Botany: An Introduction to Plant Biology - Mauseth, James D. 2017
Plant Structure
Cell Structure
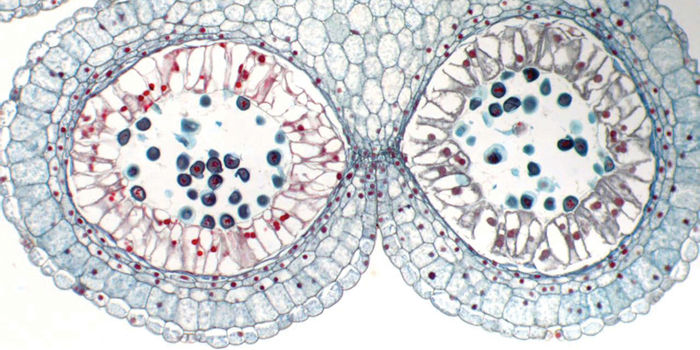
Chapter Opener Image: Several types of cells are present in this cross-section of an anther in a lily flower. In the center of each chamber are pollen grains, cells that carry genetic information during a plant’s reproduction. Each chamber is lined with a layer of tall cells that help pollen grains mature, and the large cells just below the anther’s surface will break the anther open such that the pollen grains can be carried to another flower. Each cell type has its own characteristics and modifications, and all types must work together.
OUTLINE
✵ Concepts
✵ Membranes
- Composition of Membranes
- Properties of Membranes
✵ Basic Cell Types
✵ Plant Cells
- Protoplasm
- Plasma Membrane
- Nucleus
- Central Vacuole
- Cytoplasm
- Mitochondria
- Plastids
- Ribosomes
- Endoplasmic Reticulum
- Dictyosomes
- Microbodies
- Cytosol
- Microtubules
- Microfilaments
- Storage Products
- Cell Wall
✵ Fungal Cells
✵ Associations of Cells
Box 3-1 Alternatives: Unusual Cells
Box 3-2 Plants Do Things Differently: Calcium: Strong Bones, Strong Teeth, but Not Strong Plants
Box 3-3 Botany and Beyond: The Metric System and Geometric Aspects of Cells
LEARNING OBJECTIVES
After reading this chapter, students will be able to:
✵ Describe the benefits and negative consequences of unicellular and multicellular organization.
✵ Summarize the composition of membranes.
✵ List the properties of membranes.
✵ Name the two basic cell types.
✵ Recall and define the 15 organelles found in plants.
✵ Identify the components of plant cell walls.
✵ Describe two communication methods of multicellular organisms.
 Did You Know?
Did You Know?
✵ All organisms are composed of cells.
✵ The bodies of some algae consist of just a single cell but all plants have multicellular bodies. Giant trees contain trillions of cells.
✵ Many cells must be alive to be functional (for example, cells that photosynthesize or transport sugar), whereas others must be dead (such as cells that conduct water or make up the shells of nuts).
✵ Some plant cells are large enough to be seen with the naked eye; examples are the finest strands composing cotton thread and the filaments along the torn edge of paper.
![]() Concepts
Concepts
All organisms are composed of small structures called cells. In plants, each cell consists of a box-like cell wall surrounding a mass of protoplasm, which in turn contains its own smaller parts, the organelles, such as nuclei, mitochondria, and chloroplasts (FIGURES 3-1 to 3-3 and TABLE 3-1). Cells are also the physical framework within which a plant’s metabolism occurs. Water and salts are absorbed from soil by root cells, they are transported throughout the plant by cells of the vascular tissues, and the energy of sunlight is used in leaf cells to convert carbon dioxide and water to carbohydrates. Plant reproduction is also based on cells and cell biology: Some cells in flowers produce pigments or nectar that attracts insects that carry pollen between flowers, allowing sperm cells to contact egg cells.
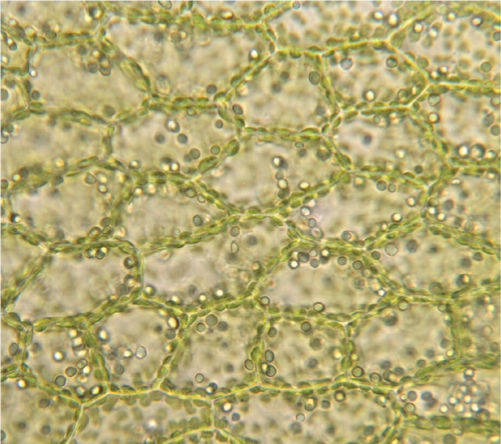
FIGURE 3-1 Light micrograph showing several basic features of plant cells. The walls and chloroplasts are easily visible. Other organelles occur but are difficult or impossible to see by light microscopy (×100).
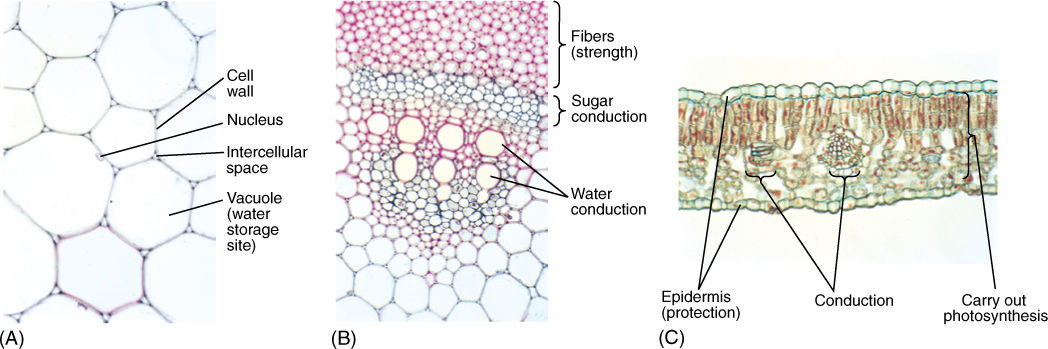
FIGURE 3-2 Plant metabolism, development, and survival depend on numerous cells working together in a coordinated, integrated fashion. (A) These cells store water in the center of a sunflower stem; they are relatively large and filled mostly with water. The cell walls and a nucleus are visible (×250). (B) Part of the system that conducts water and nutrients in a sunflower stem. Numerous types of cells occur in specific arrangements that permit efficient conduction. The large red cells in the center conduct water; the small grey cells above them conduct sugars. The cells at the top have thick red walls and provide strength to the stem (×60). (C) In this transverse section through a leaf of Ligustrum, you can see a variety of cells; those in the center carry out photosynthesis (×150).
Note: The micrograph in Figure 3-2B is 60 mm tall and has a magnification of × 60. If we were to shrink the micrograph down to life size (×1), the micrograph would be 1.0 mm tall and you would be able to see some of the cells with a magnifying glass.
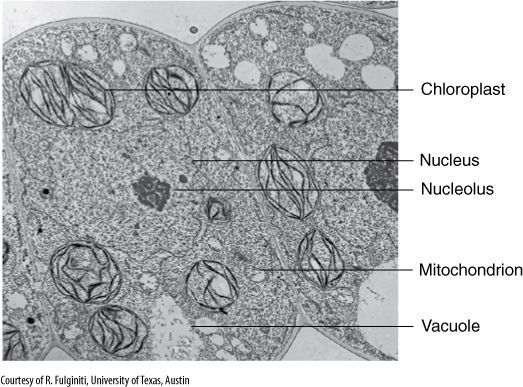
FIGURE 3-3 A micrograph made with a transmission electron microscope of leaf cells. Especially important are the numerous membranes (×17,000).
TABLE 3-1 Examples of Plant Cell Shapes and Sizes
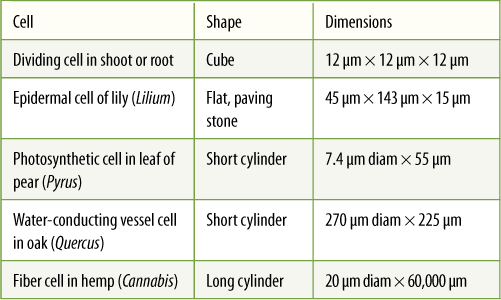
Considering the large number of living organisms and the numerous types of metabolism that must be carried out, one might suspect that there are hundreds of types of cells, but actually, just a small number of cell types exist. Most differences between organisms are due to differences in associations of their cells, not in the cells themselves. Regardless of whether a root, stem, leaf, or flower is being constructed, the same basic units—cells—are required (FIGURE 3-4). Only the cell associations and minor modifications of the cells themselves change from tissue to tissue or organ to organ.
Although only a few types of cells exist, their differences are important. Any organism composed of more than one cell (a multicellular organism rather than a unicellular one) always has several types, each specialized for different tasks (Figures 3-2 and 3-4 and TABLE 3-2). As a plant develops, the cells in various parts become especially adapted for specific tasks. This division of labor allows the entire organism to become more efficient. Unicellular organization has a significant consequence: It does not allow division of labor or specialization. Each cell must perform all tasks: sensing the environment, gathering nutrients, excreting wastes, defense, movement, and reproduction. Because each cell must perform all tasks, it cannot do any one very well. Mutations that make a cell well adapted for protection make it less adapted for other functions and therefore are selectively disadvantageous (FIGURE 3-5). The same is true for modifications that improve photosynthesis, reproduction, and so on.
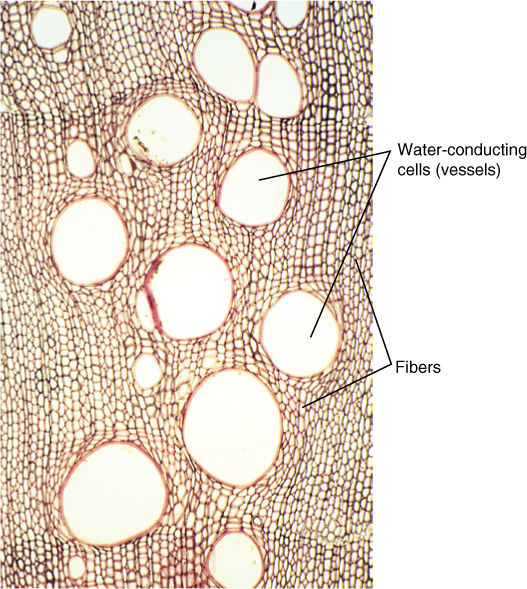
FIGURE 3-4 Transverse section of wood showing several cell types. Fibers with thick walls provide strength, and large open cells conduct water. These cells act as building blocks, and more wood is easily constructed by adding more cells. Stronger wood is produced by adding more fiber cells; more conductive wood results from adding more open cells (×100).
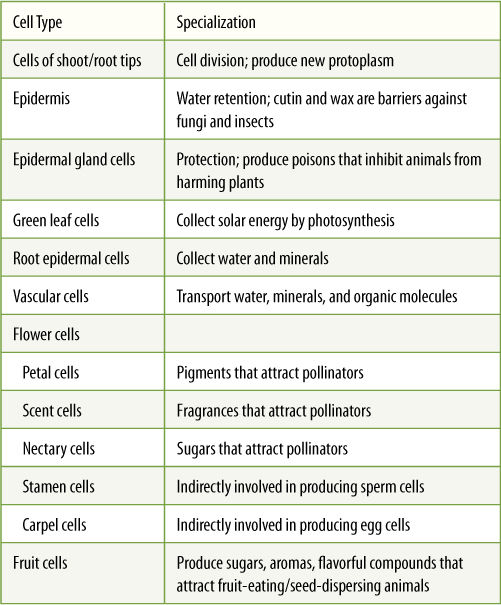
TABLE 3-2 Examples of Plant Cell Types, Specializations, and Division of Labor
Multicellularity and division of labor result in a more efficient organism, but they have negative consequences as well: As each cell becomes more specialized, it depends more on the others. If a cell evolves toward having thick walls and offering maximum protection, it must rely on other cells of the organism for photosynthesis, mineral absorption, and reproduction. Damage to one part of the organism may result in the death of all cells, even those not initially damaged, whereas in a unicellular organism, a cell dies only if it is damaged directly. Which is more advantageous selectively: a unicellular organism composed of one generalized cell or a multicellular organism composed of specialized ones? The answer is not so obvious: Both types have existed for hundreds of millions of years, and thus, both must be considered highly adaptive and successful. Also, selective advantage depends on environment: In certain habitats, unicellular organisms are better adapted; in others, multicellular organisms survive better.
Like whole individuals, cells have a life span. During their life cycle (cell cycle), cell size, shape, and metabolic activities can change dramatically. A cell is “born” as a twin when its mother cell divides, producing two daughter cells. Each daughter cell is smaller than the mother cell, and except for unusual cases, each grows until it becomes as large as the mother cell was. During this time, the cell absorbs water, sugars, amino acids, and other nutrients and assembles them into new, living protoplasm. After the cell has grown to the proper size, its metabolism shifts as it either prepares to divide or matures and differentiates into a specialized cell.
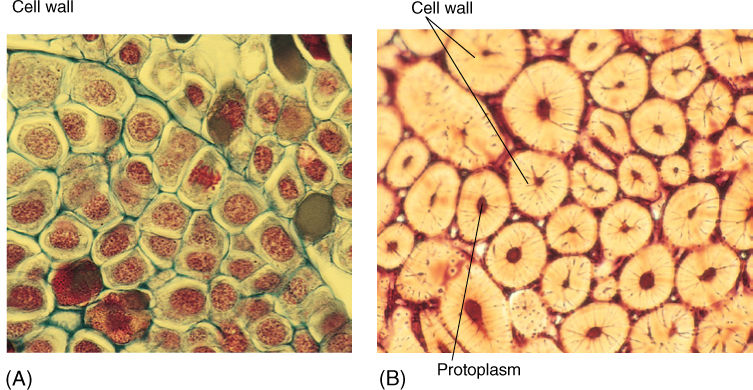
FIGURE 3-5 (A) Cells located at the growing tip of Pinus (pine) are specialized for cell growth and division. Their thin cell walls allow sugar, water, and other nutrients to enter the cells easily; however, these cells are too soft and weak to be useful as bark or the shell of a nut (×400). (B) These “stone cells” provide strength in a coconut shell (Cocos nucifera) and therefore protect the seed. Almost the entire cell volume is wall. Protoplasm was present when the cells were young, growing, and synthesizing their walls; however, after the walls were completed, cell metabolism was not necessary, and the protoplasts died (remnants are present in the tiny black hole in the center of each cell). Such modifications make it impossible for a stone cell to be a dividing, growing cell as in (A) (×200).
Both growth and development require a complex and dynamic set of interactions involving all cell parts. That cell metabolism and structure should be complex would not be surprising, but actually, they are rather simple and logical. Even the most complex cell has only a small number of parts, each responsible for a discrete, well-defined aspect of cell life.
![]() Membranes
Membranes
All cells contain at least some membranes, and cells of eukaryotes (plants, animals, fungi, and protists) contain numerous organelles composed of membranes. Membranes perform many important tasks in cell metabolism: They regulate the passage of molecules into and out of cells and organelles; they divide the cell into numerous compartments, each with its own specialized metabolism, and they act as surfaces that hold enzymes. Without membranes, life would be impossible; indeed, many things that cause death in both humans and plants—heat, cold, many poisons, alcohol—kill because they disrupt membranes.
Composition of Membranes
All biological membranes are composed of proteins and two layers of phospholipid molecules (FIGURE 3-6). If phospholipids are poured carefully onto the surface of calm water, they automatically form a single layer (a monolayer) on the water’s surface, with their hydrophilic ends forming hydrogen bonds to water molecules and their hydrophobic fatty acids projecting out of the water (FIGURE 3-7). If the water is agitated, the lipid layer doubles over and makes a bilayer in which all fatty acids are away from water and all phosphate groups are in full contact with it. Any break or tear in a bilayer membrane exposes hydrophobic fatty acids to water, and thus, membranes always reseal themselves after a rupture.
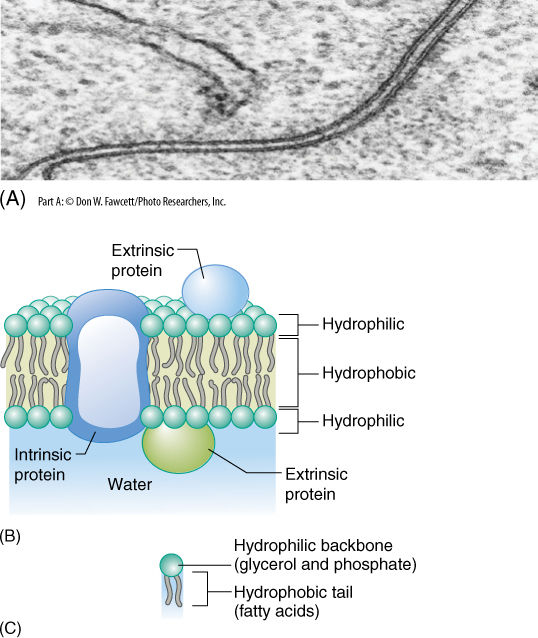
FIGURE 3-6 (A) In electron micrographs, membranes appear as two dark lines (proteins) separated by a light region (composed mostly of lipids) (×600,000). (B) Pictures such as (A) led scientists to believe that membranes consist of two layers of lipids with proteins located only on the surface; however, many proteins fit partly into the lipid bilayer or pass all the way through it. (C) A phospholipid has three parts: (1) two fatty acids, (2) a backbone of glycerol, and (3) a phosphate group.
The lipid bilayer is a very thin solution. If it contains several types of lipid, they diffuse laterally throughout the membrane because they are not bonded to each other, but they cannot diffuse vertically from the membrane into the surrounding solution.
All biological membranes contain proteins as well as lipids, usually in a ratio of 60% proteins and 40% lipid (Figure 3-6B). Most proteins have large hydrophilic regions, and thus, they associate mostly with the phospholipid phosphates and with water. Many also have large hydrophobic regions that allow them to sink into the membrane and associate with the fatty acids (FIGURE 3-8). Variations in hydrophobic and hydrophilic regions mean that various proteins sit entirely on the membrane surface, are partly immersed in it, or span it entirely, with either end projecting out of opposite sides. This allows the protein to have its active site on either or both sides of the membrane or within it. Likewise, a protein may act as a hydrophilic channel that permits small hydrophilic molecules to pass through the membrane. Proteins that are even partially immersed in the lipid bilayer are said to be intrinsic proteins. Others, extrinsic proteins (also called peripheral proteins), are located outside the membrane and merely lie next to it.
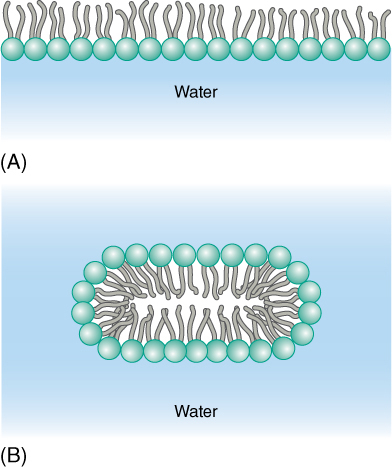
FIGURE 3-7 (A) In quiet water, phospholipids form a monolayer at the surface; by projecting away from the water, the fatty acids do not disrupt the hydrogen bonding between water molecules. (B) With agitation, phospholipids form a bilayer; this arrangement allows the greatest number of hydrogen bonds to form between phosphates and water while simultaneously minimizing the disruption of water—water hydrogen bonds by the fatty acids.
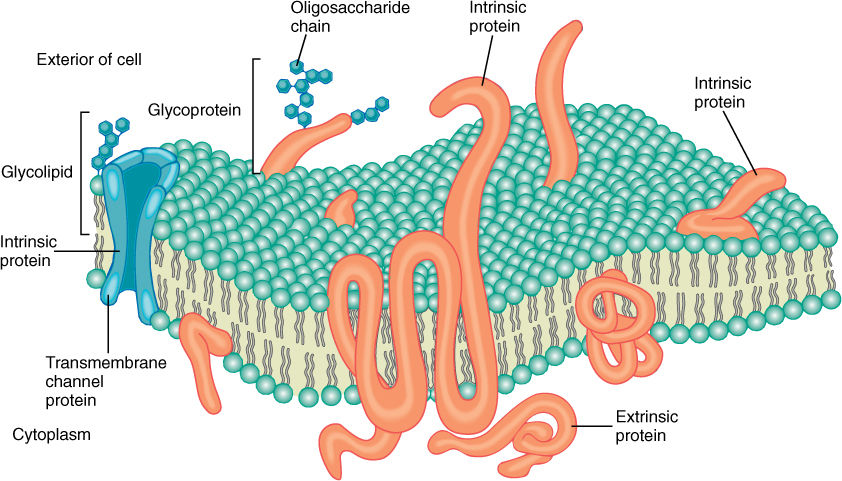
FIGURE 3-8 Intrinsic membrane proteins differ in the size, shape, and location of hydrophobic, lipid-soluble regions; consequently, some sink deep into the membrane, others span it, and some penetrate only one lipid layer.
Some intrinsic proteins contribute to the membrane’s fluid nature and, like the lipids, can diffuse laterally. Other proteins interact with adjacent proteins, forming complexes or domains (small discrete regions) different from surrounding regions of the membrane. If all membrane components could freely diffuse laterally, the membrane would become homogeneous and no differentiation could occur; however, because at least some proteins are bound to their neighbors, membranes are heterogeneous and patchy, and differentiation does take place. Because the membrane is a heterogeneous liquid, it is said to be a fluid mosaic membrane.
Some membranes contain a small amount of sugar, usually less than 8%. The sugars occur as short-chain oligosaccharides, each with about 4 to 15 sugar residues. These oligosaccharides are bound to certain intrinsic proteins, converting them into glycoproteins; rarely, sugars are attached to membrane lipids (glycolipids). Currently, we believe these polysaccharides make the membrane more distinctive and easy to recognize, an especially important feature in animals that have an immune system. Specialized protective cells attack and destroy anything that they do not recognize as a part of the animal’s own body. The glycoproteins and glycolipids occur almost exclusively on the outer surface of the membrane that covers the cell, a position that allows maximum exposure for these polysaccharides. Glycoproteins and glycolipids may be less important in plants.
Alternatives
BOX 3-1 Unusual Cells
Many plant cells are like those described in this chapter, but there are numerous types of unusual cells. Each example described here demonstrates an alternative way of organizing a cell, and of course, we can ask, “What are the consequences of each alternative?”
Dead cells. Many plant cells die as a necessary part of their development: If they remained alive they could not function as well, if at all. Cork cells in bark deposit a water-proof, enzymeresistant chemical called suberin in their cell walls, and then they die. Suberin keeps water in the plant and resists attack by microbes, but what is the consequence of dying? By breaking down their protoplasm (called programmed cell death), cork cells become much less nutritious than if they remained alive. It does an animal no good to eat cork because there are no proteins, starches, lipids, nucleic acids, vitamins, or anything else of value. An animal would waste its energy chewing and trying to digest cork. Our own epidermis cells too are dead and are more protective than they would be if they were alive and nutritious. Water-conducing cells in plants, called tracheids and vessel elements, grow to their proper size and shape then undergo programmed cell death. When fully mature, each cell consists of just a wall and an empty space where protoplasm had been. Water is pulled upward through these empty cells and if protoplasm were still present, it would impede water movement. Water-conducting cells cannot function while living—they must be dead. Other cells that function after dying are those of many plant hairs, shells around seeds, and thorns and spines.
Unusual nuclei. Cells that develop into pollen grains in flowers are surrounded by a layer of cells called the tapetum. In many species, the nucleus of each tapetum cell divides once or twice more than the cells do, so at maturity, each cell has two or four nuclei. The consequence of this seems to be that each cell then has more capacity to control a very active, dynamic cytoplasm. In contrast, cells that transport sugars throughout a plant’s body (sieve elements) destroy their nuclei during development. Unlike water-conducting cells, sugar-conducting cells cannot conduct if they are not alive. What, however, is the consequence of destroying the nucleus? We do not know why this is necessary, but we do know they become reliant on adjacent cells to control their metabolism.
Flagella. We think of plants as stationary and animals as mobile, but sperm cells of plants such as mosses, ferns, and even some seed plants (cycads) have flagella and swim. If sperm cells lacked flagella, they could not get to egg cells. There is, however, another alternative in most seed plants: Their sperm cells lack flagella but are carried passively to egg cells by elongate cells called pollen tubes, each of which grows out of a pollen grain.
Giant cells. Plant cells tend to be larger than animal cells, but both types are usually microscopic. Fortunately, several types of giant plant cells are extremely common and are so large you can see them with the naked eye. Cotton cloth and thread are made of long, slender cells, each up to 4 cm (1.5 inches) long; you can see individual cells by teasing cotton thread apart. These long cells grow from the surface of cotton seeds and then die and protect the seeds by forming a fluffy, soft layer. Most paper is made from water-conducting tracheids of wood, each tracheid being several millimeters long. These cells are easy to see by simply moistening a piece of paper and pulling it in two: The tiny filaments protruding from each edge are single cells. For these cells, a beneficial consequence of their extraordinary length is that it is easier to pull water through a few long cells as opposed to many short cells (if both types are the same width). Very unusual giant cells occur in some of the plants that ooze milky white liquid when poked (milkweeds in the family Asclepiadaceae (now classified as part of Apocynaceae) and spurges in Euphorbiaceae). The milky sap is always at least mildly toxic, often extremely toxic, and is produced by cells called laticifers (FIGURE B3-1). If quite a bit of liquid drips out, it is probably produced by long, tubular cells. Tubular laticifers begin development as ordinary, small cells, but they continue to grow even after surrounding cells have reached their full size and stop enlarging. The laticifers elongate, pushing into spaces between other cells, becoming ever longer, and often branching and growing in multiple directions. Unlike cotton cells and tracheids, these must remain alive at maturity, and have hundreds of nuclei directing the synthesis of milky sap. One consequence of being giant, branched tubular cells is that even if an animal chews into just one small part of a plant, toxic sap from a wide area of the plant will flow to that site and deter the animal.
Unusual cells are mentioned here to show that even though most plant cells are rather similar to each other, there are alternatives, and with the alternatives come consequences. Most of this book focuses on the most typical, most ordinary cells, tissues, and metabolisms, but always think about the possibility of alternatives and how those might affect the biology of the plant.
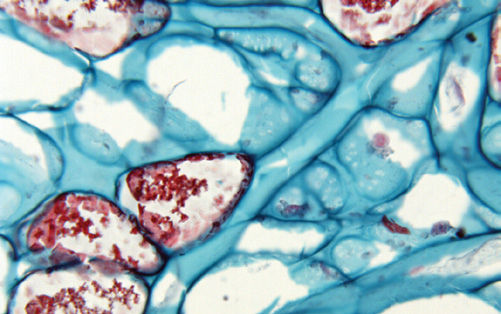
FIGURE B3-1 The long, branched tubes here are laticifers; each has branched many times (most branches are out of view of this high magnification), and all tips elongate and intrude between other cells. Individual cells are very long and have hundreds of nuclei (×200).
Properties of Membranes
Membranes have several important properties. First, they can grow. Membranes are formed molecule by molecule in certain regions of the cell; then entire pieces of membrane are moved as small bubbles or vesicles to different sites in the cell. When the vesicle of preformed membrane arrives at the growing membrane, the two fuse (FIGURE 3-9).
In addition to permitting movement of membrane pieces, membrane fusion allows the transport of material. The volume inside the vesicle (the vesicle lumen) may be filled with substances that must be accumulated, broken down, or otherwise metabolized at the vesicle’s destination. Vesicle movement may also release material to the outside of the cell (Figure 3-9). This exocytosis is a means to excrete almost anything: wastes, debris, mucilage, proteins, and polysaccharides. For example, roots slide through the soil by secreting a slippery, lubricating mucilage formed within root cells, packaged into vesicles that migrate to and fuse with the cell’s surface membrane; this mucilage is then released to the exterior. In many flowers, nectar is secreted from glands by exocytosis. Endocytosis is basically the opposite process: A small invagination forms in the outer membrane then pinches shut, creating a new vesicle that contains extracellular material. Endocytosis is especially common in algae and other microscopic organisms that take in food particles this way, but it is not known how common it is in plants. Permeability is an important property of membranes. All biological membranes are selectively permeable membranes (also called differentially permeable), meaning that certain substances cross the membrane more easily and rapidly than others (FIGURE 3-10). Because large regions of a membrane are mostly lipid, membranes are more permeable to hydrophobic substances than to anything that carries an electric charge; however, if charged compounds such as inorganic salts, sugars, and amino acids could not enter cells at all or if they could enter only slowly, cells would starve.
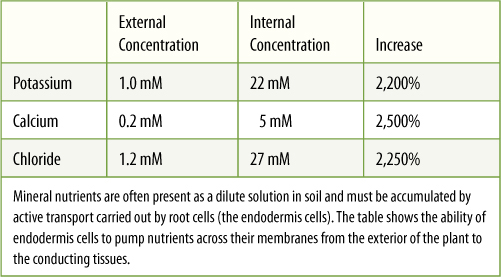
Movement of charged substances is assisted by large intrinsic proteins that span the membrane and act as hydrophilic channels through it; this is facilitated diffusion. Other proteins, called molecular pumps, actually bind to a molecule on one side of the membrane, and then by using energy, the protein changes shape and releases the molecule on the other side. By using this active pumping, called active transport, cells accumulate substances until the interior concentration of solute far exceeds the exterior concentration. A pump working in the opposite way can actively transport materials out of the cell (TABLE 3-3). Except for the pumps, the membrane must be impermeable to the molecule; it would do no good to have a molecular pump for a type of molecule that could easily leak back through.
All life depends on the principle of compartmentalization, the formation of many compartments, each specialized for a particular process such as producing a particular substance or using a particular precursor. For example, our blood is compartmentalized away from our digestive system: The two must work together, but they must not be mixed together. If all the monomers, polymers, sugars, salts, enzymes, and vitamins of an organism were mixed together, some reactions would occur at random but would not be orderly enough to be considered life. Instead, cells are filled with many compartments—the organelles—each surrounded by its own unique selectively permeable membrane. Because the protein channels and pumps are made under the guidance of the nucleus, the cell can control the numbers and types of channels and pumps it makes and inserts into each type of organelle membrane. Organelles thus have membranes with different permeabilities or pumping capacities, depending on the instructions generated by the nucleus.
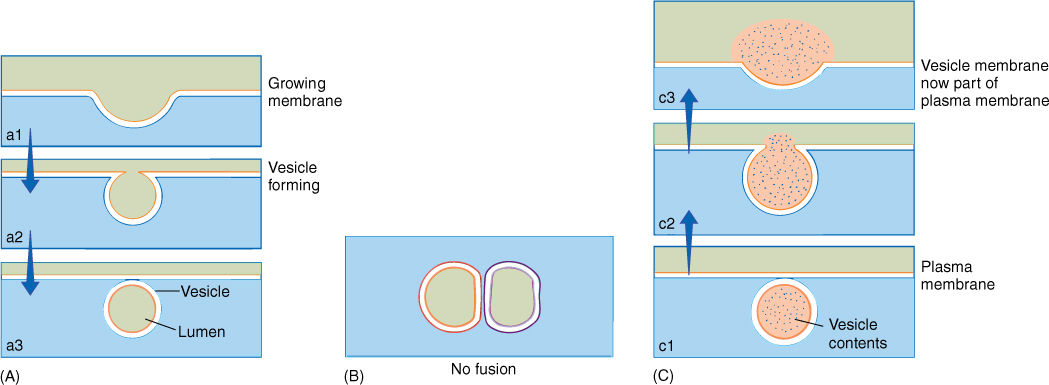
FIGURE 3-9 (A) Numerous organelles produce small vesicles that move through the cell, either remaining discrete for long periods or fusing with other organelles or vesicles. In order to fuse, two membranes must be quite similar in composition. (B) Here, a single membrane bilayer is shown as a double line, one orange and the other black. Membrane dissimilarity prevents two compartments from fusing; for example, the nucleus does not fuse with mitochondria or chloroplasts. (C) When a vesicle fuses with the membrane at the cell surface, its contents are deposited outside the cell and the vesicle membrane becomes part of the cell surface. The vesicle membrane and the plasma membrane have been drawn with the same colors to indicate their similar composition.
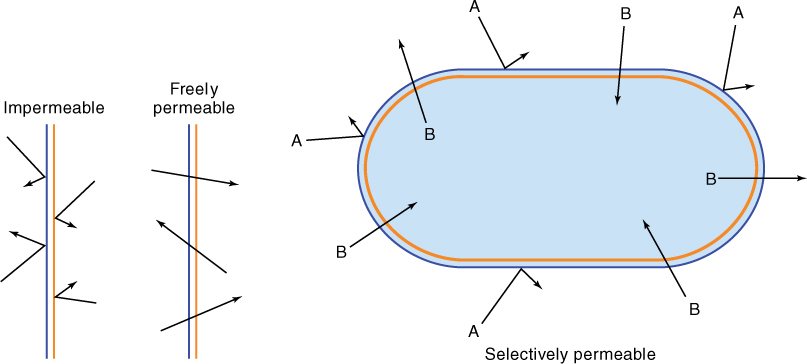
FIGURE 3-10 An impermeable membrane allows nothing to pass through, whereas a freely permeable membrane stops nothing. A selectively permeable membrane allows certain materials to pass through more readily than others, with the result that chemical concentrations on the two sides differ. This membrane is impermeable to substance A but permeable to B.
TABLE 3-3 Active Transport in Roots of Corn (Zea)
A freely permeable membrane, which allows everything to pass through quickly, would be rather useless for a cell, as would an impermeable membrane, one that does not allow anything through at all (Figure 3-10 and TABLE 3-4).
The last important property of membranes is that they are dynamic, constantly changing. Cell membranes are not simply established and left unchanged throughout the life of the cell. Instead, new components are constantly being inserted and old ones removed. If the function of the cell is always the same, the old and new components are similar, but if the cell must change its function, newly inserted components are different from the retracted ones. As the nature of the membrane changes, the nature of the cell changes also.
TABLE 3-4 Summary of Transmembrane Movement

![]() Basic Cell Types
Basic Cell Types
At the most basic level, both cells and organisms can be classified as either prokaryotic or eukaryotic. Prokaryotic cells are simpler than eukaryotic ones and are found only in domains Bacteria (bacteria and cyanobacteria) and Archaea (archaeans). It is hypothesized that prokaryotic cells represent the most archaic lines of evolution and that eukaryotic cells evolved from them.
Eukaryotic cells, found in plants, animals, fungi, and protists, are more complex than prokaryotic cells. The most striking difference, the one that gives them their name, is the presence of a true membrane-bounded nucleus in eukaryotic cells. In addition, eukaryotic cells have many organelles that allow them to be more diverse and complex, both morphologically and physiologically.
![]() Plant Cells
Plant Cells
Although various parts of a plant—roots, wood, bark, leaves, and flower parts—appear to be quite diverse, virtually all of their cells have all of the following organelles; exceptions are rare. As each type of cell develops, certain organelles may become modified and more or less abundant, but usually none is lost completely (FIGURE 3-11).
Protoplasm
All cells, either prokaryotic or eukaryotic, are made of a substance called protoplasm (Figure 3-5) (the protoplasm of a single cell is called its protoplast). This name was given early, when it was thought that protoplasm was a distinct substance such as water, oxygen, or iron and that one of its properties was life itself. We now know that protoplasm is a mass of proteins, lipids, nucleic acids, and water within a cell; except for the wall, everything in the cell is protoplasm, composed of the following organelles.
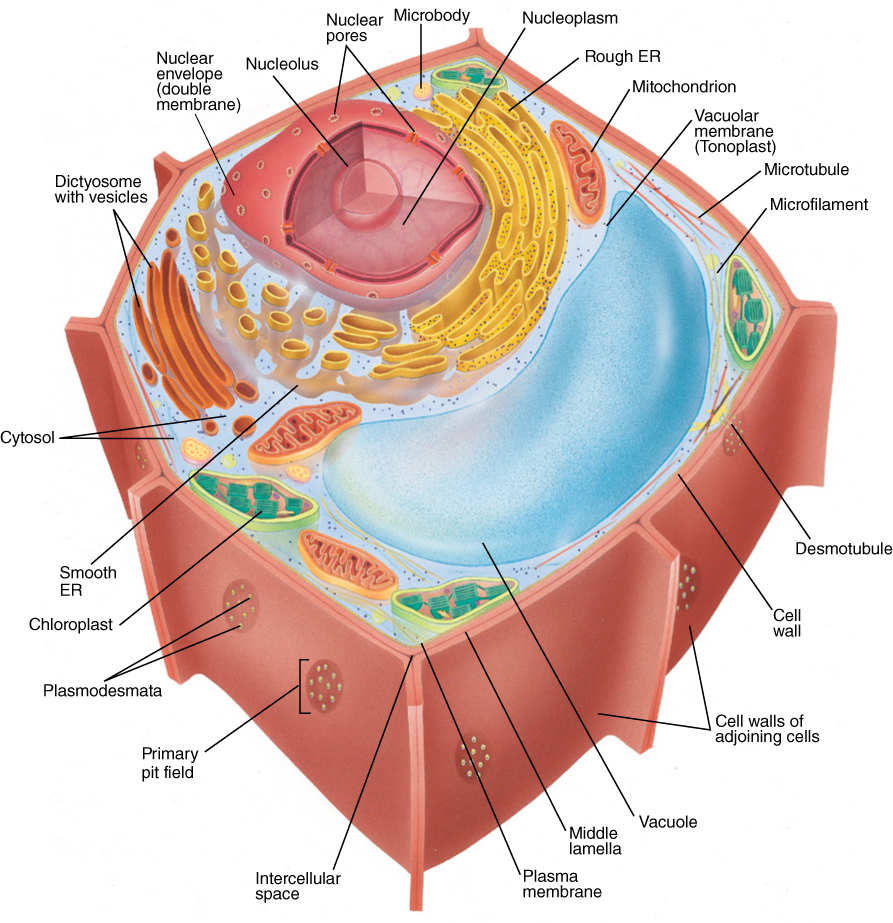
FIGURE 3-11 Generalized plant cell.
Plasma Membrane
The plasma membrane (less frequently called the plasmalemma) is the membrane that completely covers the surface of the protoplasm (FIGURE 3-12). Because it is the outermost surface of the protoplast, it must be impermeable to harmful materials and permeable to beneficial ones; therefore, it is selectively permeable. Molecular pumps in the plasma membrane actively transport needed molecules inward and pump others outward for secretion. Because one side of the plasma membrane faces the external environment and the other side faces the cell, the two sides are quite different, especially in the types of protein they contain. We know very little about the proteins and lipids that make up the plasma membrane because the membrane does not separate cleanly from the protoplast. Almost all attempts to isolate the plasma membrane for study have experienced considerable contamination from other parts of the cell.
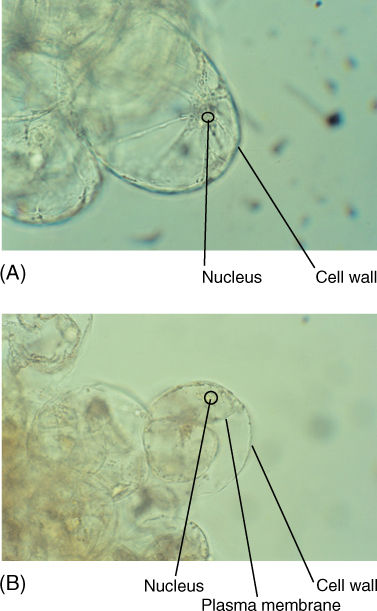
FIGURE 3-12 (A) A healthy, growing cell. Its protoplast is pressed firmly against the cell wall, and thus, its plasma membrane is not visible (×160). (B) These cells have been treated with salt to draw water out of the cell, causing the protoplast to shrink. As a result, the plasma membrane is visible (×160).
Nucleus
The nucleus (plural, nuclei) (Figure 3-11) serves as an archive, or permanent storage place, for the organism’s genetic information. This text is just a brief introduction to plant biology. No one can imagine the number of pages required to store all of the information necessary for building and maintaining a single cell; however, all of that information must be stored in the DNA inside every nucleus, and the storage must be safe and permanent1. Information is useless unless it, or copies of it, can be retrieved and used. The nucleus carries out information retrieval by making copies of specific parts of the DNA whenever that information is needed. The copy is not DNA, but a type of ribonucleic acid called messenger RNA. An exciting area of research involves understanding how the nucleus, in response to signals from the rest of the cell, searches its DNA for the needed information and then makes copies of it without accidentally copying other similar information.
The nucleus of a eukaryotic cell is always surrounded by a nuclear envelope composed of an outer membrane and an inner membrane. The nuclear envelope separates nuclear material from the rest of the cell, and it contains numerous small holes, nuclear pores (FIGURE 3-13), involved in the transport of material between the nucleus and the rest of the protoplasm. Nuclear pores have a complex structure and exert control over the movement of materials. If a nucleus is extracted from a cell and placed into water, it swells; this can happen only if the pores prevent material from oozing out as the nucleus absorbs water. Prokaryotes have no nuclear envelope; instead, DNA is simply mixed with the rest of the cell contents.
Within the nucleus is a substance called nucleoplasm. Like “protoplasm,” this name was given before the composition was understood; we now know that nucleoplasm is a complex association of (1) DNA; (2) enzymes and other factors necessary to maintain, repair, and read DNA; (3) histone proteins that support and interact with DNA; (4) several types of RNA; and (5) water and numerous other substances that are necessary for nuclear metabolism. Nuclear DNA is always closely associated with histones, and this complex of the two is known as chromatin. As a cell ages or its metabolic activities change, so do the nucleus and nucleoplasm. In cells undergoing rapid cell division (e.g., cells in root tips, shoot tips, young leaves, and flower buds), the DNA, histones, and duplicating enzymes may dominate the nucleoplasm; however, in mature cells that are not dividing, messenger molecules and reading enzymes may be more abundant.
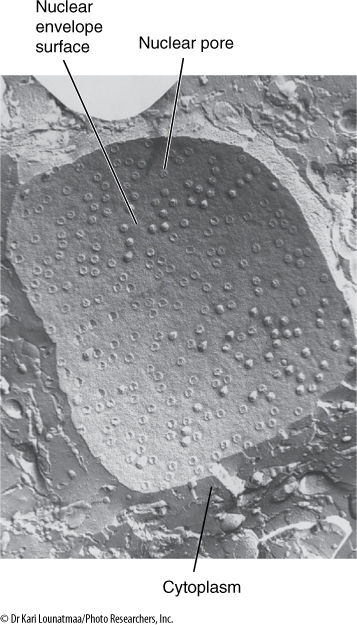
FIGURE 3-13 A preparation called a freeze-fracture. The cell is frozen and then tapped to cause it to break. The lipids of the membranes are weak when frozen, and thus, the fracturing often follows the membranes, separating one lipid layer from the other. Here the fracture passed irregularly through the cytoplasm and then entered the nuclear envelope. The nuclear envelope consists of the outer and inner membranes, with a space separating them; the two membranes fuse together at the nuclear pores. Nuclear pores may be distributed uniformly over the nuclear surface or may occur in bands or patches in some species (×60,000).
1 Actually we do know, now in 2016. The model plant Arabidopsis thaliana has 135 million “bits” of information (nucleotide base pairs composed of A, T, G, and C). Each page in this book can hold 7,670 characters, so if the genome sequence of A. thaliana were written on these pages, we would need 17,601 pages. Loblolly pine, Pinus taeda, has much larger nuclei with more information (20 thousand million base pairs, or 20 billion base pairs) and for that we would need 2,627,118 pages.
Plants Do Things Differently
BOX 3-2 Calcium: Strong Bones, Strong Teeth, but Not Strong Plants
Most plants and animals need hard parts. Wood is strong enough to support the weight of a tree, and bones play a similar role in animals. Seeds are often protected by resistant shells such as those of walnuts and almonds, and animal shells protect clams and oysters. Our teeth are so tough that they can chew through almost anything. Although plants and animals use hard parts for similar roles, plants rely on thick, tough cell walls, whereas animals use calcium salts.
Would it be possible for plants to use bone-like material? We can analyze this as a set of alternatives and their consequences. The present alternative—wood—consists of cellulose and a chemical called lignin. Both are carbohydrates that a plant itself makes through photosynthesis, and thus, they are readily available. And both are remarkably inert, having little impact on other aspects of the plant’s metabolism. In contrast, calcium and its salts participate in many metabolic pathways, and building or resorbing shells, bones, or teeth has a broad impact on cell physiology. Shells consist of calcium carbonate, and as animals use carbonate ion (CO32-) to build a shell, the acidity of the protoplasm is altered. Furthermore, animals can digest part of their shells if they need the calcium elsewhere, and this liberation of carbonate will again affect the pH. This is tolerable for marine organisms because they use carbonate from the surrounding seawater rather than from their own protoplasm so their pH is not affected. If the shell is resorbed later, the liberated carbonate is likewise dumped outside the animal into the seawater.
Animals like us—with an internal skeleton—use calcium phosphate in our bones and teeth. Calcium carbonate’s tendency to alter pH is too dangerous for us, and our skeleton cannot use seawater as a carbonate reservoir. The phosphate ion (PO42-) that we use has little effect on a cell’s acidity, and, furthermore, if we resorb bone for some reason, the liberated phosphate is a resource for many other metabolic reactions. It is not a liability at all.
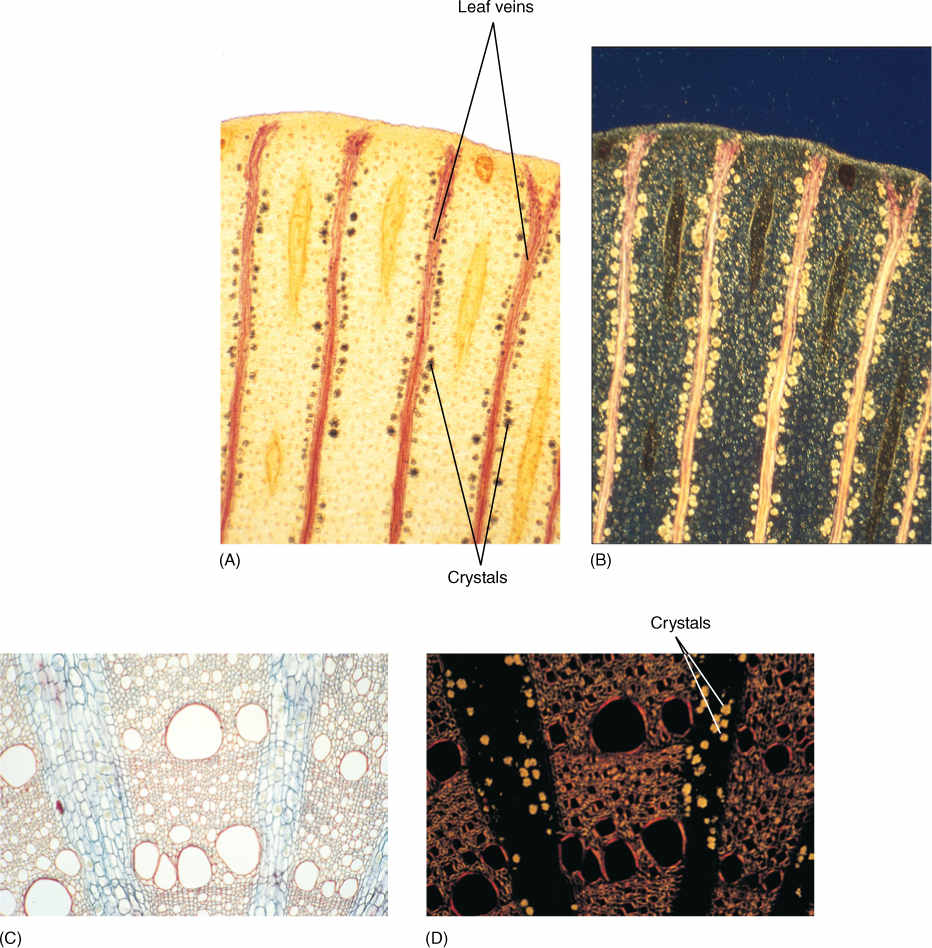
FIGURE B3-2 (A) A leaf clearing of maidenhair tree (Ginkgo), showing several red-stained leaf veins that conduct sugars out of the leaf. Such veins are the targets of aphids and other sucking insects (×15). (B) The same tissue, in polarized light (×15).(C) A cross-section of Aristolochia wood; crystals are present in the two bands of tissue with blue-stained walls, see (D). This is a soft tissue in wood and is the site where sugars and other nutrients are stored (×50). (D) The same tissue as (C), but with polarized light (×50).
Plants too must be careful with calcium and its salts. Calcium carbonate’s disturbance of cell acidity is just as dangerous for plants as it is for us, but plants do not use calcium phosphate either, perhaps because it requires too much phosphate. Most soils have just marginal amounts of phosphate and not quite enough to let plants grow optimally. If plant cells had walls made of bone, they would need much more phosphate than is available in soil.
Like animals, plants must carefully control the concentration of dissolved calcium within their cells. The concentration must be kept at extremely low levels; otherwise, it would interfere with metabolism. Roots block the entry of calcium, but so much leaks in accidentally that the plant must get rid of the excess somehow. Plants crystallize the calcium by producing oxalic acid. Calcium and oxalic acid react to form calcium oxalate, which is insoluble and forms crystals. Crystallized calcium is more or less inert and cannot interfere with the plant’s metabolism. Calcium oxalate forms only tiny individual crystals rather than big bone-like structures.
Wood with lignin is not as strong as bones or teeth, but its synthesis does not affect cell acidity. Plants can make all of the wood they need without relying on rare minerals from the soil. Plants and animals may do things differently, but there are sound biological principles underlying the differences.
Inside every nucleus is one, two, or rarely several bodies called nucleoli (singular, nucleolus) (FIGURE 3-14), areas where the components of ribosomes are synthesized and partially assembled. Each ribosome contains a large amount of ribosomal RNA copied from ribosomal genes in the chromatin.
Nuclei are large, complex organelles, and they occupy a major fraction of the volume (up to 50%) of small dividing cells. However, most cells enlarge greatly after they stop dividing and become mature (for example as a young leaf expands to become a mature leaf); as cells expand, the nucleus becomes a smaller percentage of cell volume (TABLE 3-5). In certain conducting cells (sieve tube members in phloem), the nucleus breaks down during cell differentiation, and the mature cell functions for several weeks or months without a nucleus. Our red blood cells are also enucleate (without a nucleus) at maturity. Both of these cell types are exceptional; they are enucleate only while performing a very limited type of metabolism, and they die shortly after losing the nucleus. Several types of plant cells are multinucleate.
Central Vacuole
Within young, small cells are organelles, vacuoles, that have just a single membrane, the vacuole membrane, also called the tonoplast. Vacuoles often appear to be empty (Figure 3-11 and TABLE 3-6) because they store mostly water and salts that cannot be preserved for microscopy; however, they sometimes contain visible crystals, starch, protein bodies, and various types of granules or fibrous materials in addition to water and salts.
As a cell grows and enlarges, vacuoles expand and merge until there is just one large central vacuole. Because it contains primarily water and salts, the central vacuole can expand rapidly, forcing the cell to grow rapidly as well (FIGURE 3-15). Animal cells must synthesize complete protoplasm to grow, but plant cells need only increase the amount of vacuolar water. For example, as flower buds open, petals expand from tiny to full-sized in just a few hours by simply enlarging their vacuoles, not by making new cells. Over a long period, plants must produce additional proteins, membranes, and organelles or else they would become almost pure water.
In addition to cell growth, the central vacuole functions in storage of both nutrient reserves and waste products. In seed cells, vacuoles may be filled with protein that will be used when the seed germinates, perhaps 10 to 50 years after the material was deposited in the vacuole. Calcium regulates the activity of many enzymes, and plant cells keep protoplasmic calcium concentrations at the proper level by moving calcium into the vacuole, where it reacts with oxalic acid and crystallizes into an inert form. Other nutrients such as potassium may move in and out of the vacuole on a daily basis. The water-soluble pigments in many flowers, fruits, and red beets occur in vacuoles as well (FIGURE 3-16).
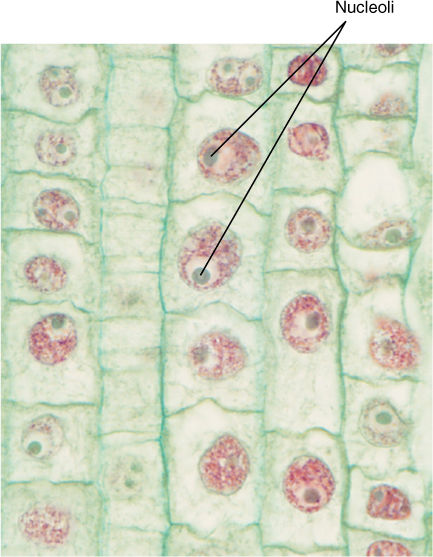
FIGURE 3-14 Nucleoli usually stain more intensely than the rest of the nucleus; occasionally, they stain a distinct color. Root tip of hyacinth (×250).
TABLE 3-5 The Relative Volumes of Organelles in Plant Cells
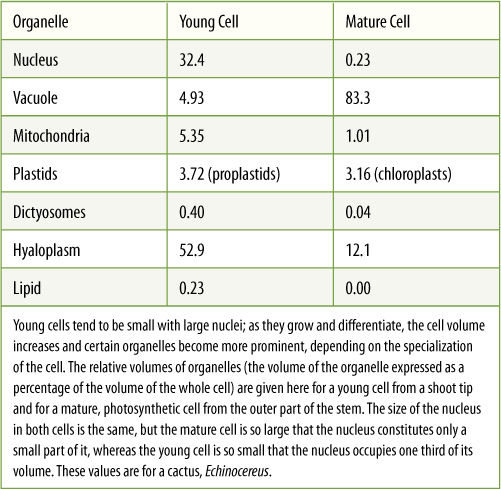
TABLE 3-6 Concentrations of Nutrients in Vacuoles Isolated from Barley (Hordeum) Leaf Cells
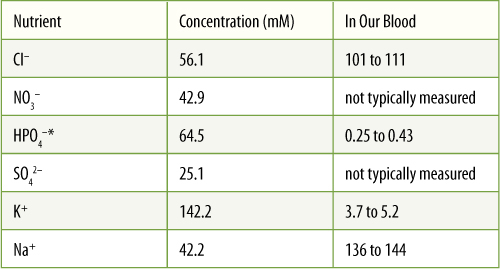
*Often abbreviated Pi
A system to excrete wastes never evolved in plants; instead, metabolic waste products are pumped across the vacuole membrane and stored permanently in the central vacuole. The tonoplast is otherwise impermeable to these wastes; therefore, they cannot leak back into the cytoplasm where they would be harmful. Holding waste inside forever does not sound like an optimal situation, but it actually is selectively advantageous: Because most of these compounds are noxious and bitter, they deter animals from eating the plants. Mutations that result in excretion might make the cells taste good, which would be selectively disadvantageous.

FIGURE 3-15 Azalea flower buds open overnight, the petals and stamens expanding rapidly as each cell fills its central vacuole with water. The flower maintains its shape by hydrostatic pressure: central vacuoles pressing cytoplasm firmly against cell walls.
The central vacuole is a digestive organelle as well. As other organelles age and become impaired, they fuse with the tonoplast and are transported into the central vacuole, where digestive enzymes break them down. The liberated monomers are transported back into the rest of the cell, where they are used again. In animal cells, which do not have central vacuoles, this task is carried out by small vacuoles called lysosomes.
Cytoplasm
If the nucleus and vacuole are excluded from the protoplasm, the remaining material is referred to as cytoplasm and contains the following structures.
Mitochondria
Cells store energy as highly energetic but fairly unreactive compounds, such as sugars and starches. To use the energy, such compounds must be broken down and their energy must be used to synthesize new compounds that are both highly energetic and very reactive, the most common being adenosine triphosphate (ATP). Because these reactions involve high-energy, reactive compounds, they occur more safely within a discrete organelle than freely in the cytoplasm, where the intermediates might accidentally react with other cell components. Mitochondria (singular, mitochondrion) are the organelles that carry out this cell respiration (FIGURE 3-17).
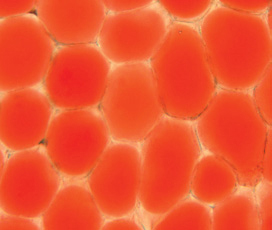
FIGURE 3-16 The central vacuoles of these petal cells contain dissolved pigments, making the vacuoles visible. The pale area between each oval of pigment is intercellular space, cell walls, nuclei, and all of the protoplasm other than vacuole (×300).
Many steps of respiration are mediated by enzymes bound to mitochondrial membranes. The enzymes are located adjacent to each other so that the product of one reaction (which becomes the reactant in the next reaction) is passed directly to the next enzyme; this controls the highly reactive intermediates (FIGURE 3-18). Mitochondrial membranes are folded, forming large sheets or tubes known as cristae (pronounced CHRIS tee; singular, crista). This folding provides room for large numbers of enzymes. Reactions that do not involve highly reactive intermediates take place in the liquid matrix between the cristae. Around this complex of cristae and matrix is a second membrane, the outer mitochondrial membrane, which gives shape and a little rigidity to the mitochondrion. The outer membrane is rather freely permeable, but the inner mitochondrial membrane, which forms the cristae, is selectively permeable and has numerous pumps and channels.
Mitochondria have their own DNA and ribosomes, both of which are different from those of the rest of the cell. Mitochondrial DNA is a circular molecule and lacks histones; the ribosomes are small and resemble those found in prokaryotes.
Mitochondria are dynamic organelles; they can grow larger, either as the cell does or as the cell’s need for respiration increases. They are often about 1 µm in diameter but up to 5 µm in length and are even much longer in some cases. Mitochondria can also divide into two, increasing the number of mitochondria per cell. Under rare conditions, the mitochondria in some species fuse together, forming a single giant mitochondrion in each cell. Mitochondria occupy between 1% and 25% of the cell volume (Table 3-5); the higher values are similar to those for animal cells. Because of their ability to divide and fuse, the actual number of mitochondria per cell (usually in the range of 100 to 10,000) may not be as important as their volume.
Plastids
Plastids are a group of dynamic organelles able to perform many functions. One prominent activity is photosynthesis, carried out by the green plastids, chloroplasts (Figure 3-1). Diverse types of metabolism occur in other classes of plastid: synthesis, storage, and export of specialized lipid molecules; storage of carbohydrates and iron; and formation of colors in some flowers and fruits. Plastids are the site of synthesis of amino acids: isoleucine, valine, and those that contain aromatic rings (phenylalanine, tryptophan, and tyrosine); or are derived from aspartate (lysine, threonine, and methionine). Each metabolism is associated with a particular type of plastid; as an organ changes, its plastids may also change, extensively altering their membranes and proteins. Plastids are found in all plants and algae but never occur in animals, fungi, or prokaryotes.
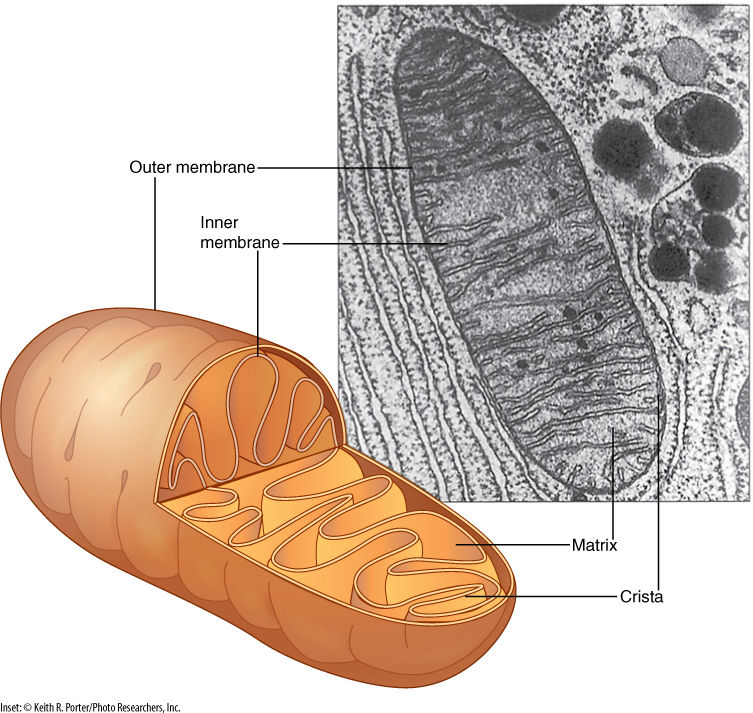
FIGURE 3-17 The inner mitochondrial membrane is folded into plate-like cristae, giving it a large surface area; many respiratory enzymes are intrinsic proteins embedded in the crista membrane. The large surface area makes it possible to contain many copies of each enzyme, and more reactions can occur simultaneously.
FIGURE 3-18 Many enzymes must work together. The product b of enzyme 1 is the reactant of enzyme 2. A) If the enzymes are in solution, they may not be close together and b must diffuse at random until it encounters enzyme 2; in the meantime, it may accidentally react with something else. B) If they are located side by side on a membrane, enzyme 1 can pass b directly to enzyme 2, not only speeding up the overall reaction but also eliminating the chance of accidental reactions.
Like mitochondria, plastids always have an inner membrane and an outer membrane and an inner fluid called stroma (FIGURE 3-19). Plastids also have ribosomes and circular DNA that is not associated with histones. Plastids grow and reproduce by pulling apart. Plastids of young, rapidly dividing cells are called proplastids and are very simple. The inner membrane has a few folds but little surface area. When exposed to light, proplastids develop into chloroplasts, which are green owing to the presence of the photosynthetic pigment chlorophyll (TABLE 3-7). Because many intermediates of photosynthesis are highly reactive, energetic compounds, the controlling enzymes must be incorporated into the membranes, just as in mitochondria. This requires the inner membrane to become more extensive and elaborately folded. Membrane sheets, thylakoids (pronounced THIGH la koids), project into the stroma. This increase in membrane area also provides more space for the insertion of the photosynthetic pigments: Chlorophyll has a lipid-soluble tail and, thus, is part of the membrane. In certain regions, the thylakoids form small bag-like vesicles that stack together. The stack of vesicles is called a granum (plural, grana). A key feature of photosynthesis is the active transport of protons (H+) into a small space to build up an electrical charge; the grana vesicles are needed to accumulate these protons from the stroma. The actual conversion of carbon dioxide to carbohydrate occurs in the stroma, catalyzed by enzymes free in solution rather than bound to any of the membranes.
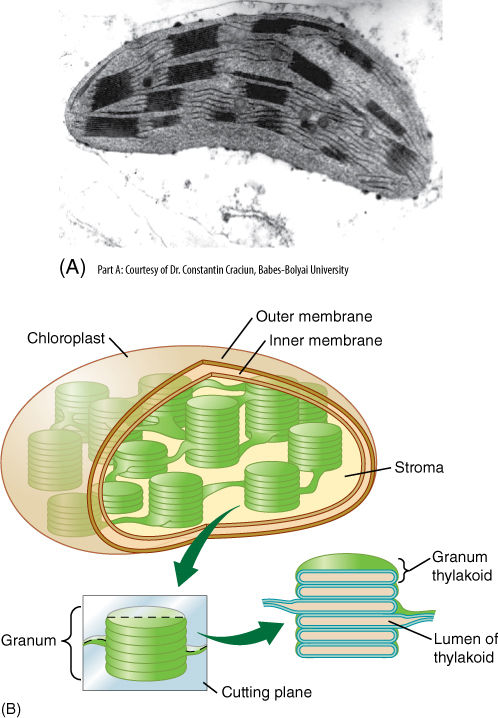
FIGURE 3-19 (A) All plastids have an outer and an inner membrane, but in chloroplasts, the inner membrane is extensive and highly folded. The single membranes are thylakoids, and the multiple membranes are actually stacks of flattened thylakoid vesicles; the stacks are called grana. Like mitochondrial cristae, the extra surface area of the inner membrane provides room for many copies of each enzyme. The photosynthetic pigment chlorophyll is part of the membrane lipid layer (approximately × 30,000). (B) The grana are interconnected by thylakoid membranes, and the liquid stroma surrounds the grana. Because the thylakoid membranes are selectively permeable, the concentrations of chemicals inside the thylakoid space differ from those in the stroma. This is essential for photosynthesis.
TABLE 3-7 Types of Plastids

Chloroplasts are larger than mitochondria, approximately 4 to 6 µm in diameter. An individual leaf cell may contain as many as 50 chloroplasts. When chloroplasts photosynthesize rapidly, they produce sugar faster than the cell can use it, and thus, it is temporarily polymerized into starch grains inside the chloroplasts.
In plant tissues that cannot photosynthesize (roots, bark, wood), proplastids develop into amyloplasts, which accumulate sugar and store it as starch for months or years as in starchy seeds like wheat, rice, corn, or vegetables such as potatoes and yams (FIGURE 3-20). Each amyloplast produces large starch grains that virtually fill the stroma; few internal membranes are present. In starchy vegetables, amyloplasts constitute the bulk of the tissue. If exposed to light, amyloplasts can be converted to chloroplasts. Nonphotosynthetic parasitic plants such as Conopholis lack chloroplasts but do have amyloplasts; no plant is ever without some form of plastid.
In some flowers and fruits—for example, tomatoes and yellow squash—bright red, yellow, or orange lipids accumulate in plastids as they differentiate into highly colored chromoplasts (FIGURE 3-21). An extensive, undulate system of membranes is present, but no grana, and the pigments may be present either as part of the membrane or as discrete droplets, plastoglobuli (singular, plastoglobulus). As fruits ripen, chloroplasts often synthesize large amounts of lipid pigments, alter their thylakoids, and convert to chromoplasts, as when apples and tomatoes change from green to red. Lipid pigments are present in low amounts as small plastoglobuli even in leaf chloroplasts, but they are masked by the abundant green chlorophyll; they can be seen in autumn when cold weather causes chlorophyll, but not lipids, to be broken down and the leaves turn red or yellow.
Many cells have large, unpigmented plastids that have neitherchlorophyll nor lipid pigments. These leucoplasts may be involved in various types of synthesis: Many types of fats and lipids are synthesized only in plastids and then transported to other organelles and inserted into their membranes. “Leucoplast” is a purely descriptive term—any colorless plastids, including proplastids and amyloplasts, can be considered leucoplasts.
Iron is an essential nutrient for plants and animals and is stored attached to a large protein. Although almost identical to human ferritin, the plant protein is called phytoferritin. Ferritin is found throughout animal cytoplasm and nuclei, but phytoferritin is stored almost exclusively in plastids. As much as 80% of the iron in leaves may be found in chloroplasts, and the leucoplasts and amyloplasts of seeds often have especially large amounts.
Ribosomes
Immersed in the protoplasm are ribosomes, particles responsible for protein synthesis (FIGURE 3-22). They are complex aggregates of three molecules of RNA (ribosomal RNA) and approximately 50 types of protein that associate and form two subunits. Compared with animal cells such as those in the liver and pancreas, most plant cells synthesize little protein and have few ribosomes; however, some do produce large amounts of proteins and are rich in ribosomes, such as the protein-rich seeds of legumes including peas and beans, and the cells that secrete the digestive enzymes of insectivorous plants. Each molecule of messenger RNA is long enough for 6 to 10 ribosomes to attach to it and read it simultaneously. The ribosomes are thus bound together by the messenger RNA, forming a cluster called a polysome.
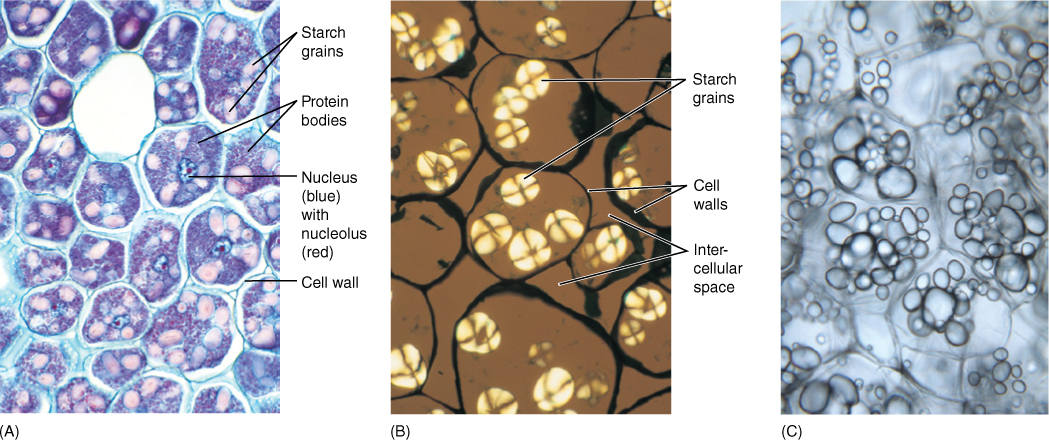
FIGURE 3-20 (A) Cells from a developing bean seed. The large pink bodies are starch grains; the small bluish-red ones are protein bodies. These colors are not natural. The material was stained to make the colorless starch and protein visible. The starch grains occur in plastids called amyloplasts; as the bean germinates, the starch is used as a source of energy. When the starch decreases, the amyloplasts convert to chloroplasts and turn green (×150). (B) Starch grains that have been photographed in polarized light rather than being stained. Substances that have an orderly, crystalline structure shine brightly in polarized light (×200). (C) Free-hand section of fresh potato; each cell is filled with numerous oval starch grains. The thin walls of these cells are just barely visible (×200).
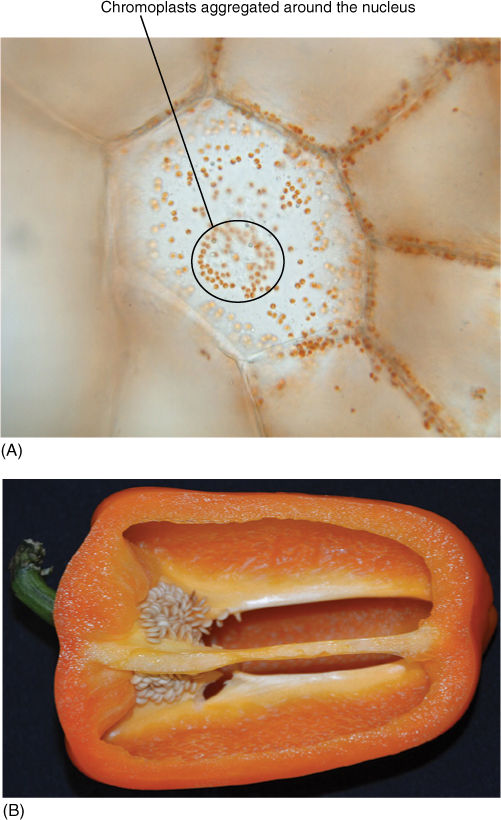
FIGURE 3-21 (A) Each tiny orange dot is a chromoplast; this is from an intensely bright orange pepper. (B) Just a few chromoplasts provide brilliant color. Chromoplasts are located in the periphery of the cell because the central vacuole occupies most of the cell center. The largest cell appears to have chromoplasts everywhere, but we are just viewing the nearest wall, not the center of the cell. Many chromoplasts are aggregated around the nucleus.
Endoplasmic Reticulum
A typical plant cell is so small that diffusion, the random movement caused by molecular motion, carries a molecule to many different parts of the cell every second. Diffusion may be the only means by which small molecules like monosaccharides and cofactors move around the cell, but some large molecules such as proteins are carried by the endoplasmic reticulum (ER), a system of narrow tubes and sheets of membrane that form a network throughout the cytoplasm (Figure 3-22).
A large proportion of a cell’s ribosomes are attached to the ER, giving it a rough appearance; consequently, this ER is called rough ER (RER). As an attached ribosome synthesizes a protein, it passes through the membrane and collects in the lumen. If the protein is a storage product, as in seeds of legumes, it merely remains in the ER, which may become quite swollen; however, if the protein is to be secreted (digestive enzymes, adhesive proteins), then its accumulation causes regions of the ER to form vesicles. These detach, move to the plasma membrane, and fuse with it, releasing their contents to the cell’s exterior by exocytosis (Figure 3-9). In many cases, the protein must be modified before export; if so, the ER pinches off only very small vesicles from regions where the ER is close to another organelle, the dictyosome, which carries out the protein modification.
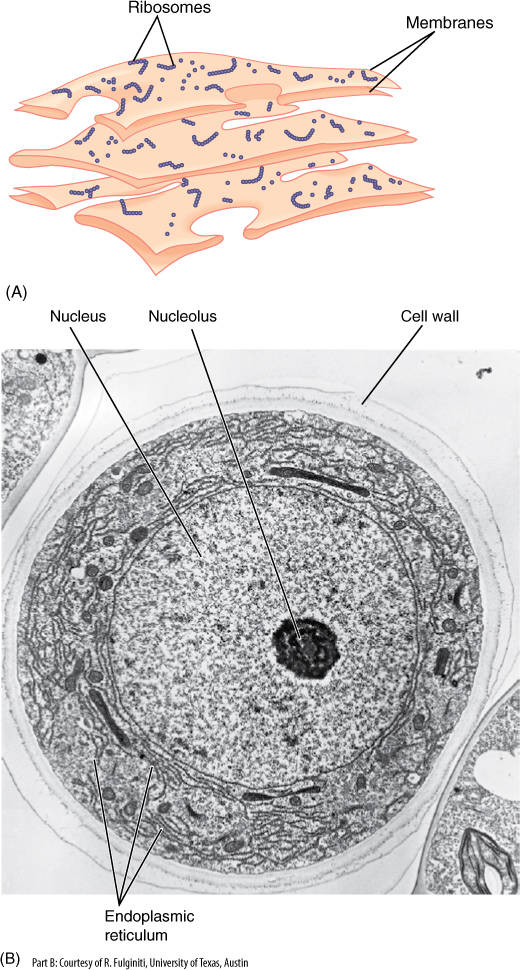
FIGURE 3-22 (A) Ribosomes only rarely occur free in the cytoplasm; instead, they are usually attached to membranes such as the type shown here, called endoplasmic reticulum (ER). The amount of ribosomes attached is greater in cells that produce abundant protein (such as protein-rich seeds) and less in cells that synthesize little protein (such as the water-storage cells of Figure 3-2A). (B) Electron micrograph of a cross-section of a hair cell that secretes protein-rich mucilage. The mucilage is synthesized in the ER, which is unusually abundant for a plant cell (×3,000).
ER that lacks ribosomes is smooth ER (SER), and it is involved in lipid synthesis and membrane assembly. The lipids range from simple to extremely complex; as they are produced, they are inserted into the membrane, and then vesicles form and pinch off, carrying the new membrane to other parts of the cell. After the ER-derived vesicles reach the correct organelle, they fuse with it, and the vesicles become a new patch of membrane in the organelle. SER is abundant only in cells that produce large amounts of fatty acids (cutin and wax on epidermal cells), oils (palm oil, coconut oil, and safflower oil), and fragrances of many flowers.
Dictyosomes
Much of the material secreted by a cell must first be modified by a dictyosome, a stack of thin vesicles held together in a flat or curved array (FIGURE 3-23). ER vesicles accumulate on one side of the dictyosome and then fuse together and form a wide, thin vesicle called a cisterna (plural, cisternae) that becomes attached to the dictyosome (FIGURE 3-24). Soon more ER vesicles gather next to this one and form a new cisterna. The first cisterna becomes embedded more deeply in the dictyosome as more vesicles accumulate on that side, which for obvious reasons is the forming face. At the other side, the maturing face, vesicles are being released; their contents have been processed. After separation, vesicles can move to the plasma membrane and release their contents (FIGURE 3-25). The outer edges of dictyosomes form an interconnected network of curving tubes, and these may absorb the contents from the center of the dictyosome cisterna and then detach and move away. It is not known why some dictyosomes concentrate material in the central cisternae, whereas others use the peripheral ones.
Dictyosomes can form large, complex associations. In animal cells that secrete very large amounts of protein, hundreds of dictyosomes associate side by side and form a cup-shaped structure called a Golgi body or Golgi apparatus. In the Golgi body, the dictyosomes’ maturing faces are on the inner side of the cup, whereas the forming faces and associated ER are on the outside. Dictyosomes only rarely aggregate into Golgi bodies in plants, one example being in root hairs: All of the dictyosomes are part of a giant Golgi body located at the tip of the hair where growth and cell wall formation occur.
Different types of processing may occur within a dictyosome: modification of the vesicle’s membrane or modification of its contents. If the vesicle is to fuse with the plasma membrane after release, the vesicle membrane must be made similar to the plasma membrane. If the vesicle is to remain separate from all other organelles, acting as a storage vesicle, its membrane must be made unique and incapable of fusion.
Alteration of the vesicle’s contents involves addition of sugars to proteins, forming glycoproteins. Sugar-containing proteins occur in the plasma membrane, the cell wall, and as storage products in seeds. Strong evidence is accumulating that dictyosomes also polymerize sugars into polysaccharides used in cell wall construction.
Movement of vesicles from ER to dictyosomes and then to other sites was hypothesized on the basis of the presence of numerous vesicles located between ER and dictyosomes and the similarity of the contents of dictyosome cisternae to those of vesicles found fusing with the plasma membrane. This hypothesis predicts that if a secretory cell is given a very brief dose of radioactive sugar or protein precursors, much of the radioactivity is soon found in the ER and later in the dictyosomes. Even later, both are nonradioactive, but vesicles at the plasma membrane and external material are radioactive. Experiments have verified both this prediction and the related hypothesis that new membrane is synthesized in the ER and then transported by vesicles to growing organelles. Although organelles appear to be distinct entities when viewed by light or electron microscopy, they are actually highly interrelated by this membrane flow. All membranes of the cell, except the inner membranes of mitochondria and plastids, actually constitute just one extensive system, the endomembrane system.
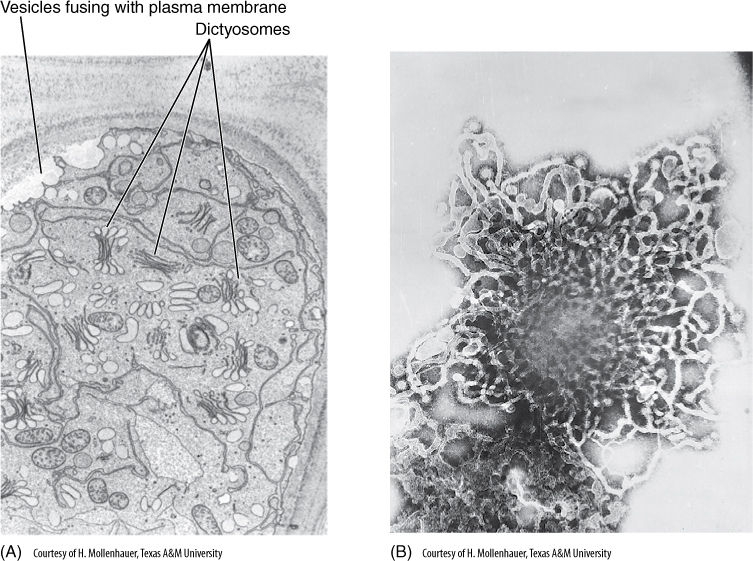
FIGURE 3-23 (A) With rapid dictyosome activity, as in these mucilage-secreting cells, the cytoplasm may almost fill with dictyosome vesicles, and fusion of vesicles with the plasma membrane is so abundant that the plasma membrane appears scalloped (×12,000). (B) Face view of dictyosome isolated from a plant cell; the network of peripheral tubules is visible (×60,000).
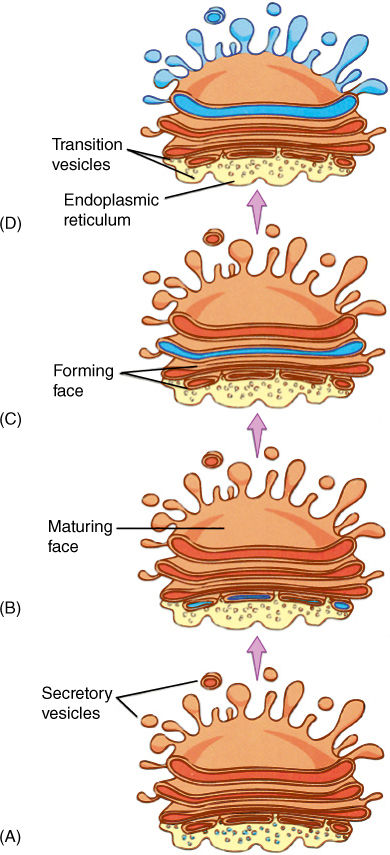
FIGURE 3-24 (A) Vesicles derived from ER migrate a short distance to a dictyosome-forming face. (B) At the forming face, the vesicles fuse into a new dictyosome vesicle, called either a vesicle or a cisterna. (C) The cisterna “moves through” the dictyosome as more vesicles form on one side while other vesicles are released from the maturing face (D).
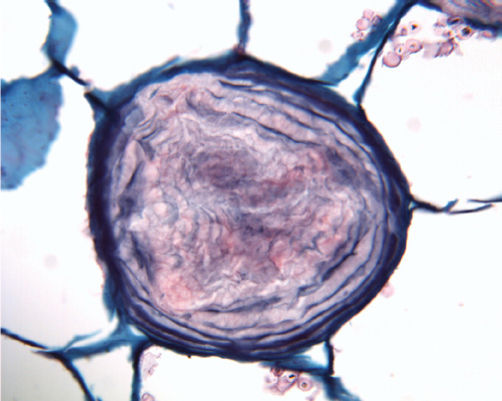
FIGURE 3-25 This is a mucilage cell. Mucilage-filled dictyosome vesicles secreted mucilage (stained purple) outside the protoplast (see Figures 3-9C and 3-24A). Mucilage could not penetrate the cell wall, so as it accumulated, the protoplast had to decrease in volume by removing water from the central vacuole. The cell died (probably by programmed cell death), and now the cell consists only of cell wall and mucilage (× 250).
In a few cases, it has been possible to measure the rate of membrane flow. The insectivorous plant Drosophyllum has leaves covered with glands that produce a sticky secretion. The fluid contains digestive enzymes that are processed by dictyosomes in the gland’s cells. After an insect has been caught, each gland produces a visible drop of digestive fluid at a rate of 1.3 µm3 per minute. Dictyosome vesicles are about 0.15 µm in radius; thus, each has a volume of 0.014 µm3, and approximately 100 must fuse with the plasma membrane every minute to deliver the 1.3 µm3 of fluid and enzymes. Each vesicle has a surface area of 0.27 µm2, so 27 µm2 of vesicle membrane fuses with the plasma membrane every minute during secretion. From calculations of the average cell volume and surface area of the plasma membrane, these secretory cells could double their plasma membrane every 20 seconds. It is important for these cells to be able to retract membrane material from the plasma membrane and recycle it. The mucilage-secreting cells of root tips are more leisurely: Each cell has approximately 800 dictyosomes that together contribute 14 to 26 µm2 of membrane per minute to the plasma membrane, which has a surface area of about 1,000 µm2. Each dictyosome receives a new cisterna every 20 to 40 minutes.
Microbodies
Viewed by electron microscopy, cells are seen to contain numerous small, spherical bodies approximately 0.5 to 1.5 µm in diameter (FIGURE 3-26). These are so nondescript that it is not easy to tell exactly what they are: ER vesicles, dictyosome vesicles, or distinct organelles; however, by using special chemical reactions and stains, it has been possible to determine the contents and even the types of reactions in these bodies. Some are organelles now called microbodies, and there are two classes: peroxisomes and glyoxysomes. Both types isolate reactions that either produce or use the dangerous compound peroxide, H2O2. If peroxide were to escape through the microbody membrane, it would damage almost anything it encountered; however, both types of microbody contain the enzyme catalase, which detoxifies peroxide by converting it to water and oxygen.
Botany and Beyond
BOX 3-3 The Metric System and Geometric Aspects of Cells
![]() The Metric System
The Metric System
In 1791 the French Academy proposed that a new system of measurement be adopted to simplify and regularize weights and measures. This metric system is used not only for all scientific measurements but also for engineering, commercial, and ordinary purposes in every country of the world except the United States (see the conversion tables at the end of this box). The fundamental unit is the meter; however, for long distances, the kilometer is used, and for small objects, centimeters, millimeters, and so on are more convenient. Volumes are also based on the length measurements, and weights are based on the gram, the weight of 1 cm3 of water. The metric system has two advantages: (1) Measurements and objects made in one country can be understood or used immediately in any other country, except the United States, and (2) measurements in one unit can be converted easily to other units: 35,645 meters is 35.645 kilometers, but 35,645 feet is how many yards or miles?
1 kilometer (km) = 1,000 meters (m)
1 meter = 1,000 millimeters (mm)
1 millimeter = 1,000 micrometers (µm)
1 micrometer = 1,000 nanometers (nm)
109 nm = 106 µm = 103 mm = 100 m = 10—3 km
These are based on multiples of 1,000. The centimeter (cm) is only 1/100 meter or 10 mm, but it is such a convenient size for measuring plants and animals that it is used even though it disrupts the otherwise perfect regularity of increments of 1,000.
![]() Geometry of Cells
Geometry of Cells
A quantitative understanding of plants provides extremely valuable insight into their biology. The following formulas are simple but will allow you to think about plants more precisely:
area of a rectangle = length × width
area of a triangle = 1/2 length × width
volume of a cubic space = length × width × height
diameter of a circle = 2 × radius
circumference of a circle = 2π × radius
area of a circle = π × radius2
surface area of a sphere = 4π × radius2
volume of a sphere = 4π/3 × radius3
volume of a cylinder
= area of base × height
= π × radius2 × height
surface area of a cylinder
= circumference of circle × height + ends
= π × diameter × height + ends
Many plant cells are almost cube shaped and have edges about 20 µm long; the volume of such a cell is the volume of a cube
= L × W × H
= 20 μm × 20 μm × 20 μm
= 8,000 μm3
Do not forget to multiply the units to get µm3 (see FIGURES B3-3A—D).
Now consider the nucleus of the cell. Many plant nuclei have a diameter of about 8 µm, and thus, their radius is 4 µm. The volume of a nucleus is V = (4)(3.141)/3 × (4 µm)3 = 4.188 × 64 µm3 = 268 µm3. The nucleus, although an important part of a cell, may not be a large part. In our average cell of 8,000 µm3 volume, the proportion that is nucleus is (volume of nucleus)/(volume of cell) × 100% = 268 µm3/8,000 µm3 × 100% = 3.3%, a very small part of the cell volume.
Mitochondria are about 1 µm in diameter and are quite variable in length; let us assume that we have measured many and found the mean length to be 5 µm. Mitochondria are almost cylinders, so we can use that formula to calculate their volume:
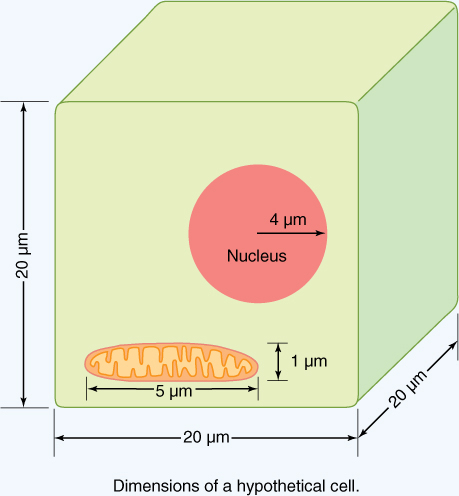
FIGURE B3-3A Dimensions of a hypothetical cell. Notice that the mitochondrion is not drawn to scale.

FIGURE B3-3B Most of us have a set of standard metric weights readily available. A dollar bill (of any denomination) weighs exactly 1 gram (g), 2 pennies weigh 5 grams, 15 quarters and 3 pennies weigh almost exactly 100 grams, and a small Post-it Note® weighs one-tenth (0.1) of a gram.

FIGURE B3-3C Many rulers have centimeters (cm) along one side and inches along the other.
volume of a cylinder
= area of a circular cross-section × height
= (3.141) × (0.5 µm)2 × (5 µm)
= 0.7852 µm2 × (5 µm)
= 3.93 µm3
Mitochondria often constitute about 7.5% of a cell’s volume; for our typical cell, that would be 8,000 µm3 × 0.075 = 600 µm3. How many actual mitochondria would there be in each cell? If each mitochondrion has a volume of 3.93 µm3, the number is (total volume)/(volume of each mitochondrion) = 600 µm3/3.93 µm3 = 153 mitochondria in each cell.
The surface of a cell or organelle is the space through which material enters and leaves the object by diffusion, facilitated diffusion, or active transport. Objects with large surface areas can absorb or lose material more rapidly than those with smaller surface areas. Also, surface area is a measure of the room available for placing membrane-bound enzymes and other intrinsic membrane proteins. In our hypothetical cubical cell, each surface has an area of 20 µm × 20 µm = 400 µm2. A cube has six sides, and thus, the total surface area of the plasma membrane is 6 × 400 µm2 = 2,400 µm2. That is about 1/30 of the surface area of the period at the end of this sentence. The nucleus of the cell has a surface area of 4π × (radius)2 = 12.56 × 16 µm2 = 201 µm2, less than 1/10 that of the cell. The cylindrical mitochondria each have a surface area of their outer membrane of 3.141 × 1.0 µm × 5 µm = 15.7 µm2 (we are ignoring the ends). What, however, of their inner membrane? Assume that measurements reveal that each crista is a cylindrical tube 0.2 µm wide and 0.9 µm long and each mitochondrion contains an average of 45 cristae; then the surface area of each is 3.141 × 0.2 µm × 0.9 µm = 0.565 µm2. The 45 in each mitochondrion have a total surface area of 45 × 0.565 µm2 = 25.4 µm2 available for respiratory enzymes. The 177 mitochondria of the cell would have a total inner surface area of 4,496 µm2, greater than that of the plasma membrane area (2,400 µm2). Thus, folding the inner membrane into cristae provides a large amount of extra surface area for enzymes.

FIGURE B3-3D The metric system measures volumes in liters (L) and milliliters (mL). A tablespoon (foreground) is 14.7 mL, and a teaspoon (the next one back) is exactly 5 mL. One cup is slightly less than 250 mL. Drops of milk are irregular, but 20 drops of water equal 1.0 mL. The measuring cup and the beaker use the abbreviation “ml” but the correct abbreviation is “mL.”
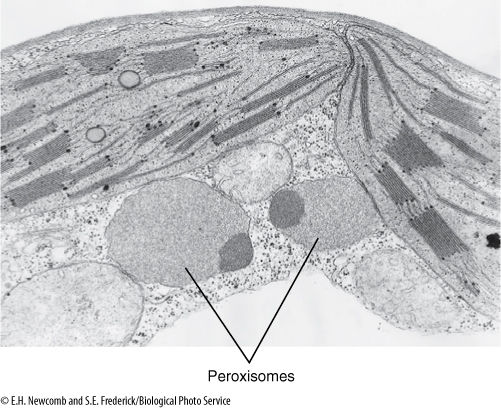
FIGURE 3-26 Electron micrograph showing two chloroplasts with two peroxisome-type microbodies next to them. In many plants, photosynthesis is accompanied by a process called photorespiration, in which peroxisomes produce the amino acid glycine (approximately × 66,000).
Peroxisomes are involved in detoxifying certain byproducts of photosynthesis and are found closely associated with chloroplasts; in animals, peroxisomes are abundant in liver and kidney cells, where they break down foreign compounds that contaminate our food. Glyoxysomes, which occur only in plants, are involved in converting stored fats into sugars. They are important during the germination of fat-rich, oily seeds such as peanut, sunflower, and coconut.
All the previously mentioned, except ribosomes, are organelles composed of membranes; the following are nonmembranous organelles.
Cytosol
Most of the volume of cytoplasm is a clear substance called cytosol or hyaloplasm. It is mostly water, enzymes, and numerous chemical precursors, intermediates, and products of enzymatic reactions (TABLE 3-8). Within cytosol are free ribosomes (not attached to RER), as well as skeletal structures—the microtubules and microfilaments.
Microtubules
Microtubules (FIGURE 3-27) are the most abundant and easily studied of the structural elements of a cell, and they have many functions. They act as a “cytoskeleton,” holding certain regions of the cell surface back while other parts expand. Without them, cells would be just spheres, but by reinforcing specific areas, cell growth and expansion are directed to weaker areas. In other cases, microtubules assemble into arrays like an antenna which either catch vesicles and guide them to specific sites or cover a region, thereby excluding the vesicles (FIGURE 3-28). Finally, microtubules are the means of motility for both organelles and whole cells. The framework that moves chromosomes during division of the nucleus is composed of microtubules, and microtubules can attach to and move whole nuclei, mitochondria, and other organelles.
TABLE 3-8 Subunits of the Cell
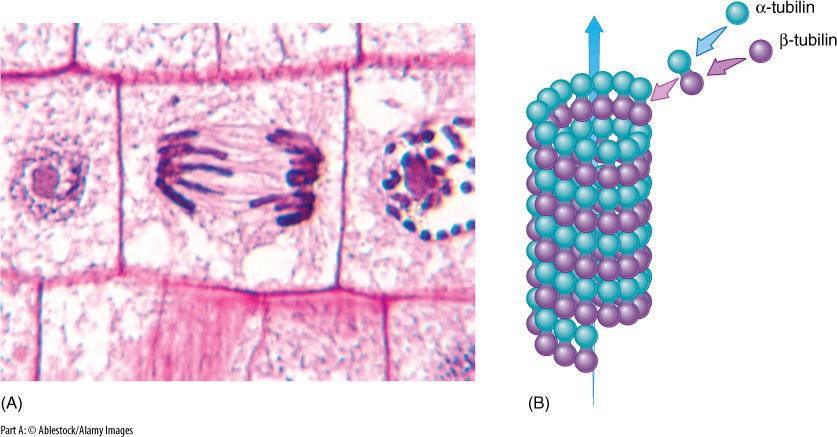
FIGURE 3-27 (A) Microtubules that are part of a dividing nucleus and are involved in pulling chromosomes to the ends of the cells. (B) Alpha- and beta-tubulin associate into a dimer called tubulin, and dimers aggregate into a microtubule. When no longer needed, microtubules depolymerize back to the monomers, which are recycled to build other microtubules.
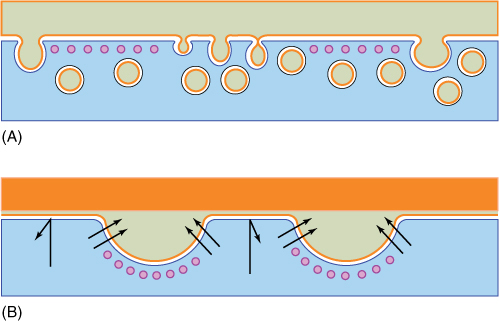
FIGURE 3-28 Microtubules (depicted here in purple) are often located next to the plasma membrane. (A) They may act as a screen that keeps vesicles away from the plasma membrane, or (B) they may pull the plasma membrane away from the cell wall, allowing material to accumulate there.
Microtubules are composed of two types of protein with a globular tertiary structure: alpha-tubulin and beta-tubulin. These associate as dimers called tubulin that further crystallize into a straight tubule with a diameter of 20 to 25 nm. This is a reversible process; when a microtubule is no longer needed, it depolymerizes back into its component monomers, which disperse into the cytosol until the cell needs to assemble a new microtubule. When microtubules occur as individuals or small clusters, new tubulin dimers are added to or removed from one end automatically. In other instances, microtubules occur in large clusters, often in a highly ordered arrangement, and a small body is usually associated with the orderly production of microtubules. For example, when the nucleus undergoes division, an array of microtubules called the spindle is formed in the middle of the cell, and spindle microtubules push and pull the chromosomes to their proper positions.
In all animals and in some fungi and algae, a pair of organelles called centrioles is associated with the formation of the spindle. A centriole is made up of nine sets of three short microtubules (FIGURE 3-29); the nine triplets are held together by fine protein spokes. Centrioles were assumed to be responsible for the organization and polymerization of the spindle microtubules even though plants never have centrioles.
Much more elaborate sets of precisely arranged microtubules occur in cilia (pronounced SILLY uh; singular, cilium) and flagella (pronounced fla GEL uh; singular, flagellum) (FIGURES 3-30 and 3-31), which appear to be identical except that cilia are short (about 2 µm) and occur in groups, whereas flagella tend to be much longer (up to several micro meters) and usually occur either singly or in sets of two or four. Both are present on many types of algae and motile fungi, but in plants, only the sperm cells have flagella. In flowering plants and conifers, no cells, not even sperm cells, ever have cilia or flagella.
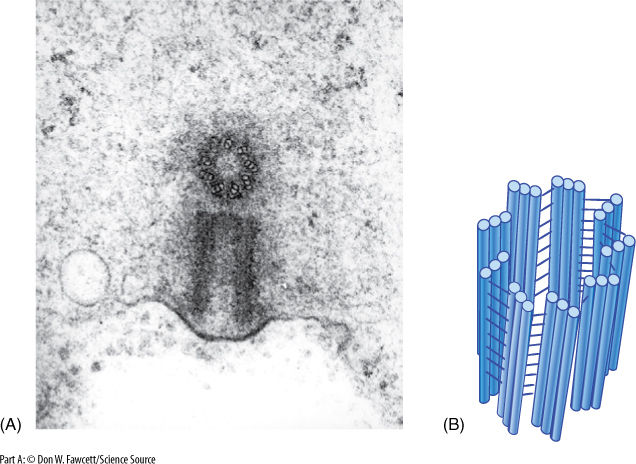
FIGURE 3-29 (A) The two members of a pair of centrioles are always located perpendicular to each other. Here, one is seen in transverse section, the other in longitudinal section (× 100,000). (B) Each part of a centriole contains a circle of nine sets of three microtubules attached to each other by fine filaments along their sides.

FIGURE 3-30 Flagella occur on many algal and fungal cells, especially the unicellular organisms. This is the alga Dunalliela. Most cells of multicellular organisms are not flagellated, but their sperm cells are (× 500).
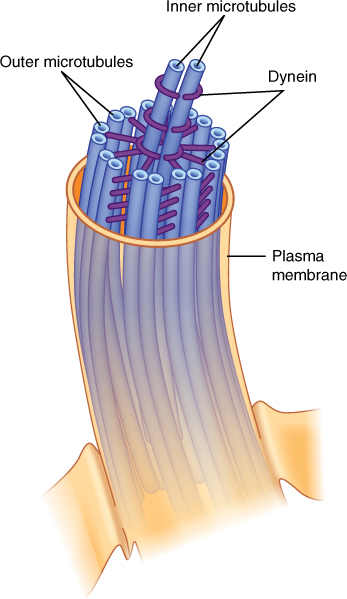
FIGURE 3-31 Cilia and flagella are composed of two central single microtubules surrounded by nine sets of two microtubules. The doublets are not merely attached to each other; they actually share tubulin monomers. The outer member of each doublet has two short arms.
In cross-section, each cilium or flagellum has a “9 + 2” arrangement: Nine pairs of fused microtubules surround two individual microtubules. The outer doublets each have two arms composed of the protein dynein, and each doublet is connected to the central pair of microtubules by protein spokes. The dynein arms convert the chemical energy of ATP into kinetic energy and bend, “walking” along the adjacent microtubule doublet. One set of doublets slides relative to the adjacent set, causing that side of the cilium or flagellum to become shorter and bend the structure. Then, as another set of microtubules slides, the cilium or flagellum bends in a different direction. The result is a powerful beating motion. If the cilia or flagella are on small organisms such as algae, fungi, or protozoans, the organism swims rapidly and gracefully.
Cilia and flagella can polymerize and depolymerize rapidly, but they never form autonomously; each is always associated with a basal body. Basal bodies appear to be identical to centrioles by electron microscopy. It was assumed that basal bodies organize the formation of flagellar microtubules, but recent studies have shown that as a flagellum grows, new monomers of tubulin and dynein are added to the tip, not the base where the basal body is located. The exact relationship between flagella and basal bodies is still not known.
Microfilaments
Like microtubules, microfilaments are constructed by the assembly of globular proteins—in this case, just one type, actin. Microfilaments are narrower than microtubules (only 3 to 6 nm in diameter), and they have been implicated in different types of structure and movement.
Storage Products
Many cells exist in an environment in which resources alternate between abundance and scarcity. To survive times of scarcity, cells must accumulate and store extra nutrients. Most often, the reserves consist of sugars that have been polymerized into starch in amyloplasts or converted into lipids and stored as large oil droplets in oily material like peanuts and sunflower seeds (FIGURE 3-32); however, with many other storage products, the function and advantages are not so obvious. Many plants store crystals of calcium oxalate or calcium carbonate (FIGURE 3-33); others accumulate large amounts of silica, tannins (FIGURE 3-34), or phenols. Because plants have no excretory mechanism, numerous waste products must be stored within the cells.
Cell Wall
Almost all plant cells have a cell wall; only sperm cells of some seed plants lack one. It is often regarded as simply an inert secretion, providing only strength and protection to the protoplasm inside; however, considerable metabolism occurs in the wall, and it should be considered a dynamic, active organelle.
The cell wall contains a considerable amount of the polysaccharide cellulose (Figure 3-5B and FIGURE 3-35A). Adjacent, parallel cellulose molecules crystallize into an extremely strong microfibril 10 to 25 nm wide. Numerous microfibrils are wound around the cell, completely covering the plasma membrane. Each cellulose polymer grows only at one end, where a complex of enzymes adds new glucose residues, one molecule at a time (Figure 3-35B). The enzymes are intrinsic proteins of the plasma membrane, and as they add new sugars to the chain, the enzymes float forward in the membrane (the chain is too heavy to be pushed backward). New cellulose molecules can be added only on the inner side of the wall, adjacent to the plasma membrane.
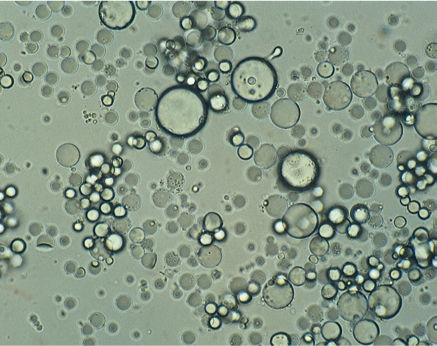
FIGURE 3-32 Avocado contains very large numbers of oil bodies; this view was prepared merely by smearing a small amount of ripe avocado fruit on a microscope slide. Virtually every visible thing is an oil body; other organelles are much less abundant (× 200).

FIGURE 3-33 Calcium oxalate crystals. When calcium is part of a crystal, it is physiologically inactive and cannot affect membranes or the tertiary structure of proteins. These are cubic crystals in wood (× 1,000).
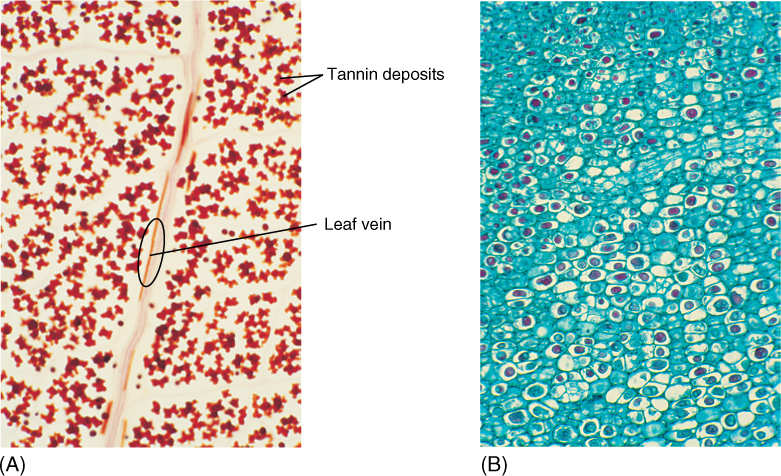
FIGURE 3-34 (A) This type of preparation is called a leaf clearing. A leaf was treated to make its tissues transparent, and then it was stained to reveal certain contents. The dark red, irregularly shaped vacuolar contents are tannins, which can denature proteins. They have a bitter taste and damage the mouth and stomach proteins of insects that try to eat them. The band running vertically through the micrograph is a leaf vein (× 200). (B) The bark of a cactus. As its cells aged, they accumulated irregular masses of tannin that have stained red (× 80).
Cellulose microfibrils are bound together by other polysaccharides called hemicelluloses, which are produced in dictyosomes and brought to the wall by dictyosome vesicles. Hemicelluloses are deposited between the cellulose microfibrils and bind to cellulose with hydrogen bonds, producing a solid structure that resembles reinforced concrete (FIGURE 3-36). In multicellular plants, the wall of one cell is glued to the walls of adjacent cells by an adhesive layer, the middle lamella, composed of a third class of polysaccharides, pectic substances.
All plant cells have a thin wall called the primary cell wall (see Figure 3-5A). In certain cells that must be unusually strong, the protoplast deposits a secondary cell wall between the primary wall and the plasma membrane (see Figure 3-5B). The secondary wall is usually much thicker than the primary wall and is almost always impregnated with the compound lignin, which makes the wall even stronger than hemicelluloses alone can make it. Lignin resists chemical, fungal, and bacterial attack. Both primary and secondary cell walls are permanent; once deposited, they are almost never degraded or depolymerized, as can be done with microtubules and microfilaments.
![]() Fungal Cells
Fungal Cells
Cells of fungi are similar to plant cells, with two important differences: (1) They do not contain plastids of any type, and (2) their walls contain chitin, not cellulose. Chitin is physically similar to cellulose, being tough, inflexible, and insoluble in water, but it contains nitrogen and is synthesized by a different mechanism than that used for cellulose. Whereas plant cells tend to be box shaped or fiber-like, fungal cells are often extremely narrow, long tubes with many, tiny nuclei.
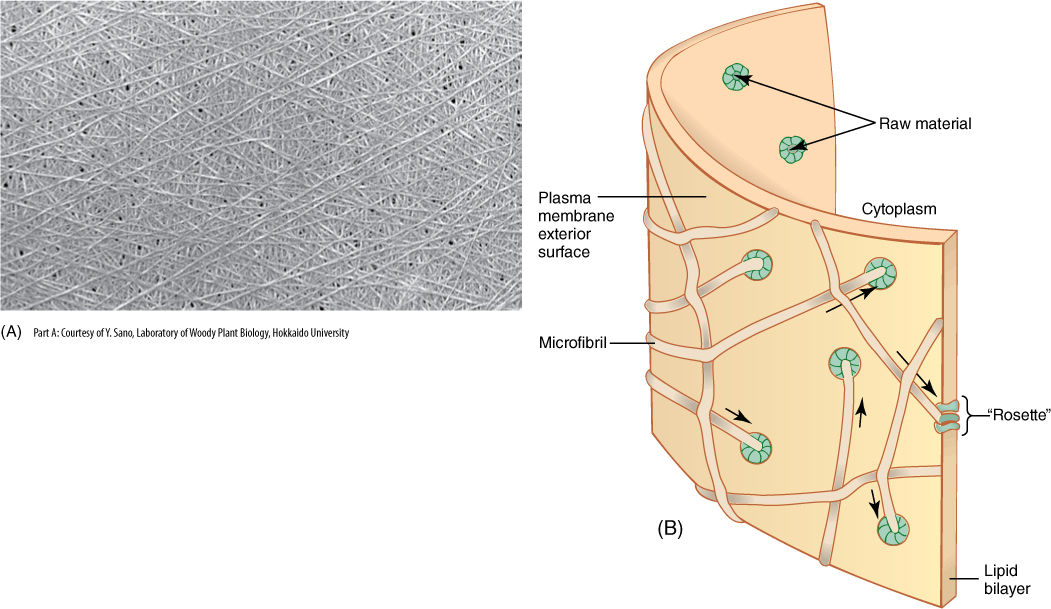
FIGURE 3-35 (A) Several layers of cellulose microfibrils are visible; other layers are present deeper in the wall. Each layer provides strength in the direction parallel to the microfibrils. Other wall components have been dissolved away to reveal the cellulose (× 7,000). (B) Cellulose-synthesizing enzymes are large proteins embedded in the plasma membrane; several proteins form a cluster called a rosette. Sugar monomers are absorbed by the enzymes of the membrane inner face and then passed across the membrane and polymerized into cellulose on the outer face. The growing microfibril extends out from the rosette and lies against microfibrils already in place. Because the microfibril cannot push into the wall, the rosette is pushed through the membrane in the direction of the arrows as the microfibril grows.
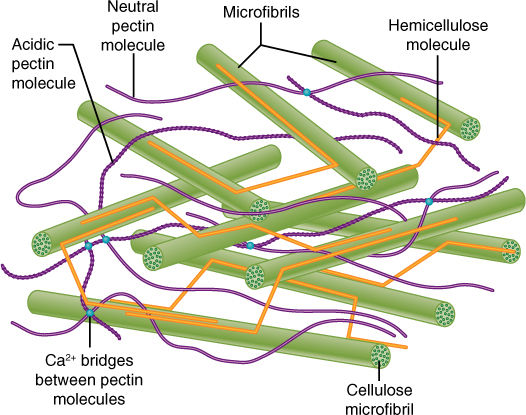
FIGURE 3-36 Although cellulose microfibrils lie close together, they do not bond with each other, and thus, a wall of pure cellulose is weak. Hemicelluloses and pectins are short, branched molecules that interact with several adjacent microfibrils, linking them together and forming a solid, three-dimensional mesh. The relative amounts of these components can vary, resulting in some walls that are flexible and others that are rigid; some strong, and others weak.
![]() Associations of Cells
Associations of Cells
In unicellular organisms, such as simple algae, protozoans, and most prokaryotes, each cell is a complete organism and does not interact directly with other cells. Such cells can communicate indirectly, at least to a small extent, by releasing chemicals that affect the other cells, but such interactions are rare, usually occurring only during the attraction of other cells during sexual reproduction.
In multicellular organisms each cell automatically, unavoidably interacts with its neighboring cells; they must share the same sources of photosynthate, oxygen, carbon dioxide, salts, and water. Whereas a unicellular organism can merely excrete its wastes across its plasma membrane, cells of a multicellular organism are not so free to do so.
Just as important are cellular interactions by which cells not only sense that they are part of a larger organism but also identify which part they are and how they should differentiate. In a developing embryo, the proper cells must be instructed to begin forming an embryonic shoot while other cells are induced to form the embryonic root, vascular tissues, seed leaves, and so on. This requires extensive, sophisticated intercellular communication.
One communication method is like that used by unicellular organisms: A cell secretes specific compounds that “inform” the surrounding cells of what it is doing metabolically and developmentally. A second method is connections between the cells. Direct physical contact between cells, as exists in animals, cannot occur in plants because the two primary walls and middle lamella are located between any two adjacent protoplasts; however, plant cells are interconnected by fine holes (plasmodesmata; singular, plasmodesma) in the walls (FIGURE 3-37). A plasmodesma is only about 40 nm in diameter, but the plasma membrane of one cell passes through it and is continuous with the plasma membrane of the adjacent cell. A small channel of cytosol also passes through, as does a short section of specialized ER.
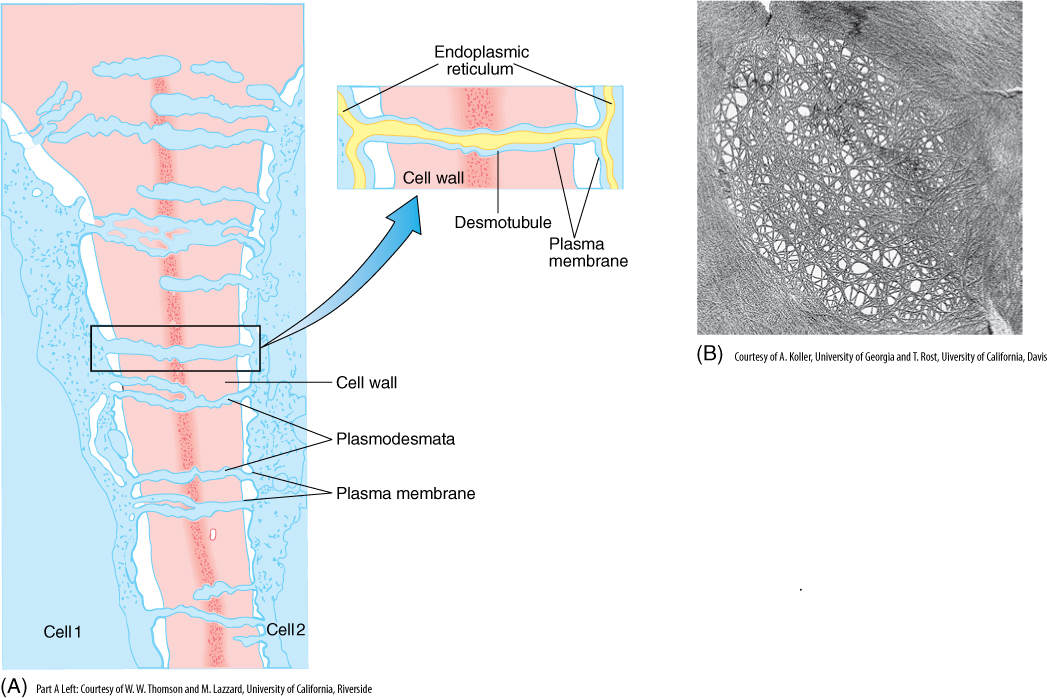
FIGURE 3-37 (A) Plasmodesmata are complexes that consist of fine holes in the primary walls; they also contain plasma membrane, liquid, and a tubule (called a desmotubule) attached to the ER (× 68,000). (B) Face view of a cell wall, showing an oval-shaped thin area, a primary pit field: The small open areas outlined by single cellulose microfibrils are holes where plasmodesmata were located. Protoplasm was removed during specimen preparation (× 45,000).
The abundance of plasmodesmata in a wall is quite variable; they can occur singly or in clusters of 10 to 20 or more (TABLE 3-9). In regions of clustered plasmodesmata, the two primary walls are often particularly thin, and the area is called a primary pit field (Figure 3-37B and 3-38). In tissues in which considerable transport occurs (into glandular or conducting cells), many primary pit fields and plasmodesmata are present, but in regions where little movement of material happens, few plasmodesmata are found.
TABLE 3-9 Frequency of Plasmodesmata
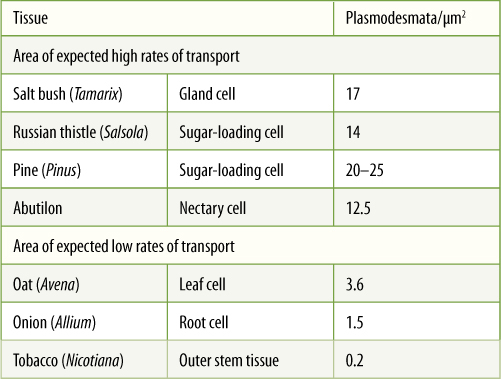
Because plasmodesmata are actually cytoplasmic channels from one protoplast to another, the individuality of the cells is diminished; all of the protoplasm within a single plant is part of one interconnected mass, the symplast. In some of the nutrient-conducting tissues (sieve tube members and companion cells) and also in some of the reproductive tissue (microspore mother cells and endosperm), plasmodesmata are so large and numerous that the cells act more like one single large cell than like individual cells. In most tissues, however, any two adjacent cells act rather independently; the communication through the plasmodesmata is not enough to prevent individuality.
The fact that walls keep protoplasts physically separated from each other has another consequence. Walls act as a second, nonliving compartment inside plants. Water moves through cell walls by capillary action, just as it moves through tissue paper. In addition, many cells do not abut each other tightly but instead have intercellular spaces between them, at least at their corners (FIGURE 3-38). In some tissues, especially leaves, cells are so loosely connected that most tissue volume is intercellular space; less than half is actually symplast. These spaces plus the cell wall constitute the apoplast; the apoplast and the symplast together make up the entire plant. The apoplast acts as a series of channels and spaces that permit the rapid diffusion of gases, which is necessary because plants do not have lungs. Diffusion through a gas-filled space is approximately 10,000 times faster than through a liquid-filled space. Without an apoplast, large, bulky plant tissues such as tree trunks, tubers, and fruits would be impossible; the interior tissues would suffocate.
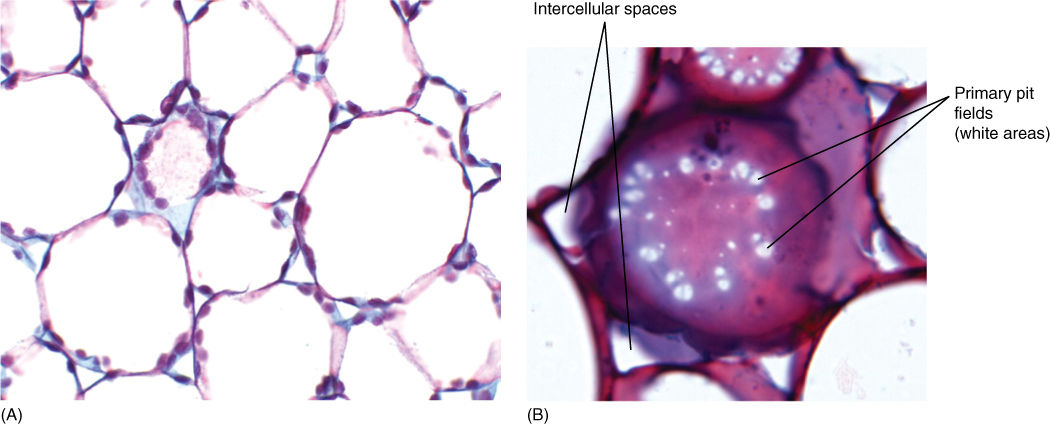
FIGURE 3-38 (A) In regions where cells press against each other, their surfaces form flat contact faces. Primary pit fields and plasmodesmata occur in contact faces. Notice that chloroplasts avoid contact faces and instead are located next to intercellular spaces, which are regions where they can obtain carbon dioxide. (× 175). (B) High magnification view of a contact face in face view. The white areas appear to be holes, but they are primary pit fields, areas where the walls are especially thin and plasmodesmata (not visible) are numerous. If this same region were viewed with an electron microscope, each small white dot would look like Figure 3-37B. The “noncontact face” is the region of cell wall that faces intercellular space rather than being pressed against an adjacent cell (× 450).
 At the Next Level
At the Next Level
1. Evolution of plastids and mitochondria. It is now relatively quick, easy, and inexpensive to sequence the DNA circles of organelles. We know the sequence and composition of every gene in the plastids of many plants, and more plastid genomes are being sequenced every week. Botanists are using this information to analyze the evolution of plastids: Algae and early plants had many more plastid genes, but in some plant groups, certain plastid genes have been transferred to the nucleus, unnecessary genes have been eliminated, and various rearrangements of genes on the plastid DNA circles are being studied. Through this we are learning how plastids and nuclear genes work together. Plant mitochondria are less well-known, but they are being studied in the same way. Remember that nuclear genes are eukaryotic, whereas plastid genes are prokaryotic, being descended from cyanobacterial genes.
2. Organization and function of plant cell cytoskeleton. Plant cytoplasm is extremely dynamic: Cytoplasm moves from place to place in the cell, as do mitochondria, plastids, and nuclei. The cytoskeleton, composed of structural and motor proteins, is involved in this, but many details are only now becoming clear. The plant cytoskeleton appears to also be involved in the ability of a plant to detect that it has touched something, for example that a tendril has touched something and should begin to wind around it or that a fly has landed on the trap of Venus flytrap.
3. History of our understanding of cells. Cells were discovered immediately after microscopes were invented in the 1660s. Two excellent books that describe the history of our knowledge about cell structure are The Birth of the Cell by Henry Harris (1999, Yale University Press) and Views of the Cell: A Pictorial History by Joseph G. Gall (1996, The American Society for Cell Biology).
SUMMARY
1. All organisms are composed of cells, the fundamental units of life. Most cells contain the same types of organelles, and there are only a few basic types of cells.
2. In multicellular organisms, cells become specialized for specific tasks, making the entire organism more efficient; however, each cell then becomes dependent on the other cells of the organism.
3. Selectively permeable membranes and active transport are critically important for life; they make compartmentalization possible, in which an enclosed compartment has a chemical composition and metabolism distinct from those outside the compartment.
4. The outermost membrane of a cell is the plasma membrane; the protoplasm it surrounds is a protoplast.
5. Plant cells contain the following membrane-bounded organelles: nucleus—heredity and control of metabolism; vacuole—temporary or long-term storage, growth; mitochondria—respiration; plastids—photosynthesis (chloroplasts); ER—intracellular transport, lipid synthesis; dictyosomes—material processing; and microbodies—detoxification or lipid metabolism.
6. Plant cells contain these nonmembranous organelles: ribosomes—protein synthesis; cytosol—the liquid matrix surrounding all other organelles; microtubules and microfilaments—skeletal structure, movement of organelles; cell wall—structure, cell shape; and various storage products such as crystals and lipid droplets.
7. Animal cells do not have a cell wall, plastids, glyoxysomes, or a central vacuole; fungal cells do have a central vacuole, but they have no plastids and their wall is made of chitin, not cellulose.
8. A plant is composed of both its protoplasm (symplast) and its intercellular spaces and walls (apoplast). The apoplast can be important for diffusion of air and liquids.
IMPORTANT TERMS
apoplast
bilayer
cell wall
centriole
chloroplasts
chromoplasts
cilia
compartmentalization
cytoplasm
cytosol
dictyosome
endocytosis
endoplasmic reticulum
eukaryotic
exocytosis
extrinsic proteins
flagella
fluid mosaic membrane
freely permeable membrane
glycoproteins
Golgi apparatus
Golgi body
impermeable membrane
intercellular spaces
intrinsic proteins
lumen
microbodies
microfilaments
microtubules
middle lamella
mitochondria
molecular pumps
nucleolus
nucleus
organelles
plasmodesmata
plastids
primary pit field
prokaryotic
ribosomes
selectively permeable membrane
symplast
tonoplast
tubulin
vacuoles
REVIEW QUESTIONS
1. In plants, each cell consists of a box-like _________________________ surrounding a mass of _______________, which in turn contains its own smaller parts, the __________________________.
2. In Table 3-1, the size of water-conducting vessel cells in oak are______________ _ _______________________. Each millimeter contains 1,000 µm. Approximately how many of these cells could be laid end to end (or side by side) in 1 mm? Do you think such a cell would be visible if it were placed in your hand? How long are the fiber cells in hemp? How long is this in millimeters? How long is it in centimeters (1 cm = 10 mm)? Could you see one of these cells if it were in your hand?
3. Figure 3-1 is a light micrograph of a leaf cell, and it is typical of what you might see if you were to examine a moss leaf. What two cell structures can you see?
4. The division of labor among various types of cells in plants is important. Examine Table 3-2, and name the cells responsible for the following tasks:
a. Collecting solar energy
b. Transporting water
c. Cell division and the production of new protoplasm
d. Water retention
e. Having pigments that attract pollinators
f. Producing poisons that inhibit animals from harming the plants
5. Stone cells in coconut shells are part of the division of labor in plants. What modifications do they have that make it impossible for them to be dividing and growing cells (hint: see Figure 3-5)?
6. All biological membranes are composed of _____________ and two layers of __________________ molecules. One of these components occurs as a thin solution only two molecules thick. A layer only two molecules thick is a _______________.
7. Some of the proteins that are part of a cell’s membranes are actually immersed in the membrane. Others are located outside the membrane and merely lie next to it. What are the names of each type?
8. Cell membranes are described as being fluid mosaics. What does that mean? Are all membrane components free to diffuse laterally anywhere within the membrane?
9. What are the two basic components of membranes? What are three ways that material can move from one side of a membrane to the other? Which method requires the plant to use energy?
10. Pieces of membranes frequently move from one area of a cell to another, but never as just flat patches, only as vesicles. Study Figure 3-9, and then draw examples of endocytosis and exocytosis using different colors for the two sides of the membranes. Notice that the black side of the membrane bilayer always touches the blue part of the cell in Figure 3-9. Can you come up with any type of vesicle fusion that would result in the black side of the membrane touching the green part of the cell?
11. Define each of these terms: a freely permeable membrane, an impermeable membrane, and a selectively permeable membrane. During your next exam, you may begin to sweat a solution of sodium, chloride, and water, but not proteins, sugars, or nucleotides. Are the membranes of your sweat gland cells freely permeable, impermeable, or selectively permeable?
12. Circle the correct answer: Plants, animals, fungi, and protists have (prokaryotic, eukaryotic) cells. What is a key feature of eukaryotic cells?
13. The _________________ __________________ (less frequently called the ____________________________)is the membrane that completely covers the surface of the protoplasm. It is (circle one: freely permeable, impermeable, selectively permeable), and it (circle one: does, does not) contain molecular pumps.
14. The nucleus serves as an archive for the organism’s _________________ _________________, all of which is stored as __ __ __ inside every nucleus.
15. Which organelle functions as storage of both nutrient reserves and waste products? What is the name of its membrane?
16. Which organelle carries out cell respiration? Each of these organelles has two membranes—a liquid portion and folded, sheet-like membrane structures. What are the names of these four components?
17. Plastids can grow and develop into many different types of structure. What is the name for the plastids that do each of the following:
a. Carry out photosynthesis
b. Synthesize lipids and other materials
c. Store starch
d. Contain abundant colored lipids
e. Occur when tissues are grown without light
18. Which organelle carries out protein synthesis?
19. What is the difference between RER and SER? SER is not very common in plants, but several examples are given in the text. What are they?
20. What is the role of dictyosomes in cell metabolism? What is their forming face? Their maturing face?
21. Draw a “typical” plant cell and include all of the organelles mentioned in this chapter. Most cells have specialized functions and increased proportions of certain organelles. How would you change your drawing if the cell is involved in photosynthesis? Is part of a yellow or orange petal? Secretes protein-rich digestive fluid? Needs a great deal of ATP? Stores starch? Has just grown a great amount?
22. Which of the organelles in Question 21 are composed primarily of membranes? What are some of the ways that the membranes participate in the metabolic activities of those organelles?
23. Describe how material is brought to a dictyosome. What happens to material while it is in a dictyosome, and how is it released? What are some of the things that might happen to it after it is released?
24. What are alpha-tubulin and beta-tubulin? When they aggregate, what do they form? What structure in dividing cells is made with this substance (Figure 3-27)? Algal, fungal, and animal cells move with structures made of this. What is the name of the structure?
25. Many animals and algae have cells with flagella, but do any plants have flagella? For example, do fern cells or moss cells have flagella?
26. How do microtubules act as a cytoskeleton? Are there times when they are particularly abundant? What are centrioles, basal bodies, and flagella?
27. Examine Figures 3-32 to 3-34. What types of substances occur as storage products in plant cells?
28. What chemical do clams use to make strong shells? What do we use for our bones? Do plants use either of these chemicals in the construction of strong cell walls? What problem would plants have if they used either the shell material or the bone material?
29. In cell walls, adjacent, parallel cellulose molecules crystallize into an extremely strong _________________________ 10 to 25 nm wide. These are bound together by polysaccharides called ______________________________, which are produced in dictyosomes.
30. What are the most abundant components of a plant cell wall? How do the components interact, and how are they arranged with respect to each other? What are the symplast and the apoplast in plants?
31. What is the type of cell wall present in all plant cells? What is the type present in certain cells that must be unusually strong?
32. What are the extremely fine holes that interconnect plant cells? What is the name of the especially thin areas of walls where these holes are particularly common (hint: Figure 3-37)?
33. What is an intercellular space? Diffusion through a gas-filled space is approximately ________________times faster than through a liquid-filled space. Living cells deep inside a bulky object such as an apple need oxygen. Do you think the oxygen could diffuse to those cells fast enough if there were no intercellular spaces? Would you guess (you will have to guess, as the answer is not in the text) that all or most of the intercellular spaces are interconnected such that oxygen could diffuse deep into an apple without ever having to dissolve into protoplasm?
Design Credits: Hummingbird: © Tongho58/Moment/Getty; Green plant cells: © ShutterStock, Inc./Nataliya Hora; Purple tulip: © ShutterStock, Inc./Marie C Fields; Dandelion: © ShutterStock, Inc./danielkreissl; Poppy: © ShutterStock, Inc./Saruri; Plant icon: © ShutterStock, Inc./Vector; Digging man icon: © ShutterStock, Inc./Z-art