Botany: An Introduction to Plant Biology - Mauseth, James D. 2017
Plant Structure
Leaves

Chapter Opener Image: Foliage leaves carry out photosynthesis, which requires that they be exposed to sunlight. But the vine maple (Acer circinatum) shown here is adapted to live in the shade of larger trees in dense forests. In this image, you can see that most leaves receive only dim light and other leaves receive almost no light at all. In contrast, the leaves of the taller trees—the ones that are shading this maple—are exposed to full, intense sunlight. Leaves of different species have evolved to be adapted to extremely diverse habitats.
OUTLINE
✵ Concepts
✵ External Structure of Foliage Leaves
✵ Internal Structure of Foliage Leaves
- Epidermis
- Mesophyll
- Vascular Tissues
- Petiole
✵ Initiation and Development of Leaves
- Basal Angiosperms and Eudicots
- Monocots
✵ Morphology and Anatomy of Other Leaf Types
- Succulent Leaves
- Sclerophyllous Foliage Leaves
- Leaves of Conifers
- Bud Scales
- Spines
- Tendrils
- Leaves with Kranz Anatomy
- Insect Traps
Box 6-1 Plants and People: Leaves, Food, and Death
Box 6-2 Botany and Beyond: Leaf Structure, Layer by Layer
Box 6-3 Alternatives: Photosynthesis Without Leaves
LEARNING OBJECTIVES
After reading this chapter, students will be able to:
✵ List three functions of leaves.
✵ Identify the parts of a leaf.
✵ Compare a simple leaf to a compound leaf.
✵ Explain the functions of the epidermis, mesophyll, vascular tissues, and petiole.
✵ Name the steps of leaf development.
✵ Compare development of monocot and eudicot leaves.
✵ Discuss the following leaf types: succulent, sclerophyllus foliage, conifer, bud scales, spines, tendrils, Kranz anatomy, and insect traps.
 Did You Know?
Did You Know?
✵ Some plants, such as lettuce and cabbage, consist almost entirely of leaves, having only a short stem and a few roots.
✵ The thin, flat shapes of most foliage leaves are adaptations that facilitate the absorption of sunlight and carbon dioxide.
✵ All foliage leaves have chlorophyll; some plants have red or purple foliage leaves due to such large amounts of red pigment that the chlorophyll is hidden.
✵ Leaves have such dense networks of vascular bundles (veins) that every leaf cell is close to cells of xylem and phloem.
![]() Concepts
Concepts
The term “leaf” usually calls to mind foliage leaves—the large, flat, green structures involved in photosynthesis. However, natural selection has resulted in numerous types of leaves that are selectively advantageous because they provide protection (bud scales, spines), support (tendrils), storage (fleshy leaves of bulbs), and even nitrogen procurement (trapping and digesting insects). Because protecting a bud from freezing or drying is very different from photosynthesizing, leaf structures and metabolisms that are selectively advantageous for a bud scale are different from those that are selectively advantageous for a foliage leaf. This principle is true for all types of leaves: For each function, certain modifications are advantageous, whereas others are not. Studying the various leaf types, their modifications, and the roles they play in the plant’s biology leads to an understanding of how structure, metabolism, and function are related.
The shoot system, containing both stems and leaves, demonstrates both division of labor and integration of distinct plant organs. Stems and leaves must function together if the plant is to survive and reproduce, but the optimal features for each organ are quite distinct. Leaves should be flat and thin for maximum absorption of light and carbon dioxide, and most of their tissues must be alive and differentiated into chlorophyll-rich chlorenchyma to carry out photosynthesis. Stems elevate leaves and conduct material to and from them, among other activities. Maximum conduction and support with minimum expenditure of construction material require a cylindrical stem; also, much of the stem—all of its tracheary elements and most fibers—must die to be functional. In woody species, new cells are added to the stem xylem and phloem each year, creating a massive wood and bark and an increasingly stronger and more conductive stem. Yearly accumulation of new cells onto leaves is not feasible; leaves typically live for only several months and are rather lightly constructed; however, in open sunny habitats, elevating leaves on a tall stem is often unnecessary, and a plant may consist almost exclusively of leaves and roots. In lettuce and cabbage, leaves are short lived and delicate, but in Agave and Yucca, they are massive (to more than 2 m long), thick, and perennial (see Figure 6-4A). Almost all leaves contain only primary tissues; secondary production of wood and bark in leaves is extremely rare.
The initial discussion in this chapter centers on foliage leaves because they are the most familiar. Then other leaf types are described and analyzed; by considering how these leaves differ functionally from foliage leaves, it is possible to determine the environmental factors important to leaf structure and how natural selection has produced diverse classes of leaves.
![]() External Structure of Foliage Leaves
External Structure of Foliage Leaves
The most obvious function of foliage leaves is photosynthesis, but other functions often taken for granted are just as important. Leaves must not lose excessive amounts of water. They must not allow entry of fungi, bacteria, or epifoliar algae. They must not be so nutritious and delicious to animals that they are a liability to the plant. They must not be such effective sails that the plant is blown over in a mild wind, and they must be cheap enough that the plant spends less carbohydrate building them than it recovers by their photosynthesis. If a foliage leaf fails in any of these aspects, the plant dies and the leaf’s photosynthesis has been useless. Many structural and physiological aspects of leaves that make them waterproof, pathogen resistant, and so on in some way interfere with photosynthesis. Although we might assume that foliage leaves are maximally adapted for photosynthesis, these other factors affect leaf biology and must be considered as well.
During photosynthesis, leaves absorb carbon dioxide and convert it to carbohydrate by using light energy. Because sunlight comes from one direction at a time, it is best for a portion of the leaf to be flat and wide, allowing maximum exposure to light and maximum surface area for carbon dioxide absorption (FIGURE 6-1). Because chlorophyll absorbs sunlight efficiently, light penetrates only a short distance, and thus, leaves can be quite thin; in a thick layer of chlorenchyma, the lowest cells would be in almost complete darkness, unable to photosynthesize. This flat, light-harvesting portion is the leaf blade (also called the lamina). The blade’s lower side is its dorsal surface, and larger veins protrude like backbones (synonym: abaxial side); the upper side is the ventral surface (synonym, adaxial) and is usually rather smooth.
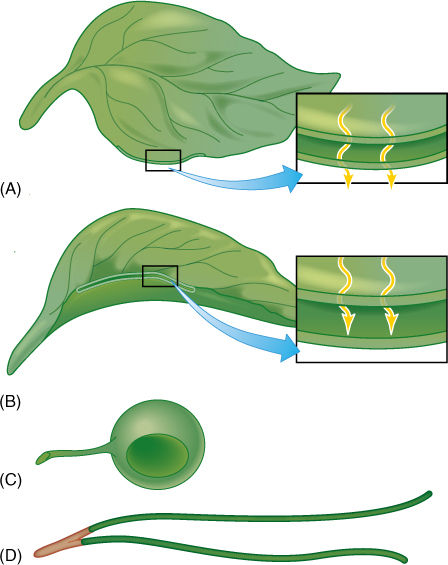
FIGURE 6-1 Several possible leaf shapes, all with equal volumes. (A) The common type with a thin, flat blade. All cells are exposed to light—there is a large surface area that absorbs CO2, and none of the veins are long. (B) In a thick leaf, light cannot penetrate to the bottom of the leaf, and those cells cannot photosynthesize. Relative to the volume, little surface area is available for CO2 absorption, but this helps conserve water. (C) A spherical leaf has maximum internal self-shading, and only a small fraction of the cells receives enough light for photosynthesis. Surface area for gas exchange is minimal. (D) A cylindrical leaf could be thin with a large surface area and no self-shading, as in (A); however, to have the same volume as (A), it would be extremely long, and conduction through the veins might be difficult.
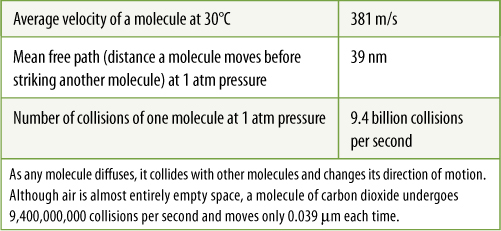
Most leaves have a petiole (stalk) that holds the blade out into the light (FIGURE 6-2). This prevents shading of leaf blades by those above them; self-shading would defeat the basic usefulness of the leaf. Petioles have other consequences. Long, thin, flexible petioles allow the blade to flutter in wind, cooling the leaf and bringing fresh air to its surface. If the leaf and air are still, carbon dioxide is absorbed from the vicinity of the leaf faster than diffusion brings more to it, depleting the carbon dioxide and decreasing photosynthesis (TABLE 6-1). Leaf flutter also makes it difficult for insects to land on a leaf and knocks off some that have already alighted.

FIGURE 6-2 The petioles of each leaf of poplar (Populus alba) are long enough that the leaves do not shade each other, and all are fully exposed to light. The lower (abaxial) side of each leaf blade is covered with white, dead trichomes, the upper (adaxial) surface is hairless and green.
TABLE 6-1 Movement of Carbon Dioxide
If leaves are small or very long and narrow, self-shading is not a problem, and there may be no petiole; the leaf is then called a sessile leaf instead of petiolate. For example, Aeonium grows in arid, sunny regions, and its fleshy leaves are packed close together (FIGURE 6-3). Because the sunlight is so intense, even with self-shading the leaves receive enough light for adequate photosynthesis. Close packing of these leaves helps trap water molecules and prevent their escape from the plant.
In many monocots, such as grasses, irises, lilies, agaves, and yuccas, foliage leaves tend to be very long and tapered, and self-shading occurs only at the base; most of the blade is well exposed to light (FIGURE 6-4 and see Figure 6-15). They also typically lack a petiole; instead, the leaf base wraps around the stem to form a sheathing leaf base. The blade can flex and flutter even without a petiole. Not all monocots have linear, grass-like leaves. Palms, aroids (such as Monstera and Philodendron), and bird-of-paradise plants have what appear to be petioles and laminas, but the leaves of these species are believed to have evolved from grass-like leaves, and the “petiole” may actually be a modified portion of the lamina (FIGURE 6-5).

FIGURE 6-3 Aeonium tabuliforme and its relatives grow in regions of intense sunlight; self-shading is not a problem, but water conservation is. Close packing minimizes water loss from stomata.

FIGURE 6-4 (A) Long, narrow leaves are common in many monocots of intensely sunny areas. Shading is no problem, although leaf bases probably do not photosynthesize at maximum efficiency. Sotol (Dasylision) (B) These are the leaves of St. Augustine grass (Stenotaphrum); the blades project horizontally and are attached to the stem by sheathing leaf bases. (C) In cattails (Typha), the sheathing leaf base can be half a meter long and provide firm but flexible attachment to the stem.
A leaf blade may be either simple or compound. A simple leaf has a blade of just one part (FIGURE 6-6), whereas a compound leaf has a blade divided into several individual parts (FIGURE 6-7 and FIGURE 6-8). Think of how this might be affected by wind: If the blade is either large or flimsy, it twists and flexes and may tear. Tearing can be prevented by making the leaf small or tough or “pretorn” (compound). A compound leaf has many small blades (leaflets), each attached by a petiolule to an extension of the petiole, the rachis. Leaves may be palmately compound, with all leaflets attached at the same point, or pinnately compound, with leaflets attached individually along the rachis (Figure 6-7). Even a large compound leaf has only small leaflets, all of which can flex individually without tearing. Flowing water is even denser than blowing air: Plants that grow immersed in streams typically have compound leaves with thread-like leaflets (FIGURE 6-9).

FIGURE 6-5 Maranta, a common houseplant, is a monocot that has broad leaves.
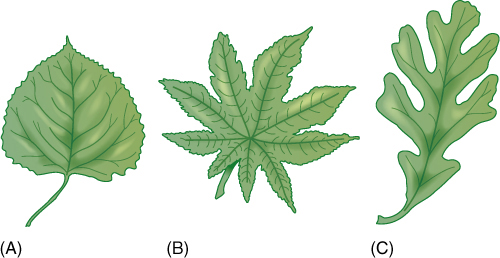
FIGURE 6-6 Simple leaves have numerous shapes, some so deeply lobed as to be almost compound. Poplar (A) and oak (C) have one dominant midrib from which lateral vascular bundles extend. In castor bean (B), petiolar bundles immediately diverge into several main veins.
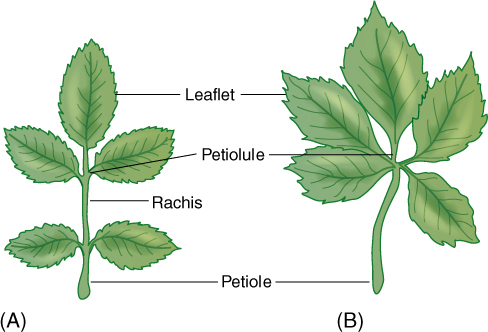
FIGURE 6-7 Leaflets of compound leaves have the same structure and metabolism as simple leaves. In pinnately compound leaves (rose, A) the petiolules of leaflets are attached to the rachis, but in palmately compound leaves (Virginia creeper, B) they attach to the end of the petiole. In either type, any leaflet can be damaged or can abscise without affecting the remainder of the leaf.
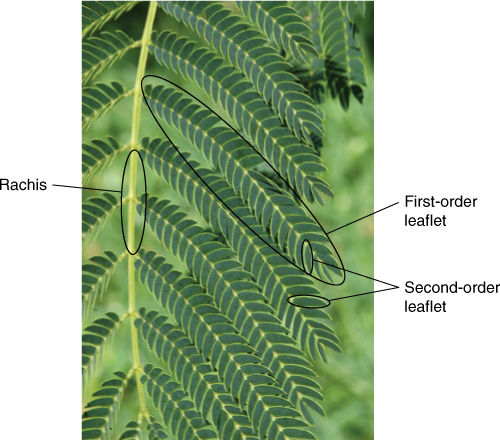
FIGURE 6-8 This is a portion of one doubly compound leaf of Mimosa: The central rachis (midrib) bears numerous leaflets, but each leaflet itself is pinnately compound.
Compound leaves have other advantages. When a mild breeze blows over two leaves of equal size, one simple and the other compound, it tends to flow smoothly over the simple leaf but turbulently over the more complex surface of the compound leaf. Turbulence brings in carbon dioxide and removes excess heat. Also, an insect can crawl or a fungus can spread across the entire blade of a simple leaf rather easily, but the edges and petiolule of a leaflet act as barriers that either prevent or at least slow movement to the rest of the leaf. In some plants, if pathogens severely damage a leaf, it abscises, carrying the pathogens with it and helping to keep pests away from healthy leaves.
Plants and People
BOX 6-1 Leaves, Food, and Death
Leaves impact our lives every day. Examples that come to mind readily are leafy foods such as artichokes (Cynara scolymus), cabbage (Brassica oleracea variety capitata), celery (petioles; Apium graveolens), lettuce (Lactuca sativa), onions (Allium cepa), and spinach (Spinacia oleracea). Also important are numerous herbs and spices: basil (Ocimum basilicum), bay leaves (Laurus nobilis), marjoram (Origanum majorana), oregano (Origanum vulgare), parsley (Petroselinum crispum), sage (Salvia officinalis), tarragon (Artemisia dracunculus), thyme (Thymus vulgaris), and the flavorings mint (several species of Mentha), spearmint (Mentha spicata), and peppermint (Mentha piperita) (FIGURE B6-1A-C). The flavors and pungency of these are due to chemicals located within the leaves themselves or in trichomes of the leaf epidermis. Most of these chemicals probably serve as antiherbivore defensive compounds, causing animals other than humans to avoid the plants. Many classes of antiherbivore chemicals have evolved, ranging from only mildly effective ones, such as these flavors, to others that are much more powerful, such as alkaloids, many of which are toxic in small amounts and kill quickly. The alkaloids in poison hemlock and death camas are particularly effective.
An alkaloid of the leaves of one plant in particular is of interest to us—it is quite lethal but acts only slowly: nicotine in the leaves of tobacco (Nicotiana tabacum) (FIGURE B6-1D). If tobacco leaves are eaten, they cause vomiting, diarrhea, and even death caused by respiratory failure; however, most tobacco leaves are smoked, of course. Americans smoked an all-time high of 4,345 cigarettes per capita in 1963. Since then, consumption has declined for white males but recently has increased for other groups. Tobacco leaves contain between 0.6% and 9.0% nicotine, and an ordinary filter cigarette has 20 to 30 mg of the alkaloid, approximately 10% of which is absorbed by the lungs. Nicotine dissolves readily into our mucous membranes and passes quickly into our blood stream. Because it is transferred across the placenta, women who smoke during pregnancy may give birth to babies addicted to nicotine. Blood-borne nicotine also affects the heart, causing coronary problems: People who smoke a pack or more a day are over three times more likely than nonsmokers to die of heart disease. After nicotine is taken into the cells of the mouth and lungs of a smoker, it can cause cancer—of the lungs especially, but also of the throat, larynx, and mouth. If detected early enough, nicotine-induced lung cancer can be combated with surgery and chemotherapy, but after the cancer has spread to the lymph system, the prognosis is not good. Lung cancer causes more than 400,000 deaths per year in the United States.

FIGURE B6-1 (A) Basil (the dots are glands that provide its flavor). (B) Spearmint. (C) Rosemary. (D) Tobacco.

FIGURE 6-9 Myriophyllum heterophyllum grows immersed in flowing streams; its leaves are very delicate and pinnately compound with thread-like leaflets (leaflets have no lamina). If the stream is quiet, some stems emerge into the air and produce thicker, tougher leaves (not present here), hence the species name “heterophyllum.”
Is it reasonable to conclude that plants with compound leaves are better adapted than plants with simple ones? Why are there any simple leaves at all? Most very large leaves of plants are compound, apparently the only feasible architecture. The large leaves of palms and bananas are formed with a large, single lamina, but they have numerous special lines of weakness and are quickly torn to a “compound” condition by wind. The largest simple leaves, those of the Victoria water lily, float on water and thus do not need large amounts of sclerenchyma for support, and adhesion to the water’s surface prevents wind damage. Among medium and small leaves, simple ones are common, perhaps because a greater percentage of a simple leaf is composed of photosynthetic cells, whereas a compound leaf has a great deal of nonphotosynthetic rachis, petiolules, and edges.
Although most pinnately compound leaves are easily recognizable as leaves, some can be mistaken for a stem with simple leaves. Close examination reveals that leaflets never bear buds in the axils of their petiolules, and the tip of the rachis never has a terminal bud. Leaflets are always arranged in two rows, never in a spiral, whorled, or decussate phyllotaxy.
Some of the tremendous range of shapes of leaves and leaflets is shown in FIGURE 6-10. There are hypotheses but no total agreement on how different leaf shapes may be adaptive; many biologists believe that all function so well that none is clearly superior or strongly disadvantageous selectively. A large number of species, however, have several types of leaves (FIGURE 6-11A). In the simplest cases, the first few leaves of a seedling are distinctly different from all leaves produced later; if they were merely smaller, it could be argued that the seedling does not have enough stored energy resources to construct the large leaves that a mature plant can afford. But very often leaves of juvenile plants differ from those of adults not only in size but also shape, texture, and even simple versus compound structure. In species such as ivy or citrus, transition from juvenile foliage to adult leaves does not take place until the plant is old enough to flower, perhaps more than 10 years old. It seems reasonable to hypothesize that juvenile leaves of seedlings are adaptive in the microhabitat close to the soil surface where a seedling is located, whereas adult leaves are adapted to the more aerial microhabitat inhabited by an older, larger plant.
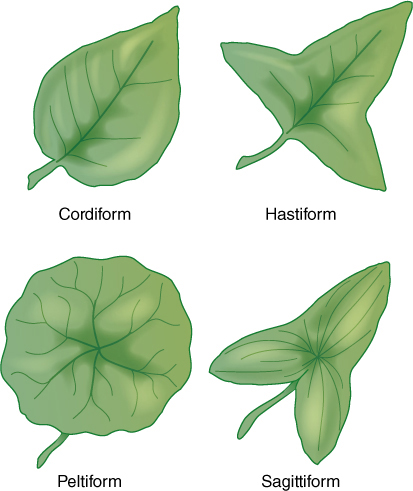
FIGURE 6-10 Four common leaf shapes; numerous other types also exist.

FIGURE 6-11 (A) Beans have two types of leaves: The very first leaf formed by the seedling is simple (right), but all later leaves are compound (left). (B) This Azara lanceolata shoot appears to have two types of leaves: The larger leaves are true leaves, whereas the smaller ones are actually enlarged stipules.
Other species produce two types of leaves simultaneously (FIGURE 6-11B), often one type on long shoots and the other on short spur shoots. In cacti, the spines are short-shoot leaves, whereas the long-shoot leaves are green, fleshy, and in most species almost microscopic. In a small percentage of species, leaf shape and size are irregularly variable even on one stem, being influenced by environmental conditions that occur at the time of leaf initiation and expansion; amounts of sunlight and minerals are perhaps the most important factors.
Despite numerous possibilities for variation within a species, leaf shape is a valuable tool for plant identification: Overall shape, including leaf base and apex, is important, as is the margin. The margin may be entire (smooth), toothed, lobed, or otherwise modified (FIGURE 6-12).
Within a leaf are veins or bundles of vascular tissue. These distribute water from the stem into the leaf and simultaneously collect sugars produced by photosynthesis and carry them to the stem for use or storage elsewhere. In basal angiosperms and eudicots, they occur in a netted pattern called reticulate venation (FIGURE 6-13 and FIGURE 6-14). In monocots with long, strap-shaped leaves, the larger veins run side by side with few obvious interconnections: This is parallel venation (FIGURE 6-15 and FIGURE 6-16).
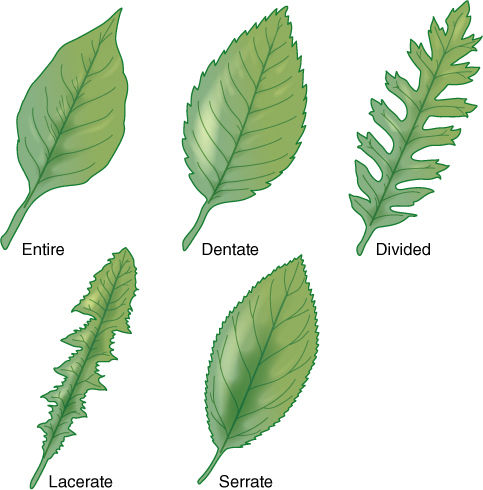
FIGURE 6-12 Several common types of leaf margins; in nature, hundreds of types exist, many being intermediate between two or among several other types.

FIGURE 6-13 A fig leaf (Ficus) held up to the light reveals five prominent main veins, numerous lateral veins and the reticulate venation of all smaller veins in this eudicot leaf; no part of the leaf is without veins.
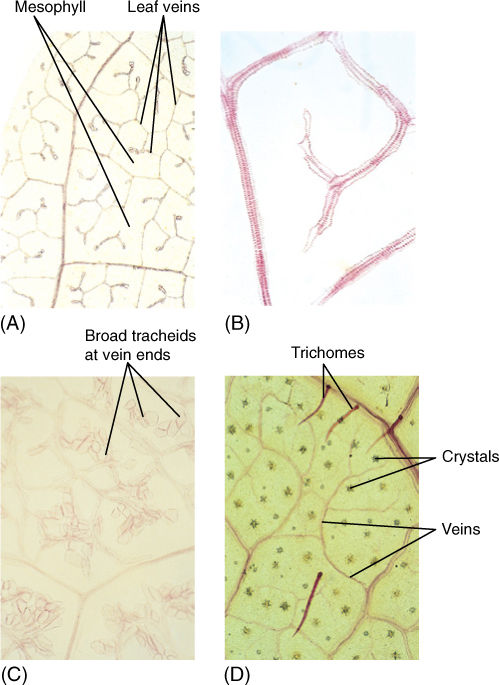
FIGURE 6-14 These leaves were treated (cleared) to make their cells transparent; then xylem was stained to make leaf veins visible. (A) In passionflower (Passiflora) leaves, veins are narrow and highly branched. At this magnification (×40), the height of the photograph represents 1 mm, and veins occur close together. Leaf veins typically occur closer together than do capillaries in our bones, tendons, and cartilage. (B) Higher magnification of privet leaf. At any point, minor veins are only two or three tracheary elements wide (×150). (C) These minor veins of crown of thorns (Euphorbia millii, a species that grows in deserts) are broad and have many wide tracheids at the ends (×50). (D) Acalypha has numerous crystals within the leaf and trichomes on the leaf surface (×80).

FIGURE 6-15 Cattails (Typha latifolia) have long, strap-like leaves common in monocots. Vascular bundles run parallel to each other from leaf base to tip.
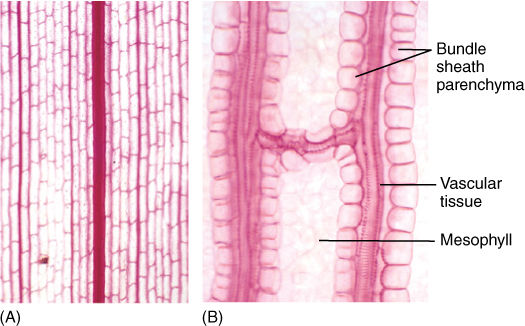
FIGURE 6-16 (A) Leaf clearing of corn (Zea), a monocot, with several sizes of longitudinal, parallel veins. If a longitudinal vein is broken by insect damage or any other problem, conduction can detour around the site by means of the fine transverse veins (×50). (B) Higher magnification showing the parenchyma cells that surround the leaf bundles in corn (×250).
At the leaf base, usually in the petiole, is an abscission zone oriented perpendicular to the petiole; its cells are involved in cutting off the leaf when its useful life is over (FIGURE 6-17). In autumn, as deciduous leaves begin to die, abscission zone cells release enzymes that weaken their walls. As the leaf twists in the wind, cell walls break, and the leaf falls off. Adjacent undamaged cells swell and become corky, forming protective scar tissue, the leaf scar, across the wound. Without an abscission zone, dead leaves might tear off irregularly, leaving an open wound vulnerable to pathogens. In many monocots, ferns, and cycads, an abscission zone is not formed; instead, a protective corky layer protects living stem tissues, but the dead leaf persists on the plant until it gradually decomposes (FIGURE 6-18).
![]() Internal Structure of Foliage Leaves
Internal Structure of Foliage Leaves
Epidermis
Flat, thin foliage leaves (optimal for light interception) have a large surface area through which water can be lost. Water loss through the epidermis is called transpiration and is a serious problem if the soil is so dry that roots cannot replace lost water. The epidermis must be reasonably waterproof but simultaneously translucent, and it must allow entry of carbon dioxide. Leaf and stem epidermises are basically similar, consisting of a large percentage of flat, tabular (shaped like paving stones), ordinary epidermal cells; guard cells and trichomes (either glandular or nonglandular) may be abundant. The dorsiventral nature of leaves, however, causes their upper and lower epidermis to exist in significantly different microclimates. On a sunny day, a leaf is usually warmer than surrounding air; thus, it heats the air and convection currents rise from it. If stomata in the upper epidermis are open and losing water, the water molecules are swept away by this convection. However, air tends to be trapped on the underside of a leaf, so water loss from stomata there is not so great: Water molecules are trapped in quiet air and may diffuse back into the stomata. In most leaves, the number of stomata per square centimeter is much greater in the lower than in the upper epidermis; in many species, no stomata at all are found in the upper epidermis (TABLE 6-2).
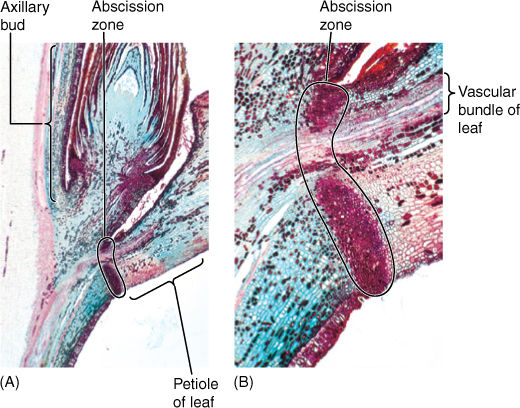
FIGURE 6-17 (A) Longitudinal section through a leaf axil, showing the abscission zone in the petiole and the axillary bud just above (×15). (B) Magnification of the abscission zone while the leaf is still healthy. The xylem and phloem are not interrupted until the leaf actually falls away (×80).

FIGURE 6-18 As leaves of Joshua tree (Yucca brevifolia) age, they flex downward and do not abscise; this creates a sheath of tough, fibrous dead leaves with many sharp spines that prevent rats, mice and other small animals from climbing the plant and eating young leaves and buds.
TABLE 6-2 Frequency of Stomata in Leaf Upper and Lower Epidermis
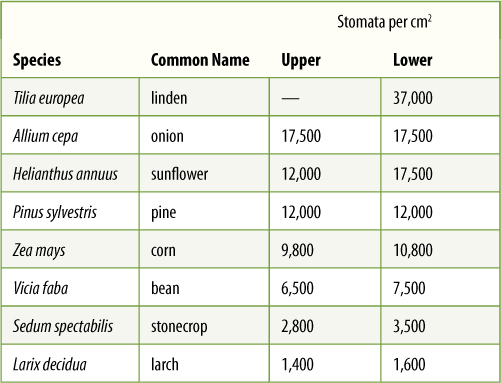
Such unilateral distribution of stomata has other beneficial consequences. Air-borne spores of fungi and bacteria are continually landing on leaves. Rye leaves, for example, typically have 10,000 fungal spores per square centimeter, but most are on the upper surface where stomata are rare; thus, few fungi can penetrate the leaf.
Leaf stomata are frequently sunken into epidermal cavities that create a small region of nonmoving air. In oleander, the lower leaf epidermis contains numerous crypts (areas where the epidermis is depressed into the leaf), and stomata and trichomes are abundant in the epidermis that lines the crypts (FIGURE 6-19). Both the recessed nature of the crypt and the presence of trichomes keep air in the crypt extremely quiet; many water molecules that diffuse out of open stomatal pores bounce around the crypt and re-enter the stomata rather than being blown away.
Leaf epidermises are often remarkably hairy, and trichomes affect leaf biology in numerous ways. What are some of the consequences of having trichomes on leaves? They provide some shade on the upper surface of the leaf, deflecting excessive sunlight, an adaptation common in desert plants (FIGURE 6-20). On the lower surface, they prevent rapid air movement and slow water loss from stomata (Figure 6-2). In any position, trichomes make walking or chewing difficult for insects, and many glandular trichomes secrete powerful stinging compounds that prevent even large animals from eating the leaf. Insects either do not bother with the leaf or must walk so slowly on it that they become more vulnerable to their own predators. Conversely, hairs provide excellent footholds against leaf flutter for insects of the appropriate size. Poisonous glandular trichomes in stinging nettle and other species prevent mammals from eating leaf tissue but also protect leaf-borne insects small enough to walk, feed, and reproduce safely between the stinging trichomes.
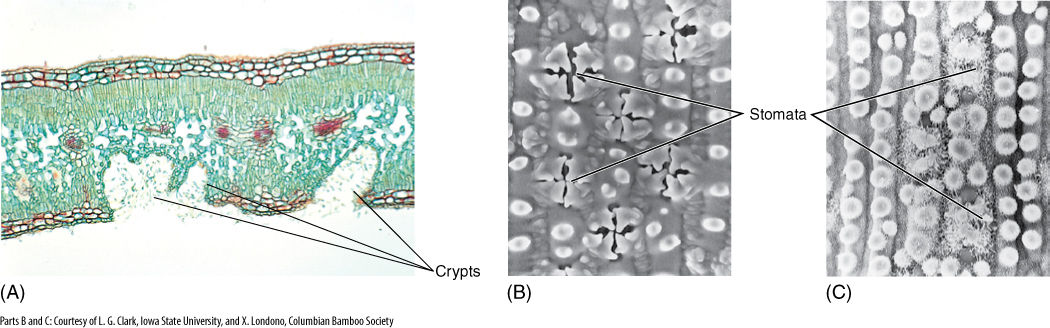
FIGURE 6-19 (A) Oleander (Nerium oleander) leaves have stomatal crypts filled with trichomes and stomata. This minimizes air movement near stomata, and water molecules that diffuse out of the leaf may re-enter it by random motion. Crypts occur only in the lower surface (×80). (B) Each stoma of this grass (Elytrostrachys) is overarched by four protrusions from surrounding cells. This reduces air movement near the stoma (×940). (C) In this bamboo (Chusqua), stomata are protected by protrusions and wax rods (×940). In all three examples, the trichomes and waxes prevent water droplets from blocking the stomatal pores.
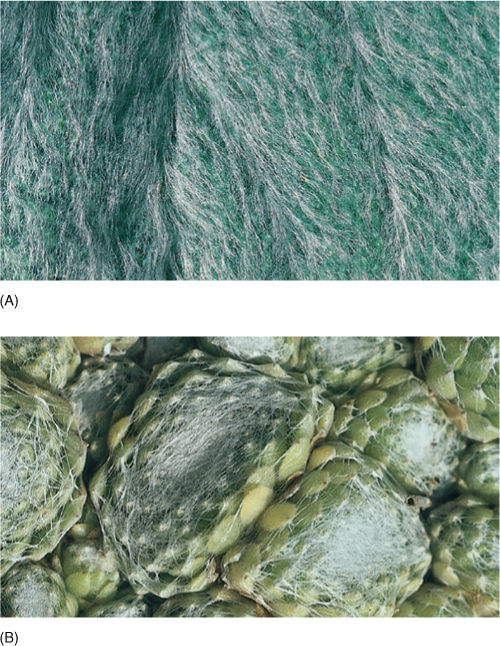
FIGURE 6-20 (A) Leaves of lamb’s ears (Stachys) are so densely covered with hairs that they often appear white. The hairs reduce the amount of heat these leaves absorb from sunlight. (B) Plants of Sempervivum are native to high alpine regions where ultraviolet radiation is intense. The hairs of the leaf epidermis reflect part of the ultraviolet radiation away from the plant.
Like stem epidermis, leaf epidermal cells contain a coating of cutin and usually also wax on their outer walls. These retain water and make digestion by fungi difficult. Their smooth, slippery surface prevents spores from sticking or allows them to be washed off by rain.
In areas of dew and fog, water droplets condense on trichomes rather than on the epidermis surface; this prevents a film of water from forming across the stomatal pores, which would block movement of carbon dioxide and oxygen until the water evaporated.
Mesophyll
The ground tissues interior to the leaf epidermis are collectively called mesophyll (FIGURE 6-21 and FIGURE 6-22). Along the upper surface of most leaves is a layer of cells, the palisade parenchyma (also called palisade mesophyll), which is the main photosynthetic tissue of most plants. Palisade cells are separated slightly so that each cell has most of its surface exposed to the intercellular spaces. Because carbon dioxide dissolves into cytoplasm slowly, the large surface gives maximum area for dissolution; tightly packed cells could not absorb enough carbon dioxide for efficient photosynthesis (Figure 6-22). Palisade parenchyma is often only one layer thick, but in regions with intense, penetrating sunlight, it may be three or four layers thick (FIGURE 6-23).
In the lower portion of the leaf is the spongy mesophyll—open, loose aerenchyma that permits carbon dioxide to diffuse rapidly away from stomata into all parts of the leaf’s interior (Figures 6-21 and 6-23). If stomata were surrounded by closely packed cells, a molecule of carbon dioxide might simply bounce off a cell and escape back out the stomatal pore.
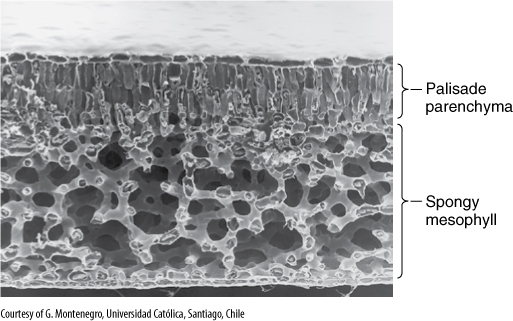
FIGURE 6-21 This leaf of Laurelia has a layer of palisade parenchyma that is two cells deep, and the spongy mesophyll is an extensive aerenchyma (×180).
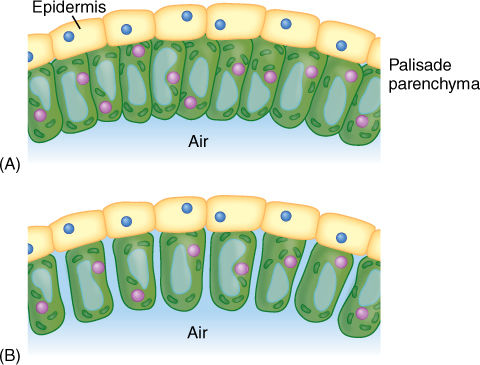
FIGURE 6-22 (A) If palisade cells are closely packed with no space between them, only their bottoms are exposed to the carbon dioxide in the spongy mesophyll. (B) With only slight separation of the cells, the volume of the palisade is almost unchanged, but the surface area available is increased enormously; carbon dioxide can be absorbed much more rapidly. Assume that the palisade parenchyma cells are rectangular columns 20 × 20 × 100 μm long; compute the surface area exposed to CO2 by 100 cells arranged as in (A) versus (B).
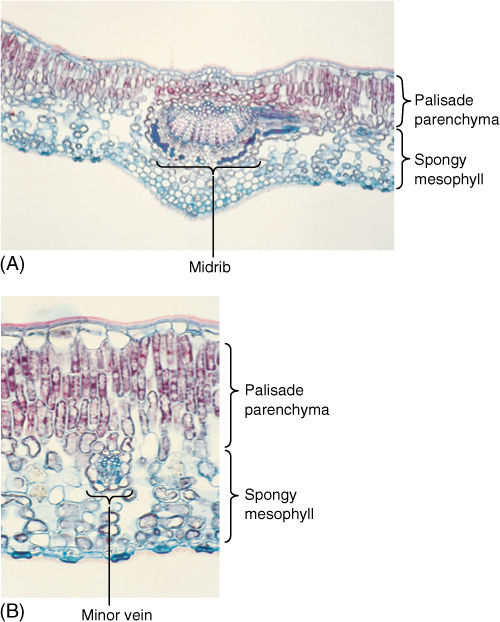
FIGURE 6-23 (A) and (B) Leaves of holly (Ilex) have an extremely thick palisade parenchyma consisting of at least three layers of columnar cells. Cells of the lowest palisade layer are relatively widely separated, permitting rapid diffusion of CO2 from spongy mesophyll to upper palisade (A) ×50; (B) ×150).
Although this arrangement is the most common, some plants have a layer of palisade parenchyma along both leaf surfaces; spongy mesophyll either occurs in the center or is lacking (FIGURE 6-24). The relationship between leaf position and the sun is important: For leaves that are held horizontally (most leaves), the sun is usually overhead, so having palisade parenchyma near the upper surface permits maximum absorption of light and photosynthesis. For plants that hold their leaves vertically (Iris, Gladiolus, and Eucalyptus), both sides are equally illuminated, and palisade parenchyma is equally functional on either side.
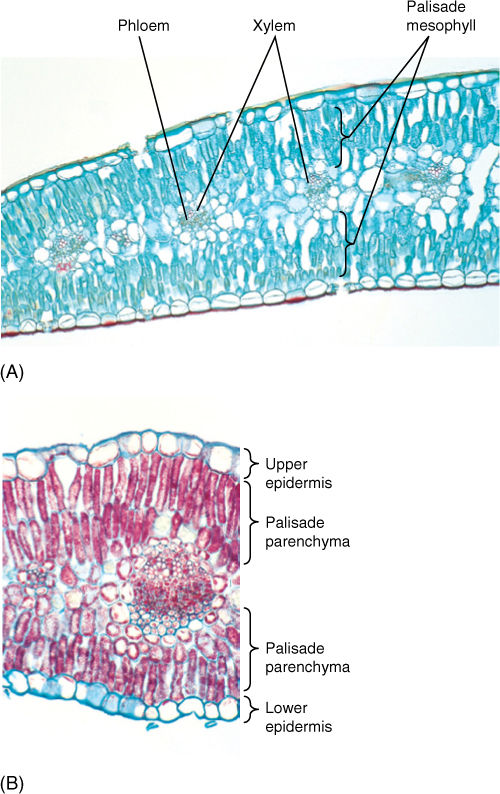
FIGURE 6-24 Leaves of both carnation (A) and creosote bush (B) have palisade parenchyma along both surfaces. The little spongy mesophyll present is located in the middle of the leaf (A) ×75; (B) ×150).
Vascular Tissues
Between the palisade parenchyma and spongy mesophyll are vascular tissues. A eudicot leaf usually has one large midrib, also called a midvein, from which lateral veins emerge that branch into narrow minor veins (see Figures 6-13 and 6-14). Minor veins are most important for releasing water from xylem and loading sugar into phloem, whereas the midrib and lateral veins are involved mostly in conduction. Vein structure changes with size: The midrib and lateral veins always contain both primary xylem on the upper side and primary phloem on the lower side (FIGURE 6-25). Because they both conduct and support the leaf blade, they may have many fibers arranged as a sheath, called a bundle sheath, around the vascular tissues (FIGURE 6-26). A sheath also makes it difficult for insects to chew into the vascular tissues. Many other types of nonconducting cells such as mucilage, tannin, or starch storage cells may be present in larger veins. Minor veins are the sites of material exchange with the rest of the mesophyll and must have a large surface area in contact with palisade and spongy mesophyll; they do not contain fibers or other nonconducting cells whose presence would interrupt this contact (FIGURE 6-27). The endings of the minor veins in some species consist of only xylem and in others only phloem, but they typically contain both.
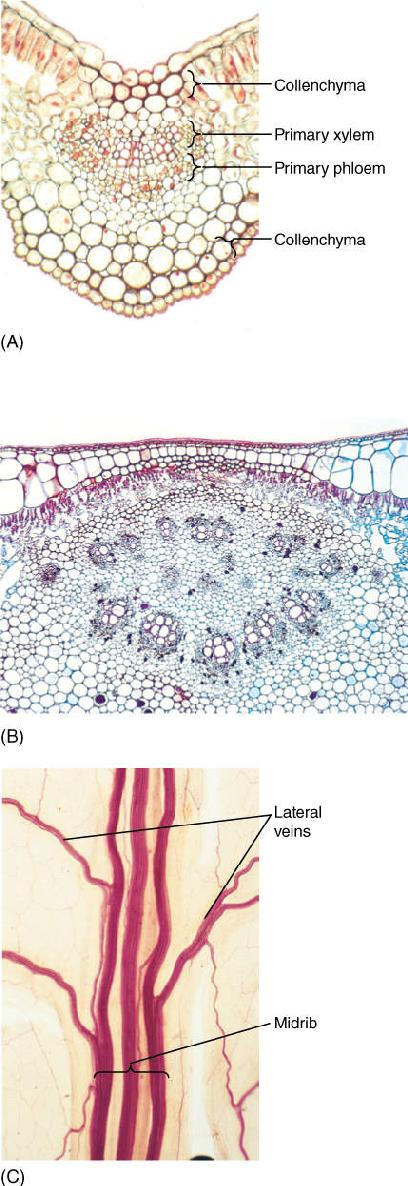
FIGURE 6-25 Transverse sections through leaf midribs. (A) This midrib of Ligustrum is rather simple, containing a single vascular bundle. Xylem is present along the top, phloem along the bottom. A small amount of collenchyma occurs on the top and bottom of the midrib (×150). (B) This midrib of rubber tree has numerous separate bundles embedded in a large mass of mesophyll (×50). (C) This is a midrib prepared as a leaf clearing (similar to Figure 6-14); the three distinct bundles have no interconnections in this region. Each transports to and from a particular portion of the blade (×15).
Veins, especially larger ones, often have a mass of fibers above, below, or both—the bundle sheath extension (Figure 6-26). Such fibers help give rigidity to the blade and are believed to provide an additional means by which water moves from the bundle out to the mesophyll. Apparently, water moves by capillary action around the fibers rather than through them.
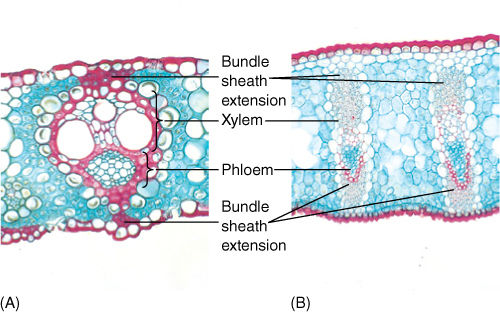
FIGURE 6-26 (A) This leaf bundle of sugar cane has bundle sheath extensions contacting both the upper and lower epidermis. Water diffuses from the xylem to the epidermis by capillary action, moving around the surfaces of the fibers (×100). (B) The leaves of flax have large bundle sheath extensions consisting of many layers of fibers (×45).
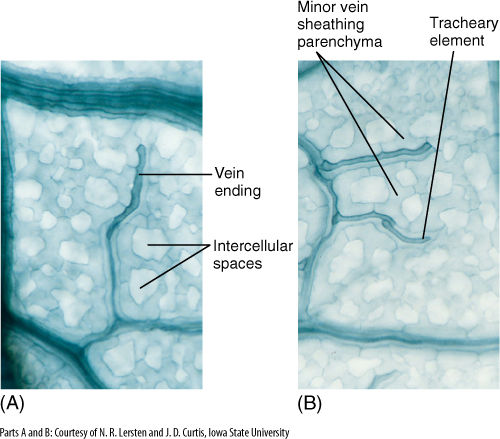
FIGURE 6-27 (A) and (B) These are minor veins, treated to reveal individual cells. Long, dark cells are tracheary elements, and phloem is out of focus, in line with the xylem. Notice how narrow these veins are: Xylem is only one element wide, so there is maximum surface area for releasing water to the leaf mesophyll. In this species, each minor vein is lined with parenchyma cells that extend outward into the spongy mesophyll (×250).
Petiole
Petioles may be tiny but are massive in plants like palms, rhubarb, celery, and water lilies. They are considered to be part of the leaf and are the transition between the stem and the lamina; consequently, the arrangement of tissues differs at the two ends. The epidermis may be similar to that on the lamina but often contains fewer stomata and trichomes. Petiole mesophyll is rather like cortex—somewhat compact and not especially aerenchymatous—and considerable collenchyma is present if the petiole supports a heavy lamina. Vascular tissues are the most variable; one, three, five, or more vascular bundles, called leaf traces, branch from stem vascular bundles and diverge toward the petiole (FIGURE 6-28A). They may remain distinct or fuse into a single trace at, near, or in the petiole. In some species, they divide into numerous bundles, and 10 or 20 (in large palm leaves, several hundred) may be found in the petiole (FIGURE 6-28B). Vascular bundles may either fuse with each other within the petiole or branch further; they can be arranged in a ring, a plate, or a number of other patterns. If the lamina has a strong midrib, most petiole bundles fuse together and form the midvein, but other bundles may enter the lamina as small lateral veins.
We do not understand the significance of all the bundle patterns in petioles. Certain patterns may ensure the proper distribution of sugars out of different parts of the lamina, especially of large leaves, into the various bundles of the stem. However, it may be that almost any pattern functions well, and thus, mutations that cause new patterns are not selectively disadvantageous. On the other hand, future studies may show correlations between lamina, petiole, and stem.
In many species, the petiole bears two small flaps of tissue at its base called stipules, which serve various functions. They may protect the shoot apical meristem while the leaf is young and small. In other plants, they are large enough to contribute a significant amount of photosynthesis, but usually, when the leaf is mature, the stipules are still small, and they die early.
![]() Initiation and Development of Leaves
Initiation and Development of Leaves
Basal Angiosperms and Eudicots
Leaves are produced only through the activity of a shoot apical meristem. At the base of the meristem, cells just interior to the protoderm grow outward, forming a protrusion known as a leaf primordium (FIGURE 6-29) that extends upward as a narrow cone, growing so rapidly that it becomes taller than the shoot apical meristem. During this stage, the primordium consists of leaf protoderm and leaf ground meristem, and all cells are meristematic, with dense cytoplasm and small vacuoles. A strand of cells in the center differentiates into provascular tissue and then into primary xylem and phloem, forming a connection with the young bundles in the stem.
Botany and Beyond
BOX 6-2 Leaf Structure, Layer by Layer
The internal organization of ordinary leaves is not very complicated but it can be difficult to visualize from looking only at transverse sections. A series of sections through a leaf of privet (Ligustrum) is presented here. These are paradermal sections; that is, they are parallel to the epidermis. The first is at the level of the palisade parenchyma, and each successive section is deeper in the leaf. Try to imagine the appearance of the leaf from the perspective of a carbon dioxide molecule.
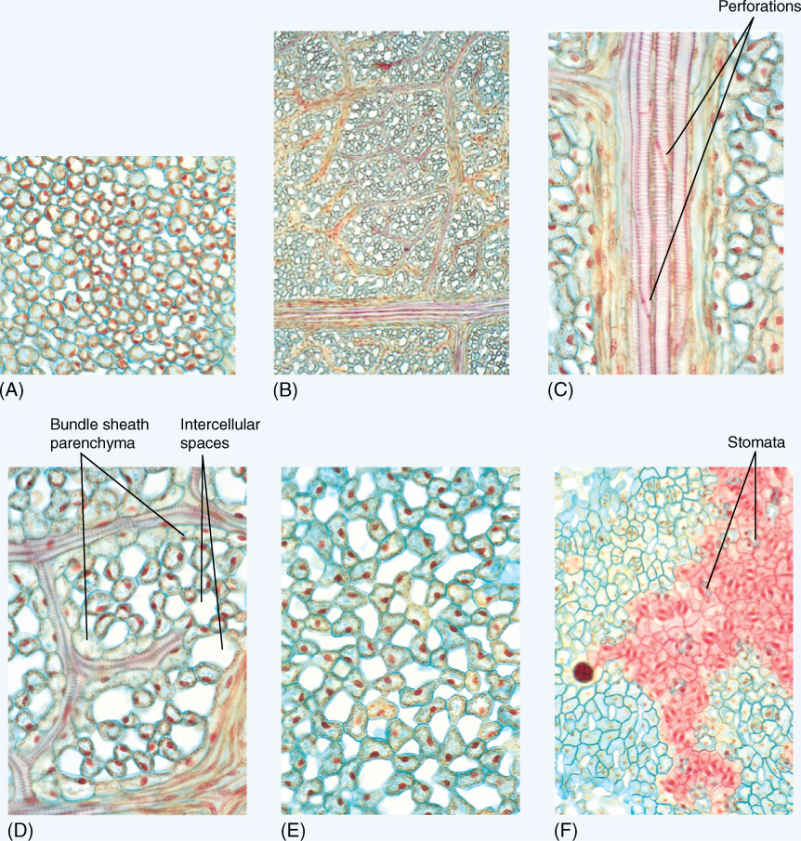
FIGURE B6-2 (A) Palisade parenchyma (×250). The cells are cylindrical and are not closely packed; carbon dioxide diffuses easily throughout this layer and is absorbed into the cells through their large surface area. (B) Leaf veins (×50). Just below the palisade parenchyma lies the network of leaf veins. Small lateral veins branch off of larger veins and in turn give off smaller veins. (C) High magnification of a large lateral vein (×250). This section passes through the xylem of the vein, and perforations are visible; thus, these are vessel elements, not tracheids. These have spiral thickenings: They could differentiate while the leaf was still small, and then as the leaf expanded, they could be stretched and still conduct water. (D) High magnification of minor veins (×250). All mesophyll cells are close to one vein or another. After sugar has been produced by photosynthesis, it does not diffuse very far before it is picked up by a sieve tube member and transported to the stem. (E) Spongy mesophyll (×250). In many leaves, more than half the spongy mesophyll consists of intercellular air space. This allows carbon dioxide molecules to diffuse rapidly into the leaf, away from the stomata. If spongy mesophyll were more dense, carbon dioxide molecules might be trapped near the stomata, then diffuse back out of the leaf before they could be absorbed by a chlorenchyma cell. (F) Lower epidermis (×150). The epidermis is slightly undulate, and as it was cut to make this slide, the knife passed through the center of the cells in some areas (green) and through the outer wall and cuticle in other areas (red). Stomata are abundant, but the actual stomatal pore is short and narrow; each is only a small opening through which carbon dioxide can enter.
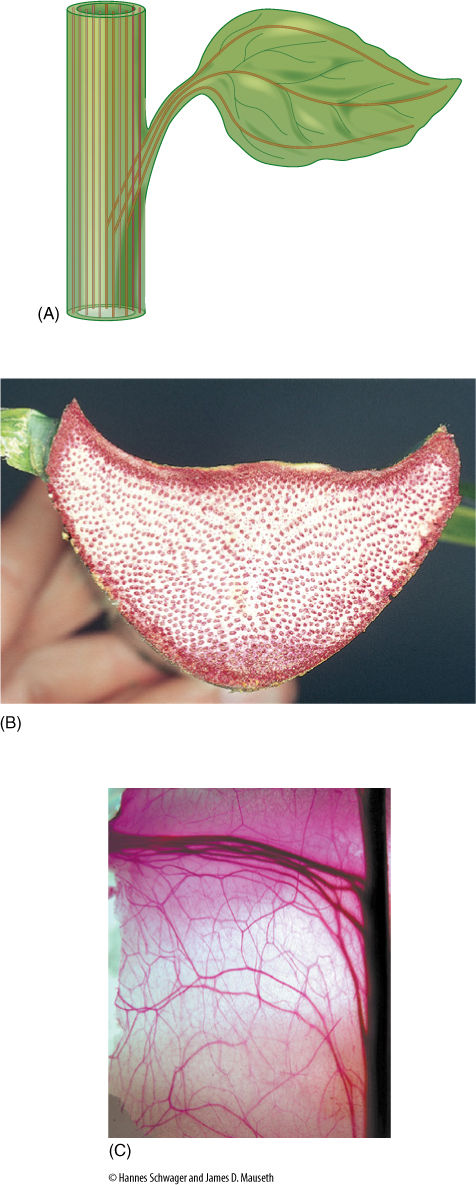
FIGURE 6-28 (A) This section of stem has 11 bundles, 3 of which produce leaf traces that enter the petiole. Later, the leaf traces diverge and spread throughout the blade. These traces are drawn without any branching or merging, although both are possible. (B) Cross-section of a petiole of the palm Attalea stained to show its many vascular bundles. Notice how numerous the bundles are compared with the leaves of Figure 6-25. This leaf, like those of most palms, is very large and must have a massive conduction system. (C) In cacti, traces that run to leaves and axillary buds are complex partly because the cortex is so thick the traces must be long, and partly because leaf and branch traces divide and bifurcate, forming an extensive set of cortical bundles that vascularize the thick, succulent cortex. The central stele of the cactus is the dark material on the right; the dark, thick traces at the top of the micrograph are leaf and bud traces that lead to a flower, and all the others are cortical bundles (×3). Hylocereus undatus.
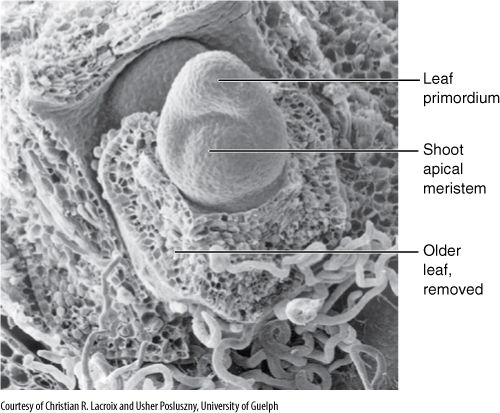
FIGURE 6-29 The shoot apical meristem of grape (Vitis riparia) can be seen in three dimensions in this scanning electron micrograph. Notice the large leaf primordium; an older leaf primordium was removed; the phyllotaxy is established at the very apex (×200).
As the leaf primordium grows upward, it increases in thickness, establishing the bulk of the midrib. A row of cells on either edge of the primordium grows outward, initiating the lamina (FIGURE 6-30). As a result of their activity, the young leaf consists of a midrib and two small, thin wings. All cells in the wings are meristematic, and their division and expansion enlarge the lamina rapidly (FIGURE 6-31). In a compound leaf, two rows of loci initiate leaflets, which then grow like simple leaves. During lamina expansion, stomata, trichomes, and vascular bundles differentiate, and the petiole becomes distinct from the midrib. The entire pattern is completed while the leaf is still very small, much less than one tenth its mature size.
In many perennial plants, leaves are initiated in the summer or autumn before they mature. After they reach the developmental state just described, they become dormant, part of a resting terminal or axillary bud. During the next growth period, usually the following spring, the bud opens and the primordial leaves expand rapidly as each cell absorbs water into its vacuole and swells (FIGURE 6-32). Little or no mitosis or cell division may occur—only maturation, especially the synthesis of chlorophyll, cutin, and wax. As the leaves expand, the immature, exposed epidermis is very vulnerable, especially to insects (FIGURE 6-33). In many species of annual plants, the process is similar, with the embryo in the seed acting like the buds of perennial plants. They initiate leaves before the seed becomes dormant and dry, while it is still inside a developing fruit, and those leaves absorb water and expand rapidly during germination.

FIGURE 6-30 (A) The uppermost leaf primordium is the oldest, and its sides have begun growing out as the lamina of a simple leaf of grape (×270). (B) In this compound leaf, rather than forming a lamina, six leaflet primordia have been established (×150).
Monocots
Monocot leaves, like those of eudicots, are initiated by the expansion of some shoot apical meristem cells to form a leaf primordium (FIGURE 6-34). Apical meristem cells adjacent to the primordium grow upward along with it, becoming part of the primordium and giving it a hood-like shape. More apical meristem cells become involved until the primordium is a cylinder that completely or almost completely encircles the shoot apical meristem. This tubular portion grows upward as a sheathing leaf base, and the original conical leaf primordium, now located on one side of the top of the tube, gives rise to the lamina. The outer surface of the tube is abaxial epidermis, the inner surface is adaxial epidermis. Meanwhile, the shoot apex enlarges, forms new stem tissue, and initiates the next leaf primordium, which will develop as a tube inside the previous leaf’s sheathing base. After the first leaf has grown to its full size, the next leaf continues elongating and emerges through the opening at the top of the previous leaf’s tubular sheathing base. Later, another, younger leaf will emerge and so on.
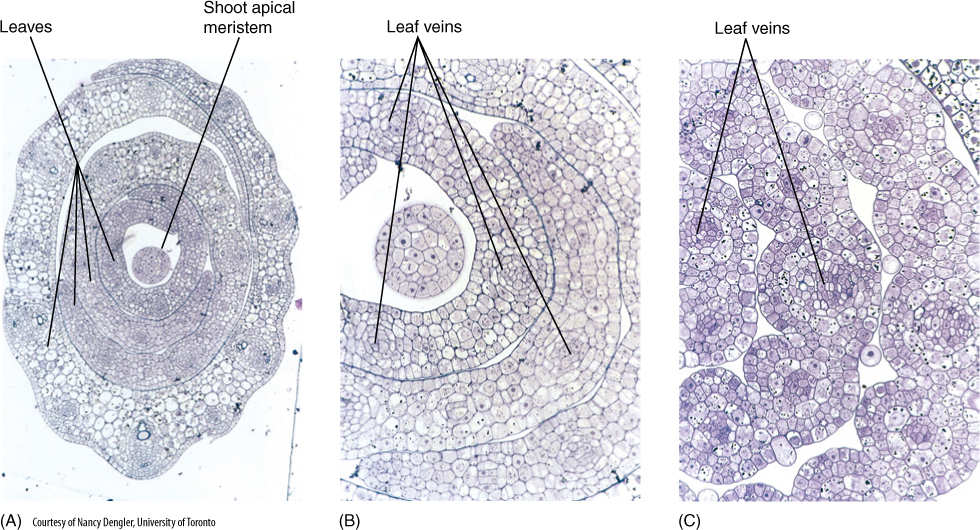
FIGURE 6-31 (A) Low-magnification transverse section through young, developing leaves of the grass Panicum effusum. The central round structure is a section of the shoot apical meristem, and the leaves encircle it (grasses have sheathing leaf bases) (×200). (B) At higher magnification, you can see that all cells in the leaves are still small and cytoplasmic, and the central vacuoles are small. At this stage, all cells are still undergoing mitosis and cytokinesis. Veins are starting to differentiate (×500). (C) At an older stage, leaf veins are more distinct. The first protoxylem tracheary elements are mature in some veins, but most cells that will differentiate into tracheary elements are still enlarging (×500).

FIGURE 6-32 Buds of buckeye (Aesculus arguta) expanding in the spring. Bud scales are being pushed back and will abscise as new foliage leaves swell. The smaller bud on the right contains only leaves, and its apical meristem may be initiating new leaf primordia that will expand a little later. The larger bud on the left has both new leaves as well as flower primordia (the small ball-like structures).
In some monocots, the lamina becomes broad and expanded like a eudicot lamina, but grasses, lilies, and many others have linear, strap-shaped leaves that grow continuously, having no predetermined size. Their lamina grows by a meristem located at its base where it attaches to the top of the sheathing leaf base. The lamina’s meristematic cells remain active mitotically, producing new cells that extend the leaf. Even if a grazing animal, a range fire, or a lawnmower destroys much of the leaf, the meristem forms more lamina. This type of regeneration is not possible in most leaves.
The constant basal expansion in monocot leaves means that protoxylem and protophloem are constantly being stretched and disrupted in the basal meristem. New vessel elements and sieve tube members differentiate rapidly enough that conduction is never interrupted. Just above the basal meristem is a region where tissues differentiate: More primary xylem and phloem are initiated, as are stomata and other features. Higher above the basal meristem, tissues are mature; differentiation is similar to that for stems but is oriented upside down in monocot leaves.

FIGURE 6-33 Aphids and other sucking insects typically attack young leaves and stems that have not yet developed any protective sclerenchyma around the vascular bundles.
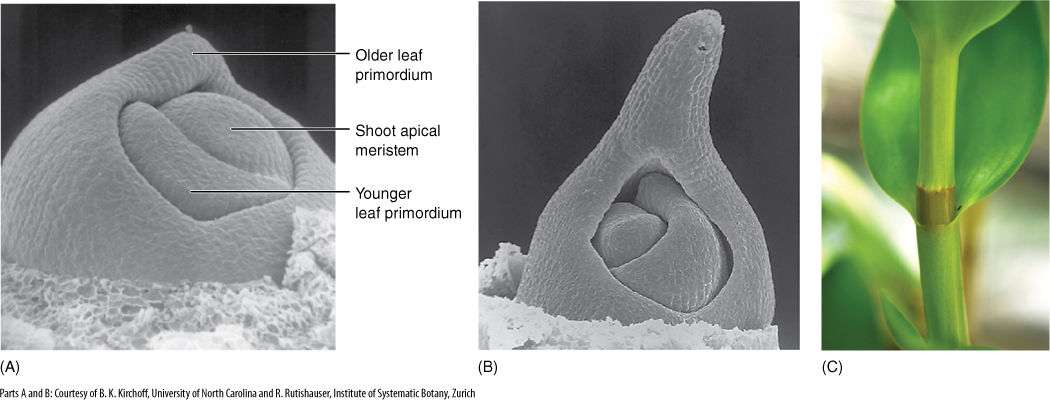
FIGURE 6-34 In a monocot, after the center of the leaf primordium has been initiated (A), neighboring regions of the shoot apical meristem become active and contribute to the primordium. This results in (B) a leaf base that encircles the stem, forming a sheath around it (A)×150; (B) ×120). (C) The leaf blade of this Dendrobium orchid has developed from a leaf primordium tip that resembled the tip in (B), and the sheathing base here developed from elongation growth of the small collar of leaf primordium tissue visible in (B).
Alternatives
BOX 6-3 Photosynthesis Without Leaves
Photosynthesis may occur in tissues other than foliage leaf mesophyll, for example, stem cortex and bark. Carrying out photosynthesis in these alternative sites has particular consequences.
Bark. A small number of trees produce green, chlorophyllous photosynthetic bark. This is rare and occurs almost exclusively in desert-adapted trees that abscise their leaves during dry periods, after which they must rely entirely on their bark for sugar production. The trees, often called palo verde (Spanish for green stick), produce cork cells in their bark like all trees do, but they also produce cells called phelloderm, which contain chloroplasts (FIGURE B6-3A). Phelloderm is usually rather compact with few intercellular spaces, and very little of the cell walls are “free surface,” that is, surface that faces intercellular space and that can absorb carbon dioxide. Consequently, carbon dioxide neither diffuses rapidly through phelloderm nor is it absorbed quickly. Furthermore, the layer of phelloderm cells is covered by several layers of cork cells, which are impermeable to gases and thus block uptake of carbon dioxide. The green bark of these trees probably carries out “recycling” photosynthesis: All living cells of the tree trunk and branches respire and give off carbon dioxide just like we animals do, and the green bark can recapture it and convert it back to carbohydrate. In trees with nonphotosynthetic bark, that carbon dioxide would simply leak out of the bark and be lost.
Cortex. Young stems of almost all plants have a chlorophyllous cortex. Most stems are too narrow to provide much surface area and thus capture little light. Whereas stomata are virtually always present in foliage leaf epidermis, they may be lacking in stem epidermis, and if so, little carbon dioxide can be absorbed. Even if stomata are present, stem cortex—like phelloderm—is typically more compact than leaf mesophyll and thus does not permit rapid diffusion or uptake of carbon dioxide.
Stem-succulent plants. Many desert plants store water in enlarged, chlorophyllous stems. Stem-succulents may produce large, thin, ordinary foliage leaves during the deserts’ brief moist period, abscising these leaves when drought begins (these are “drought-deciduous” plants), or they may produce only very tiny leaves (often incorrectly said to be leafless). Either way, stem-succulent plants usually depend on their chlorophyllous cortex for much or all their photosynthesis, and their cortex typically has many leaf-like features. Their stem epidermis has a high density of stomata, sometimes having as many stomata per square centimeter as leaf epidermis. The outermost layer or two of cortex cells may be very aerenchymatous, creating a tissue similar to spongy mesophyll, allowing carbon dioxide to diffuse rapidly away from stomata and deeply into the cortex. Chlorophyllous cortex often resembles palisade mesophyll, consisting of long columnar cells. Whereas palisade mesophyll is only rarely more than one or two cell layers thick, palisade cortex in stem succulents is often five to ten cell layers thick, being as much as 1.0 to 1.5 mm thick (about three to five times more than the entire thickness of a typical foliage leaf).
Any photosynthetic tissue must be well-vascularized, but cortex typically does not have its own set of vascular bundles. This vascular bottleneck can be overcome by several alternative arrangements. If the cortex is narrow, its chlorenchyma cells are close to the stem’s vascular bundles. If the stem has short internodes, then leaf traces are also close to each other and provide some vascularization to the cortex as they run out to the petiole base. The cortex of cacti is exceptional in that it does have its own particular vascular system called cortical bundles. This is a network of collateral vascular bundles that branch and anastomose forming a three-dimensional reticulate venation. Cactus cortex is well vascularized, and no matter how rapidly it photosynthesizes, phloem cells are close by and can quickly load the sugars and transport them away.

FIGURE B6-3 (A) Trunks and branches of trees called “palo verde” (green stick; Cercidium floridum) are green because their bark contains cells with chlorophyll. No stomata are present, so no new carbon dioxide can be absorbed from the air. (B) Trunks and branches of saguaro cactus (pronounced sa-WA-row there is no “g” sound at all) are green because the epidermis is still present and transparent, allowing us to see the green of the cortex cells inside. Stomata are present, and saguaros (Carnegiea gigantea) do bring in carbon dioxide from the air and convert it to carbohydrate. As a saguaro absorbs water after a rain, its parenchyma cells swell so the entire stem swells; the ribs flatten out and the epidermis is not torn. If it does not rain for many months, the cactus gradually loses water, despite its cuticle and wax, and shrinks; the ribs also shrink and become narrower (think of an accordion letting out air). A ribbed body like this can change volume without changing surface area.
Broad succulent stems store water, which causes them a problem not usually faced by foliage leaves: how to allow volume to increase while surface area remains constant. We animals generate new skin anywhere, anytime, which is a good thing if we become overweight: our volume increases, as does our surface area, and our skin grows rather than tearing apart and revealing our insides. However, plants only generate new epidermis at shoot and root apices and in growing leaves and flowers. Stem succulents solve this problem by having an accordion-like, pleated surface (the pleats are called ribs). As water is stored and volume increases, ribs become wider; as water is lost, ribs become narrower. Volume changes dramatically but surface does not (FIGURE B6-3B).
In these alternative photosynthetic systems, water conservation is an important factor so none of these alternatives is free to evolve to be as effective at photosynthesis as an ordinary foliage leaf is. Most modifications that increase water conservation decrease photosynthesis and vice versa. In moist habitats, water conservation is of little importance, so modifications that increase photosynthesis will increase survival of the plants even though they cause plants to lose water more rapidly. In contrast, modifications that conserve water in dry habitats are more valuable than those that allow maximum photosynthesis. Of course, desert plants are not free to maximize water conservation to the point where they do not photosynthesize at all.
![]() Morphology and Anatomy of Other Leaf Types
Morphology and Anatomy of Other Leaf Types
Succulent Leaves
Numerous adaptations permit plants to survive in desert habitats, one of the most common being production of succulent leaves (FIGURE 6-35). This is characteristic of species in the families Crassulaceae (contains Kalanchoe and Sedum), Portulacaceae (contains Portulaca and Lewisia), and Aizoaceae (ice plant), among others. Succulent leaves are thick and fleshy, a shape that reduces the surface-to-volume ratio and favors water conservation. Some leaves are cylindrical or even spherical, the optimal surface-to-volume shape. The reduction in surface area, which is advantageous for water retention, has the automatic consequence of reducing the capacity for carbon dioxide uptake.
Inside the leaf, the mesophyll contains very few air spaces, reducing the internal evaporative surface area and, in turn, water loss through stomata. A lack of air spaces also makes the mesophyll more transparent, just as pure water is more transparent than soap bubbles or foam, allowing light to penetrate farther into the leaf. Photosynthesis occurs more deeply than it would in the foliage leaves described earlier. In some members of the genera Lithops (stone plants) and Frithia, leaves are so translucent they act as optical fibers; the leaves are located almost completely underground, where it is cool and relatively damp (FIGURE 6-36). The exposed leaf tips allow sufficient light to enter and be conducted to the subterranean chlorenchyma. Although the plants live in a harsh desert in Madagascar and southern Africa, photosynthesis actually occurs in a rather mild microclimate.

FIGURE 6-35 (A) Leaves of Senecio rotundifolia are spherical, giving them an optimal surface-to-volume ratio for conserving water. (B) Leaves of the succulent Dinteranthus are hemispherical and attached to the opposite leaf, greatly reducing the exposed leaf surface. Interior tissues are mostly water-storage parenchyma. Their low levels of chlorophyll result in low levels of photosynthesis, but a lack of water is a more significant danger for these plants. Each plant consists basically of two leaves and a microscopic stem; the root system (not visible) is substantial. Dinteranthus and many related species have only two leaves at a time; as two new leaves expand, the existing two collapse and wither away. (C) This agave (probably Agave parryi) has survived a wildfire in West Texas because its succulent leaves protected the stem from the heat of the fire. The center of the plant—and especially the shoot apical meristem—is still healthy. Containing so much water, the leaves heat up slowly, and before the innermost leaves became hot, wind had blown the fire farther on and the stem and apical meristem were saved. The leaves even kept the ground at the base of the plant cool, protecting the roots.

FIGURE 6-36 Lithops, stone plant, is closely related to Dinteranthus; both are in the family Aizoaceae. Lithops also has only two leaves, but they are located almost entirely underground. The flat, translucent tips project above the soil and conduct light to the subterranean chlorenchyma.
Sclerophyllous Foliage Leaves
Foliage leaves must produce more sugars by photosynthesis than are used in their own construction and metabolism, or the plant would lose energy every time it produced a leaf. This limits the amount of sclerenchyma in foliage leaves, and most leaves therefore tend to be soft, flexible, and edible.
In some species (barberry, holly, Agave, and Yucca), leaves have evolved which are perennial, existing on a plant for 2 or more years (FIGURE 6-37). With this extended lifetime and prolonged productivity, sclerenchymatous leaves are feasible, and their hardness makes them more resistant to animals, fungi, freezing temperatures, and ultraviolet light. Such plants are sclerophyllous and the leaves are sclerophylls. The sclerenchyma is often present as a layer just below the epidermis and in the bundle sheaths, although the epidermis itself can be composed of thick-walled cells (FIGURE 6-38). The cuticle is usually very thick, and waxes are abundant on leaves of many sclerophyllous species.

FIGURE 6-37 Leaves of barberry (Berberis) are tough and hard. It is difficult for insects to bite into them or lay eggs in them; however, the plant must invest considerable glucose to make the secondary walls of sclerenchyma cells, and the leaves must therefore photosynthesize longer before they reach their break-even point.
Leaves of Conifers
In almost all species of conifers, leaves are sclerophylls; they have a thick cuticle, and their epidermis and hypodermis cells have thick walls. Most conifer leaves contain unpalatable chemicals. Conifer leaves are always simple, never compound, and have only a few forms. Needles, either short or long, occur in all pines, firs, and spruces (FIGURE 6-39). Needles of longleaf pine can be 40 cm long, although most other species of pine have needles about 10 cm long. Small, flat, scale-like leaves form a shield-like covering on stems of junipers, cypresses (Cupressus), arborvitae (Thuja), and others. In Agathis, Araucaria, and Podocarpus, all genera of the Southern Hemisphere, leaves are rather large, broad scales held away from the stem (FIGURE 6-40). Lengths up to 12.5 cm and widths of 3.5 cm have been measured in leaves of Podocarpus wallichianus.
Conifer leaves are mostly perennial, remaining on the stem for many years; consequently, the plants are evergreens. Needles of bristlecone pine live for at least 5 years; their vascular bundles can produce new phloem each year, but no new xylem (FIGURE 6-41). The small scale leaves of juniper also persist and are photosynthetically active for many years, and leaves of Agathis and Araucaria remain even on very old trunks. Three conifers have annual leaves that are shed each autumn; larches (Larix), bald cypress (Taxodium), and dawn redwood (Metasequoia) are all deciduous.
Bud Scales
One of the most common modifications of leaves is their evolutionary conversion into bud scales (FIGURE 6-42). In perennial plants, dormant shoot apical meristems are protected from low temperatures and the drying action of wind during winter by bud scales, which form a tight layer around the stem tip. Because their role is primarily protection, not photosynthesis, bud scale structure differs from that of foliage leaves. Bud scales are small and rarely compound, so mechanical wind damage is not a risk for bud scales. Their petiole is either short or absent because they must remain close to the stem and be folded over it. To be protective, they must be tougher and waxier than regular leaves; bud scales frequently produce a thin layer of corky bark, at least on exposed portions, which provides greater protection than the simple epidermis of foliage leaves.
Spines
Cacti have two types of leaves. The green cactus body has microscopic green leaves, and the clusters of spines are their axillary buds: Cactus spines are modified leaves of axillary buds. The succulent, moist cactus body would be an excellent source of water for animals were it not for the protective spines (FIGURE 6-43). As with bud scales, spines have a distinct structure related to their function. The soft, flexible blade of a photosynthetic leaf is useless as a protective device against herbivores, but spines have no blade and are needle shaped; mutations that inhibit lamina formation have been selectively advantageous. No mesophyll parenchyma or vascular tissue is present; the mesophyll instead consists of closely packed fibers. After fibers mature, they deposit lignin in their walls, which makes them hard and resistant to decay. The cells then die and dry out, hardening even further. Because cacti carry out photosynthesis in their stem cortex, loss of the leaf lamina was not selectively disadvantageous, but for plants that have no alternative photosynthetic tissues, mutations that cause loss of lamina are extremely disadvantageous. Spines of many other species have a similar structure.
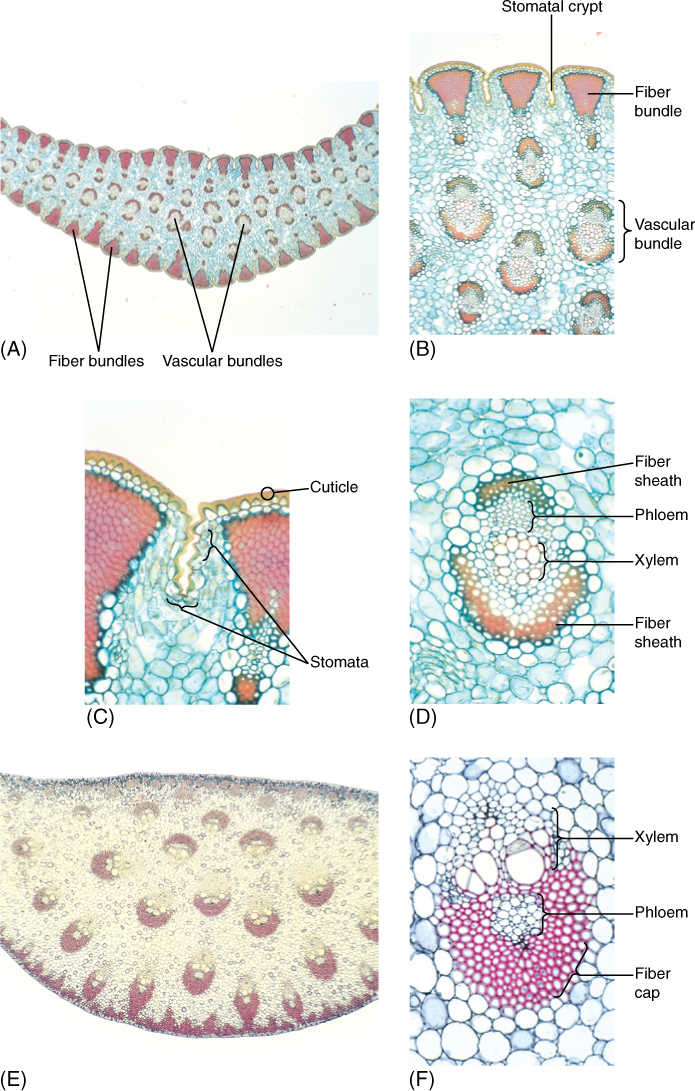
FIGURE 6-38 (A) Yucca leaves are excellent examples of sclerophylls; they are tough and fibrous, and the cuticle is thick. These features reduce the amount of light reaching the chlorenchyma, but plant growth is probably limited by scarcity of water rather than reduced photosynthesis. These leaves may function for 5 to 10 years or more (×15). (B) Bundles of fibers occur just interior to the epidermis, and the thick mesophyll surrounds many vascular bundles (×50). (C) Stomata occur in long, shallow grooves that run between fiber bundles (×150). (D) Even vascular bundles of these sclerophylls have fiber sheaths (×150). (E) Sclerophylls of Dracaena are very thick and have leaf veins throughout the mesophyll, unlike the single row in ordinary leaves (×15). (F) Leaf veins of Dracaena have extremely thick fiber sheaths (×150).

FIGURE 6-39 (A) Many conifers, such as this lodgepole pine (Pinus contorta), have needle-shaped leaves. (B) Incense cedar (Libocedrus) and many other conifers have scale-shaped leaves.

FIGURE 6-40 Scale-shaped leaves in monkey-puzzle tree (Araucaria araucana) are very large and persistent, living for many years, even as the branch enlarges and becomes woody.
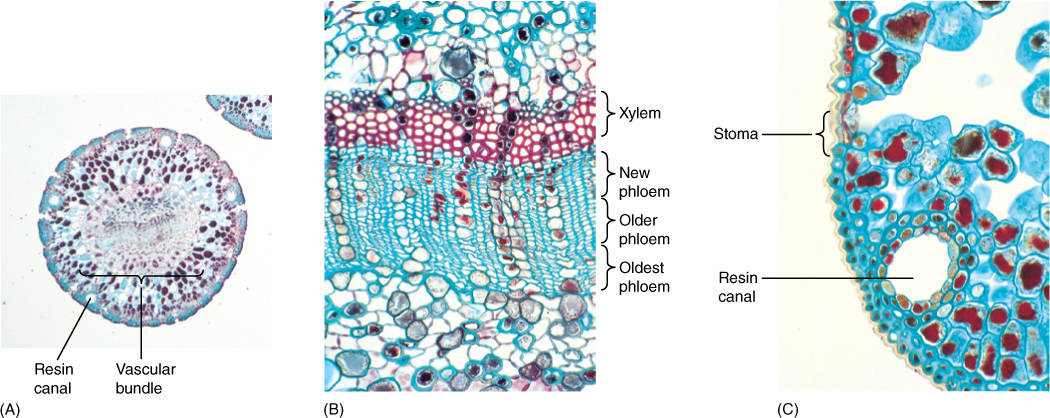
FIGURE 6-41 (A) Needle leaves of pine are round in transverse section (×15). (B) Needles last for many years, producing more phloem each year (×150). (C) Pine needles are sclerophylls: They have thick-walled epidermal cells with a thick cuticle. They also contain resin canals (×150).

FIGURE 6-42 (A) Bud scales of Rhododendron fit together tightly during winter, protecting the enclosed apical meristem and young leaves and flowers. (B) The bud scales of Magnolia have a dense covering of hair; this improves the insulation.
Tendrils
The tendrils of many plants (peas, cucumbers, and squash) are another form of modified leaf (FIGURE 6-44). Unlike photosynthetic leaves, tendrils grow indefinitely and contain cells that are capable of sensing contact with an object. When the tendril touches something, the side facing the object stops growing but the other side continues to elongate, causing the tendril to coil around the object and use it for support. A lamina would be detrimental, and none forms. Whereas many foliage leaves sense the direction of sunlight and reorient the lamina for maximum photosynthesis, tendrils respond by sensing solid objects and growing around them.
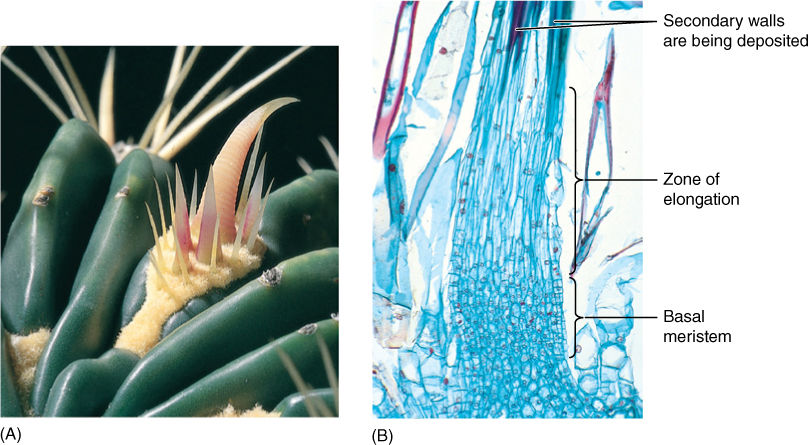
FIGURE 6-43 (A) The spines of this barrel cactus (Ferocactus) are modified leaves. The cluster of spines is actually an entire axillary bud. (B) Cactus spines have a basal meristem; as new cells are formed, older cells are pushed upward. As they move out of the meristem, they fill their central vacuole with water, elongate, then deposit a thick secondary wall and differentiate into fiber cells (×50).

FIGURE 6-44 This tendril is a highly modified leaf: It has no leaf blade, and whereas foliage leaves stop growing after they reach a specific size, this tendril continues to grow.
Leaves with Kranz Anatomy
A distinct type of leaf anatomy occurs in plants that have a special metabolism called C4 photosynthesis. These leaves lack palisade parenchyma and spongy mesophyll but have prominent bundle sheaths composed of large chlorophyllous cells. Surrounding each sheath is a ring of mesophyll cells that appear to radiate from the vascular bundle. These plants possess a mechanism of carbon dioxide transport that requires this special Kranz anatomy and adapts C4 plants to arid environments. (See Chapter 10 for details of C4 photosynthesis and its relationship to Kranz anatomy.)
Insect Traps
The ability to trap and digest insects has evolved in several families. Insectivory has evolved in plants that grow in habitats poor in nitrates and ammonia; by digesting insects, plants obtain the nitrogen they need for their amino acids and nucleotides. Trap leaves can be classified as either active traps that move during capture or passive traps incapable of movement. The best known passive traps are the pitcher leaves of Nepenthes, Darlingtonia, and Sarracenia (FIGURE 6-45A). Although the leaf appears highly modified, it is actually similar to many foliage leaves. It is thin, parenchymatous, and capable of photosynthesis. It has numerous stomata and vascular bundles as well as mesophyll containing aerenchyma and chlorenchyma. The most significant differences are that the lamina is tubular rather than flat, and it secretes a watery digestive fluid. The epidermis in the digestive region must be absorptive rather than impermeable. Also, the throat of the pitcher contains numerous trichomes that point toward the liquid; it is easier for insects to walk in the direction of the trichomes, and they are thus led to their death.
Leaves of sundew (Drosera) (FIGURE 6-45B) are active traps; they too have many features in common with foliage leaves, but their upper surface is covered with glandular trichomes that secrete a sticky digestive liquid. After an insect is caught on a single trichome, adjacent trichomes are stimulated to bend toward the victim, placing their digestive drops on it as well. The entire leaf blade curls around the insect so that many trichomes come into contact with it. The modifications in these leaves have been largely metabolic rather than structural: Trichomes and lamina must be able to sense and respond to the presence of an insect, carry on secretion and absorption, and unfold after digestion is complete.

FIGURE 6-45 (A) Nepenthes has elaborate pitcher leaves. They have an ordinary petiole and lamina, but the leaf tip is long, narrow, and pendant. Its extreme tip turns upright and develops into the hollow pitcher that contains a digestive mixture and an epidermis that can absorb nitrogen. The ultimate portion of the leaf tip is a broad flat roof that prevents rain from falling into the pitcher and diluting the digestive juices. (B) Sundew leaves have many shapes; those of Drosera capensis are long and narrow. Just after an insect is caught, the leaf curls and places numerous stalked glands on it. The drop at the end of each gland is both sticky and digestive. Once digestion is complete, the leaf uncurls and is ready for its next meal. (C) Each half of the blade of a Venus’ flytrap leaf has three hairs; if an insect touches any two, it stimulates the trap to close.
Venus’ flytraps (Dionaea muscipula) have leaves that are held flat, like most foliage leaves, but in Dionaea, this position is maintained only because motor cells along the upper side of the midrib are extremely turgid and swollen (FIGURE 6-45C). When an insect walks across the trap, it brushes against trigger hairs. If two of these are stimulated, midrib motor cells lose water quickly, and the trap rapidly closes as the two halves of the lamina move upward. On the margins are long interdigitating teeth that trap the insect; short glands begin to secrete digestive liquid. After digestion and absorption are complete, the midrib motor cells fill with water, swell, and force the trap open, ready for a new victim.
 At the Next Level
At the Next Level
1. Ecological modifications of foliage leaves. Thin, flat foliage leaves absorb sunlight and carbon dioxide, and they lose water. This shape gives a plant a large area of contact with the environment. The size and shape of leaves, their volume, and surface area all must be adaptive in the plant’s habitat. It is common to see suites of characters in most plants of particular habitats. For example, we expect to find certain leaf characters in cold, alpine, wind-swept habitats, different ones in hot, rainy tropical habitats. Topics to search for are drip tips, succulent leaves, sclerophylls, spines, and insectivorous plants.
2. Evolution of leaves. Foliage leaves evolved at least three times: in mosses, in lycophytes, and in seed plants and ferns. We know a great deal about certain aspects of leaf evolution, much less about other aspects. Search on topics such as microphyll, lycophyll, megaphyll, euphyll, telome theory, and transfusion tissue.
3. Abscission. Some leaves remain on a plant after they have died, but leaves of other species have abscission zones and cut themselves away from the plant when they can no longer photosynthesize, either because they are too old or too stressed. Abscission zones are also present and important in flowers, fruits, inflorescences, and twigs. In bald cypress (Taxodium distichum), leaves remain on twigs but whole twigs are abscised, taking the leaves with them. Abscission zones vary in their structure and placement, and also in the factors that control when they are active in causing an organ to fall.
SUMMARY
1. Natural selection has resulted in the evolution of numerous types of leaves involved in photosynthesis, protection, support, water and nutrient storage, and nitrogen absorption.
2. Foliage leaves must be resistant to pathogens and stresses, must produce more sugar than was used in their construction, and must not act as sails. Modifications related to these functions may prevent foliage leaves from being optimally adapted for photosynthesis.
3. Foliage leaves typically consist of a blade with or without a petiole; the blade may be simple or either pinnately or palmately compound.
4. Leaf variability may involve distinct juvenile and adult leaves or distinct leaves on long shoots and short shoots.
5. An abscission zone typically contains a separation layer in which cell walls rupture and a protective layer whose cells form a corky leaf scar.
6. Internally, most foliage leaves have an upper palisade parenchyma rich in chlorophyll and a lower spongy mesophyll that allows gas circulation. Vascular bundles composed of primary xylem and phloem are usually arranged in a netted pattern in basal angiosperms and eudicots and a parallel pattern in monocots. Minor veins load sugars and unload water.
7. Leaves are initiated only at shoot apical meristems, beginning as small conical leaf primordia. Eudicot leaves generally have diffuse growth in all parts simultaneously; most monocot leaf blades grow from a basal meristem.
IMPORTANT TERMS
abscission zone
bud scales
bundle sheath
compound leaf
lamina
leaf blade
leaf primordium
leaf scar
leaf traces
leaflets
mesophyll
midrib
minor veins
palisade parenchyma
parallel venation
petiole
rachis
reticulate venation
sessile leaf
simple leaf
spongy mesophyll
stipules
REVIEW QUESTIONS
1. Leaves carry out many roles in a plant’s life. Give examples of leaves that do the following:
a. Provide protection
b. Provide support
c. Provide storage
d. Obtain nitrogen
Are leaf structures and metabolisms that are selectively advantageous for one of these functions also advantageous for all of them?
2. What is the most obvious function of foliage leaves? What are some other functions that are often taken for granted?
3. Most leaves can be quite thin. Why? What would be the condition of lower layers in a thick leaf?
4. What is the stalk of a leaf called? What is the broad, flat, thin part?
5. Box 6-1, “Plants and People: Leaves, Food, and Death,” describes several ways that leaves affect animals. What is the name of the toxic chemical in leaves of poison hemlock and death camas? In leaves of tobacco? Can this poison be absorbed by the mucous membranes? Can it be passed across the placenta to an unborn baby? This chemical causes cancer of which parts of our bodies?
6. In many monocots such as grasses and irises, foliage leaves have a shape quite different from that of eudicot leaves. Their shape tends to be (circle two: long, short, tapering, wide). They lack a petiole and instead have a _________________ ______________________ that wraps around the stem. Name three monocots that have leaves that are unusual in that they appear to have petioles and laminas.
7. If a leaf has a blade that consists of one piece of tissue, we say the leaf is __________________. If the blade consists of several pieces, however, we say it is a __________________ leaf. What type of leaf is shown for a rose in Figure 6-7? For a Virginia creeper?
8. How can you distinguish between a compound leaf and a twig with several simple leaves? Assuming both a simple and a compound leaf have the same texture, which is more easily eaten by an insect larva?
9. Does a plant produce only one type of leaf during its entire life or can some plants produce various types of leaf (hint: Figure 6-11)? Explain.
10. There are two types of venation in the leaves of flowering plants. Describe the type found in eudicot leaves. Describe the type in monocot leaves.
11. What does the abscission zone do in leaves? Would you guess that there are or are not abscission zones in flowers and fruits?
12. In what ways does the upper epidermis of a leaf differ from the lower epidermis? How are these structural differences adaptive?
13. This page is 22 cm wide and 27 cm tall. Calculate its surface area (remember, just like a leaf, it has an upper and a lower surface). If this page were a leaf with the same density of stomata as a leaf of sunflower (see Table 6-2), how many stomata would be on the upper side of the page and how many on the lower side?
14. The interior tissues of a leaf are called mesophyll. In most leaves, there is an upper layer of columnar cells called the ______________________ ___________________ and a lower portion of _____________ ______________ open, loose aerenchyma. In Box 6-2, “Botany and Beyond: Leaf Structure, Layer by Layer,” Figure (A) shows the upper layer of columnar cells, but they do not look columnar. Why not? Figure (E) in the box shows the lower aerenchymatous layer. Can you tell what the shape of the intercellular spaces is from this two-dimensional micrograph (look at Figure 6-21 before you answer)?
15. Many leaves have a big, obvious vein that runs along their center, and many veins emerge from it. What is the large vein called (and can a leaf have more than one [look at Figure 6-2])? The smaller veins that emerge from it? What are the very smallest veins of a leaf called? Which veins supply water directly to mesophyll cells—the biggest veins or the smallest?
16. What is a bundle sheath in a leaf?
17. Draw a cross-section of a foliage leaf and label each part; be certain to show all the types of vascular tissues in the bundles.
18. Which parts of the leaf in Question 17 would be emphasized and which would be reduced in each of the following: tendrils, spines, bud scales, scale leaves of a bulb, and succulent leaves of a desert plant?
19. Most petioles are rather small, but they can be either massive or very long. List several examples of unusually large petiole (two are edible).
20. Shoot apical meristems make small groups of cells that protrude upward and develop into leaves. What are these small protrusions of cells called?
21. Look at the cross-sections of leaf primordia in Figure 6-31A (labeled “Leaves”). Each one wraps completely around the shoot apical meristem. Can you explain why each leaf appears to surround the meristem (hint: this is a micrograph of a grass, a monocot, and thus, it has leaves like those in Figures 6-4B and 6-4C).
22. How does the growth of eudicot and grass leaves differ? Which type would be more capable of recovering from attack by leaf-eating insects or grazing deer?
23. Dormant buds, such as the terminal and axillary buds of twigs in winter, consist of bud scales, young leaves, and leaf primordia. Which were formed by the apical meristem first and which were initiated last?
24. Figure 6-36 shows many plants of the desert plant Lithops. How many leaves does each plant have? (By the way, even if you grow these in ideal conditions, they never have more than this number.)
25. Sclerophyllous foliage leaves like those in holly, barberry (Figure 6-37), and Yucca (Figure 6-38) have many unusual features. Do they live for 1 year or longer? They are more resistant to animals that might try to eat them. Why? What type of cells do they have that most leaves do not have?
26. Leaves of conifers are perennial. Those of bristlecone pine live for at least ____ years, and their vascular bundles produce new _____________ every year. Many conifers have needle-shaped leaves, but those of incense cedar are ____________ shaped (see Figure 6-39B).
27. The spines of cacti are modified leaves, and as with bud scales, spines have a distinct structure related to their function. Why would the blade of a photosynthetic leaf be useless as a protective device? Cactus spines have no mesophyll parenchyma or vascular tissues. Instead, their mesophyll consists of _______________ ______________ __________________. What chemicals do spine cells have in their walls? Are spine cells parenchyma, collenchyma, or sclerenchyma? After spine cells are mature, do they remain alive? Look at all your answers to this question: Does this modified leaf have much in common with a foliage leaf?
28. Cacti have _________ types of leaves. The green cactus body has _____________________ green leaves, and the clusters of spines are their ________________ ________________. Spines are modified leaves of axillary buds.
29. Several species have leaves that catch and digest insects, obtaining nitrogen fertilizer from their bodies. Describe how each of the following trap leaves function—the pitchers of Nepenthes, the flypaper leaves of sundew, the traps of Venus’ flytrap. Which are active and which are passive?
Design Credits: Hummingbird: © Tongho58/Moment/Getty; Green plant cells: © ShutterStock, Inc./Nataliya Hora; Purple tulip: © ShutterStock, Inc./Marie C Fields; Dandelion: © ShutterStock, Inc./danielkreissl; Poppy: © ShutterStock, Inc./Saruri; Plant icon: © ShutterStock, Inc./Vector; Digging man icon: © ShutterStock, Inc./Z-art