion of young individuals than an old population (right side of curve where it has reached K)? Why? Does it matter whether a population has a greater proportion of young, middle-aged, or old individuals? The answer is obvious for humans, but does it matter for plants?
22. Autotrophs are known as _______________________, and they are the first step of any food web. Herbivores are _________________________________, and carnivores are __________________. Fungi and prokaryotes break down the remains of all types of organisms and are known as______________________________.
23. As plants photosynthesize, _______________________and _________________________ compounds enter the ecosystem. How do these move to the next trophic level and then to the next? How are both lost from the ecosystem?
24. Look at the pyramid of biomass in Figure 25-36 and the text on the same page. If a toxic herbicide is applied at a low rate of one part per million, what happens to the concentration of the herbicide at each higher level of the pyramid? Does this affect our food? Does it affect us?
Design Credits: Hummingbird: © Tongho58/Moment/Getty; Green Plant Cells: © ShutterStock, Inc./Nataliya Hora; Purple Tulip: © ShutterStock, Inc./Marie C Fields;
Dandelion: © ShutterStock, Inc./Danielkreissl; Poppy: © ShutterStock, Inc./Saruri; Plant Icon: © ShutterStock, Inc./Vector; Digging Man Icon: © ShutterStock, Inc./Z-Art