Botany: An Introduction to Plant Biology - Mauseth, James D. 2017
Plant Structure
Roots

Chapter Opener Image: Roots play many roles in the life of a plant, but a common function is to provide anchorage and stability. The roots shown here are old and woody, but they were initiated when the tree was just a sapling; it grew from a seed that had landed on the side of a fallen tree, so the young roots had to grow over the trunk of the dead tree for more than a meter before they ever reached the soil. In several decades, the fallen tree will decompose completely, then the roots will be holding up the trunk of their tree, more than a meter above the ground with just air between them.
OUTLINE
✵ Concepts
✵ External Structure of Roots
- Organization of Root Systems
- Structure of Individual Roots
✵ Internal Structure of Roots
- Root Cap
- Root Apical Meristem
- Zone of Elongation
- Zone of Maturation/Root Hair Zone
- Mature Portions of the Root
✵ Origin and Development of Lateral Roots
✵ Other Types of Roots and Root Modifications
- Storage Roots
- Prop Roots
- Aerial Roots of Orchids
- Contractile Roots
- Mycorrhizae
- Root Nodules and Nitrogen Fixation
- Haustorial Roots of Parasitic Flowering Plants
- Roots of Strangler Figs
Box 7-1 Plants Do Things Differently: Plants and People and Having a Weight Problem
LEARNING OBJECTIVES
After reading this chapter, students will be able to:
✵ List the three main functions of roots.
✵ Explain the properties and biological consequences of taproots, lateral roots, and fibrous root systems.
✵ Describe the structure of individual roots.
✵ Name the components of the internal root structure.
✵ Compare lateral root development to bud formation in shoots.
✵ Discuss other types of roots: storage, prop, aerial, contractile, mycorrhizae, nodules, haustorial, and strangler fig roots.
 Did You Know?
Did You Know?
✵ We know very little about roots because they are hidden in soil and we cannot easily see when they are active or dormant, or even if they have died.
✵ Root organization is similar to shoot organization: Roots have an epidermis, cortex, phloem, and xylem, but they lack pith.
✵ Certain cells in the tips of roots have dense starch grains that sink to the bottom of the cells; this indicates to the cells which way is down.
✵ Some plants (mangroves) that grow along seashores have roots that grow upward, providing submerged roots with oxygen.
![]() Concepts
Concepts
Most roots have three functions: (1) anchoring the plant firmly to a substrate, (2) absorbing water and minerals, and (3) producing hormones. Firm anchoring provides stability and is therefore important for virtually all plants. Stems, leaves, flowers, and fruits then can be properly oriented to the sun, pollinators, or fruit distributors. Without proper root attachment, trees and shrubs could not remain upright, and epiphytes would be blown from their sites in the tree canopy. A highly branched rhizomatous or stoloniferous plant might resist being blown over even without roots, but their horizontal stems are usually so flexible that roots are necessary to stabilize the aerial structure.
Although roots, like leaves, have an absorptive function, the two organs have totally different shapes. Sunlight always comes from above, but water and minerals are distributed on all sides of a root. Its cylindrical shape allows all sides to have the same absorptive capacity. Consider a leaf and a system of thin roots, both with equal volumes; the root system has a higher surface-to-volume ratio, ideal for absorption. The lower surface-to-volume ratio for leaves reduces the carbon dioxide absorption capacity, but not the absorption of light, and is actually beneficial for water conservation. Roots do not need to be adapted for light absorption, and they absorb water rather than needing to conserve it. Their cylindrical shape is also undoubtedly related to the growth of the roots through a semisolid, resistant medium. Even a light, porous soil can be most easily and thoroughly penetrated by narrow cylinders rather than thin sheets. Thus, although both leaves and roots have absorptive functions, it is selectively advantageous for them to have different shapes.
Roots are quite active in the production of several hormones; shoot growth and development depend on the hormones cytokinin and gibberellin imported from the roots. This reliance of the shoot on root-produced hormones may be a means of integrating the growth of the two systems. It is selectively advantageous for a plant to control the size of its shoot so that transpiration by its leaves does not exceed absorption by its roots, and a plant should not waste carbohydrates by constructing a larger root system than its shoot needs; the extra carbohydrate could be used for leaves or reproduction (FIGURE 7-1).
In many cases, roots have functions in addition to or instead of anchoring, absorption, and hormone production. Fleshy taproots, such as those of carrots, beets, and radishes, are the plant’s main site of carbohydrate storage during winter. As roots of willows, sorrel, and other plants spread horizontally, they produce shoot buds that grow out and act as new plants. This method of vegetative reproduction is quite similar to that of stoloniferous and rhizomatous plants, except that roots rather than stems are involved. In the palms Crysophila and Mauritia, roots grow out of the trunk and then harden into sharp spines. Ivy and many other vines have modified roots that act as holdfasts, clinging to rock or brick. Finally, many parasitic flowering plants (mistletoe and dodder) attack other plants and draw water and nutrients out of them through modified roots.
Distinct sets of characteristics are adaptive for different root functions. As roots specialize and become more efficient for particular tasks, they become poorly adapted for other tasks, and thus, several types of roots may occur in one plant, resulting in division of labor. For example, in addition to the large storage root, carrots and beets also have fine absorptive roots, and ivy has both holdfasts and absorptive roots. The characteristic types of structure and metabolism of each should be analyzed in terms of the function of the particular root.
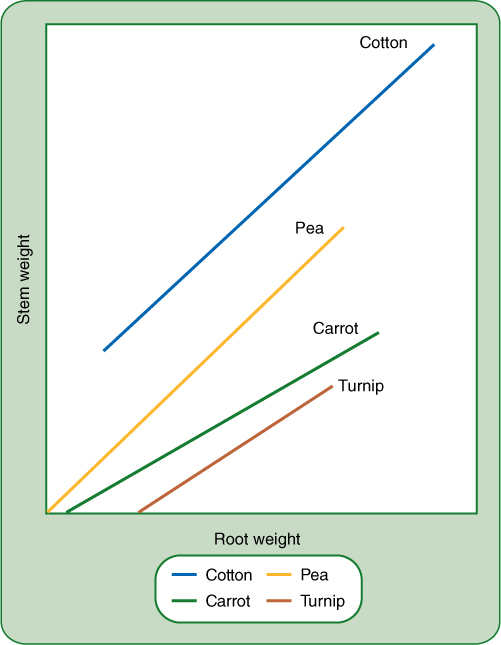
FIGURE 7-1 The ratio of root system to shoot system is critically important; for each species, as the root system becomes heavier, so does the shoot system by a characteristic amount. However, the relationship differs for each species: Obviously for a given amount of turnip root there is relatively little shoot, but there is much more shoot for the same amount of pea root, and even more for cotton.
![]() External Structure of Roots
External Structure of Roots
Organization of Root Systems
Roots must have an enormous absorptive surface; in order for a single unbranched root to have sufficient surface area, it would have to be hundreds of meters long, which would make conduction impossible (TABLE 7-1). Instead, plants have a highly branched root system (FIGURE 7-2). Most seed plants have a single prominent taproot that is much larger than all the rest and numerous small lateral roots or branch roots coming out of it (FIGURE 7-3). This taproot develops from the embryonic root, called the radicle, that was present in the seed; after germination, it grows extensively and usually becomes the largest root in the system. Carrots, beets, turnips, and other taproots sold in stores have dozens of fine lateral roots while growing, but these are removed before the products are shipped to market.
TABLE 7-1 Dimensions of the Root System of Barley (Hordeum vulgare), 4 Weeks Old
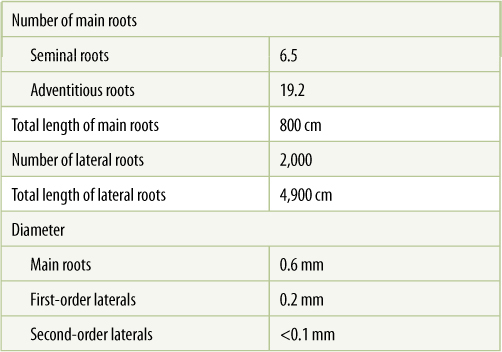
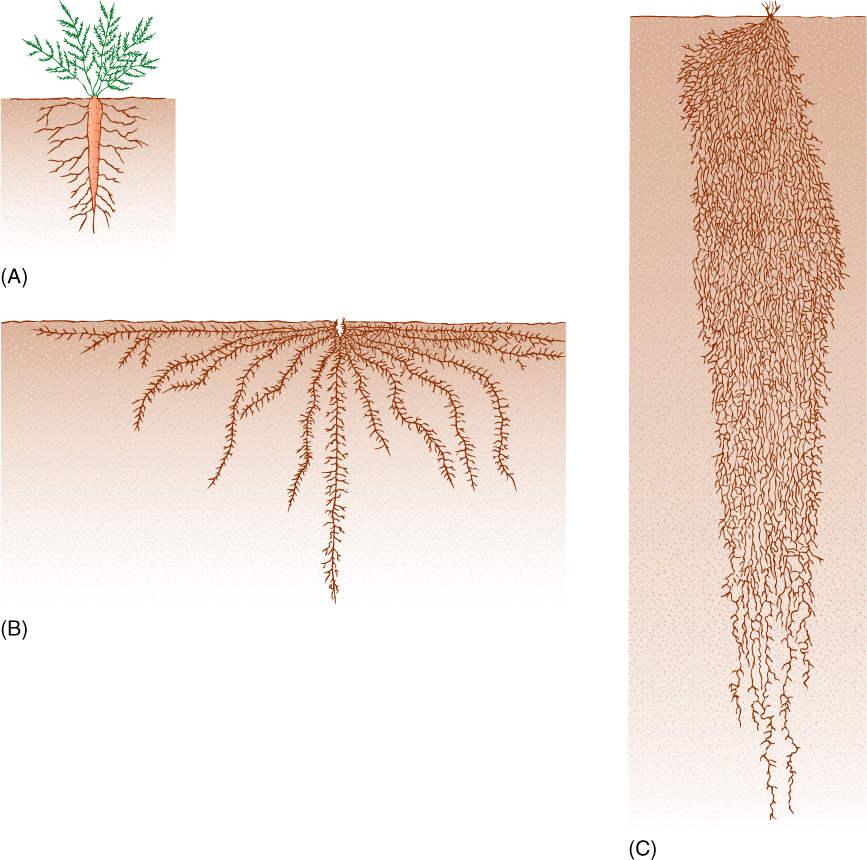
FIGURE 7-2 Some taproots, such as a carrot (A), become extremely swollen and are much larger than the numerous lateral roots, whereas in other species, such as sunflower (B), the taproot is about the same size as the laterals. The important criterion is that the taproot develops from the root of the embryo, the radicle. (C) A fibrous root system, as in this winter wheat, consists of many roots, none of which is the radicle. Instead, all of the main roots are adventitious roots that originated in stem tissue.

FIGURE 7-3 (A) The taproot of this tobacco seedling (Nicotiana tabacum) is definitely larger than the lateral roots, but more importantly, its development can be traced directly to the embryo. Relative size is not the critical factor; in many species, lateral roots are the ones that become enlarged. (B) The roots of radishes are obviously taproots.
Lateral roots may also produce more lateral roots, resulting in a highly ramified set of roots analogous to the highly branched shoot system of most plants. Lateral roots can become prominently swollen like a taproot, as in sweet potatoes and the tropical vegetable manioc (cassava). If the plant is perennial and woody, roots also undergo secondary growth, producing wood and bark.
Most monocots and some eudicots have a mass of many similarly sized roots constituting a fibrous root system (FIGURE 7-4). This arises because the radicle dies during or immediately after germination; root primordia at the base of the radicle grow out and form the first stages of the fibrous root system. As the plant ages, more root primordia are initiated in the stem tissue. Because these roots do not arise on pre-existing roots and because they are not radicles, they are known as adventitious roots. Adventitious roots increase the absorptive and transport capacities of the root system.
The functional significance of taproots versus fibrous root systems becomes apparent when the general growth forms of eudicots and monocots are considered. Many eudicots are perennial and undergo secondary growth, resulting in an increased quantity of healthy, functional wood (xylem) in both the trunk and roots (FIGURES 7-5A and 7-5B). This enlarging conduction capacity permits an increase in the number of leaves and fine, absorptive roots. This also occurs in other groups of seed plants such as conifers and cycads.
Most monocots cannot undergo secondary growth; after their stem is formed, the number of vascular bundles, tracheary elements, and sieve tubes is set, and their conducting capacity cannot be increased. Extra leaves could not be supplied with water, nor could their sugar be transported (FIGURES 7-5C and 7-5D). Such a shoot could not supply sugars to an ever-increasing taproot system. However, some monocots do increase their size by means of stolons or rhizomes: Their horizontal shoots branch and then produce adventitious roots (FIGURE 7-5E). Because these roots are initiated in the new stem tissues, they transport water directly into the new portions of the shoot, unhindered by the limited capacity of the older portions of the shoot. By this mechanism, monocot shoots can branch and grow larger, as long as they remain close enough to the substrate to produce new adventitious roots. For them, a fibrous root system is functional, whereas a taproot system is not.
The ability to form adventitious roots is not limited to monocots; many rhizomatous and stoloniferous eudicots also grow this way naturally. Furthermore, many eudicots that never produce adventitious roots in nature do so if they are cut; this is important in the process of asexual propagation by cuttings.
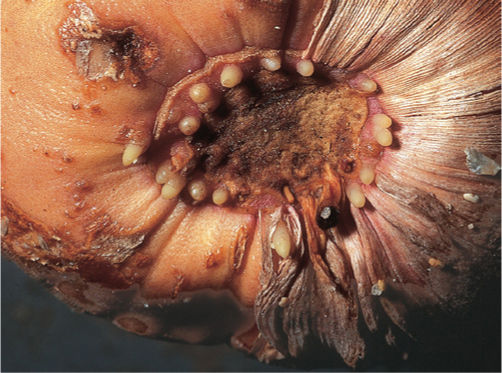
FIGURE 7-4 Corms of Gladiolus, like other monocots, have a fibrous root system. The radicle died shortly after germination; no root here has developed from the radicle. As the Gladiolus continues to grow, it will produce more roots, each independent of the others.
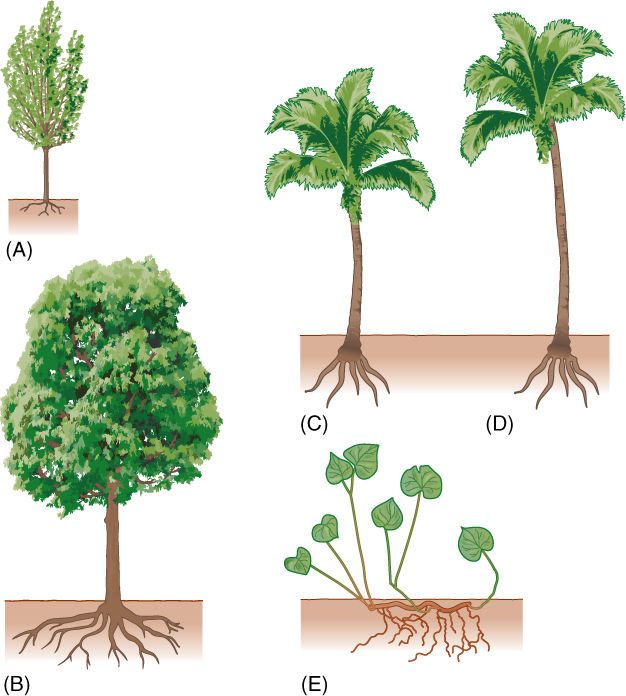
FIGURE 7-5 (A) A seedling of a woody plant has a few leaves and a small root system; the narrow trunk with a few vascular bundles can conduct water and nutrients between them. (B) An older woody plant has more leaves and a larger root system; the stem has more wood and bark, which increases its capacity to conduct water and sugar. Because most monocots do not undergo secondary growth, the stem of an older monocot is not wider (D) than that of a young plant (C), and it has no increased conducting capacity. Consequently, the old plant has no more leaves or roots than the young plant. (E) If a plant can produce adventitious roots, the bottleneck of the monocot stem does not matter. New roots originate near the aerial shoots and conduct directly into them, and little or no long-distance conduction occurs in the rhizome. No part of the monocot stem needs to conduct all of the water from all the roots to all the leaves and flowers, as does the trunk of a woody plant.
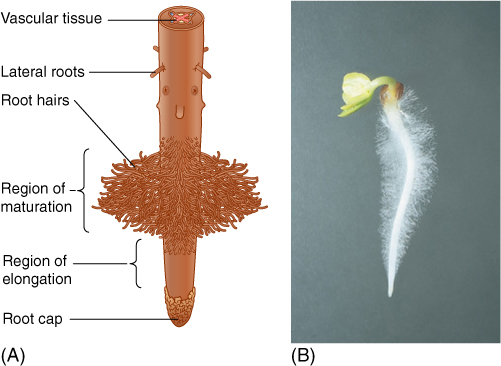
FIGURE 7-6 (A) The tip of a root consists of a root cap, the root apical meristem, a zone of elongation growth, and a region where root hairs are formed. (B) This radish seedling is only 3 days old and has abundant root hairs; those near the root tip are short because they are newer and younger than those near the shoot.
Structure of Individual Roots
An individual root is fairly simple; because it has no leaves or leaf scars, it has neither leaf axils nor axillary buds (FIGURES 7-6 and 7-7). The root tip, like that of the shoot, is the region where growth in length occurs. In roots, growth by discrete apical meristems is the only feasible type of longitudinal growth. In most animals, all parts of the body grow simultaneously (diffuse growth), whereas roots and stems elongate only at small meristematic regions (localized growth). Because the root is embedded in a solid matrix, it is impossible for all parts to extend at once; the entire root would have to slide through the soil. With apical growth, only the extreme tip pushes through soil.
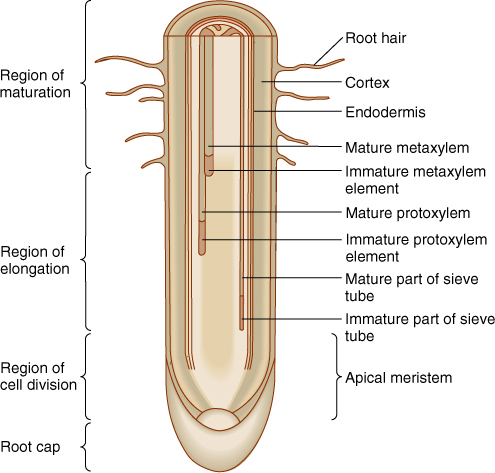
FIGURE 7-7 Although the exterior of a root appears rather uniform, several distinct zones of differentiation are present internally. Root hairs form only above the elongation zone, and the endodermis and first vascular tissues appear earlier than do root hairs.
Whereas the shoot apical meristem is protected by either bud scales or young, unexpanded foliage leaves, the root apical meristem is protected by a thick layer of cells, the root cap (Figure 7-7 and FIGURE 7-8). Although some soils appear soft and easily penetrable, on a microscopic scale, all contain sand grains, crystals, and other components that can damage the delicate apical meristem and root cap. Because the cap is forced through the soil ahead of the root body, it is constantly being worn away and must be renewed by cell multiplication.
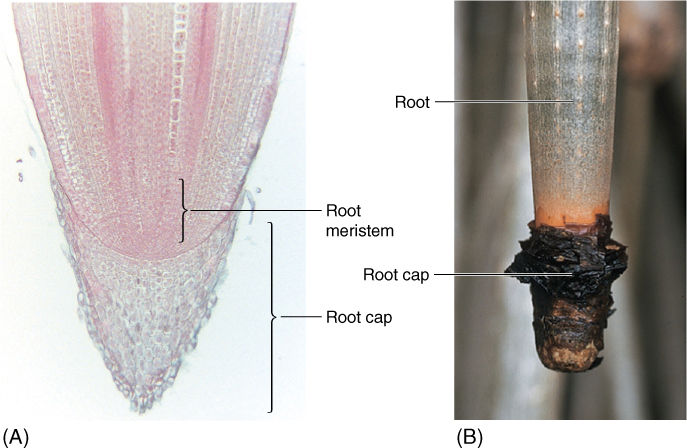
FIGURE 7-8 (A) The root cap is distinct from the root proper. The root apical meristem is located at the apex of the root proper but is buried under the root cap. Because no leaves, leaf traces, or branches are present, cells develop in an extremely orderly fashion in regular files. It is easy to see that these cells are derived directly from the meristem; thus, they make up primary tissues (×150). (B) Screw pine (Pandanus) has giant aerial roots that allow us to see the root cap more easily than with ordinary tiny roots. The root cap cells die and are sloughed off.
Dictyosomes of root cap cells secrete a complex polysaccharide called mucigel, which lubricates passage of the root through the soil. It also causes soil to release its nutrient ions and permits the ions to diffuse more rapidly toward the root. Mucigel is rich in carbohydrates and amino acids, which foster rapid growth of soil bacteria around the root tip. The metabolism of these microbes is believed to help release nutrients from the soil matrix.
Just behind the root cap and root apical meristem is a zone of elongation only a few millimeters long within which the cells undergo division and expansion (Figure 7-7). Behind it is the root hair zone, a region in which many of the epidermal cells extend out as narrow trichomes. Root hairs form only in a part of the root that is not elongating; otherwise, they would be shorn off.
Root hairs greatly increase the root’s surface area. In a study of rye, a single plant was found to have 13 million lateral roots with 500 km of root length and a surface area of 200 m2. Because of the abundant production of root hairs, however, the total surface area was doubled. Root hairs have other effects that should not be overlooked. Most pores in soil are too narrow for a root (usually at least 100 mm in diameter) to penetrate (FIGURE 7-9). Root hairs, however, being only about 10 mm in diameter, can enter any crevice and extract water and minerals from it. Furthermore, carbon dioxide given off by the respiration of root hairs combines with soil water to form carbonic acid, which helps release ions from the soil matrix. Without the acid, ions would be too firmly bound to soil particles for the root to absorb them. Root hairs are unicellular, never have thick walls, and are extremely transitory. They die and degenerate within 4 or 5 days after forming.
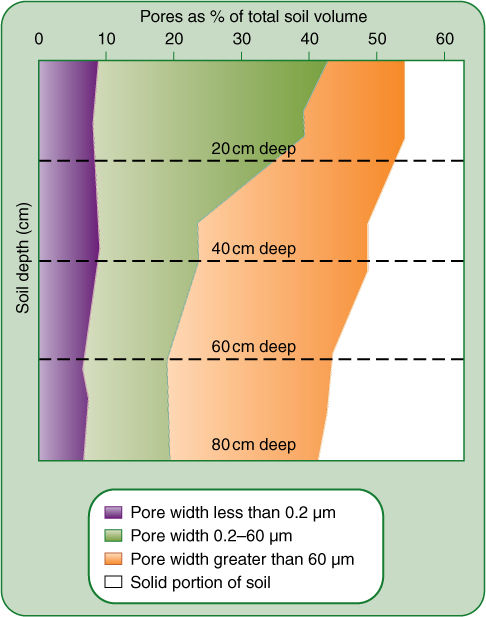
FIGURE 7-9 Much of a soil is composed of spaces between soil particles; in the uppermost 20 cm (uppermost dashed line) of a soil composed of sandy loam, 54% of the soil is pore space. Of this, 9% consists of extremely fine spaces less than 0.2 μm wide (purple), 33% of spaces ranging between 0.2 and 60 μm (green), and 12% of large spaces wider than 60 μm (orange). At deeper levels, the soil is more compacted, containing only about 42% pore space (bottom of graph); the decrease is due mostly to a compaction of intermediate-sized spaces.
Behind the root hair zone is a region where new lateral roots emerge. They may occur in rows or may appear to be randomly distributed on the parent root. Outgrowth of lateral roots often depends on the soil microenvironment. If the parent root grows into a zone of rich, moist soil, numerous lateral roots form, and the pocket is fully exploited. If the soil is poor, hard, or dry, few lateral roots emerge.
![]() Internal Structure of Roots
Internal Structure of Roots
Root Cap
To remain in place and provide effective protection for the root apical meristem, the root cap must have a specific structure and growth pattern. The cells in the layer closest to the root meristem are also meristematic, undergoing cell division with transverse walls and forming files of cells that are pushed forward (FIGURE 7-10). Simultaneously, cells on the edges of this group grow toward the side and proliferate. Although cells appear to extend around the sides of the root, the root is actually growing through the edges of the root cap.
Cells are small and meristematic when first formed at the base of the root cap, but as they are pushed forward, they develop dense starch grains and their endoplasmic reticulum becomes displaced to the forward end of the cell. These cells detect gravity because their dense starch grains settle to the lower side of the cell.
As cells are pushed closer to the edge of the cap, their structure and metabolism change dramatically. Endoplasmic reticulum becomes less conspicuous, starch grains are digested, and the cell’s dictyosomes secrete copious amounts of mucigel by exocytosis. Simultaneously, the middle lamella breaks down and releases cells, which are usually crushed by expansion of the root. It has been estimated that only 4 or 5 days pass from cell formation in the root cap to its sloughing off. Consequently, the cap is constantly regenerating itself. A dynamic equilibrium must be maintained between these two processes.
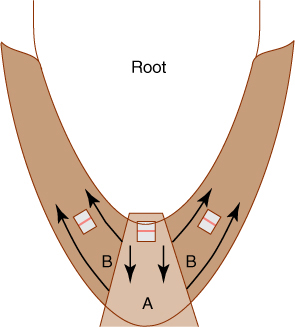
FIGURE 7-10 In the root cap, cells in the central portion (A) divide so that the two daughter cells are aligned with the root and the root cap grows forward. On the edges (B), cells divide and expand in such a way that cells flow radially outward.
Root Apical Meristem
If the root apical meristem is examined in relationship to the root tissues it produces, regular files of cells can be seen to originate in the meristem and extend into the regions of mature root tissues (Figure 7-8). The root is more orderly than the shoot because it experiences no disruptions because of leaf primordia, leaf traces, or axillary buds. Because cell files extend almost to the center of the meristem, one might assume that cell divisions are occurring throughout it. However, the use of a radioactive precursor of DNA, such as tritiated thymidine (tritium is a radioactive form of hydrogen), can demonstrate that the central cells are not synthesizing DNA: Their nuclei do not take up the thymidine and thus do not become radioactive (FIGURE 7-11). This mitotically inactive central region is known as the quiescent center. These cells are more resistant to various types of harmful agents such as radiation and toxic chemicals, and it is now believed that they act primarily as a reserve of healthy cells. If part of the root apical meristem or root cap is damaged, quiescent center cells become active and form a new apical meristem. After the new meristem is established, its central cells become inactive, forming a new quiescent center. Such a replacement mechanism is extremely important because the root apex is probably damaged frequently by various agents—sharp objects, burrowing animals, nematodes, and pathogenic fungi.
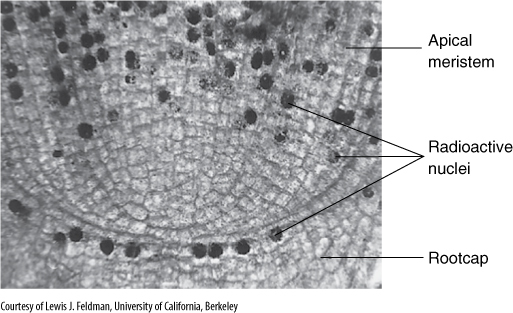
FIGURE 7-11 These roots were grown briefly in a solution of tritiated thymidine, a radioactive precursor of DNA. In cells that were undergoing the cell cycle, during S phase, the radioactive thymidine was incorporated into the nuclei. After a few hours, the root was killed, sliced into sections, and placed on photographic film in the dark. The radioactive nuclei caused black spots to form in the film next to the nuclei. The slide was then given a brief exposure to light so that the outlines of the cells and nuclei would be faintly visible (too much light would obscure the radioactivity-induced black spots). The quiescent center is the region where no nuclei became radioactive. Apparently no cell in the region passed through S phase while tritiated thymidine was available—these cells were in cell cycle arrest.
Zone of Elongation
Just behind the root apical meristem itself is the region where cells expand greatly; some meristematic activity continues, but mostly cells are enlarging. This zone of elongation is similar to the shoot’s subapical meristem region. Cells begin to differentiate into a visible pattern, although none of the cells is mature. The outermost cells are protoderm and differentiate into epidermis. In the center is provascular tissue, cells that develop into primary xylem and primary phloem. As in the stem, protoxylem and protophloem, which form earliest, are closest to the meristem. Farther from the root tip, older, larger cells develop into metaxylem and metaphloem. Between the provascular tissue and the protoderm is a ground tissue, a rather uniform parenchyma that differentiates into root cortex.
In the zone of elongation, tissues are all quite permeable. Minerals penetrate deep into the root through the apoplast simply by diffusing along the thin, fully hydrated young walls and intercellular spaces. This zone is so short that little actual absorption occurs there, and much that is absorbed is probably used directly for the root’s own growth.
Zone of Maturation/Root Hair Zone
In the maturation zone (FIGURE 7-12), several important processes occur more or less simultaneously. Root hairs grow outward, greatly increasing absorption of water and minerals. In some electron micrographs, a thin cuticle appears to be present on root epidermal cells, but this may be just a layer of fats. The zone of elongation merges gradually with the zone of maturation; no distinct boundary exists because the differences between the two represent the gradual continued differentiation of the cells.
Although cortex cells continue to enlarge, their most significant activity is transfer of minerals from the epidermis to the vascular tissue. This can be either by diffusion through the walls and intercellular spaces (called apoplastic transport) or by absorption into the cytoplasm of a cortical cell and then transferal from cell to cell, probably through plasmodesmata (symplastic transport; FIGURE 7-13). Cortical intercellular spaces are also important as an aerenchyma, allowing oxygen to diffuse throughout the root from the soil or stem.
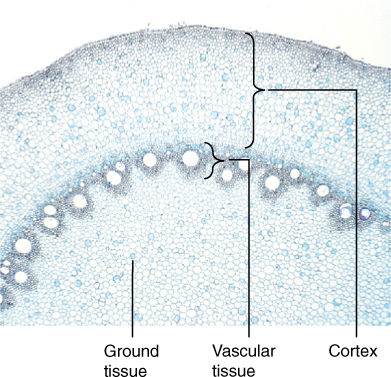
FIGURE 7-12 The cortex may be broad in many roots, and its interior boundary is the endodermis (×15).
In the zone of maturation, minerals do not have free access to the vascular tissues because the innermost layer of cortical cells differentiates into a cylinder called the endodermis (Figure 7-13A). Cells of the endodermis have tangential walls, those closest to the vascular tissue or the cortex, which are ordinary thin primary walls; however, their radial walls—the top, bottom, and side walls, that is, all the walls touching other endodermis cells—are encrusted with lignin and suberin, both of which cause the wall to be waterproof. The bands of altered walls, called Casparian strips, are involved in controlling the types of minerals that enter the xylem water stream. Cortex cells can not exert any control over movement of minerals within intercellular spaces; without an endodermis, minerals of any type could move from the soil into the spaces, then into the xylem, and then into all parts of the plant. Because Casparian strips are impermeable, however, minerals can cross the endodermis only if endodermal protoplasts absorb them from the intercellular spaces of the cortex apoplast or from cortical cells and then secrete them into the vascular tissues (Figure 7-13B). Many harmful minerals can be excluded by the endodermis. It is not a perfect barrier against uncontrolled apoplastic movement because in the zone of elongation, where the endodermis is not yet mature, minerals do have free access to the protoxylem, but this is only a small amount of uncontrolled movement. Many glands and secretory cavities also have Casparian strips, which prevent the glands’ secretory product from seeping into the surrounding tissues.
Within the vascular tissues, many of the much larger cells of the metaxylem and metaphloem become fully differentiated and functional in the zone of maturation. The arrangement of these tissues differs from that in stems: Instead of forming collateral bundles containing xylem and phloem, the xylem of almost all plants except some monocots forms a solid mass in the center, surrounded by strands of phloem; no pith is present (FIGURES 7-14A and 7-14B). In roots of many monocots, strands of xylem and phloem are distributed in ground tissue (FIGURES 7-14C and 7-14D). Within the xylem, the inner wide cells are metaxylem and the outer narrow ones are protoxylem. Two to four or more groups of protoxylem may be present, the number being greater in species with larger roots with a wider mass of xylem (FIGURE 7-15); the number of phloem strands equals the number of protoxylem strands. Within phloem strands, protophloem occurs on the outer side, and metaphloem on the inner side. Other than the arrangement, the vascular tissues of the root are similar to those of the stem and leaf. Those formed first are narrowest and most extensible, often finally being torn by continued elongation and expansion of cells around them. Those formed after adjacent cells have stopped expanding are larger and, in xylem, have heavier walls, often with bordered pits.
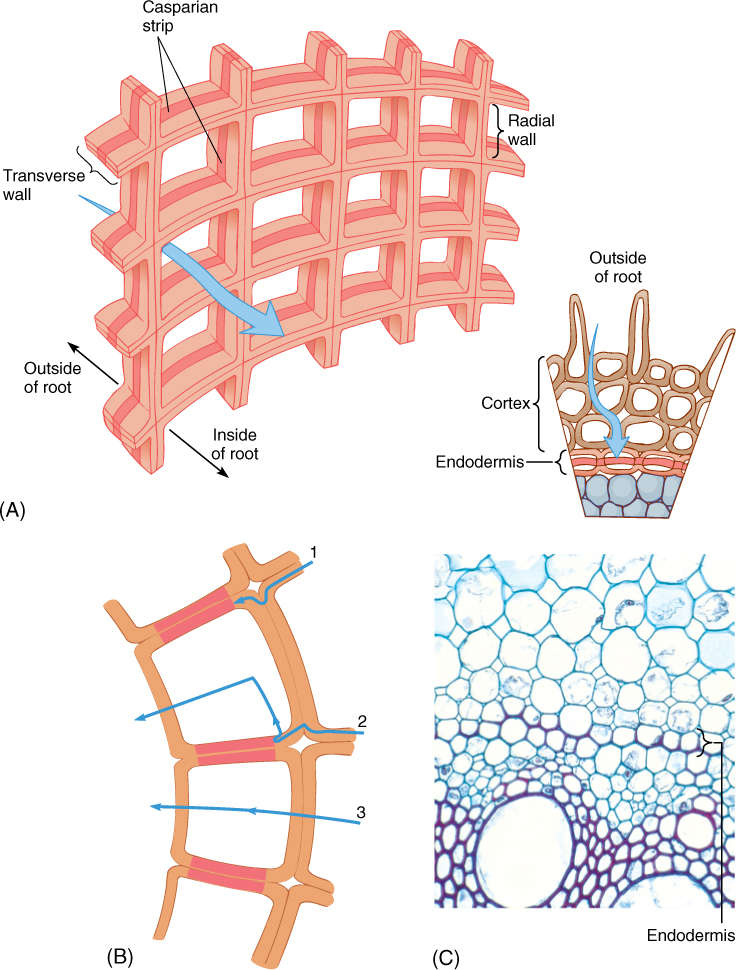
FIGURE 7-13 (A) The endodermis is a cylinder, one layer of cells thick, each with Casparian strips. The best analogy is a brick chimney: It is a cylinder, one layer thick. The cement is analogous to the Casparian strips. (B) No apoplastic transport occurs across the endodermis. Materials that have been moving apoplastically (1) are stopped and can proceed farther only if the endodermis plasma membrane accepts them (2). Symplastic transport (3) is not affected. Plasma membranes that are impermeable to a particular type of molecule prevent that molecule from crossing the endodermis. (C) The endodermis in this corn is relatively easy to see: In all cells, Casparian strips are visible as red lines on the radial walls. In a few cells, only Casparian strips are present, but in many, the radial walls and inner tangential walls have started to thicken; it is unusual for this development to start so early (×200).
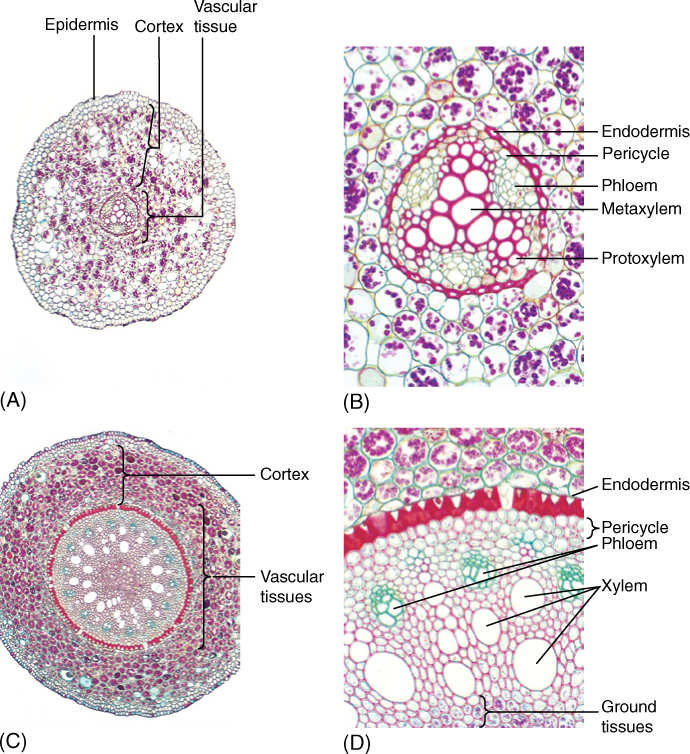
FIGURE 7-14 (A) Low-magnification view and (B) high-magnification view of a transverse section of a eudicot root (buttercup, Ranunculus) showing the broad cortex and the small set of vascular tissues. The xylem has three sets of protoxylem, the narrow cells at the tips of the arms (B). The central, larger xylem cells are metaxylem. Three masses of phloem are also present; this is a triarch root. The pericycle is the set of cells between the endodermis and the vascular cells; (A) ×50; (B) × 250. (C) Low-magnification view and (D) high-magnification view of a transverse section of a monocot root (greenbrier, Smilax). The vascular tissue consists of numerous large vessels and separate bundles of phloem. The endodermis is very easy to see; (C) ×15; (D) ×150.
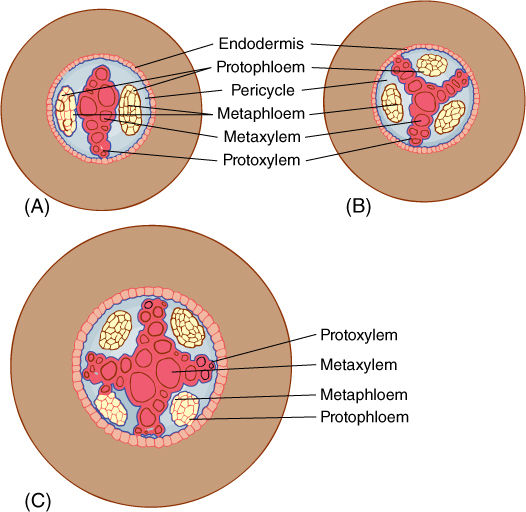
FIGURE 7-15 The number of strands of protoxylem in a root varies from one species to another and among the roots of one plant. Generally wider, more robust roots have more protoxylem masses. Having two masses of protoxylem (and two of phloem) is diarch (A). Three is triarch (B), and four is tetrarch (C). These diagrams represent three separate roots, not young and older stages of a single root; a diarch root does not become triarch and then tetrarch as it ages.
Between the vascular tissue and the endodermis are parenchyma cells that constitute an irregular region called the pericycle (Figures 7-14 and 7-15). When lateral roots are produced, they are initiated in the pericycle.
Plants Do Things Differently
BOX 7-1 Plants and People and Having a Weight Problem
Most of us probably do not spend too much time worrying about starving to death. However, throughout much of the history of civilization, people had to be careful to store up enough food to last not only through winter but through spring as well. Food supplies had to last until gardens could provide potatoes, beans, wheat, and other staples. Grocery stores are a recent luxury. How did people store food? If dry seeds like wheat and beans are kept free of moisture, they last a long time. Grapes and milk are not dry, but they can be preserved by turning the first into wine and the second into cheese. Meat and fish can be dried, smoked, or salted for long-term storage.
One particular method of storing food is particularly popular with humans and is the only means available to most animals. Eat the food whenever it is available, and store it as fat inside our body. Beans can become moldy. Rats can find our supply of dried meat, but fat in our adipose tissues is safe. For animals that hibernate, getting fat in autumn is the only way to survive. We may regret eating so much at Thanksgiving, but the pilgrims did not. Feasting was a means of storing food that might otherwise spoil during winter.
Plants too must be adapted to the availability—or the scarcity—of food. For plants, “food” is supplied by photosynthesis, and that requires only light, carbon dioxide, and water. In tropical climates where temperatures are always mild and droughts virtually never occur, plants have leaves throughout the year and photosynthesize every day, making all of the carbohydrates they need whenever they need them. Storing food reserves is not a problem. In temperate climates, evergreen trees such as pines and hollies also are able to photosynthesize most days, being inhibited only when it is extremely cold. But deciduous plants—those that drop their leaves and become dormant—are similar to animals in that they need to store food to maintain their metabolism while leafless. When a plant abscises its leaves in autumn, it is almost as if an animal were throwing away its entire digestive tract in anticipation of growing an entire new one in spring. If a plant had no nutrient reserves inside itself when it abscised its leaves, it would not even be able to make new leaves in the following spring. It would starve to death.
Energy reserves can be stored as a variety of chemical compounds. We animals sequester our reserve energy as fats. A little bit of energy is stored as a polymer called glycogen, located in our liver and muscle cells, but that is only enough to keep us going for a few hours as any runner or cyclist knows. Plants virtually never store fats; they rely on starch instead. Why do plants and animals differ on such a simple feature? Let us look at the consequences of each alternative storage molecule.
An energy storage molecule should not be too heavy, and it must be stable enough that it does not “go bad” within the plant or animal’s body. Fat is the most lightweight means of storing energy. A pound of fat—whether it is lard, oil, butter, or margarine—stores more energy than a pound of starch or protein. For an organism that needs to move, weight will be less of a problem if it stores fat rather than if it stores the same amount of energy as starch. The next time your bathroom scale reads 10 pounds more than you want, be glad we do not store starch. We would be even more overweight and have even bigger rear ends.
Plants, however, do not move around too much; thus, saving on weight is not a real necessity. For plants, the long-term stability of starch is better than the lightness of fat. Some plants save up energy for years, not just months, and starch will last that long in the plant’s body. For example, century plants bloom only once after growing for about 12 to 15 years (but not for an entire century), and fishtail palms may not bloom until they are more than 70 years old. These plants store some starch each year, then use it all at once in a massive flowering. Fat would not last that long because it becomes rancid if exposed to oxygen, and all parts of a plant are well aerated.
Pollen and seeds are exceptions. Many flowers produce pollen with a drop of oil rather than a grain of starch, which makes it lighter and easier for wind or insects to carry it to another flower. Seeds such as peanuts, cashews, and sesame store oil and thus are lighter and smaller and more easily moved by animals. Avocados are very rich in oil, but rather than being an energy storage mechanism, it is a reward that entices an animal to eat the fruit and then spit the seed out somewhere, thereby dispersing the plant’s seeds far and wide.
Plants also store a little starch here and there throughout their bodies, some in cortex cells, some in pith cells, and even wood can store a bit. However, when a plant needs to store a lot of starch, it almost always relies on its roots. The enlarged roots of beets, carrots, radishes, sweet potatoes, and similar plants are filled with starch. By using roots as a storage organ, the plants are putting their reserves underground, out of sight of hungry animals. Also, the soil is a more stable environment, being neither as hot during the day nor as cold at night as the air, and similarly, it maintains a more uniform humidity. The storage tissue in roots is usually wood—a type of xylem; thus, just as in adipose tissue of animals, root storage tissues are well vascularized.
Even though plants and animals store energy differently, the reasons are understandable when we consider the consequences of each alternative. Plants favor stability. Animals need mobility.
Mature Portions of the Root
Root hairs function only for several days, after which they die and degenerate. Absorption of water and minerals in this area is then greatly reduced but does not stop entirely. Within the endodermis, cells may remain unchanged, but usually there is a continued maturation in which a layer of suberin is applied over all radial surfaces, the inner tangential face, and sometimes even the outer tangential face. This can be followed by a layer of lignin and then more suberin (Figure 7-14D). This is an irregular process, and some cells complete it earlier than others; thus, in fairly mature parts of the root, it is possible to find occasional cells that have only Casparian strips. These cells are called passage cells because they were once thought to represent passageways for the absorption of minerals; it is now suspected that they are merely slow to develop.
The result of continued endodermis maturation is the formation of a watertight sheath around the vascular tissues. In older parts of the root, it functions to keep water in. The absorption of minerals in the root hair zone causes a powerful absorption of water, and a water pressure, called root pressure, builds up. If it were not for the mature endodermis, root pressure would force the water to leak out into the cortex of older parts of the root instead of moving up into the shoot. This is presumably also a function of the endodermis in rhizomes and stolons; if water leaked into the stem cortex and filled intercellular spaces, oxygen diffusion would be prevented and the tissues would suffocate.
Many important events occur at the endodermis. The cortex and epidermis, aside from root hairs, may be superfluous because shortly after the root hairs die, underlying cortex and epidermis often die also and are shed from the root. The endodermis becomes the root surface until a bark can form. This happens mostly in perennial roots that persist for several years. In apple trees, the cortex is shed as early as 1 week after the root hairs die, although it may persist for as long as 4 weeks; only the root tips have a cortex. The large fibrous roots of many monocots are strictly annual; they are replaced by new adventitious roots that form on new rhizomes or stolons. In these plants, the entire root dies.
![]() Origin and Development of Lateral Roots
Origin and Development of Lateral Roots
Lateral roots are initiated by cell divisions in the pericycle. Some cells become more densely cytoplasmic with smaller vacuoles and resume mitotic activity. The activity is localized to just a few cells, creating a small root primordium that organizes itself into a root apical meristem and pushes outward. As the root primordium swells into the cortex, the endodermis may be torn or crushed or may undergo cell division and form a thin covering over the primordium. As it pushes outward, the new lateral root destroys cells of the cortex and epidermis that lie in its path, ultimately breaking the endodermis (FIGURE 7-16). By the time the lateral root emerges, it has formed a root cap, and its first protoxylem and protophloem elements have begun to differentiate, establishing a connection to the vascular tissues of the parent root.
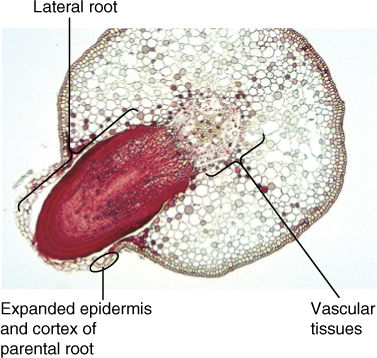
FIGURE 7-16 This young lateral root of willow (Salix) was initiated in the pericycle, next to a mass of protoxylem; its growth caused considerable damage to the parent root cortex, and it broke the surface open. Entry of pathogens is prevented by formation of wound bark around the tear it caused (×50).
Note how this differs from bud formation in shoots. Lateral roots are initiated deep within the root (endogenous origin), not at the surface as are axillary buds (superficial origin); they are formed in mature regions of the root, not right at the base of the apical meristem, and of course, they never develop into flowers.
![]() Other Types of Roots and Root Modifications
Other Types of Roots and Root Modifications
The type of root just described is the most common, generalized type, comprising some or all of the roots of most plants. In other species, some roots are modified and carry out different roles in the plant’s survival. Various structures and organizations are selectively advantageous for various functions.
Storage Roots
Roots frequently provide long-term storage for carbohydrates that accumulate during summer photosynthesis. In biennial species (which live for 2 years; for example, beets, carrots, and celery) and many perennials (Datura, Phlox, many daisies), roots are the only permanent organs: In autumn, most of the stem dies back to a few nodes located below ground at the top of the root. Carbohydrates stored in the root are used to produce a new shoot in the spring, when photosynthesis is impossible owing to the lack of leaves. It seems less economical in these species to winterproof the shoot than to replace it using nutrients stored in the roots. Annual plants can survive without such storage capacity.
Perennial plants with permanent shoots that do not die back also store significant amounts of nutrients within themselves during winter, but roots offer certain advantages. Being subterranean, roots are less available as food than are swollen, highly nutritious, easily visible stems. Roots also have a much more stable environment, subjected to less extreme changes in temperature and humidity, which may be important for survival of storage parenchyma cells.
Prop Roots
The stem of a monocot can become wider, with more vascular bundles, if it can produce adventitious roots that extend to the soil. In some cases, roots are capable of extensive growth through the air: In many palms, the exposed roots can be 20 to 50 cm long (FIGURE 7-17A). In the screwpine (Pandanus, a monocot, not a pine), they are often 3 or 4 meters long, and the roots may grow through the air for months before reaching the ground (FIGURE 7-17B-D). Less dramatic examples are found at the bases of corn and many other grasses. After prop roots do make contact with the soil, they transport additional nutrients and water to the stem. Just as importantly, they contract slightly and place some tension on the stem, thus acting as stabilizers, much like guy wires on tall television antennas. If the roots undergo secondary growth and become woody, they can be extremely strong supports, permitting a branch to extend even farther from the trunk without breaking or sagging (FIGURE 7-18A). In banyan trees (eudicots of the genus Ficus), prop roots and branches can spread and produce massive trees many meters in diameter. Roots of certain tropical trees become tall, plate-like buttress roots; their upper side grows more rapidly than other parts of the root. Buttress roots brace the trunk against being blown over by wind (FIGURE 7-18B).

FIGURE 7-17 (A) Many monocots like this small palm (Ptychosperma) produce adventitious roots near the base of the stem. These provide both extra absorptive capacity and extra stability. (B and C) Screwpine (Pandanus) produces extremely long adventitious prop roots that not only stabilize the large, heavy trunk but also bring water and minerals into the stem. (D) In older plants of screwpine, even the branches produce adventitious roots. This water and mineral supply to the branches bypasses the trunk, where vascular bundles are already conducting at full capacity.

FIGURE 7-18 (A) Banyan trees (eudicots) produce adventitious roots, which, as in monocots, provide increased support and absorptive capacity. Because the giant branches are supported along their entire length, they can become much larger and more extensive than branches that are supported only at the point of attachment to the trunk. (B) Plate-like buttress roots of Ficus subcordata. (C) These roots emerge from the basal portions of mangrove (Rhizophora mangle) stems in the Everglades National Park in Florida. Mangroves live in the intertidal zone, the band of seashore between high and low tide. At high tide, the roots are flooded with ocean water. This is a harsh environment because the seawater is so salty it can pull water out of most terrestrial plants and animals (that is why we cannot drink seawater), and the water should make the roots drown for lack of oxygen. Mangrove roots survive these conditions because their surface tissues keep seawater away from the delicate cells inside the root, and they have aerenchyma that lets oxygen diffuse into the root from the shoots.
Mangroves also have prop roots, but these seem to be selectively advantageous for other reasons (FIGURE 7-18C). The plants grow in intertidal marshes and are subjected to powerful water currents during storms and even normal tide changes; brace-like prop roots provide much more stability than a taproot system would. In addition, the aerial portion of the prop root is covered with numerous air chambers—lenticels—and its cortex is a wide aerenchyma. The subterranean portion of the root grows in a stagnant muck that has little or no oxygen; it is able to respire only because the aerenchyma permits rapid diffusion of oxygen from the aerial lenticels to the submerged root tissues. If mangrove roots were entirely subterranean, respiration would be extremely reduced. Few plants have adapted to the intertidal zone. Although ocean shores have existed for billions of years, mangroves became adapted to the intertidal zone only in the last several million years: There are no fossils of mangrove plants older than about 35 million years.
Aerial Roots of Orchids
Many orchids are epiphytic, living attached to the branches of trees. Their roots spread along the surface of the bark and often dangle freely in the air. Although these plants live in rainforests, the orchids are actually adapted to drought conditions. In the few hours when rain does not fall, the air and bark become dry and could easily pull water out of the orchid’s roots if there were no water-conserving mechanism. The root epidermis, called a velamen in these orchids, is composed of several layers of large dead cells that are white in appearance (FIGURE 7-19). Apparently, the velamen acts as a waterproof barrier, not permitting water to leave the sides of the root.

FIGURE 7-19 Aerial roots of orchids, which grow along the surface of tree bark, are covered by a white velamen that helps retain the water absorbed by the green root tip. These roots grow along the bark, not downward.
Contractile Roots
In Oxalis, Gladiolus, Crinum, and other plants (many with bulbs), roots undergo even more contraction than prop roots do. After extending through the soil and becoming firmly anchored, the uppermost portions slowly contract (FIGURE 7-20). Because the root is firmly fixed to the soil, the stem is pulled downward so that the base of the shoot is either kept at soil level or, in the case of bulbs, actually buried deeper. The contraction is caused by changes in the shape of cortex cells: They simultaneously shorten and expand radially, losing as much as one half to two thirds of their height. The vascular tissues buckle and become undulate but are able to continue conducting.

FIGURE 7-20 The roots of this hyacinth have contracted, causing their surfaces to become wrinkled.
Contractile roots may be more common than is generally appreciated. Many seeds germinate at or near the soil surface; root contraction may be the means by which the shoot becomes anchored in the soil. In bulbs, corms, rhizomes, and other subterranean stems, contractile roots can be important in keeping the stems at the proper depth.
Mycorrhizae
Roots of most species of seed plants (at least 80%) have a symbiotic relationship with soil fungi in which both organisms benefit. The associations are known as mycorrhizae (singular, mycorrhiza), and two main types of relationships are known. In nearly all woody forest plants, an ectomycorrhizal relationship exists in which fungal hyphae (slender, thread-like cells) penetrate between the outermost root cortex cells but never invade the cells themselves. Herbaceous plants have an endomycorrhizal association, in which hyphae penetrate the root cortex as far as the endodermis; they pass through the walls of the cortex cells but cannot pass the Casparian strip (FIGURE 7-21). They invade the cell but do not break the host plasma membrane or vacuole membrane. Inside the cell, they branch repeatedly, forming a small structure called an arbuscule; this fills with granules of phosphorus that later disappear as the phosphorus is absorbed by the plant. Other hyphae fill with membranous vesicles. The plant cells lack starch grains, presumably because sugars are being transferred to the fungus. The fungus is unable to live without the sugars from the plant, and in many cases, the plant is severely stunted if the fungus is killed. Apparently, this is a critical means of absorbing phosphorus into the roots, the mycorrhizal fungi being much more effective than root hairs.
All the modified roots mentioned to this point—storage roots, prop roots, contractile roots, aerial roots of orchids, and mycorrhizae-associated roots—have a structure virtually identical to that of the more common anchoring/absorbing roots described above. All grow by an apical meristem, and a cross-section through a mature region of each reveals epidermis, cortex, endodermis, pericycle, and vascular tissue. In nitrogen-fixing nodules and haustoria, however, the structure is highly modified.
Root Nodules and Nitrogen Fixation
For most plants, the scarcity of nitrogenous compounds in the soil is one of the main growth-limiting factors. Although nitrogen is abundant in air (78% of the atmosphere is N2), plants have no enzyme systems that can use that nitrogen. Only some prokaryotes can use N2 by incorporating it into their bodies as amino acids and nucleotides; when they die and decompose, the nitrogenous compounds are available to plants. The chemical conversion of atmospheric nitrogen into usable compounds is nitrogen fixation.
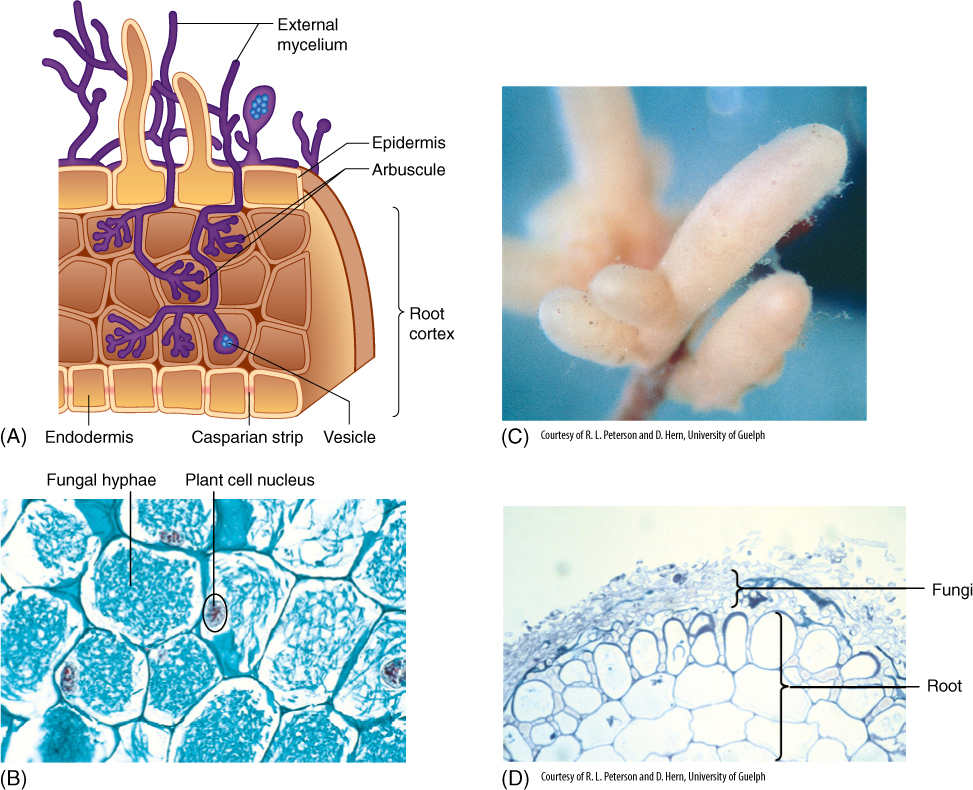
FIGURE 7-21 (A) Diagram of endomycorrhizae showing hyphae penetrating the root cortex and forming arbuscules and vesicles. (B) Root cells filled with fungal hyphae in the form of arbuscules. You can tell that this is a symbiotic fungus rather than a pathogenic one because the plant cells are healthy, with large normal nuclei (×220). (C) Roots of beech encased in ectomycorrhizae. The presence of the fungus stimulates the roots to be short and broad. (D) Transverse section of an ectomycorrhizal root showing that the fungal filaments are restricted to just the outermost part of the root; they do not penetrate the cortex.
In a small number of plants, especially legumes, a symbiotic relationship has evolved with nitrogen-fixing bacteria of the genus Rhizobium. Bacteria free in the soil secrete a substance that causes root hairs to curl sharply; a bacterium then attaches to the convex side of the hair and pushes into the cell by means of a tubelike invagination of the plant cell wall. The tube is an infection thread, and the bacterium sits in it. The infection thread, penetrating one cell after another, extends all the way into the root’s inner cortex, where adjacent cortical cells undergo mitosis and form a root nodule (FIGURE 7-22); the bacterium is released from the infection thread and enters the plant cell cytoplasm, where it proliferates rapidly, filling the plant cells with bacterial cells (known as bacteroids) capable of converting N2 into nitrogenous compounds that are released to the plant cells. Energy for this process is supplied by sugars from the legume root cells, so both the Rhizobium and the legume benefit.
The nodule may remain rather simple or may become complex, with a meristematic region, vascular tissue, and endodermis. Such nodules function for extended lengths of time. Numerous metabolic changes have also evolved: The critical bacterial enzymes are sensitive to oxygen, being immediately poisoned by even traces of free oxygen; however, it is the plant, not the bacterium, that produces a special chemical—leghemoglobin—that binds to oxygen and protects the bacterial enzymes. Root nodules represent a sophisticated symbiosis: Plants and bacteria both benefit. The bacteria are not damaging the plants, nor are the plants parasitizing the bacteria. Without the bacteria the complex development of nodules does not occur.
This process is often described in this way: “The bacterium supplies the plant with nitrogenous compounds and in return the plant gives it sugars.” This inaccurately suggests voluntary action, choice, and decision making on the part of both organisms. Instead, think in terms of natural selection: Because the bacterium receives sugars and other nutrients from the plant, any genetic mutation in the bacterium that makes the plant healthier and more vigorous is beneficial to the bacterium as well, whereas any mutation that is harmful to the plant—such as a bacterial plasma membrane that does not allow nitrogenous compounds to move to the plant—is deleterious to the bacterium. The same principle applies to the plant: Genetic mutations that aid the bacterium ultimately benefit the plant as well. We hypothesize that these organisms will continue to co-evolve and become more fully co-adapted, but this hypothesis will have to be checked by someone a few million years from now.
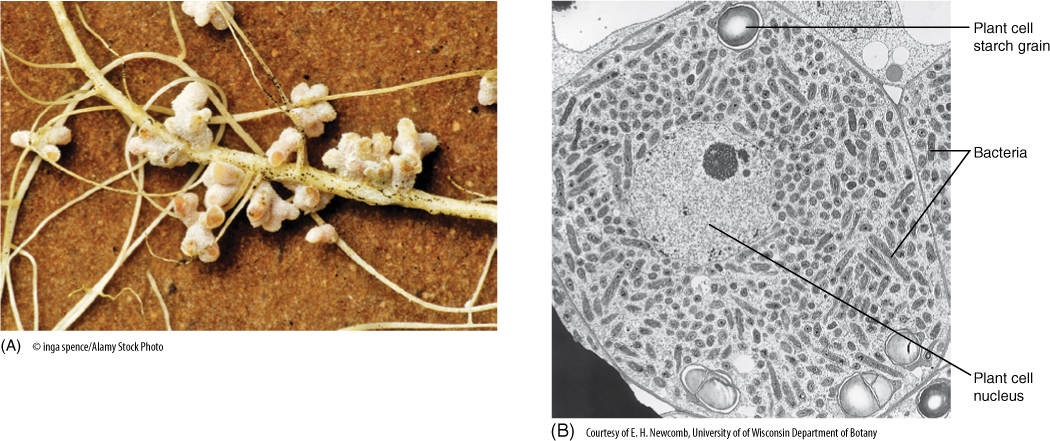
FIGURE 7-22 (A) The nodules on these alfalfa roots contain bacteria capable of absorbing atmospheric nitrogen and converting it to ammonia, which the plant can use to make amino acids. (B) Bacteria in an infected root nodule cell of cowpea, Vigna unguiculata (×5,900).
Working backward, we can imagine less sophisticated legumes and Rhizobium bacteria that did not work so well together but evolved to the present state as mutations occurred and survived by natural selection. Certain critical mutations must be rare, or perhaps several unusual mutations must occur almost simultaneously in plant and bacterium; this hypothesis is based on the observation that, although this symbiosis would probably be beneficial to virtually all plant species, only a few have it.
Haustorial Roots of Parasitic Flowering Plants
A number of angiosperms are parasites on other plants; because their substrate is the body of another plant, a normal root system would not penetrate the host or absorb materials from it effectively. Consequently, roots of parasitic plants have become highly modified and are known as haustoria (FIGURE 7-23). In most cases, very little root-like structure remains. Parasitism has evolved several times; thus, the structures that are termed haustoria are not all related to each other, and generalizations are difficult. Haustoria, however, typically must adhere firmly to their host either by secreting an adhesive or by growing around a small branch or root. Penetration occurs either by forcing a shaft of cells through the host’s dermal system or by expanding the haustorium radially, cracking the host’s epidermis. After penetration, cells of the parasite make contact with the host’s xylem. In many cases, both host and parasite cells divide and proliferate into an irregular mass of parenchyma, and then a column of cells differentiates into a series of vessel elements. This results in a continuous vessel from host to parasite constructed of cells of both. Surprisingly, many parasites attack only the xylem; they do not draw sugars from the host but carry out their own photosynthesis. Others do contact both xylem and phloem and perform little or no photosynthesis. For such parasites, mutations that result in the loss of leaves or vegetative stems are not disadvantageous.
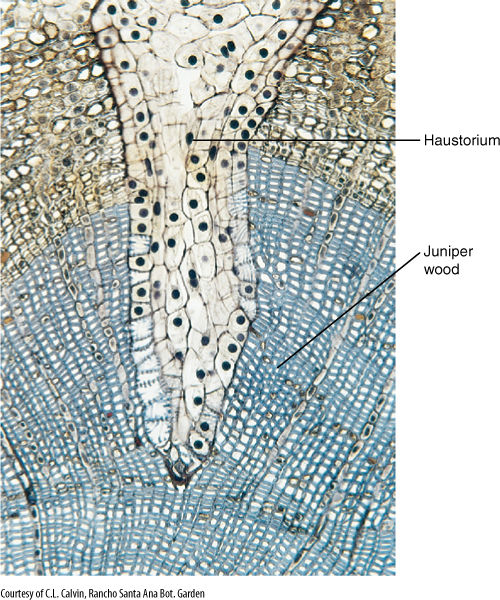
FIGURE 7-23 Transverse section of a branch of juniper being attacked by the haustorium (modified root) of a mistletoe (Phoradendron). The wood of the host juniper has been stained blue; the bark is tan. The haustorium is able to draw water and nutrients from the host vascular tissue.
A parasitic plant named Tristerix spends most of its life as nothing more than a diffuse root system—a haustorium—growing inside the body of its host plant, a cactus. The body is just a network of parenchyma cells with no stem, leaves, epidermis, cortex, or vascular tissues except when Tristerix produces flowers.
The typical organization of a root would be nonfunctional in haustoria: The root cap, root apical meristem, and cortex would all prevent vascular tissues of the parasite from contacting the appropriate tissues of the host. Conversely, haustoria are completely inadequate for growth in soil, however rich and moist it might be. For each substrate and each microhabitat, specific adaptations are selectively advantageous.
Roots of Strangler Figs
While young, plants of strangler figs (several species in the genus Ficus) grow as epiphytes perched on a branch of a host tree. Birds eat fruits of strangler figs (FIGURE 7-24) and then deposit the seeds on branches of other trees. When the strangler fig seed germinates, its roots cling to the bark of the host tree branch and then grow rapidly downward, hugging the trunk of the host tree. For some months or even years the roots have no contact with the soil, so they must absorb nutrients from rainwater that runs down the trunk of the host tree. Finally, the strangler fig’s roots do reach soil and then penetrate it rapidly and branch profusely; upper portions of the roots enlarge and become woody. Strangler fig roots branch and grow at various angles, encircling the host tree’s trunk, and the roots fuse to each other wherever they meet. Before long the trunk of the host tree is encased in numerous roots of the strangler fig. Typically, the host tree finally dies and rots away, leaving the strangler fig as a self-supporting tree with its root—shoot junction high in the air.
It was believed that the tight network of strangler fig roots prevented the host tree from growing thicker, and thus the strangler fig killed the host by “strangling” it. That may be true in some cases, but it seems that more often the fig produces so many leafy branches that it shades the host tree so much it cannot photosynthesize; the host tree dies from lack of light.

FIGURE 7-24 This tree in the Everglades National Park is encased in the roots of a strangler fig. The large “branch” of the fig at the upper left is actually the trunk of the fig, which is the point where the seed germinated. All portions below that are roots of the strangler fig. It would be easy to mistakenly assume the fig is a vine and the parts encasing the host tree are stems of the fig, but a close examination of the anatomy shows these are roots not branches. If this were really a vine, we would expect to see leafy branches emerging from many areas in this photo.
 At the Next Level
At the Next Level
1. Modified roots. This chapter touched briefly on root nodules and nitrogen-fixing bacteria and also on mycorrhizal associations of roots and phosphate-absorbing fungi. Much more is known about both these topics and you can learn a great deal by exploring on your own. Other modified roots for you to investigate are the coralloid roots of cycads (they contain nitrogen-fixing cyanobacteria) and proteoid roots (they branch profusely when they enter pockets of mineral-rich soil). Roots of many epiphytes are modified such that their growth is not controlled by gravity but instead is guided by contact with host tree bark.
2. Root ecology. Root systems vary greatly in size, branching, orientation, and so on. Some roots spread horizontally in the topsoil, others grow downward 30 m or more. Certain plants have much more root mass than shoot mass, whereas some large globular cacti have only a few scrawny, thread-like roots. We are starting to understand the factors that influence root growth, so this is a rich area for you to explore. Especially important are the enormous root systems of prairie grasses.
3. Aerenchyma and flooding. Many river valleys flood during spring time, causing roots to be flooded for days or weeks. This reduces the amount of oxygen available, and the risk of suffocation is great. Some roots and stems respond by producing aerenchyma under such conditions, allowing more rapid diffusion of any oxygen present. Marsh plants such as mangroves (Avicennia germinans) and bald cypress (Taxodium distichum) automatically produce “knees” (technically called pneumatophores; pronounced new MAT oh fours), modified aerenchymatous roots that grow upward into the air.
SUMMARY
1. Most roots have a variety of functions, including anchorage, absorption, and hormone production. Other roots may be specialized for nutrient storage, vegetative reproduction, or surviving harsh conditions, some even being modified into spines.
2. Roots have a root cap but nothing equivalent to leaves, nodes, internodes, or buds.
3. Adventitious roots form in organs other than other roots or the embryo; they are especially important in stoloniferous and rhizomatous plants.
4. Roots, like shoots, elongate by localized growth (apical meristems). Only the root tip and zone of elongation must slide between soil particles.
5. Root hairs greatly increase the absorptive surface area of the root system, and the carbonic acid that results from their respiration helps release minerals from soil particles.
6. An endodermis with Casparian strips prevents minerals from moving from the soil solution into the xylem. To enter the xylem, minerals must at some point cross a plasma membrane.
7. Lateral roots arise in the pericycle, deep inside the root, unlike axillary buds in stems, which arise in the outermost stem tissues.
8. Prop roots provide additional stabilization and transport for certain plants with narrow stems. Contractile roots aid in burying certain bulbs, corms, and rhizomes. Haustoria are modified roots that penetrate the tissues of host plants.
9. Most plants absorb much of their phosphorus from mycorrhizal fungi that form an extensive network both in the soil and within the root cortex.
10. A small number of plant species form symbiotic relationships with nitrogen-fixing prokaryotes. The bacteria or cyanobacteria often live in root nodules, passing nitrogen compounds to the plant and receiving carbohydrates and minerals.
IMPORTANT TERMS
adventitious roots
branch roots
buttress roots
Casparian strips
ectomycorrhizal relationship
endodermis
endomycorrhizal association
fibrous root system
haustoria
infection thread
lateral roots
mycorrhiza
nitrogen fixation
pericycle
prop roots
quiescent center
radicle
root apical meristem
root cap
root hairs
root nodule
taproots
zone of elongation
REVIEW QUESTIONS
1. The “Concepts” section of this chapter states that most roots have three functions. What are they? Some roots have other functions in addition to these three, or instead of them. Name an example of at least one species in which roots do the following:
a. Store carbohydrate during winter
b. Produce shoot buds that can act as new plants
c. Grow out of the trunk and harden into spines
d. Act as holdfasts
e. Attack other plants and draw water and nutrients out of them
2. What are the two types of root systems? Give several examples of plants that have each type. Which type is associated with nutrient storage in biennial species like carrots and beets? Which is associated with rhizomes and stolons?
3. Roots must have an enormous absorptive surface. Why do plants have a highly branched root system instead of just one long root?
4. Even before a seed germinates, it already has a root; what is the name of this embryonic root? In eudicots, what does this embryonic root usually develop into? In most monocots, this embryonic root does a strange thing during or immediately after germination. What does it do?
5. What is an adventitious root? In a monocot rhizome such as that of irises and bamboo, are adventitious roots common or rare? If a bamboo rhizome grows 100 feet underground and then sends up an aerial branch, where does that branch get its water—from the roots 100 feet away or from adventitious roots right at its base (Hint: look at Figure 7-5E; you might also want to look at Question 22)?
6. What does the root cap do? Do you think this structure would have evolved if roots all grew in air like shoots do, rather than growing through dirt?
7. Roots have localized growth; the only parts of the root that become longer are the root apical meristem and the __________________ of ___________________, a region only a few millimeters long within which the cells undergo division and expansion.
8. What would happen to root hairs if they formed in the zone of elongation? Would this happen if roots were growing in air or in water?
9. Root hairs greatly increase the root’s surface area. In rye, a single plant has been found to have __________________ lateral roots with ________________ km of root length and a surface area of _________________ m2. Convert the length to miles (1 mile = 1.6 km) and the area to square feet (1 foot = 0.3 m).
10. Root hairs are narrower than roots. A root is usually at least __________________ mm in diameter, but root hairs are only approximately ___________________ _____ mm in diameter. Look at Figure 7-9. Could either roots or root hairs enter the pores represented by the red part of the chart? Could either or both enter the gray part? The white part? Most roots grow near the soil surface, in the upper 20 cm, where most of the soil pores are between 0.2 to 60 mm wide. If a root did not produce any root hairs at all, could the plant obtain water and minerals from these abundant, narrow pores?
11. Which part of the root detects gravity? Which organelles enable them to do this?
12. The use of a radioactive precursor of DNA can demonstrate that the central cells of the root apical meristem are not synthesizing DNA. This mitotically inactive central region is called the _______________ _______________. It is now believed that the central cells act primarily as a reserve of ______________ ___________.
13. If you could examine a transverse section through the zone of elongation in a root, would you see any mature cells? Would there be any fully differentiated epidermis cells or vascular tissues? What would the outermost cells be called, if not epidermis? In the center of the zone of elongation, what would be the tissue that would later differentiate into xylem and phloem?
14. Examine Figure 7-13. What are the four diffusion paths that a molecule might follow as it travels through the root epidermis and cortex? Why does this diagram stop at the endodermis? What happens at the endodermis?
15. Describe the shape of the endodermis. Is it a flat plane, a cylinder, a single ring of cells? What are the Casparian strips, and on which cell walls are they located?
16. Draw cross-sections of a root showing its structure at three levels: the mature region, the root hair zone, and the zone of elongation. At which level is the endodermis complete with Casparian strips?
17. Because Casparian strips are impermeable, minerals can cross the endodermis only if the endodermal ________________ absorb them from the intercellular spaces or from cortical cells. Many _________________ minerals can be excluded by the endodermis.
18. Examine Figure 7-14. The roots of eudicots are usually noticeably different from those of monocots in transverse section. Which has a relatively wide set of vascular tissues? Which has endodermis cells with very thick walls? Which has ground tissues in the center, and which has metaxylem in the center?
19. Which part of the root produces the primordia for lateral roots? How does the vascular tissue of the lateral root connect with that of the parent root?
20. Some perennial plants store significant amounts of nutrients in the stem during winter, but roots offer certain advantages. Describe two of these advantages.
21. Animals typically use fats in adipose tissues for long-term energy storage, whereas plants use starch in roots. How do animals benefit from using fat? How do plants benefit from using starch? Name two plants that store energy for many years. How long is long-term storage for these species? What two plant parts often use fats and why? Storage tissue in enlarged roots is vascularized. How is that important to the plant?
22. Look at the prop roots of screwpine in Figure 7-17D (screwpine is a monocot). Because the branches produced their own adventitious roots (the prop roots), can the branches obtain water and minerals without depending on the xylem in the trunk (think about Question 5)?
23. Look at the prop roots of banyan trees in Figure 7-18. These prop roots supply extra water to the branch, but how else do they help the branches?
24. What are contractile roots? They are especially common in plants with what types of shoots (e.g., in rhizomes, vines, or what)?
25. What is a mycorrhizal association? What benefit does the plant derive from the association?
26. Describe the structure of a nitrogen-fixing nodule; consider especially the relationship with the plant’s vascular tissue.
27. Root nodules are ________________ associations. Plants and bacteria both benefit. The bacteria are not ______________ the plants, nor are the plants ________________________ the bacteria.
28. Many plants are parasitic on other plants. These parasitic plants have modified roots called ___________________. Describe some of the ways that this parasitic root penetrates the host stem or root. Do these parasitic roots have a root cap, a root apical meristem, or a cortex?
29. Except when it produces flowers, what is the body of the parasitic plant Tristerix? What tissues does this plant body lack, except when it produces flowers?
30. Imagine a plant that has ten roots, each 1 cm long. What is the total length of the root system? Imagine that at the tip of each root are ten lateral roots, each 1 cm long. Now what is the total length of the root system? What is the maximum distance a water molecule must travel from the farthest root tip to the base of the shoot? Distance traveled increases as a simple addition, but total absorbing capacity increases exponentially. Why is the length that is traveled important? Consider the friction of moving through tracheids and vessel elements.
Design Credits: Hummingbird: © Tongho58/Moment/Getty; Green plant cells: © ShutterStock, Inc./Nataliya Hora; Purple tulip: © ShutterStock, Inc./Marie C Fields; Dandelion: © ShutterStock, Inc./danielkreissl; Poppy: © ShutterStock, Inc./Saruri; Plant icon: © ShutterStock, Inc./Vector; Digging man icon: © ShutterStock, Inc./Z-art