CONCEPTS IN BIOLOGY
PART IV. EVOLUTION AND ECOLOGY
13. Evolution and Natural Selection
13.9. Gene-Frequency Studies and the Hardy-Weinberg Concept
In the early 1900s, an English mathematician, G. H. Hardy, and a German physician, Wilhelm Weinberg, recognized that it was possible to apply a simple mathematical relationship to the study of gene frequencies. Their basic idea was that, if certain conditions existed, gene frequencies would remain constant and the distribution of genotypes could be described by the relationship p2 + 2pq + q2 = 1, where p2 represents the frequency of the homozygous dominant genotype, 2pq represents the frequency of the heterozygous genotype, and q2 represents the frequency of the homozygous recessive genotype. Constant gene frequencies over several generations would imply that evolution was not taking place. Changing gene frequencies would indicate that evolution was taking place.
The conditions necessary for gene frequencies to remain constant are the following:
1. Mating must be completely random.
2. Mutations must not occur.
3. The migration of individual organisms into and out of the population must not occur.
4. The population must be very large.
5. All genes must have an equal chance of being passed on to the next generation. (Natural selection is not occurring.)
The Hardy-Weinberg concept states that gene frequencies will remain constant if these five conditions are met. The concept is important, because it allows a simple comparison of allele frequency to indicate if genetic changes are occurring within a population. Two different populations of the same species can be compared to see if they have the same allele frequencies, or populations can be examined at different times to see if allele frequencies are changing.
|
Possible female gametes |
|||
|
A = 0.6 |
a = 0.4 |
||
|
Possible male gametes |
A = 0.6 |
Genotype of offspring AA = 0.6 x 0.6 = 0.36 = 36% |
Genotype of offspring Aa = 0.6 x 0.4 = 0.24 = 24% |
|
a = 0.4 |
Genotype of offspring Aa = 0.4 x 0.6 = 0.24 = 24% |
Genotype of offspring aa = 0.4 x 0.4 = 0.16 = 16% |
|
Determining Genotype Frequencies
It is possible to apply the Punnett square method from chapter 10 to an entire gene pool to illustrate how the Hardy-Weinberg concept works. Consider a gene pool composed of only 2 alleles, A and a. Of the alleles in the population, 60% (0.6) are A and 40% (0.4) are a. In this hypothetical gene pool, we do not know which individuals are male or female and we do not know their genotypes. With these allele frequencies, how many of the individuals would be homozygous dominant (AA), homozygous recessive (aa), and heterozygous (Aa)? To find the answer, we treat these alleles and their frequencies as if they were individual alleles being distributed into sperm and eggs. The sperm produced by the males of the population will be 60% (0.6) A and 40% (0.4) a. The females will produce eggs with the same relative frequencies. We can now set up a Punnett square as shown at the top of this page. The Punnett square gives the frequency of occurrence of the three possible genotypes in this population: AA = 36%, Aa = 48%, and aa = 16%.
If we use the relationship p2 + 2pq + q2 = 1, p2 is the frequency of the AA genotype, 2pq is the frequency of the Aa genotype, and q2 is the frequency of the aa genotype. Then, p2 = 0.36 and p would be the square root of 0.36, which is 0.6—our original frequency for the A allele. Similarly, q2 = 0.16 and q would be the square root of 0.16, which is 0.4. In addition, 2pq would equal 2 x 0.6 x 0.4 = 0.48. If this population were to reproduce randomly, it would maintain an allele frequency of 60% A and 40% a alleles. It is important to understand that Hardy-Weinberg conditions rarely exist; therefore, there are usually changes in gene frequency over time or genetic differences in separate populations of the same species. If gene frequencies are changing, evolution is taking place.
Why Hardy-Weinberg Conditions Rarely Exist
Random mating does not occur for a variety of reasons. Many species are divided into small local populations that are isolated from one another and mating with individuals in other local populations rarely occurs. In human populations, these isolations may be geographic, political, or social. In addition, some individuals may be chosen as mates more frequently than others because of the characteristics they display. Therefore, the Hardy-Weinberg conditions are seldom met, because non-random mating is a factor that leads to changing gene frequencies.
Spontaneous mutations occur. Totally new kinds of alleles are introduced into a population, or 1 allele is converted into another, currently existing allele. Whenever an allele is changed, 1 allele is subtracted from the population and a different allele is added, thus changing the allele frequency in the gene pool. Mutations in disease-causing organisms may have significant impacts (Outlooks 13.3).

Immigration and emigration of individual organisms are common. When organisms move from one population to another, they carry their genes with them. Their genes are subtracted from the population they left and added to the population they enter, thus changing the gene pool of both populations. It is important to understand that migration is common for plants as well as animals. In many parts of the world, severe weather disturbances have lifted animals and plants (or their seeds) and moved them over great distances, isolating them from their original gene pool. In other instances, organisms have been distributed by floating on debris on the surface of the ocean. As an example of how important immigration and emigration is, consider the tiny island of Surtsey (3 km2), which emerged from the sea as a volcano near Iceland in 1963 and continued to erupt until 1967. The new island was declared a nature preserve and has been surveyed regularly to record the kinds of organisms present. The nearest possible source of new organisms is about 20 kilometers away. The first living thing observed on the island was a fly seen less than a year after the initial eruption. By 1965, the first flowering plant had been found and, by 1996, 50 species of flowering plants had been recorded on the island. In addition, several kinds of sea birds nest on the island.
Populations are not infinitely large, as assumed by the Hardy-Weinberg concept. If numbers are small, random events to a few organisms might alter gene frequencies from what was expected. Consider coin flipping as an analogy. Coins have two surfaces, so, if you flip a coin once, there is a 50:50 chance that the coin will turn up heads. If you flip two coins, you may come up with two heads, two tails, or one head and one tail. Only one of these possibilities gives the theoretical 50:50 ratio. To come closer to the statistical probability of flipping 50% heads and 50% tails, you would need to flip many coins at the same time. The more coins you flip, the more likely it is hat you will end up with 50% of all the coins showing heads and the other 50% showing tails. The number of coins flipped is important. The same is true of gene frequencies. The smaller the population, the more likely it is that random events will alter the gene pool—that is, the more likely it is that genetic drift will occur.

Finally, genes are not all equally likely to be passed to the next generation. It is important to understand that genes differ in their value to the species. Some genes result in characteristics that are important to survival and reproductive success. Other genes reduce the likelihood of survival and reproduction. For instance, many animals have cryptic color patterns that make them difficult to see. The genes that determine the cryptic color pattern would be selected for (favored), because animals that are difficult to see are not killed and eaten as often as those that are easy to see. Recall that albinism is the inability to produce pigment, so that the individual’s color is white. White animals are conspicuous, so we might expect them to be discovered more easily by predators (figure 13.12). Because not all genes have equal value, natural selection will operate and some genes will be more likely to be passed on to the next generation.

FIGURE 13.12. Albino Animals Are More Conspicuous
Pythons rely on camouflage coloring to help them catch prey. This albino form (a) is more likely to be spotted than a member of the species with normal coloration (b).
OUTLOOKS 13.3
The Reemerging of Infectious Diseases
Infectious diseases caused by bacteria, viruses, fungi, and parasitic worms continue to be a major cause of suffering and death throughout the world. They are the third leading cause of death in the United States. Reemerging infectious diseases (for example, diphtheria, malaria, whooping cough) are diseases that were once major health concerns but then declined significantly. However, they are beginning to increase in frequency.
The reemergence of many kinds of infectious diseases is the result of two primary factors: our failure to immunize against these diseases, and evolutionary changes in the microbes. People who are not being immunized against diseases are susceptible and may become ill with the disease or become asymptomatic carriers of the microbe. A further contributing factor to the reemergence of old diseases is the increased number of people with poorly functioning immune systems. HIV/ AIDS has created a huge population of people with compromised immune systems. Famine and malnutrition also impair the immune system. War and the crowding that occurs in refugee camps and prisons enhance the easy spread of disease.
The reemergence of some diseases is also the result of evolution. Mutations are necessary if evolution is to take place. As parasites and their hosts interact, they constantly react to each other in an evolutionary fashion. Hosts develop new mechanisms to combat parasites, and parasites develop new mechanisms to overcome the hosts' defenses (for example, antibiotic resistance).
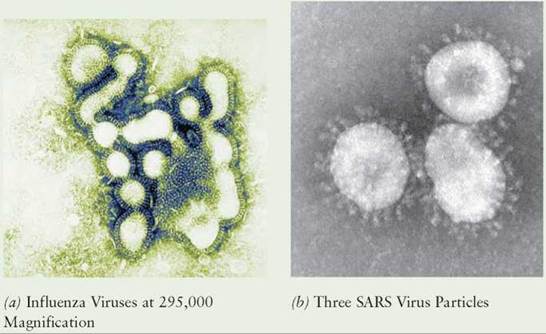
One of the mechanisms viruses use is a high rate of mutation. This ability to mutate has resulted in many new, serious human diseases. In addition, many new diseases arise when viruses that cause disease in another animal are able to establish themselves in humans. Many kinds of influenza originated in pigs, ducks, or chickens and were passed to humans through close contact with infected animals or by eating infected animals. In many parts of the world, these domesticated animals live in close contact with humans (often in the same building) making conditions favorable for the transmission of animal viruses to humans.
Each year, mutations result in new varieties of influenza and colds, which pass through the human population. Occasionally, the new varieties are deadly. In 1918, a new variety of influenza virus originated in pigs in the United States and spread throughout the world. During the 1918-1919 influenza pandemic that followed, 20 to 40 million people died. In 1997 in Hong Kong, a new kind of influenza was identified that killed 6 of the 18 people infected. When public health officials discovered the virus had come from chickens, they ordered the slaughter of all the live chickens in Hong Kong, which stopped the spread of the disease.
In early 2003, an outbreak of a new viral disease, known as severe acute respiratory syndrome (SARS), originated in China. SARS is a variation of a coronavirus, a class of virus commonly associated with the common cold, but it causes severe symptoms and, if untreated, can result in death. In June 2003, the SARS virus was isolated from an animal known as the masked palm civet (Paguma larvata). This animal is used for food in China and is a possible source of the virus that caused SARS in humans. However, other animals have also tested positive for the virus. The disease spread rapidly to several countries as people traveled by airplane from China to other parts of the world. A recognition of the seriousness of the disease and the isolation of infected persons prevented further spread, and this new disease was brought under control. However, if the virus still exists in some unknown wild animal host, it could reappear in the future.
In addition to cold and influenza, other kinds of diseases often make the leap from nonhuman to human hosts. The swine flu virus outbreaks of 2009 were traced to a population of pigs in Mexico. It is likely that genetic mixing occurred in the pigs' cells, resulting in a new kind of virus containing bird, human, and swine genes. The emergence of these new kinds of viruses enables them to more easily move from one species to another.
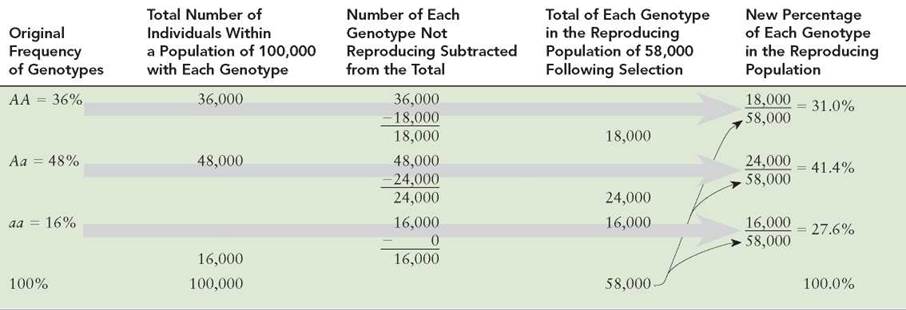
Using the Hardy-Weinberg Concept to Show Allele-Frequency Change
We can return to our original example of alleles A and a to determine how natural selection based on differences in survival can result in allele-frequency changes in only one generation. Again, assume that the parent generation has the following genotype frequencies: AA = 36%, Aa = 48%, and aa = 16%, with a total population of 100,000 individuals. Suppose that 50% of the individuals having at least one A allele do not reproduce because they are more susceptible to disease. The parent population of 100,000 would have 36,000 individuals with the AA genotype, 48,000 with the Aa genotype, and 16,000 with the aa genotype. Because only 50% of those with an A allele reproduce, only 18,000 AA individuals and 24,000 Aa individuals will reproduce. All 16,000 of the aa individuals will reproduce, however. Thus, there is a total reproducing population of only 58,000 individuals out of the entire original population of 100,000. What percentage of A and a will go into the gametes produced by these 58,000 individuals?
The percentage of A-containing gametes produced by the reproducing population will be 31% from the AA parents and 20.7% from the Aa parents (table 13.2). The frequency of the A allele in the gametes is 51.7% (31% + 20.7%). The percentage of a-containing gametes is 48.3% (20.7% from the Aa parents plus 27.6% from the aa parents). The original parental allele frequencies were A = 60% and a = 40%. These have changed to A = 51.7% and a = 48.3%. More individuals in the population will have the aa genotype, and fewer will have the AA and Aa genotypes.
TABLE 13.2. Differential Reproduction
The percentage of each genotype in the offspring differs from the percentage of each genotype in the original population as a result of differential reproduction.
If this process continued for several generations, the allele frequency would continue to shift until the A allele became rare in the population (figure 13.13). This is natural selection in action. Differential reproduction rates have changed the frequency of the A and a alleles in this population.
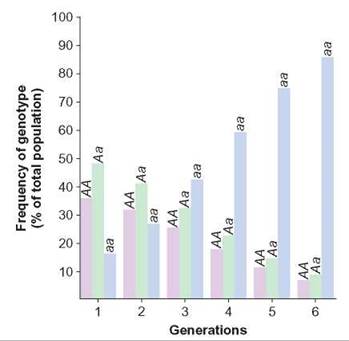
FIGURE 13.13. Changing Allele Frequency
If 50% of all individuals with the genotypes AA and Aa do not reproduce in each generation, the frequency of the a allele will increase, whereas the frequency of the A allele will decrease. Consequently, the aa genotype will increase in frequency, whereas that of the AA and Aa genotypes will decrease.
13.9. CONCEPT REVIEW
17. A gene pool has equal numbers of alleles B and b. Half of the B alleles mutate to b alleles in the original generation. What will the allele frequencies be in the next generation?
18. Hardy-Weinberg is a theoretical concept that describes gene frequencies. List five reasons why the conditions of the Hardy-Weinberg concept are rarely met.
19. The smaller the population, the more likely it is that random changes will influence gene frequencies. Why is this true?