Botany: An Introduction to Plant Biology - Mauseth, James D. 2017
Appendix
Notes on Fundamental Aspects of Chemistry
OUTLINE
✵ Concepts
✵ Atoms and Molecules
✵ Water
- Polarity
- Solubility
- Acids and Bases
✵ Chemical Reactions
✵ Carbon Compounds
✵ Organic Molecules
- Functional Groups
- Carbohydrates
- Proteins
- Nucleic Acids
- Lipids
✵ Cofactors and Carriers
- Energy Carriers
- Electron Carriers
✵ Enzymes
![]() Concepts
Concepts
Plant metabolism, like that of all other organisms, is based on the fundamental principles of physics and chemistry that govern inanimate matter. The bodies of plants and other organisms are made of atoms drawn from soil, air, or water, and the energy that drives their metabolism is produced by ordinary chemical reactions.
![]() Atoms and Molecules
Atoms and Molecules
There are 92 natural elements, each differing from the others by the number of protons in the nuclei of its atoms. Neutrons affect only the weight of the atom, not its chemical properties. Around each nucleus are electrons with a negative charge, and thus each neutralizes the positive charge of a proton. Atoms with equal numbers of protons and electrons are electrically neutral.
An atom or molecule that carries a charge is an ion; a negative ion is an anion (pronounced AN eye on); a positive ion is a cation (pronounced CAT eye on).
![]() Water
Water
Polarity
Electrons are often not shared equally between the atoms of a molecule. In water, electrons spend more time near the oxygen atom and less time near the hydrogens; therefore, the oxygen in water has a partial negative charge and each hydrogen has a partial positive charge. The molecule is thus a polar molecule, with a slightly negative end and a slightly positive end. When two water molecules come close to each other, the positive charge of one slightly attracts the negative end of the other. This is hydrogen bonding, and it is sufficient to cause water to be a rather sticky, viscous substance. It absorbs a great deal of energy without warming rapidly and requires a large amount of energy to convert to vapor. Without the stickiness caused by hydrogen bonding, water molecules could not be lifted from roots to leaves.
Substances that carry no unbalanced electrical charge are nonpolar substances, and they do not undergo hydrogen bonding. Nonpolar substances move easily past each other and flow with little viscosity. When energy is supplied, their speed quickly increases, raising their temperature. They boil and turn to a gas even at a low temperature, with little energy needed.
Solubility
Molecules that are polar or that form hydrogen bonds are water soluble: They will dissolve in water. Nonpolar substances and those that cannot form hydrogen bonds, are water insoluble, but they will dissolve in fats and oils: They are lipid soluble.
Acids and Bases
The concentration of protons (H+) in a solution is known as the acidity of a solution; it is measured as pH, which is the negative logarithm of the H+ concentration (TABLE AP-1). Any substance that increases the concentration of free protons is an acid. A base is anything that decreases the concentration of free protons; this is usually accomplished by giving off hydroxyl ions that combine with protons and form water, effectively removing free protons.
TABLE AP-1 Acids and Bases

![]() Chemical Reactions
Chemical Reactions
Atoms and molecules become more stable by the transfer or sharing of valence electrons among atoms. It is important to emphasize that by more stable we mean that the atoms have less energy. A reaction that releases energy to the environment (such as respiration) is an exergonic reaction; if energy is released as heat, the reaction is also exothermic. A reaction that absorbs energy from the environment (such as photosynthesis) is an endergonic reaction (endothermic if energy is absorbed in the form of heat). Every endergonic reaction needs a source of energy—an exergonic reaction occurring somewhere. For example, the endergonic reactions of photosynthesis are powered by energy given off by the exergonic thermonuclear reactions in the sun—even though it is 93 million miles away.
In a second-order reaction, two molecules react to form a third: A + B → AB. In order for A and B to react, they must be so close that their valence electrons can move between the two sets of orbitals, but as A approaches B, the two sets of electrons repel each other. The energy (speed) needed to overcome electron-cloud repulsion and permit chemical reaction is called the activation energy. FIGURE AP-1 shows a reaction diagram of the potential energy of the reactants and products during a reaction.
A first-order reaction involves only one molecule, not the collision of two. An example is AB → A + B, in which a compound breaks down into two parts. First-order reactions also have energies of activation that are overcome as the two parts of the molecule vibrate. They can be accelerated by heating or by catalysts.
For every reaction, an opposite back reaction can also occur; thus, not only can A and B combine to form AB, but AB can break down to form A and B. In any mixture of A and B, both reactions occur. In all reactions, a reaction equilibrium is established in which there is a mixture of some reactants and some products, some forward reaction and some back reaction.
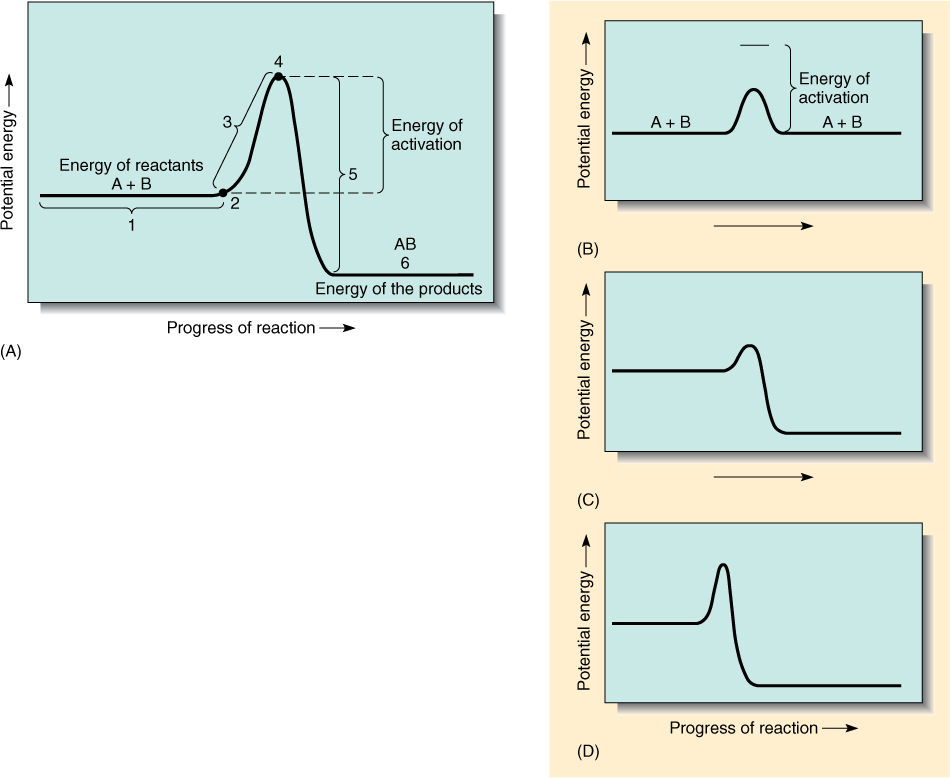
FIGURE AP-1 (A) A potential energy diagram of a second-order reaction. At 1, two atoms, A and B, are still too far apart to influence each other, but at 2, they begin to repel. During interval 3, they continue to approach because their momentum is great enough to overcome repulsion. At 4, orbitals rearrange from nonbonding to bonding orbitals, and energy is given up (5) and radiated away from the molecule. Consequently, after the reaction (6), the molecule AB has less potential energy than the two atoms had at 1. (B) If two atoms are moving too slowly, electron cloud repulsion is not overcome, and the two atoms bounce off each other. No reaction occurs. (C) A low energy-of-activation barrier. (D) A high energy-of-activation barrier.
Although there is no way to change the energy of activation of a reaction, it is possible to change the mechanism of a reaction. A substance that enters a reaction and permits it to occur rapidly at low temperature is a catalyst; in living organisms, virtually all catalysts are proteins called enzymes.
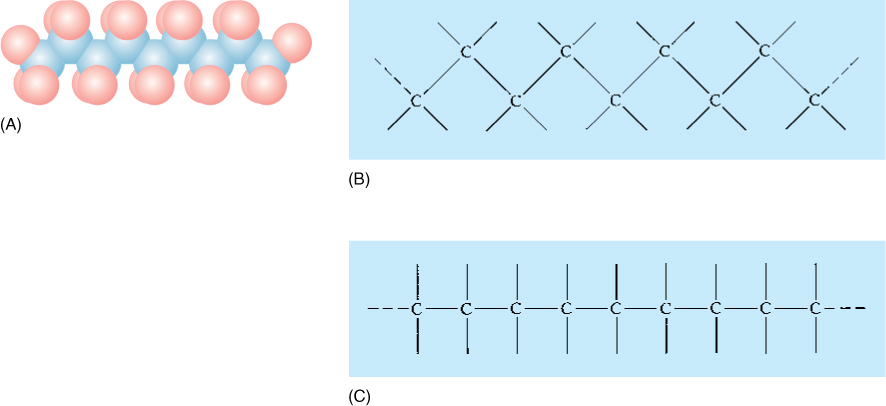
FIGURE AP-2 (A) A series of carbon atoms bonded to other carbons by single bonds would look something like this, with the blue spheres representing carbon atoms and the red spheres representing other atoms attached to the carbons. Because the bonds are arranged as a tetrahedron, the carbon backbone has a zigzag shape. (B) and (C) are simpler ways of showing the structure in (A).
![]() Carbon Compounds
Carbon Compounds
Carbon forms three types of covalent bonds. If there are four other atoms, the carbon is linked by single bonds, which are arranged in a tetrahedron. In a chain of carbons, the carbon backbone is zigzag, not straight (FIGURE AP-2).
Carbon can form a double bond by sharing two of its electrons with one other atom that also contributes two electrons. If a carbon forms one double bond and two single bonds, the molecule is flat and shaped like a Y. The double bond is extremely rigid, and the arms cannot rotate around the carbon. Many organic molecules have carbon—carbon double bonds, and because the double bond cannot rotate, there are two possible forms; with both side groups on the same side of the double bond, they are in the cis position. When on opposite sides, they are in the trans position. The cis form has physical and metabolic properties that differ from those of the trans form.
Rarely, carbon forms a triple bond, but it is not very stable (its electrons still have a great deal of energy) and is broken easily.
![]() Organic Molecules
Organic Molecules
Functional Groups
The properties of a compound are due mostly to the chemical groups, known as functional groups, attached to the carbon atoms (TABLE AP-2). The physical and chemical properties of organic molecules are most strongly determined by the functional groups present; the backbone often is not so important. Because carbon compounds can be large, each may have many functional groups of various types, being simultaneously both acidic and basic, or lipid soluble in some regions and water soluble in others.
Carbohydrates
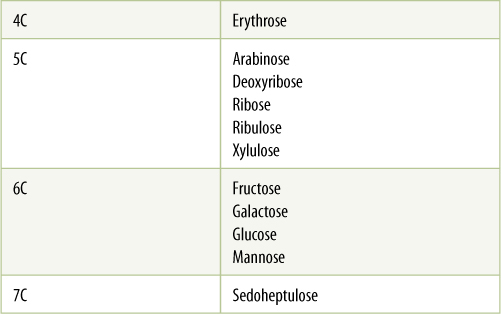
The simplest carbohydrates are the monosaccharides, or simple sugars, glucose being a familiar example. Monosaccharides are classified by the number of carbon atoms each contains; pentoses (5 carbons) and hexoses (six carbons) are the most abundant (TABLE AP-3). Monosaccharides are flexible because all their carbon—carbon bonds are single bonds. When one end of a molecule comes close enough to the other end, the two may react, forming a closed ring (FIGURE AP-3). The ring form is the more stable, more common form for a hexose dissolved in the water of a cell.
Monosaccharides can act as monomers, reacting with other monosaccharides to form polymers called polysaccharides. Sucrose is a disaccharide composed of one glucose molecule plus one fructose molecule. Extremely short polysaccharides, less then about 10 monosaccharides long, are called oligosaccharides. Many organisms attach short oligosaccharides onto proteins.
TABLE AP-2 Functional Groups
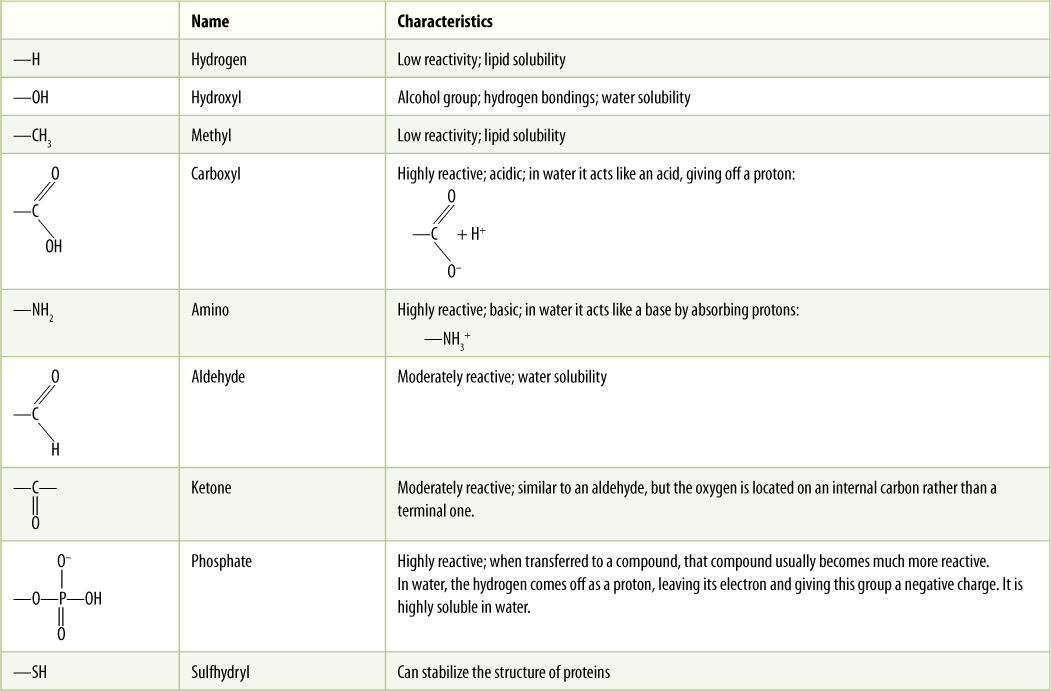
TABLE AP-3 Common Monosaccharides
Starches, technically known as amylose and amylopectin, are long polysaccharides composed only of glucose residues (FIGURE AP-4). The enzyme responsible for polymerization is starch synthetase and it makes an alpha-1, 4-glycosidic bond. Starch molecules become about 1,000 glucose residues long, but great variation exists.
A set of enzymes called cellulose synthases polymerizes glucose molecules into the polymer cellulose. The glucose residues have an alternating orientation, and the resulting bond is a beta-1, 4-glycosidic bond. Cellulose molecules form large numbers of hydrogen bonds with other cellulose molecules, crystallizing into rigid aggregates that are extremely strong.
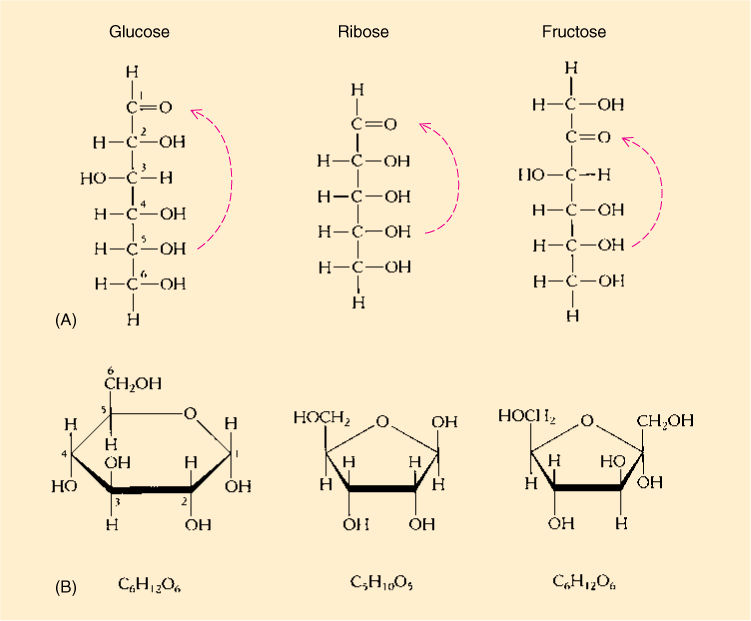
FIGURE AP-3 (A) Ribose is a 5C sugar, a pentose; glucose and fructose are both hexoses (6C). (B) Each sugar can also exist in ring form, but ring formation involves the aldehyde or ketone functional group, which are therefore not present in the ring-form sugars.
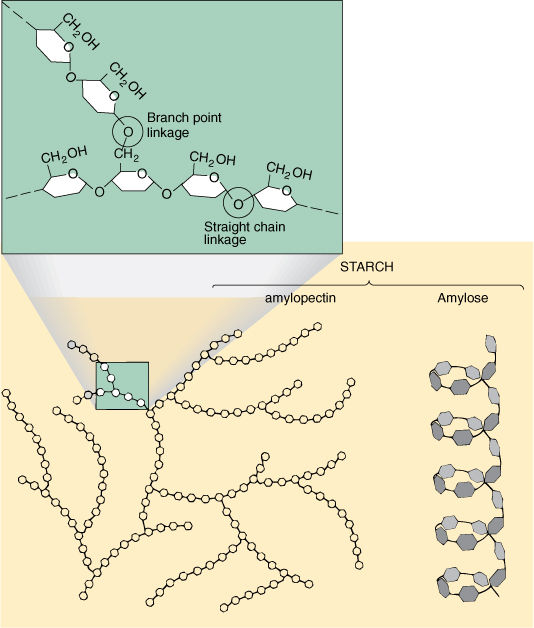
FIGURE AP-4 If glucoses are linked to other glucoses by alpha-1, 4-glycosidic bonds, the result is an unbranched, coiled chain called amylose, the main component of starch. A second enzyme occasionally makes a bond with the number 6 carbon, resulting in an alpha-1, 6-branched-chain amylopectin, also a component of starch.
Proteins
Proteins are unbranched polymers of amino acids; they tend to be about 100 to 200 amino acid residues long. Very short proteins, with fewer than about 50 amino acids, are frequently called polypeptides. Twenty amino acids are used for protein synthesis (FIGURE AP-5); each consists of one carbon that carries four functional groups: (1) —COOH, the carboxyl group that causes it to be an acid; (2) —NH2, the amino group; (3) —H; and (4) a fourth group “R” that differs from one amino acid to another. The R groups are not involved in polymerization; instead, they protrude to the sides of the protein backbone, and their properties determine the property of the protein.
The amino acid sequence of a protein is its primary structure. Because each amino acid has a unique R group, the sequence of amino acids produces a particular sequence of R groups. Because R groups are chemically diverse, a protein has extremely complex properties. Proteins are flexible, and some regions interact with other regions of the same protein, causing the entire molecule to have a characteristic shape.
If the R groups (functional groups) of an entire region of a protein are of the correct type, that region takes on a secondary structure such as an alpha helix or a beta pleated sheet. Both of these secondary structures are short and affect just portions of a single protein; it is common for certain proteins in membranes to have several alpha helices.
The physical shape of a protein in its functional mode is its tertiary structure, determined largely by primary structure. Tertiary structure is affected by small molecules and ions whose concentrations in the protoplasm affect the shape and therefore the activity of many enzymes. Tertiary structure is affected by pH and heat.
Quaternary structure refers to the interaction between two or more separate polypeptides. Quaternary structure is maintained by hydrogen bonding, the interaction of hydrophobic regions, or disulfide bridges. Many enzymes consist of two or more polypeptides that must be associated to work properly; only when all are aggregated are the active sites completely formed and functional. Quaternary structure allows self-assembly of certain structures; after the individual protein monomers are formed, they automatically associate into the proper structure such as a microtubule or an enzyme complex.
Nucleic Acids
Nucleic acids are polymers composed of monomers called nucleotides. Each nucleotide is formed by the bonding of (1) a phosphate group, (2) a five-carbon sugar, and (3) a complex ring molecule that contains nitrogen and acts like a base (FIGURES AP-6 and AP-7). Nucleic acids contain only five nitrogenous bases that fall into two groups: Pyrimidines are molecules composed of a single ring, whereas purines consist of two rings. The pyrimidines are cytosine, uracil, and thymine, and the purines are adenine and guanine.
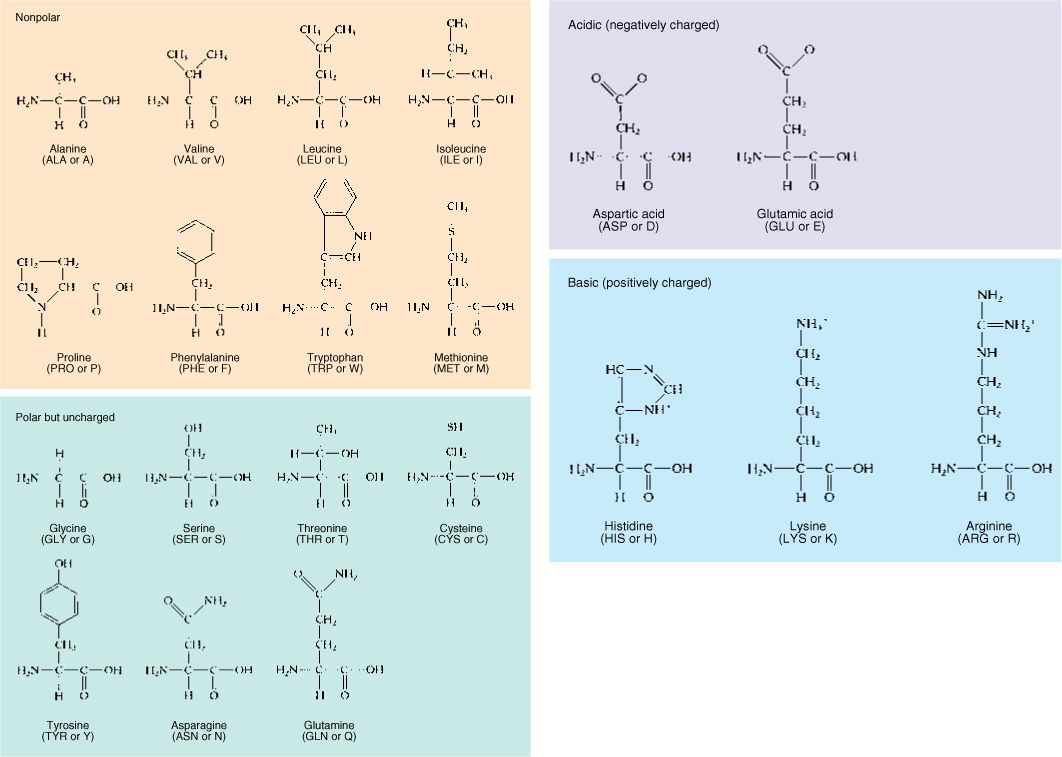
FIGURE AP-5 These are the 20 amino acids that occur in proteins. Two systems of abbreviations are used for the names of amino acids—an older system that involves three letters for each, and a newer system that uses just one letter each.
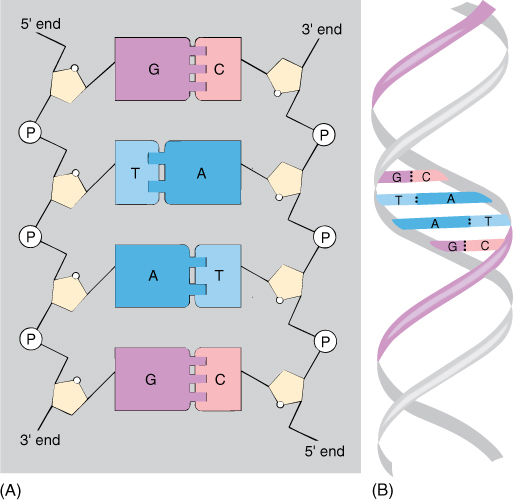
FIGURE AP-6 (A) As nucleotides polymerize into nucleic acid, the sugars are bound to each other by phosphate groups, making a long chain with the bases projecting from the side. In DNA, the bases of one nucleic acid molecule complement those of another nucleic acid, and the two form thousands of hydrogen bonds and adhere to each other, making double-stranded DNA. (B) The two nucleic acids are not straight but rather spiral around a common axis, forming a double helix.
Only two types of five-carbon sugar are ever joined to these nitrogenous bases: ribose and deoxyribose. Only four ribonucleotides (ribose-containing nucleotides) occur because thymine is never attached to ribose. Similarly, uracil is never attached to deoxyribose, so only four deoxyribonucleotides occur. The sequence of deoxyribonucleotides is the genetic information stored in the nucleus.
Lipids
Lipids are fats and oily substances that are extremely hydrophobic and water insoluble. The basic units of many lipids are fatty acids. These are long chains containing up to 26 carbon atoms with a carboxyl group at one end. If every carbon atom except the carboxyl carbon carries two hydrogens, the fatty acid is saturated; that is, it can hold no more hydrogen (FIGURE AP-8). All carbon—carbon bonds are single, and all parts of the molecule can rotate; in groups, the molecules tend to straighten and crystallize. Saturated fatty acids tend to be solid at room temperature.
If some carbons are double bonded to adjacent carbons, the fatty acid is unsaturated and the molecule has a kink. With several double bonds (polyunsaturated), the molecules are irregular in shape and cannot align well, they have little tendency to crystallize, and they melt at low temperature.
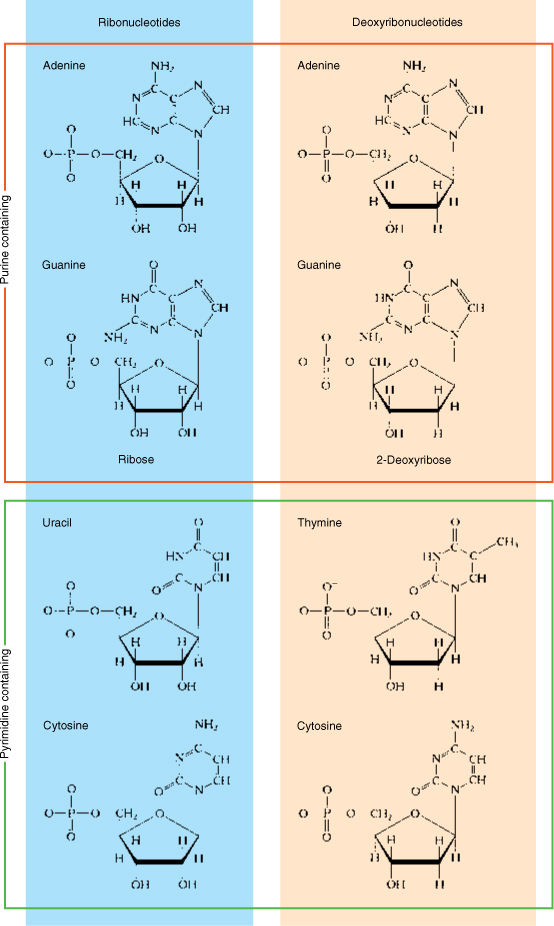
FIGURE AP-7 Purines (adenine and guanine, in the orange frame) contain two rings fused together, and pyrimidines (cytosine, uracil, and thymine, in the green frame) contain a single ring. [Note: Pyrimidines are pie shaped, and a pie can be CUT (cytosine, uracil, and thymine).] The sugar ribose is found attached only to adenine, guanine, uracil, and cytosine (blue panel), whereas deoxyribose is attached only to adenine, guanine, thymine, and cytosine (tan panel).
Fatty acids tend to polymerize readily with each other, especially when exposed to oxygen. Plants secrete fatty acids through the outer wall of their epidermal cells, and these polymerize when they come into contact with oxygen. Short fatty acids polymerize into cutin and longer fatty acids form wax.
Triglycerides are composed of three fatty acids combined with one molecule of glycerol (FIGURE AP-9A). The three fatty acids within a single triglyceride vary in length and degree of saturation and thus affect the nature of the triglyceride. Phospholipids contain glycerol, two fatty acids, and a phosphate group (FIGURE AP-9B). The phosphate group is extremely hydrophilic; therefore, these molecules have one end that tends to dissolve in water and one end that repels water. In cells, phospholipids form two-layered membranes, the hydrophobic layer of one contacting the hydrophobic layer of the other.
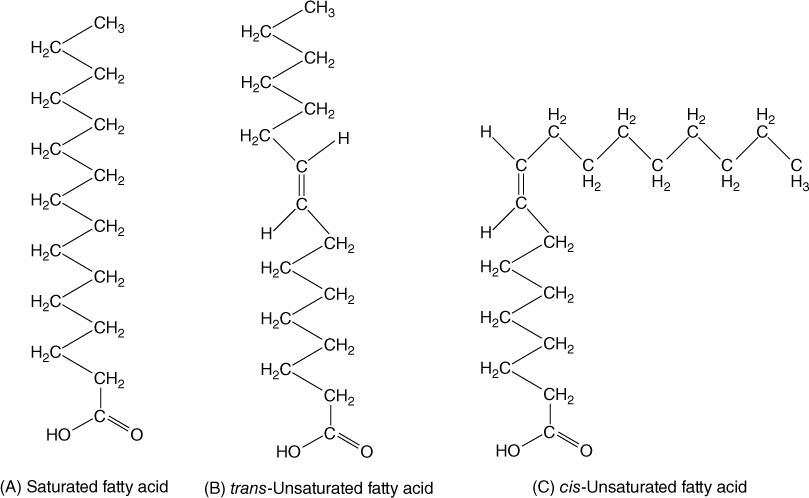
FIGURE AP-8 The various types of fatty acids. (A) Saturated, (B) trans-unsaturated, and (C) cis-unsaturated.
![]() Cofactors and Carriers
Cofactors and Carriers
Energy Carriers
The most common energy carrier is adenosine triphosphate (ATP) (FIGURE AP-10). The last two phosphate groups of ATP are attached by high-energy phosphate bonds whose bonding orbitals are not very stable. If a phosphate bond is broken and releases a phosphate group (Pi) and adenosine diphosphate (ADP), large amounts of energy are released. ADP can lose a second phosphate group to become adenosine monophosphate (AMP), which also releases large amounts of energy. The phosphate of AMP is attached by a stable, low-energy bond and cannot be used to drive an endergonic reaction. Because the conversions of ATP → ADP + Pi and ADP → AMP + Pi are both highly exergonic, they can be coupled with endergonic reactions, forcing the endergonic reactions to proceed.
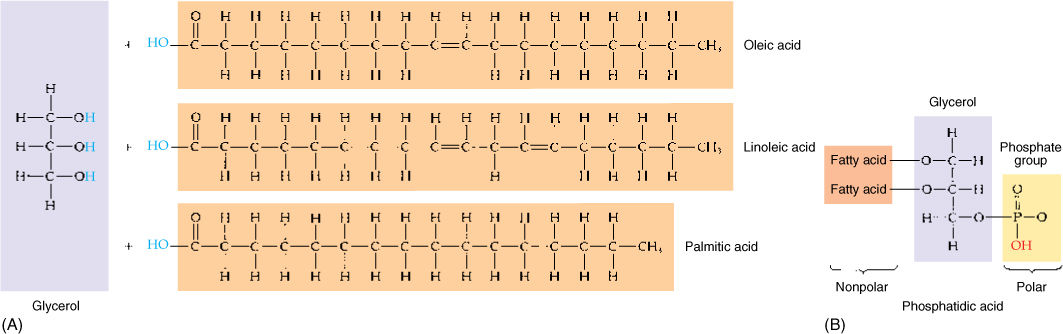
FIGURE AP-9 (A) In a triglyceride, the carboxyl group on each fatty acid reacts with a hydrogen group on the glycerol in a dehydration reaction. All parts of this molecule are nonpolar and extremely hydrophobic. (B) A phospholipid is nonpolar on one end and polar on the other. The polar end can dissolve into water; the nonpolar end can dissolve into fats.
Electron Carriers
Many metabolic reactions in plants generate molecules that have strong tendencies to donate electrons. Other reactions involve molecules needing electrons to become stabilized. Electron carriers are small molecules that react with highly electronegative compounds and take the energetic electrons away from them. The carrier itself then becomes reactive, but less so than the first molecule. When it meets the proper enzyme, it is bound to the active site and transfers the electrons to the substrate, activating it.
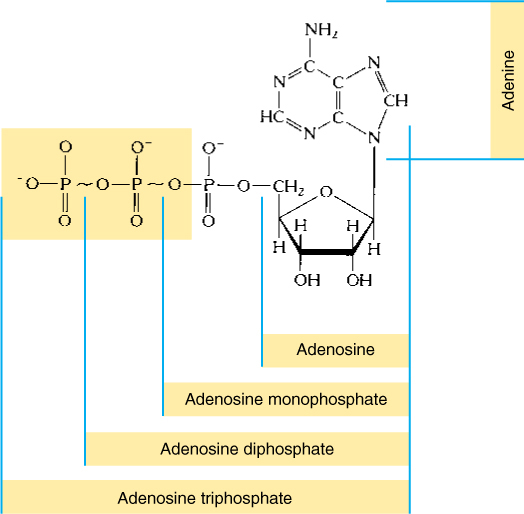
FIGURE AP-10 Adenosine triphosphate. Breaking off the last phosphate to produce ADP results in a more stable set of electron orbitals, and energy is given off. The same is true of removing the second phosphate, but not the third.
Several electron carriers exist. Nicotinamide adenine dinucleotide (NAD+) (FIGURE AP-11), nicotinamide adenine dinucleotide phosphate (NADP+), flavin adenine dinucleotide (FAD), and flavin mononucleotide (FMN). NAD+ is probably the most versatile, capable of transferring electrons between many different metabolic reactions.
The reactions that form NADH are complex, and to be strictly correct chemically, NADH should always be considered to be associated with a second proton: NADH + H+. Many textbooks do this, but others often simply write either NADH or NADH2. The same is true for NADPH.
![]() Enzymes
Enzymes
Enzymes are protein catalysts that accelerate certain chemical reactions by providing alternative mechanisms in which all energy-of-activation barriers are lower than in the original reaction mechanism. The atoms or molecules that an enzyme interacts with are its substrates, and these must fit into and be bound by the enzyme’s active site if a reaction is to occur. Many enzymes have a high substrate specificity; that is, the active site is so distinctive that only one or two specific substrates fit into it. Some active sites show specificity not for particular molecules but for specific functional groups, being able to bind to carboxyl groups, for example, or phosphate groups, with little or no regard for the nature of the rest of the molecule.
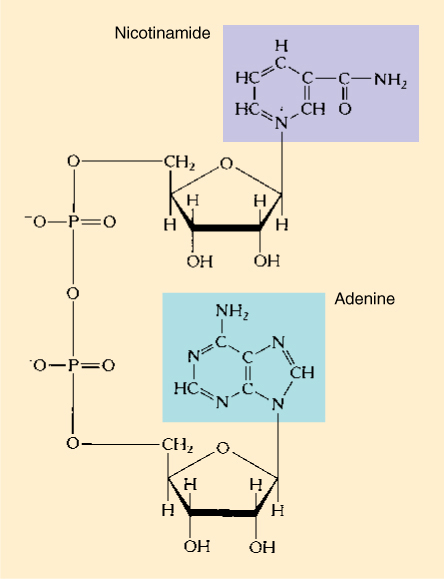
FIGURE AP-11 Nicotinamide adenine dinucleotide (NAD+). The important area is the site where the positive charge is carried (purple box); this ring can pick up two additional electrons and then deposit them elsewhere. This entire molecule is the “container” for carrying two electrons around a cell. The adenine (green box) is far removed from the site where the electrons are carried, but if the adenine is altered, NAD+ cannot enter into the proper reactions.
Cells control the activities of their enzymes by a variety of methods. One is by simply producing or not producing the enzyme. After an enzyme is present, its activity can usually be regulated by factors such as pH, temperature, and the abundance of magnesium and calcium ions. An inhibitor impairs the active site when it binds to the enzyme. A competitive inhibitor does this by occupying the active site and preventing a substrate from entering: It competes with the substrate for the same site. A noncompetitive inhibitor attaches to the enzyme at an area other than the active site; binding alters the enzyme. In many cases, the inhibitor molecule is either the product of the enzyme itself or is the end product of the metabolic pathway involving the enzyme. This is end-product inhibition.
SUMMARY
1. The metabolism of all organisms is based on universal principles of physics, chemistry, and mathematics.
2. Electronegativity is a measure of the capacity of an element to accept or donate electrons.
3. Electrons, orbitals, and entire atoms are more stable if they have less energy. If a reaction allows valence electrons to move into bonding orbitals that are more stable than the nonbonding orbitals, the product is more stable than the reactants. Energy is given off in this exergonic reaction.
4. In water, hydrogen ions are covalently bonded to oxygen, but water is a polar molecule with partially positive and partially negative ends. This permits hydrogen bonding.
5. Acids increase the concentration of protons in a solution, usually by liberating protons. Bases decrease the proton concentration, usually by liberating hydroxyl ions that bond with protons.
6. Carbon can form single, double, or triple covalent bonds. The single bonds are the most stable, whereas the bonding electrons in double or triple bonds still have considerable energy and can enter chemical reactions relatively easily.
7. If the activation—energy barrier is low, bonding orbitals can be rearranged easily, and even molecules with little energy (low temperature) can react. If orbital rearrangement is difficult, the barrier is high, and only high-energy (high-temperature) molecules can react.
8. Catalysts such as enzymes do not change the activation energy of a reaction; rather, they provide an alternative mechanism by which orbitals are more easily rearranged—all activation—energy barriers are lower. Catalysts do not change the total amount of energy liberated or absorbed during a reaction.
9. In all reactions, a reaction equilibrium is established in which there is a mixture of some reactants and some products, some forward reaction and some back reaction.
10. The physical and chemical properties of organic molecules are most strongly determined by the functional groups present; the backbone often is not so important.
11. Polymeric construction simplifies construction techniques and reduces the amount of information necessary for construction.
12. Three important carbohydrate polymers are starch, cellulose, and oligosaccharides. All are composed of monosaccharides. The type of bond and the presence or absence of branching are important.
13. The primary structure—the amino acid sequence—of proteins determines how the protein folds (secondary and tertiary structure) and how separate proteins aggregate (quaternary structure).
14. Nucleic acids are of two types: deoxyribonucleic acid, a polymer of deoxyribonucleotides, and ribonucleic acid, a polymer of ribonucleotides.
15. Many lipids contain fatty acids, either saturated or unsaturated. A triglyceride consists of glycerol bonded to three fatty acids; a phospholipid consists of a glycerol bonded to two fatty acids and a phosphate group.
16. Many small molecules cycle repeatedly between two states and serve as carriers. ATP carries energy; NADH, NADPH, FAD, and FMN carry electrons.