Botany: An Introduction to Plant Biology - Mauseth, James D. 2017
Plant Structure
Growth and Division of the Cell

Chapter Opener Image: The bodies of both plants and animals grow as their cells and nuclei divide and multiply. Almost all cells multiply by mitosis (duplication division), such that every cell in the body has the same sets of genes as every other cell. This is true of most parts of flowers as well, but some of the cells of the stamens (yellow structures) and the carpels (red structures) multiply by meiosis (reduction division), such that the reproductive cells each have only one set of genes. Prickly poppy, Argemone.
OUTLINE
✵ Concepts
✵ Growth Phase of the Cell Cycle
- G1 Phase
- S Phase
- G2 Phase
✵ Division Phase of the Cell Cycle
- Mitosis
- Cytokinesis
- Meiosis
- Meiosis I
- Meiosis II
✵ Less Common Types of Division in Plants
✵ Cell Division in Algae
- Nuclei
- Cytokinesis
✵ Cell Division of Prokaryotes
✵ Division of Chloroplasts and Mitochondria
Box 4-1 Plants and People: Controlled Growth Versus Cancerous Growth
Box 4-2 Alternatives: Rates of Growth
Box 4-3 Botany and Beyond: Chloroplast Division During Leaf Growth
LEARNING OBJECTIVES
After reading this chapter, students will be able to:
✵ State the two phases of the cell cycle.
✵ Order and explain the three phases within interphase.
✵ List the four phases of chromatid separation during mitosis.
✵ Explain mitosis and meiosis.
✵ Order the five stages of prophase I of meiosis.
✵ Summarize the cell division process in algae.
✵ List one difference in the cell division process between prokaryotes and eukaryotes.
 Did You Know?
Did You Know?
✵ The processes of nuclear division are almost identical in plants and animals.
✵ The processes by which plant cells divide differ greatly from the processes used by animals.
✵ Cells in mature plant organs such as fully expanded leaves or open flowers typically do not divide; only cells in very immature organs divide.
✵ Mitochondria and plastids replicate their DNA and divide using processes similar to those of bacteria.
![]() Concepts
Concepts
The life cycle of individual organisms includes stages of initiation, growth, and death. Individual cells also have a life cycle, the cell cycle. Cells are initiated by division of a mother cell, grow for a period, and usually cease to exist by dividing and producing two daughter cells (FIGURE 4-1). This is not a real death because the substance of the mother cell continues to exist in the daughter cells.
As a multicellular organism grows, this type of cell cycle is common (about 25 million cell divisions occur every second in your own body), but as parts of a plant reach their final form, most cells stop dividing (cell cycle arrest) and enter an extended period of growth, during which they differentiate and mature (FIGURE 4-2). For example, leaf cells stop dividing when the leaf is only a few millimeters long; they continue to grow as the leaf expands and then function for the rest of the leaf’s life. Leaf cells cannot normally be stimulated to divide again; if the leaf is damaged, little regeneration is possible. In contrast, cortex cells in the stem or root also stop dividing before the organ is mature, but if damaged, the remaining cells resume division and remain active until the damaged portion has been protected with a layer of cork.
Some cells live for many years, even hundreds of years, but others die shortly after they mature. Bark cells are more protective if dead (FIGURE 4-3); many flower parts die only a few days after the flower first opens, and gland cells often die after a brief period of secretion.
Some cells never stop dividing. Cells in the growing points at the tips of roots and shoots are constantly cycling (FIGURES 4-4 and 4-5), as are those that form wood and bark (the cambium). When a cambial cell divides, one of the new daughters becomes a wood or bark cell, and the other remains part of the dividing layer. The wood or bark cell divides a few times and then differentiates, matures, and usually does not divide again. The daughter cell that stays in the dividing layer grows back to its original size and divides again.
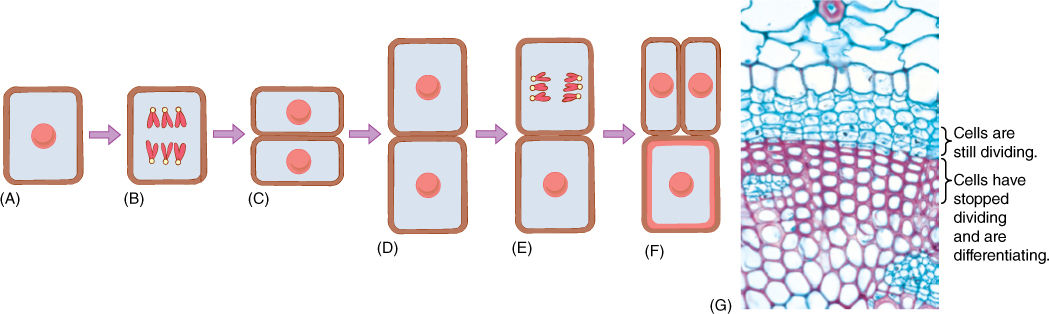
FIGURE 4-1 The cell cycle consists of division and growth phases. (A) The cell has grown and is ready for division; (B) first the nucleus divides, (C) and then the cytoplasm is divided by the formation of a new wall. (D) The new cells enter a new cell cycle by growing. (E) Then one divides again, but the other may begin to differentiate; in this example, its wall (red) thickens as it matures into a fiber. (F) The upper cell has finished dividing, and both daughter cells may enter new cell cycles. The lower cell continues differentiation; in some fiber cells the nucleus may divide once or twice, but usually once fiber differentiation begins, the nucleus never divides again. (G) A cross-section of a set of cells similar to those depicted in (A) to (F); the red cells have stopped dividing and are differentiating into fiber cells. The blue cells with thin walls are still capable of growth and cell division (× 250).

FIGURE 4-2 Palms are extreme examples of plants that have only a few dividing cells; in this entire palm shoot, cell division only occurs at (1) the extreme apex, (2) in developing leaves, and (3) in developing flowers. Of all the millions of cells in the visible trunk here, none is dividing; however, most cells are still alive. Cells at the base of the trunk were produced when the plant was just a seedling and have lived for about a century.
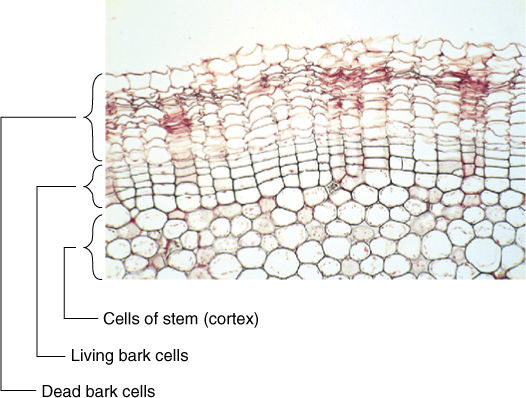
FIGURE 4-3 Cells in this geranium bark divide only a few times and then differentiate by placing antimicrobial compounds in their walls along with waterproofing compounds. The cells then die and degenerate to cell walls without protoplasts. Lack of a protoplast is selectively advantageous because if a fungus or insect does penetrate the wall, it encounters no nutritious protoplasm and starves (× 120).

FIGURE 4-4 In contrast to palms, this conifer has many groups of dividing cells. Each bud (eight are visible on just this twig) has a group of dividing cells (an apical meristem) at its apex. The needle-shaped leaves about to emerge probably formed most of their cells while enclosed in the bud and will now just undergo cell enlargement and maturation. Inside the woody brown stem is a layer of dividing cells (a vascular cambium) that will add more wood and bark cells, making this part of the stem wider but not longer.

FIGURE 4-5 The growing shoot tip of a cactus (Cleistocactus). All of its cells are alive and undergo repeated cell division. One of the basic roles of shoot tips is to convert nutrients into new cells that can later differentiate into shoot tissues. At the base, cells are lightly stained because their large vacuoles do not absorb the stain; these cells cycle more slowly than the smaller, less vacuolate cells at the tip (× 80).
The cell cycle can be divided into a growth phase and a division phase.
![]() Growth Phase of the Cell Cycle
Growth Phase of the Cell Cycle
In the 1800s, when the cell cycle was first being studied intensively, researchers gave the greatest attention to the division activities because many events could be identified. They assumed that between divisions cells were “resting,” so the growth phase was called the resting phase, or interphase (FIGURE 4-6). We now know that the cell is active during interphase, and it is possible to detect three distinct phases within interphase: G1, S, and G2.
G1 Phase
In G1 (or gap 1), the first stage after division, the cell is recovering from division and conducting most of its normal metabolism (Figure 4-6). One important process is the synthesis of nucleotides used for the next round of DNA replication. The length of a cell cycle varies tremendously, depending on the type of cell, the type of plant, its health, its age, the temperature, and many other factors. In singlecelled organisms such as some algae, the cell cycle may be as brief as several hours; short cycle times also occur in rapidly growing embryos and roots. On the other hand, cell cycle times of 2 to 3 days or even months are not unusual in tissues or plants that grow slowly. During winter dormancy, the cell cycle may last from autumn until spring; when dormancy is broken and metabolism accelerates, the cell cycle may last only a few hours. The actual length of time that any cell spends in G1 is similarly variable. Despite exceptions, in general, G1 is the longest part of the cell cycle (TABLE 4-1). After a cell undergoes cell cycle arrest, stops dividing, and begins to differentiate and mature, it may enter a state similar to G1 and remain in it for life.
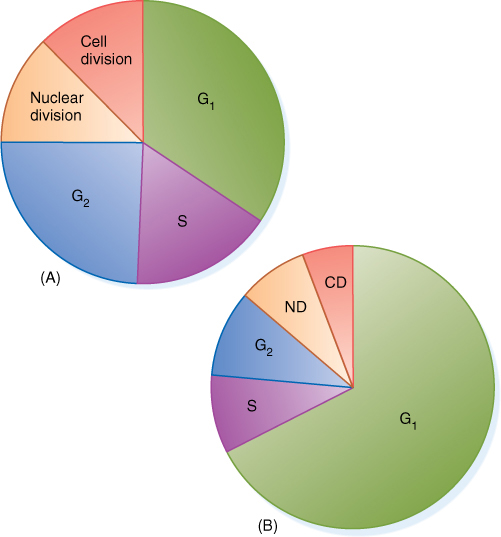
FIGURE 4-6 Comparison of cell cycles of different tissues or species. In a shoot tip (A), cell division may occur as soon as the cell grows to its proper size, and G1 is only a small part of the cell cycle. Below the shoot tip (B), cells still divide but spend more of their time growing and carrying out nondivision metabolism; G1 is a greater proportion of the cell cycle. Both (A) and (B) give only the relative amount of time spent in each phase, not the absolute time in hours or days. The cell cycle in cells like (B) is often much longer than in shoot tips (A).
S Phase
During S (or synthesis) phase, the genes in the nucleus are replicated. A gene is a polymer of nucleotides, and each gene has a unique sequence of nucleotides. Although many genes may have similar sizes, different genes never have exactly the same sequence of nucleotides. Many higher plants and animals need about 30,000 types of genes to store the information required to make the proper enzymes, structural proteins, and hormones necessary for the organism’s life. The entire complex of genes for an organism is its genome.
Obviously, if each individual gene floated around the cell or nucleus as an independent piece of DNA, then during division, it would be extraordinarily difficult to find each of those 60,000 genes (30,000 originals and 30,000 copies) and make certain that each daughter cell received one of each (FIGURE 4-7). This is not a problem, however, because all genes are attached to other genes by short pieces of linking DNA. Thousands of genes are attached together in a linear sequence; the entire structure is a chromosome (FIGURE 4-8). It might seem reasonable for all genes to link together in a single chromosome, but such a long chromosome would probably become hopelessly tangled. Only a few plants have as few as 2 chromosomes for their genome; most have between 5 and 30 chromosomes (TABLE 4-2).
TABLE 4-1 Cell Cycle Phase Durations in Root Tip Cells and Cell Cultures
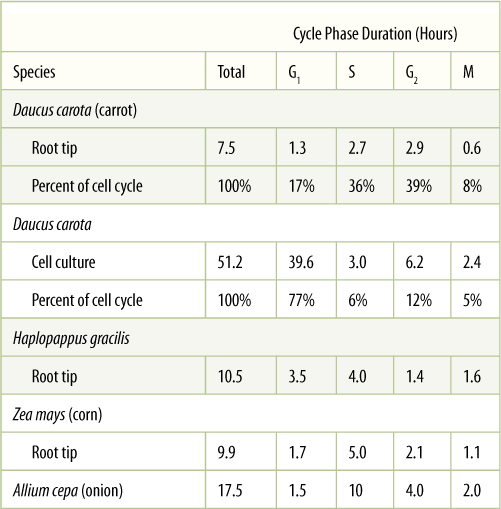
Note that when carrot cells are grown in cell culture, the cell cycle becomes much longer, mostly because of G1, which goes from being only 17% of the cell cycle in roots to 77% in cell culture.
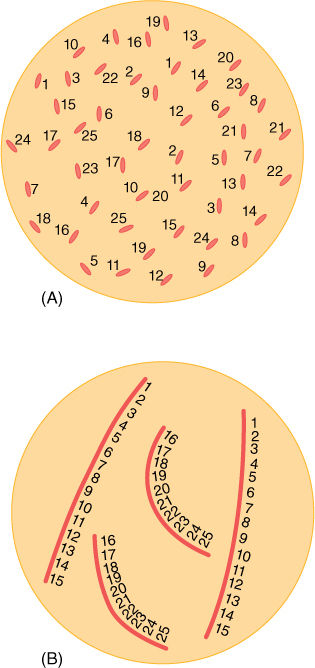
FIGURE 4-7 (A) If each gene were a distinct piece of DNA, ensuring replication of each would be difficult, and ensuring that each daughter nucleus received one copy of each would be even more difficult. (B) Grouping the genes into chromosomes makes them easier to manipulate. Here only two types of chromosomes are shown: long ones with genes 1 to 15 and short ones with genes 16 to 25.
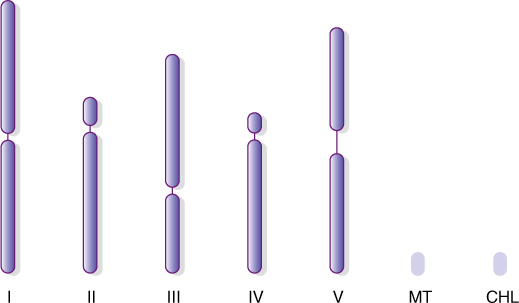
FIGURE 4-8 Each nucleus in the cells of Arabidopsis thaliana has two of each of these five types of chromosomes, one set inherited from the paternal and one from the maternal parent. The slender area in each represents the centromere. The lines labeled MT and CHL represent the DNA circles of mitochondria and plastids. The entire genome has been sequenced: We know which DNA nucleotide occurs at each site in each chromosome.
Even with 20 different chromosomes, an organism with 30,000 genes would have an average of 1,500 genes on each chromosome, resulting in long pieces of DNA that might break if unprotected. In onion, the DNA in each nucleus is 10.5 meters long when all DNA molecules are measured; in lilies, it is 21.8 meters long. In eukaryotes, a special class of proteins called histones complexes with DNA and gives it both protection and structure (FIGURE 4-9). Chromosomes also have another structural feature, the centromere, which is usually located near the center of the chromosome (FIGURE 4-10). Each end is capped by a telomere.
TABLE 4-2 Number of Chromosomes in a Haploid Set
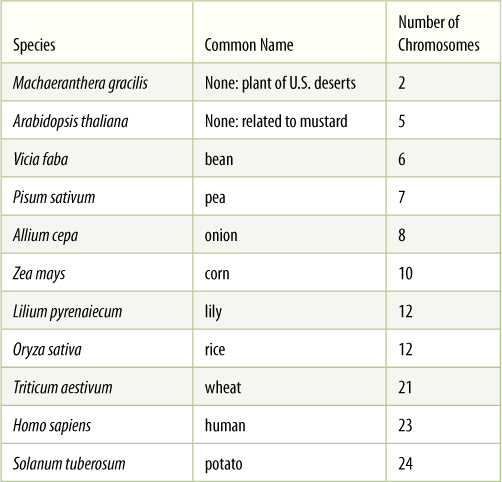
A haploid set is a single set of chromosomes, as carried by sperm cells and egg cells. A fertilized egg has two sets and is diploid.
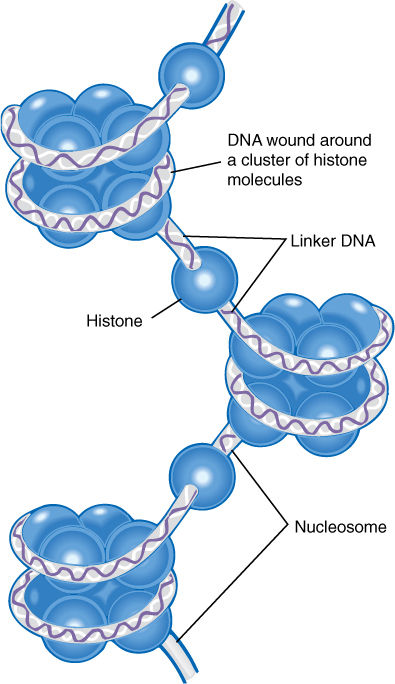
FIGURE 4-9 Histone proteins associate into short cylinders and then DNA winds around each cylinder. The DNA-histone particles then associate into a compact arrangement.
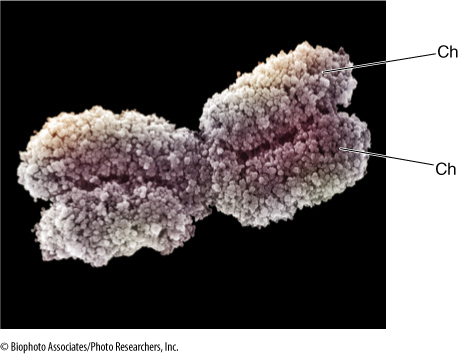
FIGURE 4-10 This chromosome was taken from a cell during division. It had gone through S phase of the cell cycle, so it has two chromatids (Ch), not just one. The chromatin has coiled tightly (condensed) and is visible as loops. Condensation has made the chromosome short and thick, and it can be moved around the cell easily. The pinched region in the center is the centromere.
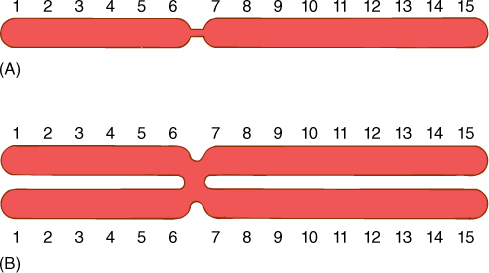
FIGURE 4-11 (A) Before S phase, each chromosome has one chromatid and one copy of each gene. (B) After replication in S phase, each chromosome has two chromatids and two copies of each gene. The constriction represents the centromere.
During S phase, linking pieces of DNA as well as genes are replicated, and new histone molecules complex with the new DNA. Thus, entire chromosomes, not just DNA, are replicated (FIGURE 4-11). After replication is finished, the duplicate DNA molecules remain attached at the centromere, connected by a protein called cohesin. Although it would be justifiable to call this a “double chromosome” or a “pair of chromosomes,” it is also called a chromosome. Now that we know how a chromosome changes during S phase, we call each half of the doubled chromosome a chromatid. After S phase, each chromosome has two identical chromatids; before S phase, each chromosome has just one chromatid. A chromosome after S phase is twice as large as it was before S phase.
Although many cells stop in G1 when they cease dividing and begin to mature, most plant cells enter S phase and replicate their DNA before they begin to differentiate. This may involve just a single cycle of DNA replication, resulting in a nucleus twice as large as would be expected, or it may continue for many rounds of DNA synthesis and the nucleus becomes gigantic (FIGURE 4-12). This process is endoreduplication, estimated to occur in 80% of all maturing plant cells (TABLE 4-3). The resulting nucleus has many copies of each gene—as many as 8,192 copies in some cells of kidney beans (Phaseolus vulgaris). Endoreduplication occurs most often in hairs, glandular cells, and other cells that must have an extremely rapid, intense metabolism. The normal complement of two copies of each gene does not seem to make messenger RNA rapidly enough for such active cells. Such cells are also rich in ribosomes and produce large amounts of protein.
Gene amplification is similar to endoreduplication, but only some genes are repeatedly replicated. The amplified genes are those needed for the specialized metabolism of the mature cell (FIGURE 4-13). For example, as a protein-rich seed develops, its cells need large quantities of the messenger RNA that codes for the storage protein, and those genes are amplified; however, it would be a waste of energy and resources to replicate all of the other genes as well. The most extreme case of gene amplification known is in Arum maculatum (a relative of Philodendron): Each nucleus has enough DNA for 24,576 copies of every gene. We do not know how many genes are being amplified and how many are present as only two copies per nucleus, and thus, we cannot compute the actual number of copies of each amplified gene; however, it is certainly more than 24,576.
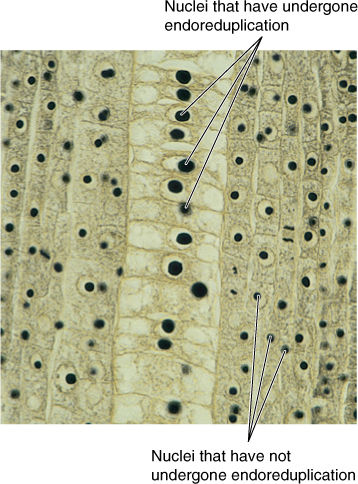
FIGURE 4-12 In this root of Scirpus (bulrush, sedge), cells in the central column have enormous nuclei that have undergone endoreduplication. Each nucleus now has many copies of each gene so they can produce messenger RNA very rapidly. Consequently, the cytoplasmic ribosomes produce protein quickly as well (× 200).
TABLE 4-3 Examples of Endoreduplication and Gene Amplification
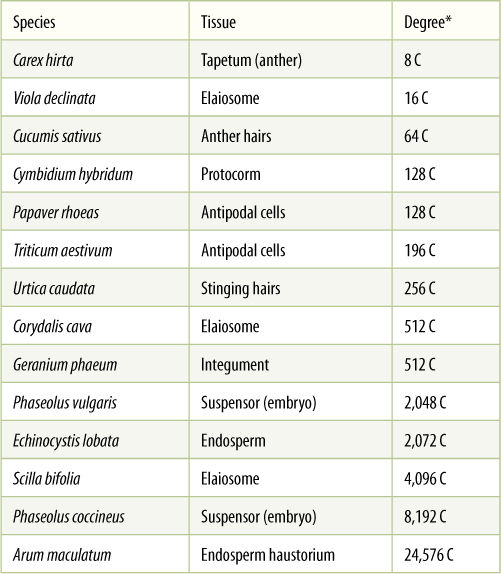
* “C” is a symbol that represents the amount of DNA present in a nucleus; a normal body cell has 2 C before S phase and 4 C after S phase. Values that are a power of 2 represent endoreduplication; those that are not represent gene amplification.
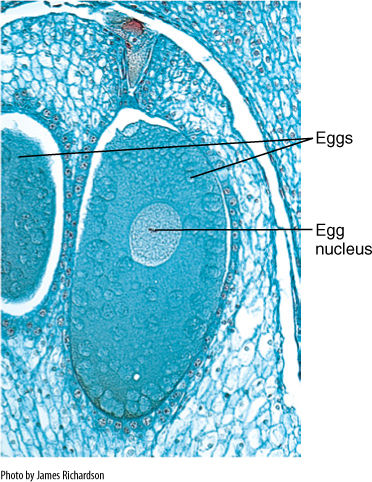
FIGURE 4-13 Egg cells of pines and other conifers become enormous; their nuclei are gigantic due to having large amounts of RNA. Like any egg cell nucleus, this must fuse with a sperm cell nucleus before it can divide (× 20).
G2 Phase
After S phase, the cell progresses into G2 phase (G2), during which cells prepare for division. This phase lasts only about 3 to 5 hours. The α- and β-tubulins necessary for spindle microtubules are synthesized, and the cell produces proteins necessary for processing chromosomes and breaking down the nuclear envelope. In cultured animal tissue, if a cell whose nucleus is dividing is forced to fuse with one in G1, the second cell’s nucleus also begins the first steps of nuclear division. This is evidence that during G2 the first cell produced factors necessary to start nuclear division and that these factors are located in the cytoplasm.
G1, S, and G2 constitute the interphase portion of the cell cycle. After G2, division can occur.
![]() Division Phase of the Cell Cycle
Division Phase of the Cell Cycle
The actual division involves two processes: (1) division of the nucleus, called karyokinesis, and (2) division of the cytoplasm, called cytokinesis. There are two types of karyokinesis: mitosis and meiosis.
Mitosis
Mitosis is duplication division. It is the more common type of karyokinesis, the method any multicellular organism uses as its body grows and the number of its cells increases. It is also used by eukaryotic unicellular organisms when they are not undergoing sexual reproduction. Mitosis is called duplication division because the nuclear genes are first copied; then one set of genes is separated from the other, and each is packed into its own nucleus (FIGURE 4-14). Each daughter nucleus is basically a duplicate of the original mother nucleus and a twin of the other. Mitosis produces nuclei that are more or less exact copies of the original nucleus, except for occasional errors.
As mentioned, thousands of genes are linked together into just a few chromosomes, making separation of gene sets much easier: It is necessary to transport only one of each type of chromosome to each daughter nucleus. This is made even easier because the two new chromatids remain attached together by their shared centromere as the chromosome is replicated in S phase. It is necessary only to make certain that one half of the doubled, large chromosome goes to one end of the cell and the other half to the other end. If that happens with each chromosome, each end of the cell automatically receives one full set of genes. The mechanism that ensures this orderly separation of chromatids consists of the following four phases:
Prophase
During interphase, the DNA of a chromosome exists as a long, extended double helix associated with histone protein (Figure 4-9). This open configuration allows enzyme complexes to find specific genes that must be read for the information they contain, but in this condition, a chromosome may be several centimeters long. It could be wrapped around the cell hundreds of times, making it impossible to pull one chromatid from the other. During prophase, however, the chromosomes condense—they undergo a change in the way the histones associate. Chromosomes begin to coil repeatedly (Figure 4-10), becoming shorter and thicker each time. In addition, a protein framework develops to which DNA binds. Chromosome condensation continues until chromosomes are only 2 to 5 μm long; in this form, they can be moved around the cell more easily. In this condensed form, we can actually see the chromosomes; the uncoiled molecules of interphase are too narrow to be resolved by light microscopy.
As chromosomes condense and become visible during prophase, other events occur as well (Figure 4-14). The nucleolus becomes less distinct and usually completely disappears by the end of prophase. The nuclear envelope breaks into numerous vesicles, each attached to rough endoplasmic reticulum. In those organisms that have a set of centrioles (mostly algae, fungi, and animals), the daughter sets that were duplicated during the previous interphase now migrate to opposite poles (sides) of the cell. It had been thought that they simultaneously acted as microtubule-organizing centers and produced a long set of microtubules, the spindle, between themselves, but even if centrioles are destroyed by laser microbeams in animals, the spindle still forms. It could be that the centrioles themselves are separated by spindle formation, and the association is a mechanism that ensures that each daughter cell receives a centriole.
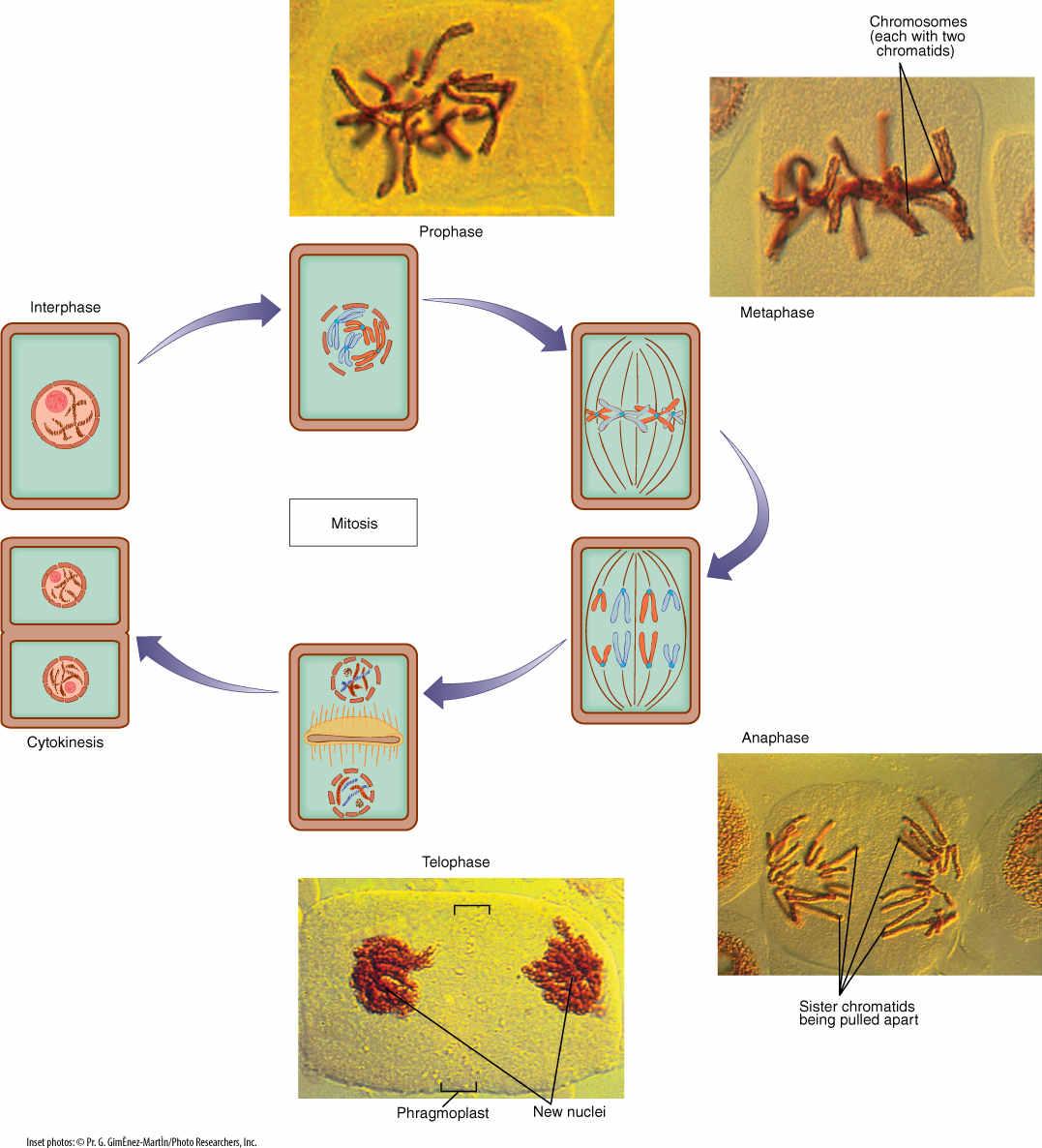
FIGURE 4-14 During mitotic nuclear division, one chromatid from each chromosome is pulled to one end of the cell, and the other chromatid of each chromosome goes to the other end. Details are given in the text.
The spindle is composed of hundreds or thousands of microtubules. Some microtubules extend from one pole to the center of the cell where their ends overlap the ends of other microtubules that extend from the opposite pole. The two sets together, overlapping in the center, form a large framework (FIGURES 4-15 and 4-16). Other microtubules run from a pole to a centromere. The point of attachment is a kinetochore; a structure consisting of two layers of proteins, one layer bound tightly to centromere DNA and the other attached to spindle microtubules. Each centromere has two kinetochore faces, one attached by approximately 15 to 35 microtubules to one end of the spindle and the other face attached by a similar number to the other end of the spindle (Figure 4-15).
Plants and People
BOX 4-1 Controlled Growth Versus Cancerous Growth
The actual steps of karyokinesis and cytokinesis must be controlled if cells are to divide properly, but other aspects must be regulated as well. First, the rate and frequency of cell division are important in determining how rapidly or slowly one cell produces a mass of progeny cells. Second, orientation of both cell division and cell growth affects the shape of the growing mass of cells: If all cells divide with their new walls parallel to each other, the mass grows as a column, but if new cell walls occur in any plane, the mass grows in all three dimensions. The mass grows as a sheet if new cell walls can occur in two planes but not in the third. Finally, it is important to control which cells divide: If only some cells undergo cell division, they may produce a lump or outgrowth while the rest of the mass of cells remains unchanged.
These factors are controlled accurately in plants. In a young embryonic plant, all cells divide but later, cells at the tips of roots and shoots become centers of cell division and growth while the rest of the stem and root tissues mature and carry out their functions. By controlling rate, orientation, and location of cell division, plants produce cylindrical stems and roots, thin flat leaves and petals, and massive, three-dimensional fruits.
Plants impose quiescence on certain cells and then reactivate them to growth and division later: Plants produce buds that are forced to remain quiescent for a long time, even years, but can then be stimulated by the plant to grow out as a branch or a flower. In young stems, epidermal cells grow rapidly enough to keep the stem covered, but then they mature and remain mitotically quiescent as they protect the plant. In many species, these can be reactivated years later, undergo cell division, and produce bark cells.
Cell division and growth must be controlled in animals as well. During early stages of fetal development, all cells of human embryos divide and grow. Later, some cells divide more rapidly than others, but basically, most of our cells are mitotically active during much of our growth before birth. Then cells in certain tissues and organs undergo cell cycle arrest; they mature and never divide again such as the cells of our eyes and our brains. Other cells never stop dividing and are active until we die, such as the layer of cells that produces our skin and hair and the bone marrow that generates most of our blood cells. Just as in plants, some of our cells enter a prolonged state of quiescence and later are activated to division. For example, surgical removal of part of the liver causes cells of the remaining portion to divide and restore the organ to an adequate size. Of course, this is not true of most of our organs.
Some of our cells release themselves from cell cycle arrest and begin growing uncontrolled by the rest of the body. This is cancerous growth, and its severity depends on which types of cells and organs are involved, how rapidly the cells divide, and whether the cells can migrate from their original site and invade surrounding tissues. It is well known that certain environmental factors act as carcinogens—agents that cause cancer by interfering with cell cycle arrest. Cigarette smoke is known to cause cancer of the lung and throat, and ultraviolet light triggers skin cancer.
Whereas uncontrolled cancerous growth in humans may be fatal, it does not seem to be a problem in plants. Irregular lumps and growths, called galls, may occur, but these are often caused by insects or microbes, not by the plant’s own cells undergoing a spontaneous, self-induced release from cell cycle arrest. It may be that plants do form cancerous growths but that they are not a serious problem for several reasons. First, cells cannot migrate through a plant body the way that our cells migrate through our bodies; consequently, any uncontrolled growth is localized, not invasive. Second, whereas we have many organs that are each critical to our life and which occur singly (heart, brain) or in pairs (kidneys, lungs), plants have many leaves, roots, and flowers, and no single one is indispensable. Damage to one part of the plant may have little effect on the rest of the plant. By lacking the highly differentiated, complex, and tightly integrated body of humans and many animals, plants are not so threatened by diseases involving control of nuclear and cellular division.
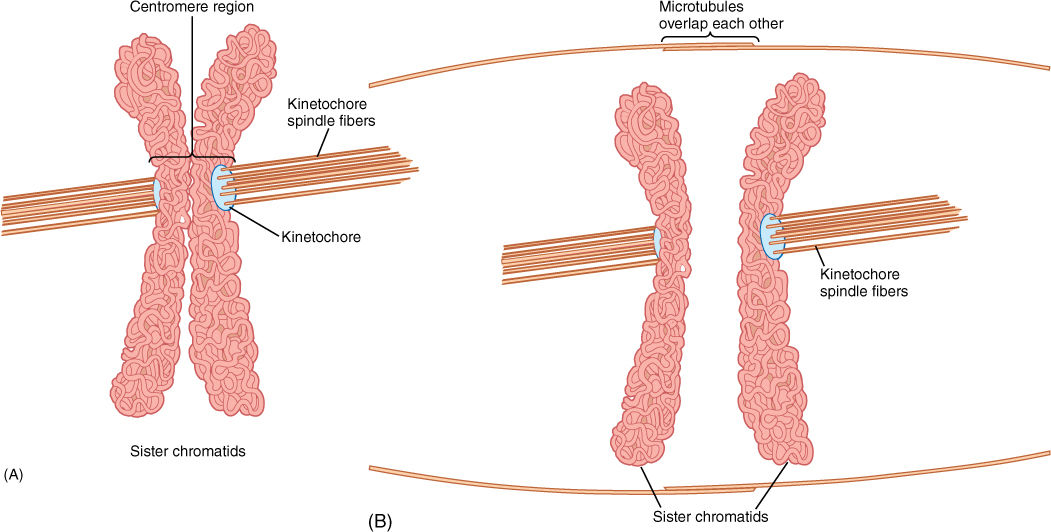
FIGURE 4-15 (A) Early in mitosis, spindle microtubules attach to chromosomes at the kinetochore of the centromere. Each chromatid is attached only to microtubules coming from one end of the cell. (B) Other microtubules pass from pole to pole or overlap the ends of other microtubules at the center of the spindle. As the kinetochore-attached microtubules shorten, the two chromatids of each chromosome are pulled in opposite directions. The long microtubules that do not attach to chromatids act as a framework such that chromatids are pulled apart rather than having the spindle poles pulled into the center of the cell.
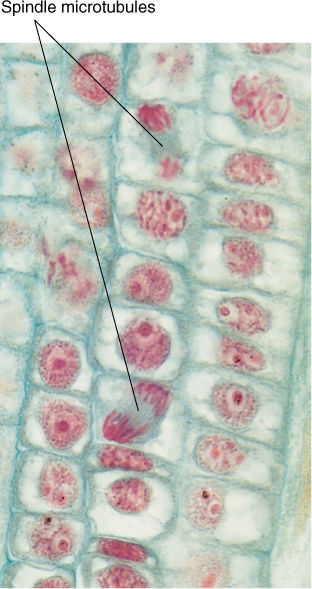
FIGURE 4-16 Anaphase in hyacinth root tips. The gray material between the red chromosomes consists of masses of spindle microtubules. These roots grow quickly and cell cycle times are short, and thus, at any particular time, many cells are in mitosis rather than just in interphase (× 250).
Metaphase
After spindle microtubules attach to centromeres, they push and pull on the chromosomes and gradually move them to the cell center; their arrangement there is called a metaphase plate (Figure 4-14). Viewed from the side, the chromosomes appear to be aligned in the very center, but viewed from the end, they are seen to be distributed throughout the central plane of the cell.
No distinct boundary occurs between prophase and metaphase; chromosomes gradually become visible and gradually move to the metaphase plate. Fragments of nucleolus and nuclear envelope may persist well into metaphase; however, a distinct boundary is evident between metaphase and the following anaphase. At the end of metaphase, a protein-degrading enzyme called separase is activated, and it digests cohesin: The two chromatids of each chromosome are suddenly free of each other. In this step, the number of chromosomes is doubled, but the size of each chromosome is halved. Each chromosome is like it was in G1, before S phase. Although the number of chromosomes in the cell is suddenly doubled, the doubling is transitory because the cell soon divides.
Anaphase
Anaphase begins just after cohesin releases the centromeres, freeing the two kinetochore faces from each other. Spindle microtubules that run to the kinetochores shorten, depolymerizing at the end near the spindle pole, pulling each daughter chromosome away from its twin. Agents that inhibit microtubule depolymerization, such as deuterium oxide, also inhibit chromosome movement, whereas those that speed depolymerization, such as low levels of colchicine, speed movement. The energy necessary to move a chromosome from the metaphase plate to the end of the spindle is small: just 20 ATP molecules. Long chromosomes may tangle somewhat, but microtubules exert sufficient pull to untangle them and drag them to the ends of the spindle. Because the spindle is shaped like a football, as chromosomes on each side get closer to the end, they are pulled together into a compact space.
Telophase
As chromosomes approach the ends of the spindle, fragments of nuclear envelope appear near them, connect with each other, and form complete nuclear envelopes at each end of the cell. The total surface area of the two new nuclei is larger than that of the envelope of the original mother nucleus; extra membrane may be derived from endoplasmic reticulum (ER). Chromosomes become less distinct because they start to uncoil. Gradually, new nucleoli appear as the ribosomal genes become active and produce ribosome subunits. The spindle depolymerizes completely and disappears. Most events in telophase are reversals of those in prophase.
To summarize mitosis: After G2 is completed at the end of interphase, each chromosome has been replicated and consists of twin chromatids. Spindle microtubules from opposite poles attach to the centromeres then pull the twin chromatids of each chromosome away from each other. Two new nuclei form, each containing a full set of chromosomes and each chromosome having one chromatid. The new nuclei are identical to each other and to the original nucleus that began mitosis. The nuclei can then enter G1 of interphase. The steps of mitosis (prophase, metaphase, anaphase, and telophase) are part of a continuous process and intergrade with each other. Similarly, prophase and telophase intergrade with interphase.
Cytokinesis
The division of the protoplast is much simpler than the division of the nucleus. Although it is necessary for each daughter cell to receive some of each type of organelle, random distribution of organelles in the mother cell usually ensures this. No matter how the cell divides, each half typically contains some mitochondria, some plastids, some ER, some vacuoles, and so on. It is not necessary for each daughter cell to get exactly half of each. A single mitochondrion can divide, or a fragment of ER can grow until the cell has an adequate amount. The same is not true for genes: If one daughter cell is missing a gene or chromosome, the other genes cannot regenerate the missing information.
During prophase, a set of microtubules and actin filaments aggregates into a band running around the cell, just interior to the plasma membrane. This preprophase band identifies the plane of division, and it marks the region where the new cell wall will attach to the existing wall. If the preprophase band forms around the middle of a long cell, the cell will divide into two equal-sized cells, but if it forms at one end, then division will produce a large and a small cell. The preprophase band is transitory, and its microtubules quickly disassemble and are recycled into the mitotic spindle. If nuclear division occurs without cell division, a preprophase band is not formed.
Cytokinesis in plants involves formation of a phragmoplast, a set of short microtubules aligned parallel to the spindle microtubules. Actin filaments are also present. The phragmoplast forms in the cell center where the metaphase plate had been (FIGURES 4-17 and 4-18). Phragmoplast microtubules trap dictyosome vesicles that then fuse into a large, flat, plate-like vesicle in which two new primary walls and a middle lamella begin to form. The phragmoplast grows outward toward the walls of the original cell. It may take half a day to reach the walls of large cells. Phragmoplast microtubules do not actually migrate toward the walls. Instead, new microtubules polymerize near the outer edge of the phragmoplast, whereas those on the inner side depolymerize. New dictyosome vesicles are trapped and added to the edge of the large vesicle; it too grows outward, following the phragmoplast. Similarly, the new walls extend outward along their edges. The phragmoplast, vesicle, and walls are called the cell plate. This process continues until the large vesicle meets the mother cell’s plasma membrane, the two fuse, and the vesicle membrane becomes a part of the plasma membranes of the two daughter cells. Simultaneously, the new walls meet and fuse with the wall of the mother cell, completing the division of the mother cell into two daughter cells.
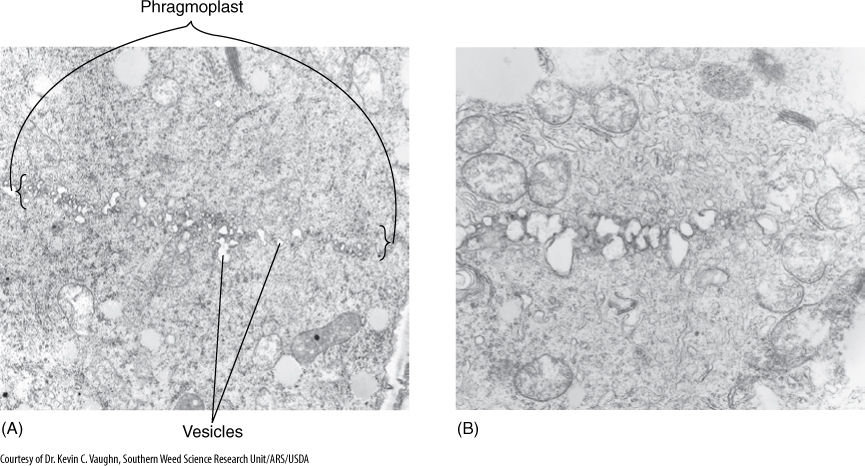
FIGURE 4-17 (A) Late telophase in onion. The row of small vesicles is the beginning of a cell plate. At the edges are short segments of microtubules, the phragmoplast. (B) Cell division has been interrupted by the herbicide DCPA (dimethyl tetrachloroterephalate) (both × 20,000).
Alternatives
BOX 4-2 Rates of Growth
An organism, or a part of an organism, can produce more cells in a variety of ways. The two ways that are most important for you to understand are called arithmetic and geometric increase.
In arithmetic increase, only one cell is allowed to divide. Of the two resulting progeny cells, one continues to divide, but the other undergoes cell cycle arrest and begins to develop, differentiate, and mature. After each round of cell division, only a single cell remains capable of division, and one new body cell exists. For example, starting with a single cell, after round 1 of cell division, there is one dividing cell and one body cell. After round 2, there are two body cells, and after round 3, there are three, and so on. To obtain a plant body containing 1 million cells (which would be a very small body), the plant’s single dividing cell would undergo 1 million rounds of nuclear and cellular division. If each round requires 1 day, arithmetic increase would require 1 million days, or 2,739.7 years. As you can imagine, arithmetic increase is too slow to ever produce an entire plant or animal; however, it is capable of producing the small number of cells present in very small parts of plants. For example, the hairs on many leaves and stems consist of just a single row of cells produced by the division of the basal cell, the cell at the bottom of the hair next to the other epidermal cells. The hair may contain 5 to 10 cells, and thus, all of its cells could be produced in just 5 to 10 days by divisions of the basal cell; this would be quick compared with the rest of leaf development, which might take weeks or months.
An alternative pattern of growth by cell division, geometric increase, results if all cells of the organism or tissue are active mitotically. Again starting with a single cell, after round 1 of cell division, there are two cells capable of cell division. After round 2, there are four, and after round 3, there are eight, and so on. The number of cells increases extremely rapidly and can be calculated as a power of 2. After round 3, there are 23 = 8 cells, and after round 10, there are 210 = 1,024 cells (these calculations should be easy for you on any pocket calculator). After round 20, there are already 220 = 1,048,576 cells. If, as in our previous example, each round of cell division requires 1 day, it now takes the plant only 20 days, not thousands of years, to reach a size of 1 million cells. With geometric increase, a large plant or animal body can be produced quickly.
In fact, geometric growth is common in animals but rarely occurs in plants except when they are extremely young and small. The problem is that in plants, dividing cells are typically simple and do not carry out many of the specialized metabolic processes necessary for plant survival. They cannot be the tough, hard fiber cells that make wood strong enough to hold up a tree; they cannot be the waxy, waterproofing cells of the plant’s skin that protect it from fungi, bacteria, and loss of water.
Plants grow by a combination of arithmetic and geometric increase. A young, embryonic plant grows geometrically and rapidly, with all of its cells dividing. Then cell division becomes restricted to a small group of cells at the tips of roots and shoots. After this point, growth is of the slower arithmetic type: Half of the new cells produced in each round of division remain capable of division, and the other half of the new cells develop into their mature condition and begin carrying out specialized types of metabolism. Plants are thus a mixture of older, mature cells and young, dividing cells.
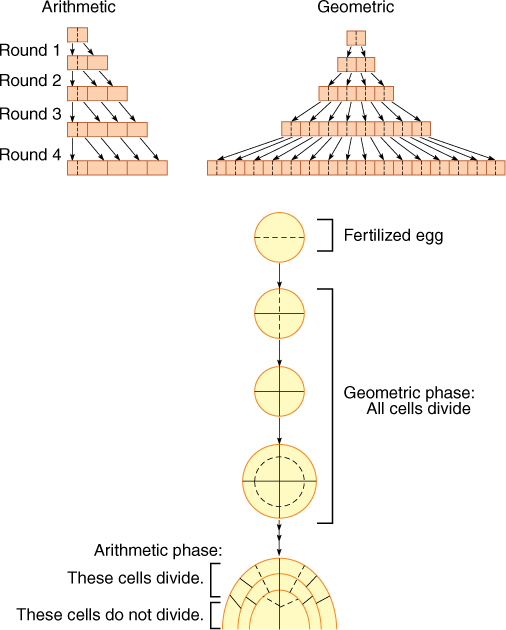
FIGURE B4-1 Possible types of cell division.
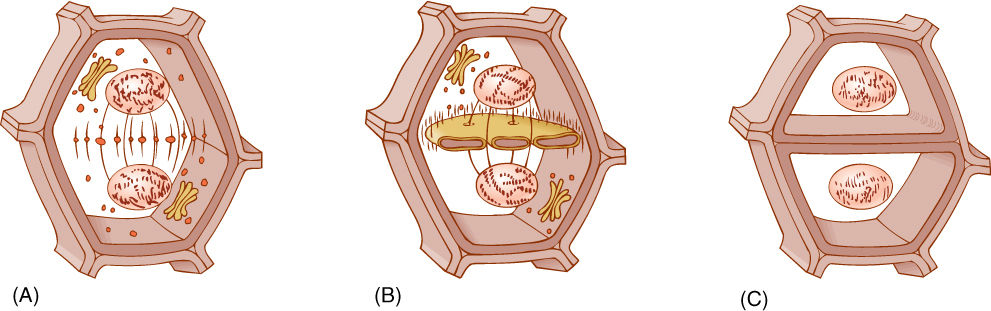
FIGURE 4-18 (A) Cytokinesis begins as dictyosome vesicles are trapped by phragmoplast microtubules in the space between the two new nuclei. (B) The small vesicles fuse into one large vesicle in which the new middle lamella and two primary walls will form. Plasmodesmata form at this time. (C) The cell plate enlarges toward the existing cell wall as more dictyosome vesicles fuse with the edges of the cell plate vesicle. When the cell plate reaches the existing cell walls, the vesicle membrane fuses with the plasma membrane and thus becomes plasma membrane itself. The new cell plate abuts the old cell wall, and the two become glued together with hemicelluloses and pectins.
Many meristematic cells are small and cytoplasmic with only a few small vacuoles, but some highly vacuolate cells also divide, for example, vascular cambium cells and large cortex cells forming scar tissue over a wound. In these, the vacuole itself must be divided before either the nucleus or the cell can divide. Vacuole division is accomplished with a phragmosome, a set of microtubules, actin filaments, and cytoplasm. First, the microtubules and actin filaments pull the nucleus to the site where it will divide, either along the edge of the cell or often suspended in a tube of cytoplasm that extends through the vacuole. The phragmosome then enlarges as a thin plate-like sheet of cytoplasm and cytoskeleton, extending outward from the vicinity of the nucleus and pushing into the vacuole, dividing it in two. As the phragmosome advances through the vacuole, the nucleus divides, the phragmoplast and cell plate form, and then they too extend outward through the thin sheet of cytoplasm established by the phragmosome. Just after the phragmosome reaches the far side of the vacuole, the phragmoplast and cell plate pass through and join with the plasma membrane and cell wall; the cell has been divided.
Meiosis
In mitosis, daughter nuclei are replicates of the original mother nucleus. This is necessary for the growth of an organism but creates a problem when sexual reproduction occurs. Two sex cells (gametes) fuse together, forming a zygote, which then grows into a new adult. Each gamete contains one complete set of chromosomes (FIGURE 4-19). Nuclei, cells, and organisms with one set of chromosomes in each nucleus are said to be haploid. The zygote has two complete sets (one from each gamete), so it is diploid. In most species, the zygote grows into an adult by cell divisions in which the nucleus divides by mitosis. As a result, all cells of the adult are diploid because all nuclei are replicates of the mother nucleus.
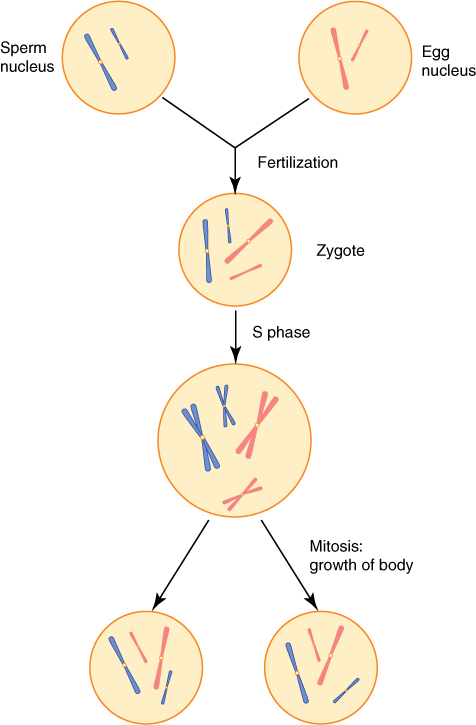
FIGURE 4-19 Sperm cells and egg cells are haploid, each having just one set of chromosomes; in this case, the set contains a long chromosome and a short one. After fertilization, the zygote is diploid with two complete sets: two long chromosomes and two short ones. The zygote grows into a mature plant by mitosis, so all nuclei are duplicates of the original zygote nucleus. Each is diploid, with one set each of paternal and maternal chromosomes.
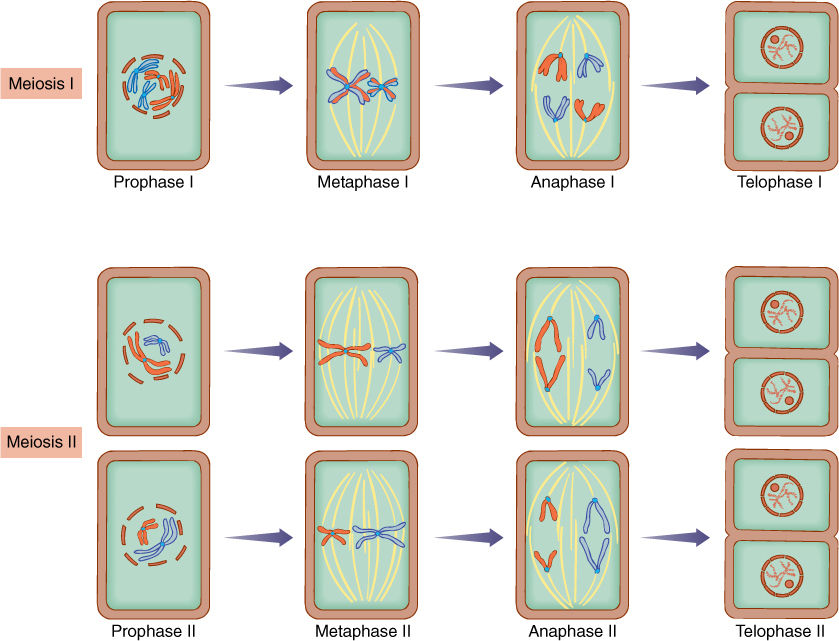
FIGURE 4-20 Meiotic nuclear division consists of two divisions without an intervening S phase. Details are given in text.
If the adult were to produce gametes by mitosis, the gametes would be diploid as well, and the next zygote would be tetraploid with four sets. It is not possible biologically to double the number of sets per nucleus with each new generation; instead, it is necessary for a reduction division, called meiosis, to occur somewhere. Meiosis involves two rounds of division without allowing the S phase to occur after the first division. The two divisions are called meiosis I and meiosis II, and each contains four phases similar to those of mitosis (FIGURE 4-20).
Meiosis occurs only in the production of reproductive cells: gametes in animals and some algae and fungi and spores in plants and other algae and fungi. In seed plants, meiosis occurs in only a few cells in the stamens and ovaries. Meiosis is never used in the growth of the body of any organism.
Meiosis I
Prophase I
All events that characterize prophase of mitosis also occur in prophase I: Nucleolus and nuclear membrane break down; centrioles (if present) separate; a spindle forms; microtubules attach to centromeres, and chromosomes condense and become visible (Figure 4-20). In addition, special interactions of chromosomes occur that are unique to prophase I as opposed to prophase of mitosis. Because of this, prophase I is divided into five stages:
1. In leptotene, chromosomes begin to condense and become distinguishable, although they appear indistinct.
2. During zygotene, a remarkable pairing of chromosomes occurs (FIGURE 4-21A). There are two sets of chromosomes, one from the paternal and one from the maternal gamete. Because these have gone through S phase in the preceding interphase, each chromosome has two chromatids; altogether there are four sets of genes on two sets of chromosomes. During zygotene, each chromosome of one set finds and pairs with the equivalent chromosome (its homologous chromosome, or homolog) of the other set; this pairing is synapsis. With remarkable accuracy, the two homologous chromosomes in each pair become almost perfectly aligned from end to end. A structure, the synaptonemal complex, is present between the paired homologous chromosomes (FIGURE 4-21E). This complex is composed of a linear central protein element connected by fine transverse fibers to two lateral elements bound to the DNA of the homologous chromosomes. A synapsed pair of homologous chromosomes is called a bivalent.
3. As chromosomes continue to condense, they become shorter and thicker; this stage is pachytene. The synaptonemal complex seems to be involved in the crossing-over that occurs now: In several places in each chromosome, the DNA of each homolog breaks (Figure 4-21). Breaks occur in almost identical places in the paired homologs, and the enzymes that repair the breaks hook the “wrong” pieces together. A piece of the maternal homolog is attached to the paternal homolog, and the equivalent piece of the paternal homolog is attached to the maternal one. The full consequences of this are explained in texts on genetics and evolution. If the maternal and paternal chromosomes are absolutely identical, nothing significant has happened, but if the genes on the paternal and maternal chromosomes are slightly different, the new chromosomes that result from synapsis and crossing-over are slightly different from the original chromosomes. There are no new genes, but rather, there are new combinations of genes on each chromatid. This greatly increases the genetic diversity of all haploid cells produced: Every sperm cell or egg cell produced by any single plant or animal (yourself included) is unique. This genetic diversity is important for evolution.
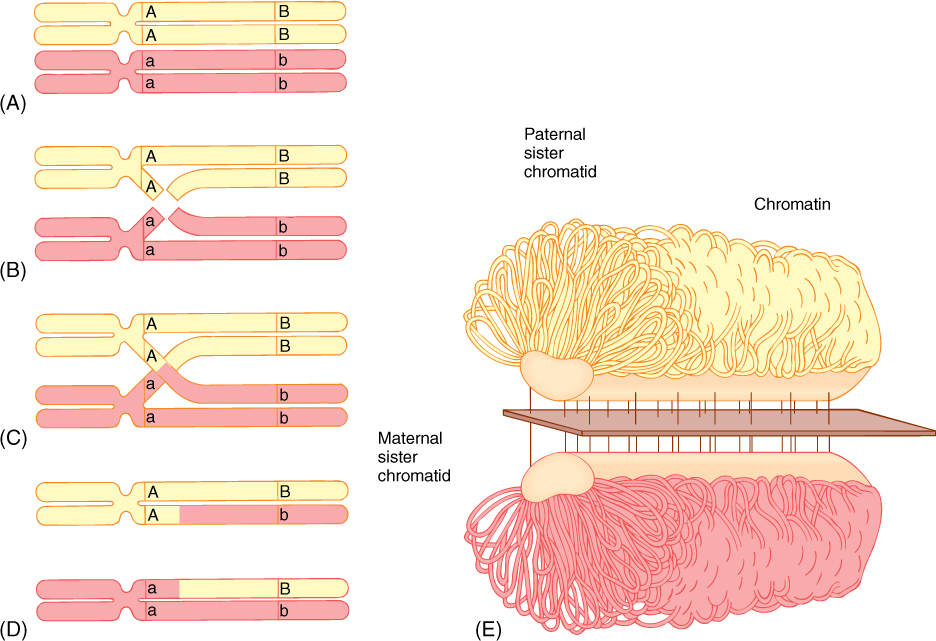
FIGURE 4-21 (A) During prophase I of meiosis, each paternal chromosome somehow finds and pairs with the equivalent maternal chromosome; they lie parallel to each other. (B) and (C) Breaks occur in similar sites on equivalent chromatids, and (D) repair enzymes attach maternal pieces to paternal pieces, resulting in new chromatids. (E) The synaptonemal complex between paired homologous chromosomes. It is not known how it facilitates crossing over.
4. After pachytene is diplotene. The homologous chromosomes of each bivalent begin to move away from each other but do not separate completely because they are held together at their paired centromeres and at points (chiasmata; singular, chiasma) where they appear to be tangled together. Some biologists believe that chiasmata are the points at which crossing-over occurred, but evidence is accumulating that chiasmata are only tangles and are not related to crossing-over at all. Under good conditions, it is possible to see all four chromatids of the paired homologous chromosomes; they are called tetrads at this stage.
5. In the final stage, diakinesis, homologs continue to separate, and chiasmata are pushed to the ends of the chromosome. The homologous chromosomes become untangled and are paired only at the centromeres.
Prophase I is the most complicated stage of meiosis; the remaining stages are simple and quite similar to the stages of mitosis.
Metaphase I
Spindle microtubules move the tetrads to the center of the cell, forming a metaphase plate (Figure 4-20).
Anaphase I
The homologous chromosomes separate completely from each other, moving to opposite ends of the spindle. Cohesin is not digested; thus, centromeres do not divide, and each chromosome continues to consist of two chromatids. Notice how this is different from the metaphase-anaphase transition of mitosis. In mitosis, separase degrades cohesin, releasing centromeres from each other, and each chromosome divides into two chromosomes, each with just one chromatid. However, in the metaphase I—anaphase I transition, homologous chromosomes separate from each other, and each still has two chromatids. One set of chromosomes is pulled away from the other set, and two new nuclei are formed. These nuclei are now haploid because each has only one set of chromosomes; the homolog of each chromosome in one nucleus is now in the other nucleus. Note that each chromosome in anaphase I has two chromatids; thus, each nucleus has two sets of genes, but this does not make them diploid. Any cell in G2 has at least two sets of genes; the critical factor is to have two sets of chromosomes.
Telophase I
Because the chromosomes are still doubled (each has two chromatids) and are in a G2 state, they do not need to undergo an interphase with its G1, S, and G2. Also, because telophase I is basically the opposite of prophase II, some organisms go directly from anaphase I to metaphase II, skipping telophase I and prophase II. In most organisms, however, there is at least a partial telophase I in which chromosomes start to uncoil, and the nucleolus and nuclear envelope start to reappear. If the cells actually progress fully to interphase, no replication of the DNA occurs—the S phase is completely missing. This interval is called interkinesis. Cytokinesis may occur, but it is not unusual for this to be absent also and for both daughter nuclei (i.e., both masses of chromatin) to stay in the original, undivided mother cell.
Meiosis II
If a telophase I occurs, then prophase II is necessary to prepare the nucleus for division. Prophase II is not subdivided into stages like prophase I. Metaphase II is short, and at the end of it, the centromeres divide, thereby separating each chromosome into two chromosomes, just as in metaphase of mitosis, but different from metaphase I. Anaphase II then separates each new chromosome from its replicate, and in telophase II, new nuclei are formed. Each nucleus contains just one set of chromosomes, each with a single chromatid.
To summarize, during meiosis I, each chromosome of the paternal chromosomes undergoes synapsis with the homologous chromosome of the maternal set. Crossing-over results in new combinations of genes on the chromosomes. Spindle microtubules pull the paired homologs to opposite poles; therefore, at each pole, there is only one set of chromosomes, not two. The new nuclei that form temporarily after meiosis I are thus haploid. When these nuclei undergo meiosis II, the two chromatids of each chromosome are separated, as in mitosis, and the resulting nuclei each have a haploid set of chromosomes, each chromosome with only one chromatid.
In meiosis, nuclear division and cell division are often not directly linked. In some species, cytokinesis happens after both meiosis I and meiosis II, and four haploid cells result from each original diploid mother cell. In other species, no cytokinesis occurs after meiosis I, but a double cytokinesis occurs after meiosis II, again resulting in four haploid cells. For example, pollen grains can form either way, and meiosis results in four haploid cells; however, in many organisms, no cytokinesis occurs at all during the meiosis that leads to the formation of eggs. The final cell is tetranucleate and may remain that way, or three of the nuclei may degenerate and produce only a single uninucleate haploid cell by meiosis (FIGURE 4-22). It seems to be selectively advantageous to produce one large cell rather than four small ones.
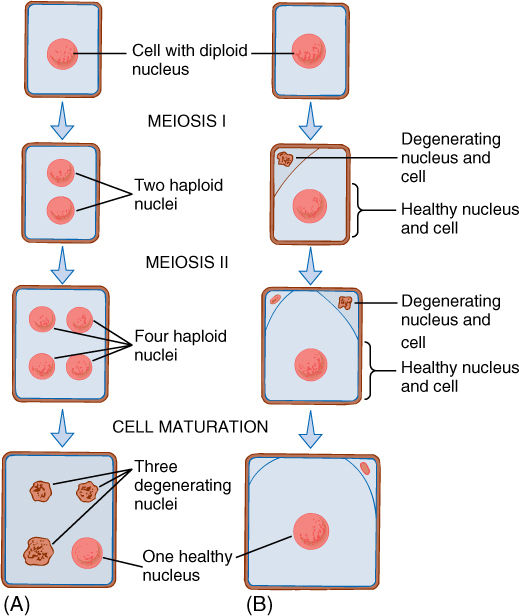
FIGURE 4-22 In animals and some plants, the meiosis that is part of egg production produces only one large haploid cell. (A) In some species, meiosis results in four haploid nuclei; no cytokinesis occurs and all four nuclei are temporarily in one cell until degeneration eliminates three of them. (B) In other species, after meiosis I occurs, cytokinesis places one nucleus in a small cell, and both nucleus and cell degenerate before they can undergo meiosis II. The nucleus of the larger cell does undergo meiosis II, and again, one daughter nucleus is placed in a small cell that degenerates. Only three nuclei are ever formed, and only one survives.
![]() Less Common Types of Division in Plants
Less Common Types of Division in Plants
Cytokinesis and karyokinesis are often so closely associated that we tend to think of them as two aspects of one process. In numerous tissues, however, karyokinesis occurs without cytokinesis, and multinucleate cells are formed. The nutritive tissue of many seeds goes through a phase in which its cells are multinucleate (FIGURE 4-23), and in many algae, all cells are like this. If the cell becomes very large and has hundreds or thousands of nuclei, it is called a coenocyte.
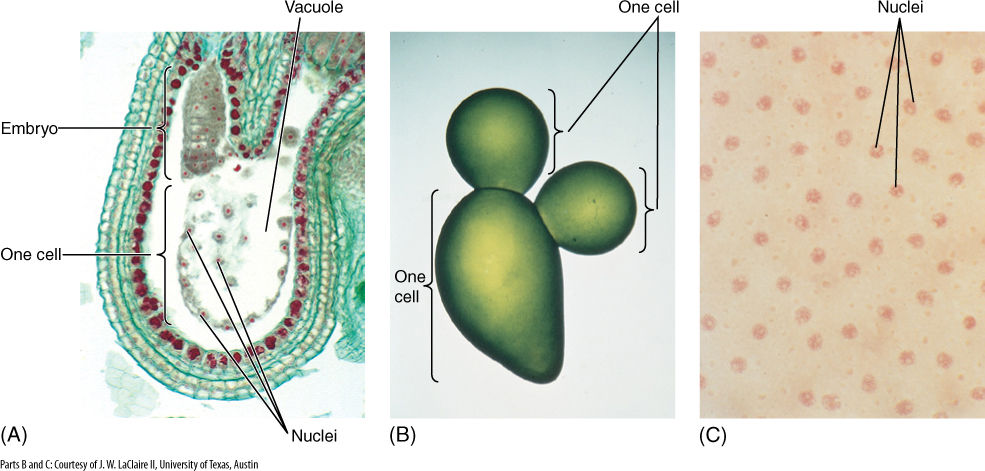
FIGURE 4-23 (A) A multinucleate cell in the developing seeds of Capsella (shepherd’s purse). Although numerous nuclei have undergone mitotic divisions, the cell has not divided but has enlarged and now has a giant vacuole. The small ball of cells is the embryo, which pushes into the multinucleate coenocyte and draws nutrients from it (× 80). (B) Multinucleate cells are typical of many algae. This is Valonia; the entire alga pictured here consists of just three giant cells (this photo is only four times life size), but each has thousands of nuclei. (C) A magnification of (B), showing some of the many nuclei, which have been stained pink (× 320).
On the other hand, cell division may occur without nuclear division; this is most common in algae, fungi, and the nutritive tissues of seeds. The cells first become multinucleate and persist in that condition for some time; then cell division begins as new walls are organized around each nucleus and its accompanying cytoplasm.
In lily flowers, each haploid cell involved in producing an egg cell undergoes two rounds of mitosis without cytokinesis, resulting in a tetranucleate cell (called the first four-nucleate stage; FIGURE 4-24). Three nuclei migrate to one end of the cell, the fourth nucleus migrates to the other, then all four undergo mitosis again. The isolated nucleus divides into two nuclei as would be expected; however, as the three crowded nuclei begin to divide, their spindles fuse into a single structure. Thus, all three sets of chromatids on one side go to one pole of the spindle, and all three on the other side go to the other spindle: The result is not six haploid nuclei but instead two triploid nuclei. The cell now has two small haploid nuclei and two large triploid nuclei (called the second four-nucleate stage). The egg is always formed from one of the haploid nuclei, never one of the triploid ones.
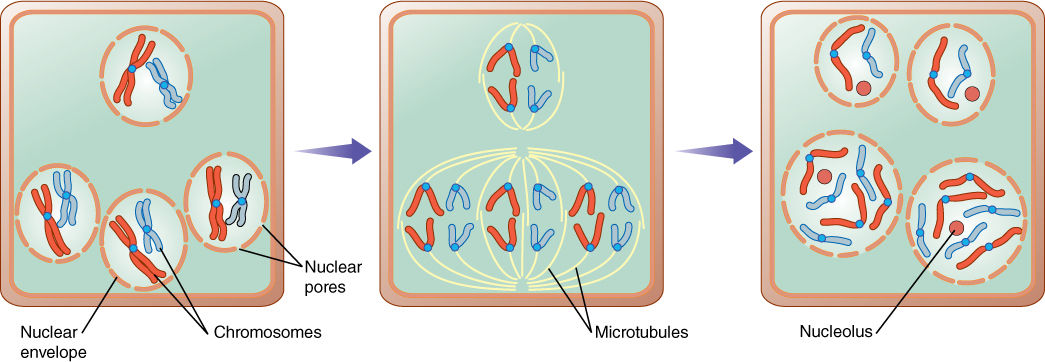
FIGURE 4-24 As part of egg production in lilies, certain cells go through an odd karyokinesis in which three haploid nuclei divide, but because they lie so close to each other, their spindles fuse and two new triploid nuclei are formed instead of six haploid ones.
![]() Cell Division in Algae
Cell Division in Algae
Nuclei
The nuclei of plants are virtually identical to those of animals in terms of structure, metabolism, mitosis, and meiosis. Apparently, the plant and animal lines of evolution diverged only after the eukaryotic nucleus had become quite sophisticated; however, nuclei are complex organelles, and mitosis and meiosis are intricate processes with many steps. Certainly plant and animal nuclei could not have arisen from prokaryotic nucleoids quickly; many intermediate steps spanning hundreds of millions of years must have been involved.
Several groups of organisms with unusual nuclear characteristics may represent lines of evolution that originated earlier in the history of eukaryotes than did plants and animals. Nuclei of many fungi have an unusual mitosis, some having both an intranuclear and an extranuclear spindle. In many fungal species, the nuclear envelope and nucleolus either do not break down at all or do so only late in mitosis. Similar behavior occurs in many groups of algae, including dinoflagellates, brown algae, euglenoids, and some green algae (FIGURE 4-25A). Gaps form in the nuclear envelope, and bundles of microtubules pass completely through the nucleus. Even more significantly, nuclei of dinoflagellates have no histones. Their chromosomes remain condensed at all times, undergoing only a slight uncoiling during interphase (FIGURE 4-25B). Because histone and nucleosome structure is so constant in all other eukaryotes, this unique situation in dinoflagellates is considered highly significant.
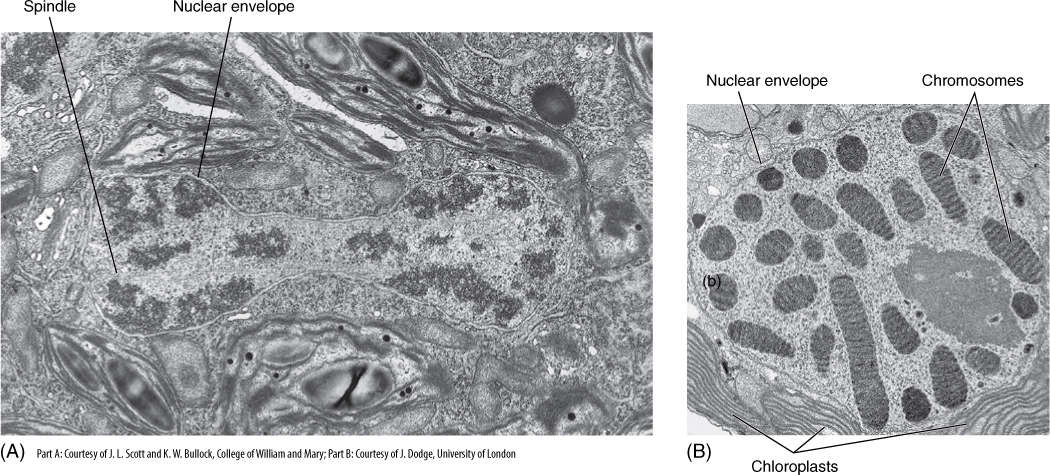
FIGURE 4-25 (A) Late telophase in the green alga Cladophora; not only is the nuclear membrane completely intact, but a portion of the intranuclear spindle is visible (× 26,656). (B) An interphase nucleus of dinoflagellate. Chromosomes never completely decondense. It is not known how chemical messengers, DNA replicases, and RNA synthetases gain access to the DNA molecule (× 21,000).
Cytokinesis
Several types of cytokinesis occur in algae. Many groups of unicellular algae have no wall, their plasma membrane being their outermost surface. During cell division, the plasma membrane pinches in two, being pulled inward as a cleavage furrow, a process remarkably similar to cytokinesis in animals (FIGURE 4-26A).
In many algae with walls, cell division is similar to that of plants: After telophase, a phragmoplast forms, and dictyosome vesicles establish a cell plate, which then grows radially outward—centrifugal growth—until it meets and fuses with the wall of the parental cell.
In some green algae cytokinesis occurs by a different method; the mitotic spindle depolymerizes quickly, and the two daughter nuclei lie close together. A new set of microtubules appears between them, oriented parallel to the plane where the new wall will form, which is perpendicular to the orientation of the spindle. This group of microtubules is a phycoplast and may be associated with division either by furrowing or by cell plate formation (FIGURE 4-26C and 4-26D). Phycoplasts are associated with those green algae in which the nuclear envelope does not break down during mitosis, whereas phragmoplasts are associated with the loss of the nuclear envelope. Plants arose from green algae that divide with a phragmoplast rather than a phycoplast.
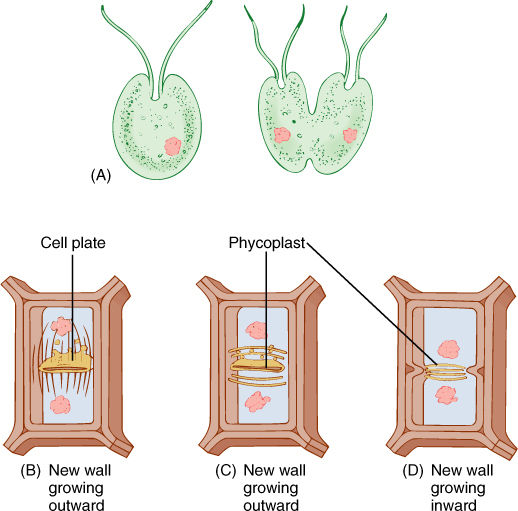
FIGURE 4-26 (A) In unicellular, wall-less forms, cell division occurs by infurrowing of the plasma membrane. (B) Most algae with walls divide by a phragmoplast and cell plate, just as plant cells do. (C) and (D) Some green algae have a phycoplast; the mitotic spindle disorganizes quickly, after which a new set of microtubules polymerizes perpendicular to the spindle axis. Division may then occur by cell plate growth (C) or by infurrowing (D).
In red algae cell division occurs by a phragmoplast, but the new wall grows inward from the pre-existing walls. It stops growing while still incomplete, having a large hole called a pit connection, which is filled with protein and carbohydrate material (a pit plug) (FIGURE 4-27). Otherwise, most aspects of wall formation are similar to those of other walled algae.
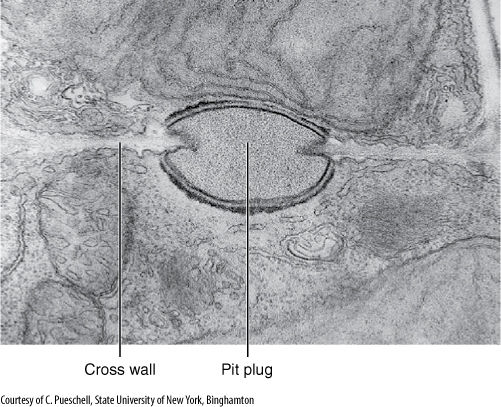
FIGURE 4-27 Cross walls of red algae are unusual in being incomplete—having a hole in their center. This is closed by a pit plug. Scinaia confusa (× 15,000).
![]() Cell Division of Prokaryotes
Cell Division of Prokaryotes
The events of mitosis and meiosis occur only in eukaryotes. In prokaryotes—bacteria, cyanobacteria, and archaea—mitosis and meiosis do not occur, and cytokinesis is much simpler. All genetic material occurs as one or several rings of DNA, which does not have any protective histones. The loops of DNA are attached to the cell’s plasma membrane, and when the DNA is replicated, the two daughter loops also are both attached to the membrane (FIGURE 4-28). Because no centromere holds them together, the circles of DNA are pulled apart as the cell and its membranes grow. Several new rounds of replication may occur before cell division takes place, and thus, each cell may have as many as 20 identical circles.
Cytokinesis occurs by a process of infurrowing: The plasma membrane pulls inward and finally pinches in two. As the plasma membrane furrows inward, a new cross wall grows inward, starting from the existing wall. After completion, the cross wall splits, becoming two walls and releasing the two daughter cells; multicellular aggregates are rarely formed in bacteria and archaea. In cyanobacteria, cross walls do not break apart, and multicellular bodies are formed. The cell cycle of many bacteria can be short—only 20 minutes under ideal conditions. For many species, however, the cycle may last several days or weeks even under optimal conditions.
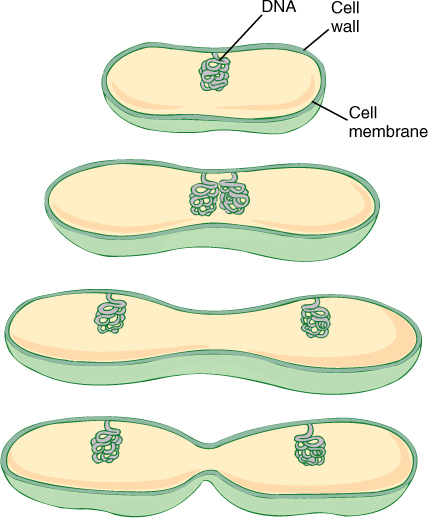
FIGURE 4-28 In prokaryotes, the circles of DNA are attached directly to the plasma membrane. As the cell and its membrane grow, the attached circles of DNA are separated. Cell division then occurs between the circles.
Botany and Beyond
BOX 4-3 Chloroplast Division During Leaf Growth
Elegant studies have been done of the growth and division of plastids in relationship to growth and development of leaves in spinach. In very small leaves, 1 mm long or less, plastid DNA constitutes 7% of the total cell DNA, and an average of 76 DNA circles are present in each plastid (TABLE B4-3). As the leaf doubles in size to 2 mm long, plastid DNA is replicated at approximately the same rate as nuclear DNA, and thus, it remains low, approximately 8% of total cellular DNA. As the leaf continues to expand to 20 mm long, plastid DNA is replicated much more rapidly than nuclear DNA and increases to 23% of total cellular DNA. At the same time, the number of plastids per cell triples from 10 to 29, and thus, neither plastid division nor plastid DNA replication is controlled by the mechanisms that govern cell or nuclear DNA replication. At this point, each plastid has 190 DNA circles, and each cell has a total of 5,510. In the next stage of leaf growth, to 100 mm long, no synthesis of DNA occurs, and no new cells form. Instead, those already present expand; however, plastids continue to divide even though they are not making any more DNA. Consequently, the number of DNA circles per plastid drops from 190 to 32, whereas the number of plastids per cell increases from 29 to 171. From these data, it is reasonable to form the hypothesis that plastid growth, DNA replication, division, and development are correlated predominantly with tissue or organ development rather than with the cell cycle.
TABLE B4-3 Development of the Plastid Genome in Spinach Leaves
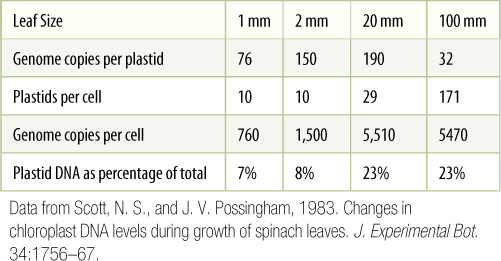
Data from Scott, N. S., and J. V. Possingham, 1983. Changes in chloroplast DNA levels during growth of spinach leaves. J. Experimental Bot. 34:1756—67.
![]() Division of Chloroplasts and Mitochondria
Division of Chloroplasts and Mitochondria
Mitochondria and plastids are constructed similarly to prokaryotes; they also contain circles of naked DNA that become separated by membrane growth. Division of the organelles is accomplished either by infurrowing or by being pulled in two (FIGURE 4-29). Because they contain DNA necessary for their growth and functioning, each daughter cell must receive at least one mitochondrion and one plastid during cytokinesis; if not, the cell lacks that organellar portion of its genome and cannot produce the organelle. This happens occasionally with plastids and is often not a serious problem; the cell survives by importing sugar from neighboring cells and can grow along with the tissue. All daughter cells also lack plastids, so if this occurs in a young leaf, a white spot forms. The same phenomenon probably occurs with mitochondria but is more difficult to detect.
The metabolic stimulus that triggers the replication of nuclear DNA is not the stimulus that controls replication of organellar DNA. During the cell cycle, replication of nuclear DNA is episodic, occurring as discrete episodes that occupy a small portion of the total cell cycle, S phase; however, the “duplication” of the rest of the cell seems to be continuous: The volumes of plastids, mitochondria, cytosol, endoplasmic reticulum, and other organelles appear to increase gradually and steadily throughout interphase rather than in discrete episodes (Box 4-3).
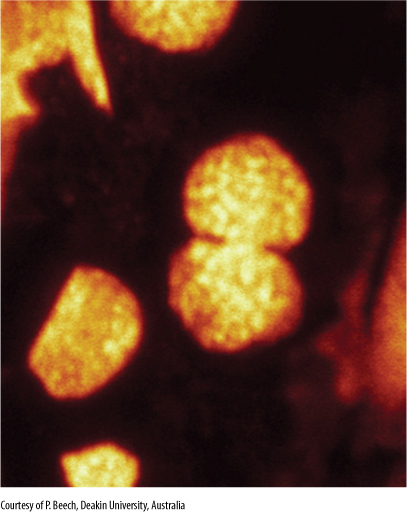
FIGURE 4-29 Micrograph of a chloroplast dividing in Arabidopsis thaliana. As the chloroplast divides, its central region is pinched inward by a contracting ring of proteins, which constricts the chloroplast into a dumbbell shape. The chloroplast shines brightly because of the way its chlorophyll reacts to the light used by the microscope: It absorbs the light and re-emits it as a different color (this is called fluorescence microscopy) (× 30,000).
 At the Next Level
At the Next Level
1. Orientation of cell division. The orientation of cell division in plants determines the positions of the cells as they differentiate. Plant cells cannot migrate to new positions in the way animal cells can, they must be produced in the proper place. Cells usually divide into two cells that are more or less equal in size, but during the formation of stomata in the epidermis, cells often divide asymmetrically into a large cell and a small one. Also, when a plant is wounded, surviving cells near the would typically divide in such a way as to produce a layer of protective scar cells (called wound cork) near the wound. The mechanisms that determine where the pre-prophase band forms are critical in establishing the plane of cell division.
2. Control of the cell cycle. Chemical factors control when cells replicate their DNA, when they start to divide, how rapidly they divide, and when they stop. Cells and growing tissues (for example in the tips of roots and shoots and in very young leaves) divide repeatedly, but at some point, some or all these cells must stop dividing and begin to differentiate into specialized, mature cells. Also during autumn or when stressed, cell division must be slowed or stopped, whereas during seed germination, certain factors influence certain cells to initiate the division cycle. Another important but poorly understood process determines whether cells undergo mitosis or meiosis.
3. Cell formation without nuclear division. During seed development in angiosperms, a tissue called endosperm develops in an unusual manner. In the first stage, endosperm nuclei divide frequently but there is no cell division, so a giant multinucleate cell is formed. The center of an unripe coconut is one of these giant cells, the coconut milk is the cell’s protoplasm. In the second stage, hundreds of cell walls form, trapping some cytoplasm and nuclei, creating hundreds or thousands of cells; this is “cellularization of endosperm” and it produces the white “coconut meat” we use in candy, cake, and pies.
SUMMARY
1. Cells have a “life cycle,” the cell cycle, consisting of alternating phases of growth and division. In organs that live just briefly, cells cycle only while the tissue is young and then enlarge and differentiate.
2. The interphase portion of the cell cycle consists of G1, S, and G2. During G1, the nucleus controls cell metabolism. During S, the DNA is replicated; in G2, the cell prepares for division.
3. Each DNA molecule contains thousands of regions—genes—that contain the information for proteins. A chromosome is a complex in which histone proteins bind to DNA and stabilize it. Each chromosome consists of a centromere and either one chromatid before S phase or two chromatids after S phase. Chromatid ends are telomeres.
4. Cell division is cytokinesis, and nuclear division is karyokinesis. The words “mitosis” and “meiosis” technically refer to only karyokinesis.
5. During mitosis, spindle microtubules pull the two chromatids of each replicated chromosome to opposite ends of the cell. Because the two chromatids are virtually identical, the two new nuclei are also almost identical to each other.
6. Nuclei of any ploidy level—haploid, diploid, and higher ploidies—can undergo mitosis. During growth of the plant body, all nuclear divisions are mitotic divisions.
7. Meiosis is reduction division: diploid cells produce haploid cells. Just as important, genetic diversity is increased by meiosis.
8. During prophase I, homologous chromosomes pair (synapse) and exchange pieces of chromatid (crossing-over). At the end of prophase I, the nucleus no longer has purely paternal and purely maternal chromosomes. During anaphase I, homologous chromosomes are pulled away from each other, reducing the number of sets of chromosomes to half the original number. Haploid, triploid, and all other odd-ploid nuclei cannot undergo meiosis.
9. Meiosis occurs only in the production of reproductive cells, either gametes or spores. Meiosis is never used in the growth of the body of any organism.
10. Cytokinesis in plants and algae occurs by the formation of a large vesicle between the forming daughter nuclei. Within the vesicle, the middle lamella and two new walls form and expand until they reach existing walls.
11. Cytokinesis and karyokinesis are usually closely coordinated, but nuclear division can occur without cell division, resulting in multinucleate cells.
IMPORTANT TERMS
anaphase
arithmetic increase
cell cycle
cell plate
centromere
chromatid
chromosome
chromosome condensation
crossing-over
cytokinesis
diploid
duplication division
G1 phase (gap 1)
G2 phase (gap 2)
gametes
genome
geometric increase
haploid
histones
homologous chromosome
interphase
karyokinesis
kinetochore
meiosis
metaphase
metaphase plate
mitosis
phragmoplast
phragmosome
phycoplast
preprophase band
prophase
reduction division
resting phase
S phase
spindle
spindle microtubules
synapsis
telophase
tetraploid
zygote
REVIEW QUESTIONS
1. In a woody plant such as a tree, which parts have cells that live only briefly and die quickly? Which parts have cells that live for several years?
2. Some cells never stop dividing. Give two examples of cells like this.
3. Interphase is also called the resting phase of the cell cycle. Why was it given that name?
4. What are the main activities of a cell while it is in G1 phase? What kinds of organisms have short G1 phases, and how long does a short G1 phase last? What kinds of organisms have long G1 phases, and how long do they last?
5. What is the main activity of the S phase of the cell cycle? What does “S” stand for?
6. Table 4-2 gives the number of chromosomes in a haploid set of chromosomes. What is the lowest number in the table? What is the highest number? Most cells in plants are diploid, having two sets of chromosomes, so the number per nucleus should be doubled. How many chromosomes are present in each diploid potato nucleus? If a potato plant has 1 million cells, each with one diploid nucleus, how many chromosomes are present in the entire plant?
7. How many chromatids does a chromosome have before S phase of the cell cycle? How many does it have after S phase?
8. Examine Table 4-1. How many hours does the cell cycle last in the root tips of corn and in onion? Which plant has cells that divide more quickly? How many cell cycles could corn and onion roots undergo in 4 weeks (the cell cycles in Table 4-1 are given in hours, not days)? If after a cell divides both daughter cells could divide and then their daughter cells could divide, how many corn cells and how many onion cells would be present at the end of 4 weeks if you started with just one cell of each?
9. What are the four phases of the cell cycle? What is the principal activity in the cell during each phase? Can any phase be eliminated or bypassed?
10. Why is mitosis called duplication division and meiosis called reduction division? What is reduced and what is duplicated: chromosomes, number of chromosomes, or number of sets of chromosomes?
11. What does it mean when chromosomes are said to condense during prophase of mitosis? How long are chromosomes after condensation is complete? How big is a typical dividing cell in a root or shoot? If a chromosome were still 20 μm long after condensation, would it be possible for division to pull half of it to one end of the cell and the other half to the other end?
12. What is the name of the set of microtubules that pull chromosomes apart? What is the name of the attachment point between microtubules and chromosomes? What is the name of the ends of the cells where the chromosomes are pulled?
13. What is a metaphase plate? What is duplicated at the end of metaphase? When this is duplicated, chromatids become free of each other. How many chromatids are there per chromosome in prophase? How many chromatids per chromosome after metaphase?
14. Imagine a nucleus that has 10 chromosomes. How many chromosomes does it have before prophase begins? How many chromosomes does each daughter nucleus have after telophase has been completed? How many chromatids are present in the one mother nucleus before prophase begins? How many chromatids are present in each daughter nucleus after telophase has been completed? How many chromatids are present if you add together all chromatids in both daughter nuclei after telophase has been completed?
15. What are the four phases of mitosis, and what is the principal activity in the nucleus during each phase?
16. Draw a single, imaginary chromosome as it would appear just as mitosis is ending. Now describe what happens to it during interphase and then during mitosis. Be especially careful to consider how many chromatids and how many copies of each gene it has at each stage.
17. How does cytokinesis occur in plants? Which organelle produces vesicles that fuse to form the cell plate? What membrane is transformed into new plasma membrane?
18. Many people consider algae to be plants even though algae do not have roots, stems, and leaves, but in many algae, cell division is different from that in true plants. Do most algae have a phragmoplast? What is the name of the structure they use?
19. Sex cells are also called ____________________. If a species has males and females (not all species do), males produce sex cells ____________________ called cells, and females produce sex cells called ____________________ cells.
20. How many sets of chromosomes does each sperm cell carry? How many does each egg cell have? Gametes such as sperm cells and egg cells are said to be (circle one: diploid, haploid). After a sperm and an egg have fused, the new cell is called a ____________________, and it is (circle one: diploid, haploid).
21. If a plant has diploid cells with 20 chromosomes in each nucleus, how many chromosomes does each of its gametes have? What is the name of the division that reduces the number of chromosomes from 20?
22. What are the five stages of prophase I, and what is the principal activity of the nucleus and chromosomes during each stage?
23. During zygotene stage of meiosis I, a remarkable pairing of chromosomes occurs. Look at the bottom part of Figure 4-19 in which “mitosis: growth of the body” has produced cells with four different types of chromosomes. The black ones came from the sperm nucleus. The red ones came from the egg nucleus. If one of these nuclei were to undergo meiosis, which chromosomes would pair during zygotene (which are homologous)—the long black one with the short black one or the long black one with the long red one?
24. What is the name of the structure that holds homologous chromosomes together after they have paired during zygotene?
25. During pachytene, crossing over occurs. Do chromosomes actually break during this process?
26. How does anaphase I of meiosis differ from anaphase of mitosis? Does the number of chromatids per chromosome change in anaphase I? Does the number of sets of chromosomes per nucleus change in anaphase I?
27. Draw all stages in the cell cycle and meiosis for a nucleus that has just one pair of homologous chromosomes, and then do the same for a nucleus that has three different types of chromosomes (six chromosomes in three sets of homologs). Draw all stages (this is not easy).
28. If a cell undergoes nuclear division but not cell division, it becomes multinucleate. If it has hundreds of nuclei, it is called a____________________.
29. Do prokaryotes undergo meiosis or mitosis? How do their cells divide?
30. Describe the process of division that produces more plastids and mitochondria.
Design Credits: Hummingbird: © Tongho58/Moment/Getty; Green plant cells: © ShutterStock, Inc./Nataliya Hora; Purple tulip: © ShutterStock, Inc./Marie C Fields; Dandelion: © ShutterStock, Inc./danielkreissl; Poppy: © ShutterStock, Inc./Saruri; Plant icon: © ShutterStock, Inc./Vector; Digging man icon: © ShutterStock, Inc./Z-art