Must Know High School Biology - Kellie Ploeger Cox 2019
PART TWO Cells
Signal Transduction: Cell Communication
MUST ![]() KNOW
KNOW
![]() In signal transduction, the original signal never enters the cell.
In signal transduction, the original signal never enters the cell.
![]() The purpose of signal transduction is to move the message into the cell in order to elicit a response.
The purpose of signal transduction is to move the message into the cell in order to elicit a response.
![]() The G protein-coupled receptor is found in all eukaryotes.
The G protein-coupled receptor is found in all eukaryotes.
Cells are in constant communication with their surroundings, and the signals they receive are frequently in the form of chemicals. Considering that the cell membrane provides a barrier separating the inside of the cell from the surrounding environment, the signal must somehow pass through the membrane and into the cell’s interior. If the chemical signal is lipid soluble, it is easy! The saying “like-dissolves-like” applies here, because if the signal is a lipid and the membrane is made of lipids, the signal molecule will just pass through. More often than not, however, the signal is made of protein (many hormones are proteins) and thus cannot pass through the cell membrane. For example, the hormone epinephrine (also called adrenaline) is responsible for telling cells in the liver and muscles to release glucose from the storage polymer glycogen. If epinephrine is a protein and cannot pass into the cell, how is it supposed to cause the hydrolysis of glycogen within the target cell? As our must know concept states, the original signal never enters the cell. The answer to this problem lies in the process of signal transduction.
Three stages are needed for a signal molecule to illicit a cellular response without actually entering the cell:
1. Reception
2. Transduction
3. Response
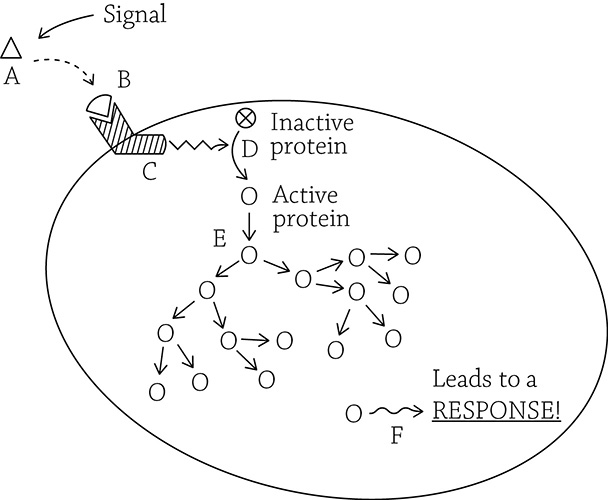
Signal transduction overview
Let’s take a look at what’s happening in the figure above. The initial signal (A) never enters the cell. Instead, it binds (B) with a transmembrane receptor protein. As soon as the receptor binds the signal, the inside portion changes shape (C) and activates another protein (D). The activated protein activates other proteins in a chain reaction (E). Eventually, the swarm of activated proteins lead to some sort of cellular response (F).
Step 1: Reception
Reception is what it sounds like: the signal is “received” by the cell. The cell hears the signal when the ligand binds with a membrane receptor. The receptor is a transmembrane protein that spans the width of the cell membrane. The part that extends to the outside of the cell has a specific shape that fits perfectly with the ligand. This is important because it ensures only the correct target cell (with the correct receptor) actually hears and responds to the signal. Something happens at this step that magically moves the message into the cell without the actual signal getting through. Once the transmembrane receptor binds the signal, it changes shape.
![]()
Here is another example of a protein’s tertiary (3D) structure being very important to its function. Not only that, the ability of a protein to be flexible and change shape is also very important. An enzyme’s function relies on these structural qualities, as does our signal transduction membrane receptors.
When the inside portion of the receptor protein changes shape, it essentially moves the signal into the cell. But before we get to the next step of transduction, let’s look at two specific types of receptors: G protein-coupled receptor and the ligand-gated ion channel.
G Protein-Coupled Receptor
This is probably the most important type of receptor protein to study. The G protein-coupled receptor is the main signaling pathway in a wide variety of eukaryotes, which suggests it evolved very early and has been conserved through evolutionary history.
![]()
The phrase “conserved through evolutionary history” means an adaption was so helpful to the survival of the organism that the trait was maintained through countless rounds of natural selection. If a species has a characteristic that isn’t very helpful (or, even worse, is disadvantageous), then those individuals would not survive and pass down those characters to the next generation. The G protein-coupled receptor, however, was so pivotal and critical to proper cellular functioning, it was maintained through many generations. Since the G protein-coupled receptor first arose in an early eukaryotic ancestor, it is now in almost all eukaryotes: fungi, protists, and animals (and maybe plants, though this is not yet known for sure).
When a signal molecule binds to the “outside-the-cell” portion of the G protein-coupled receptor, the receptor protein becomes activated and changes its shape on the cytoplasmic (inside) portion. There is a separate protein called (appropriately enough) a G protein that hangs out next to the cytoplasmic portion of the receptor. Once the receptor is activated, it also activates its buddy, the G protein. Finally, this activated G protein slides along the cell membrane over to an inactive enzyme and activates it! The reaction this enzyme then catalyzes depends on the specific cell type.
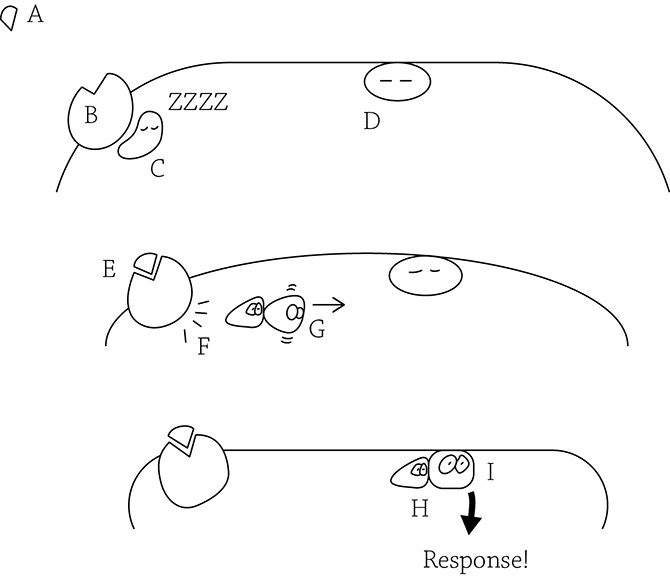
G protein-coupled receptor
As we can see in the figure above, if the signal (A) has not been yet grabbed by the receptor (B), then the partner G protein (C) remains quiet and inactive. There is an inactive enzyme (D) waiting for its own signal. Once the receptor binds the signal (E), the receptor activates the G protein (F), and the activated G protein slides along the inner membrane (G). The G protein binds to the awaiting enzyme (I), activating it and leading to a response.
Because it is the most abundant type of mammalian cell-surface receptor, there are numerous cool examples of G protein-linked receptors in action. For example, some bacterial toxins wreak havoc with G protein function and cause diseases such as cholera, whooping cough, and botulism. Another cool (and less deadly) example is some folks’ ability to taste the chemical phenylthiocarbamide (PTC). The gene TAS2R38 codes for a G protein-coupled taste receptor located in the cells of the taste buds. This receptor binds to the chemical PTC, resulting in a large number of proteins being made that stimulate neighboring neurons to send a “bitter” signal to the brain. For many people, the gene that codes for the PTC receptor is mutated. These people don’t “hear” the bitter signal of PTC. A long time ago (during our hunting/gathering days), having the ability to easily detect bitterness could help reduce the chances of ingesting toxic plant materials (plant toxins often have a bitter taste). Nowadays, a robust ability to detect the PTC signal tends to influence our food preferences and make some items—such as broccoli and rhubarb—unpalatable and bitter.
Ligand-Gated Ion Channel
The ligand-gated ion channel acts as a doorway for ions such as Na+ or Ca2+. The key to unlocking this door is the signal molecule, which itself does not pass into the cell. The “message” is instead the influx of ions that just occurred because the signal molecule swung open the gate!
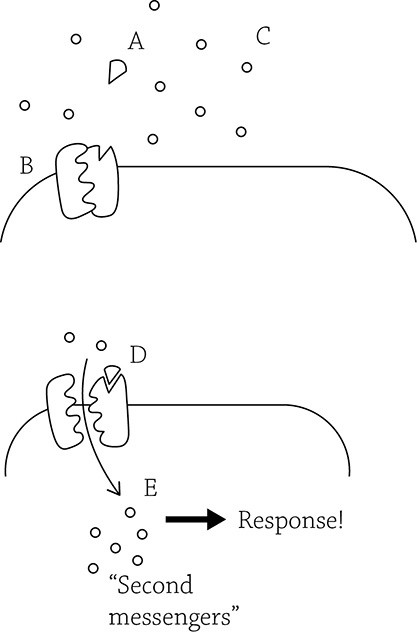
Ligand-gated ion channel
From the figure above, we can see that before the signal (also called the ligand, A) binds the receptor, the receptor remains closed (B), blocking the entry of extracellular ions (C). Once the signal binds (D), the receptor gate swings open and the ions (also called “second messengers”) rush in (E), leading to a response.
Ligand-gated ion channels are key to nervous system function. When a signal must traverse the open space between two adjacent neurons (the space is called the synapse), the chemical messenger is a neurotransmitter. Neurotransmitters aren’t allowed into the cell, and instead they latch onto ligand-gated ion channel receptors. Once this happens, it signals the second neuron that it needs to “fire” in the forms of a wave of positive charge that whooshes down the cell. This wave occurs because the ligand-gated ion channels open and Na+ flows in (more about neuron function in Chapter 26).
Step 2: Transduction
Now that the cell has “heard” the signal, it must be amplified. This is a super helpful step because even if only one, single, signal molecule managed to find its way to the target cell, it is enough to create a large coordinated response. Once the message has been moved into the cell, it can now be amplified and made louder through transduction. The transduction step is based on a cascading series of molecular interactions that boosts the signal as it is moved through the cell. It’s like a cascade of dominoes!
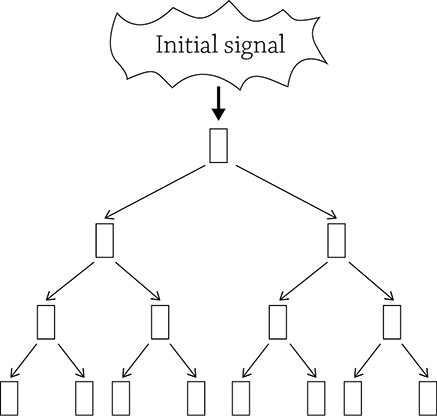
Cascades amplify a signal
Usually, each step involves activating a protein and making it change its shape. This shape change is caused by phosphorylation, or adding a phosphate group to a protein. Phosphorylation (and dephosphorylation) of proteins is a widespread means of regulating the activity of the proteins. The enzyme responsible for transferring a phosphate group from ATP and sticking it onto the target protein is called a kinase.
Protein kinases are enzymes that transfer phosphate groups from ATP to a protein. Protein kinases play a major role in signal transduction pathways. They often activate other protein kinases and create a cascade, just like the domino analogy. If you swap out the dominos with protein kinase enzymes, you have created a phosphorylation cascade.
Since adding a phosphate changes a protein from inactive to active, you also need a way to stop the activation. Protein phosphatases are enzymes that remove phosphate groups from proteins, inactivating the pathway. This is important because if the initial signal is no longer present, you don’t need the response to keep going!
![]()
If you see an enzyme and it has “kinase” in its name, it’s a hint to the reaction it catalyzes. Kinase enzymes specialize in transferring phosphate groups to (or from) ATP. The glycolytic pathway alone has many different kinase enzymes.
Step 3: Response
Finally, we get to the point of all this … the response. It can be any cellular activity that occurs in the nucleus or in the cytoplasm. Many signaling pathways result in a gene being expressed, because the initial signal told the cell “Make this specific protein! Now!” The pathway may instead result in the activity of a protein that’s already waiting in the cytoplasm. Earlier, I mentioned epinephrine as an example signal, and the response was the activation of an enzyme that cleaved apart glycogen in order to release glucose molecules. Your liver cells shouldn’t send out a flood of glucose molecules unless it hears the proper signal (epinephrine) to do so.
Yeast and Shmoo Love
Here is a romantic example of the entire process of signal transduction: yeast mating. And yes, yeast can “mate.” Usually, they reproduce by asexually budding, but what’s the fun (and genetic variation) in that? The simple baker’s yeast—Saccharomyces cerevisiae—has the option to mix up its alleles (and thus increase variation) by fusing with another cell. Each yeast cell has two mating types: a and α. To ensure maximum variation, yeasty beasties prefer to mate with the other mating type: a mates with an α. In order for a yeast cell to “sense” another nearby yeast cell of the correct mating type, it relies on chemical pheromones that are released by the cells into the surroundings:
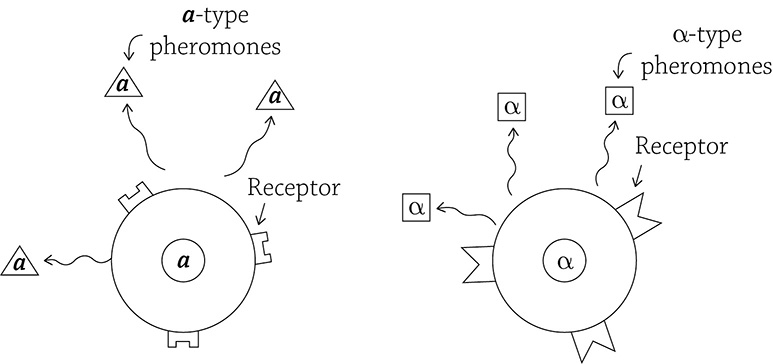
Yeast cells sending out mating signals
In order for an a cell to “hear” the signal from a nearby α cell, the α’s signal must bind to the a cell’s G protein-linked receptor (and vice versa).
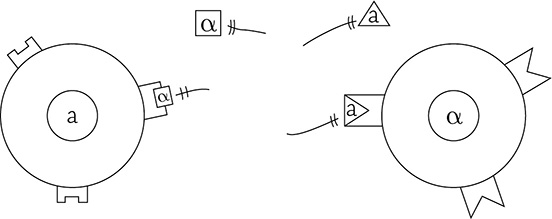
Yeast cells “hearing” the signal from the other mating type
The protein mating signal, once bound, activates a signal transduction pathway. This causes a signal cascade of a bunch of protein kinases activating other protein kinases, and so on and so forth. The kinases eventually descend upon the cell’s nucleus and spark the activity of a certain transcription factor (more on that in Part Three: Genetics) that initiates transcription of genes necessary for the cells to elongate and grow toward their mating partner. Specifically, the shape change occurs in the direction toward the highest concentration of pheromone, and the spherical little yeast cell forms a pear-like shape called … wait for it … a shmoo!
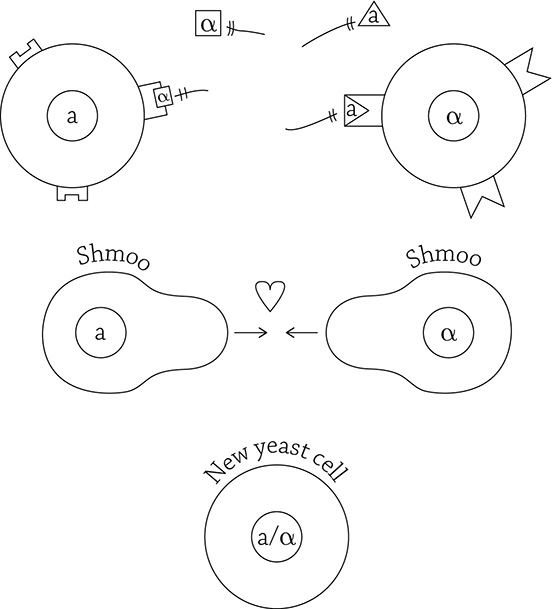
Yeast mating and the signal response pathway
Shmooing takes a lot of energy! The signal transduction pathway ensures that the cells only respond when a mating partner is close enough to make the attempt. Making a love connection is exhausting.
REVIEW QUESTIONS
1. Which category of chemical signal (lipid or protein) relies on the process of signal transduction to move the message into the cell?
2. Why does the receptor have to be a transmembrane protein, instead of a surface protein?
3. Put the following steps of G protein-coupled receptor function in order:
a. Cytoplasmic side of the receptor changes shape
b. G protein activates an enzyme on the inside of the cell
c. Signal binds the receptor
d. G protein is activated
4. What is the purpose of the transduction step of signal transduction?
5. List the three stages of signal transduction.
6. As soon as a signal binds the receptor, what occurs to move the message into the cell?
7. Put the following steps of ligand-gated ion channel function in order:
a. Second messengers rush into the cell
b. Receptor gate opens
c. Receptor is closed
d. Signal binds the receptor
8. Which receptor type is key for nervous system function?
9. Protein kinase enzymes play an important role in signal transduction because they ________________the message by creating a __________________ cascade.
