THE LIVING WORLD
Unit two. The Living Cell
4.8. The Cytoskeleton: Interior Framework of the Cell
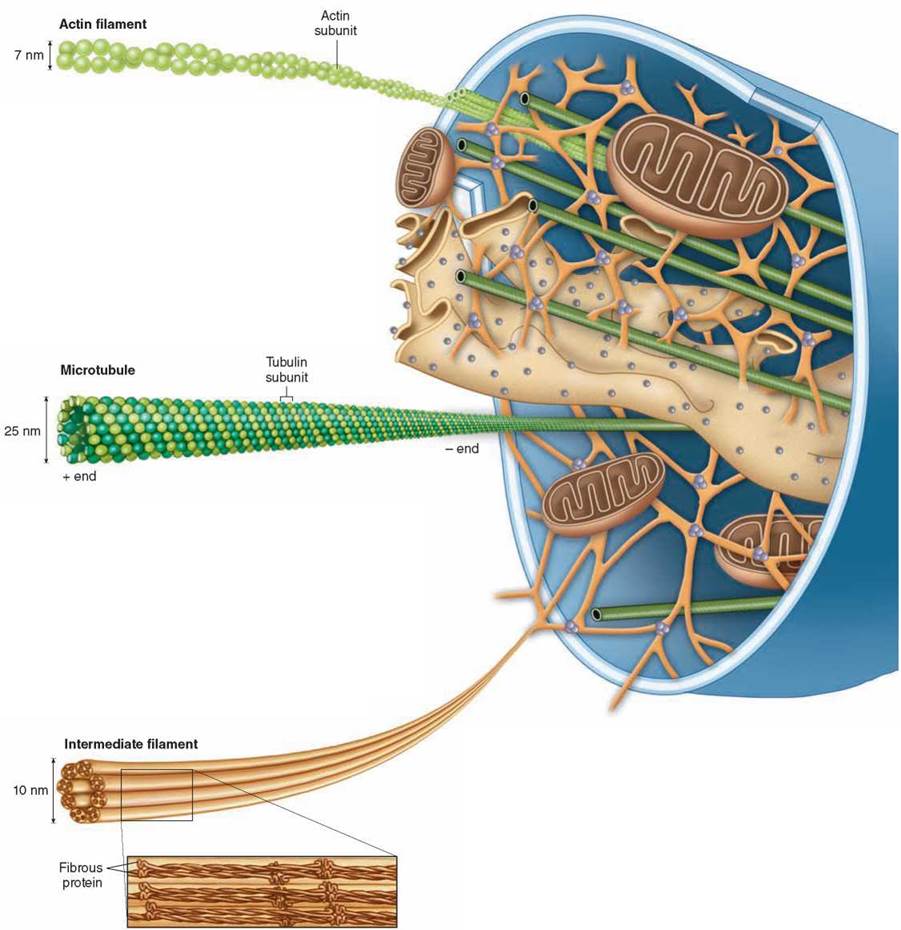
If you were to shrink down and enter into the interior of a eukaryotic cell, your view would be similar to what you see in the illustration shown here: A dense network of protein fibers called the cytoskeleton provides a framework that supports the shape of the cell. The cytoskeleton also anchors organelles like the mitochondria to fixed locations within the cell interior. The protein fibers of the cytoskeleton are a dynamic system, constantly being formed and disassembled. There are three different kinds of protein fibers that make up the cytoskeleton, shown as enlargements below and in table 4.2.
Microfilaments (Actin Filaments). Actin filaments are long fibers about 7 nanometers in diameter. Each filament is composed of two protein chains loosely twined together like two strands of pearls. Each “pearl,” or subunit, on the chains is the globular protein actin. Actin filaments are found throughout the cell but are most highly concentrated just inside the plasma membrane. Actin filaments are responsible for cellular movements such as contraction, crawling, “pinching” during division, and formation of cellular extensions.
Microtubules. Microtubules are hollow tubes about 25 nanometers in diameter composed of tubulin protein subunits arranged side by side to form a tube. In many cells, microtubules form from nucleation centers near the center of the cell and radiate toward the periphery. The ends of the microtubule are designated as “+” (away from the nucleation center) or “-” (toward the nucleation center). Microtubules are comparatively stiff cytoskeletal elements that serve to organize metabolism and intracellular transport in the nondividing cell and to stabilize cell structure. They are also responsible for the movement of chromosomes in mitosis.
Intermediate Filaments. Intermediate filaments are composed of overlapping staggered tetramers of protein. These tetramers are then bundled into cables. This molecular arrangement allows for a ropelike structure that imparts tremendous mechanical strength to the cell. Intermediate filaments are characteristically 8 to 10 nanometers in diameter, intermediate in size between actin filaments and microtubules (which is why they are called intermediate filaments). Once formed, intermediate filaments are stable and usually do not break down. They provide structural reinforcement to the cell and organelles.
The cytoskeleton plays a major role in determining the shape of animal cells, which lack rigid cell walls. Because filaments can form and dissolve readily, the shape of an animal cell can change rapidly. If you examine the surface of an animal cell with a microscope, you will often find it alive with motion, projections shooting out from the surface and then retracting, only to shoot out elsewhere moments later.
The cytoskeleton is not only responsible for the cell’s shape, but it also provides a scaffold both for ribosomes to carry out protein synthesis and for enzymes to be localized within defined areas of the cytoplasm. By anchoring particular enzymes near one another, the cytoskeleton participates with organelles in organizing the cell’s activities.
Centrioles
Complex structures called centrioles assemble microtubules from tubulin subunits in the cells of animals and most proists. Centrioles occur in pairs within the cytoplasm, usually located at right angles to one another as you can see in figure 4.12. They are usually found near the nuclear envelope and are among the most structurally complex microtubular assemblies of the cell. In cells that contain flagella or cilia, each flagellum or cilium is anchored by a form of centriole called a basal body. Most animal and protist cells have both centrioles and basal bodies; higher plants and fungi lack them, instead organizing microtubules without such structures. Although they lack a membrane, centrioles resemble spirochete bacteria in many other respects. Some biologists believe that centrioles, like mitochondria and chloroplasts, originated as symbiotic bacteria.

Figure 4.12. Centrioles.
Centrioles anchor and assemble microtubules. Centrioles usually occur in pairs and are composed of nine triplets of microtubules.
Vacuoles: Storage Compartments
Within the interiors of plant and many protist cells, the cytoskeleton positions not only organelles, but also storage compartments that are membrane-bounded, called vacuoles. The center of the plant cell shown above in figure 4.13, as in all plant cells, contains a large, apparently empty space, called the central vacuole. This vacuole is not really empty; it contains large amounts of water and other materials, such as sugars, ions, and pigments. The central vacuole functions as a storage center for these important substances.
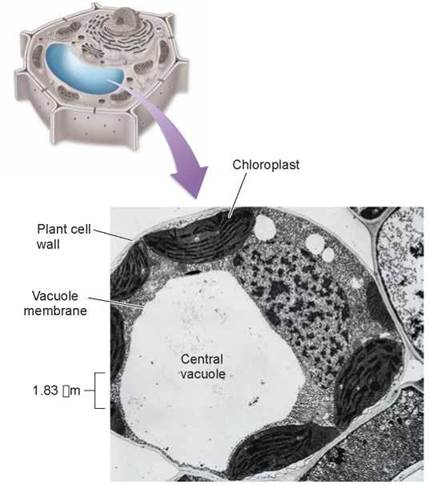
Figure 4.13. A plant central vacuole.
A plant's central vacuole stores dissolved substances and can increase in size to increase the surface area of a plant cell.
Certain protists, like Paramecium in figure 4.14, contain a contractile vacuole near the cell surface that accumulates excess water. This vacuole is bounded by actin filaments and has a small pore that opens to the outside of the cell. By rhythmic ATP-powered contractions, it pumps accumulated water out through the pore.
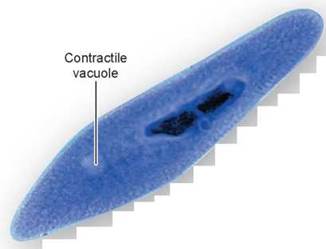
Figure 4.14. A contractile vacuole in Paramecium.
The contractile vacuole in Paramecium collects and eliminates excess water in the cell. Once filled, the contractile vacuole contracts and expels the water through a pore.
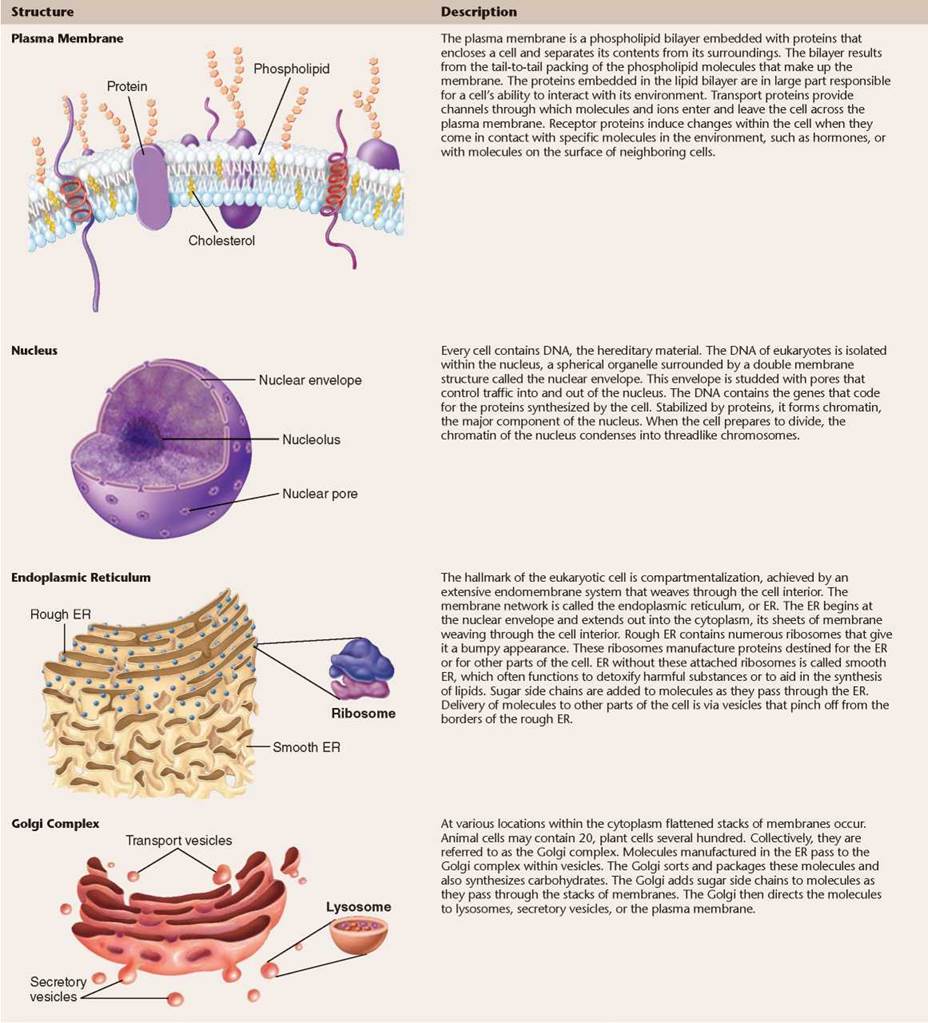
TABLE 4.2. EUKARYOTIC CELL STRUCTURES AND THEIR FUNCTIONS
Cell Movement
Essentially, all cell motion is tied to the movement of actin filaments, microtubules, or both. Intermediate filaments act as intracellular tendons, preventing excessive stretching of cells, and actin filaments play a major role in determining the shape of cells. Because actin filaments can form and dissolve so readily, they enable some cells to change shape quickly. If you look at the surfaces of such cells under a microscope, you will find them moving and changing shape.
Some Cells Crawl. It is the arrangement of actin filaments within the cell cytoplasm that allows cells to “crawl,” literally! Crawling is a significant cellular phenomenon, essential to inflammation, clotting, wound healing, and the spread of cancer. White blood cells in particular exhibit this ability. Produced in the bone marrow, these cells are released into the circulatory system and then eventually crawl out of capillaries and into the tissues to destroy potential pathogens. The crawling mechanism is an exquisite example of cellular coordination.
Cytoskeletal fibers play a role in other types of cell movement. For example, during animal cell reproduction (see chapters 8 and 9), chromosomes move to opposite sides of a dividing cell because they are attached to shortening microtubules. The cell then pinches in two when a belt of actin filaments contracts like a purse string.
Muscle cells also use actin filaments to contract their cytoskeletons. The fluttering of an eyelash, the flight of an eagle, and the awkward crawling of a baby all depend on these cyto- skeletal movements within muscle cells.
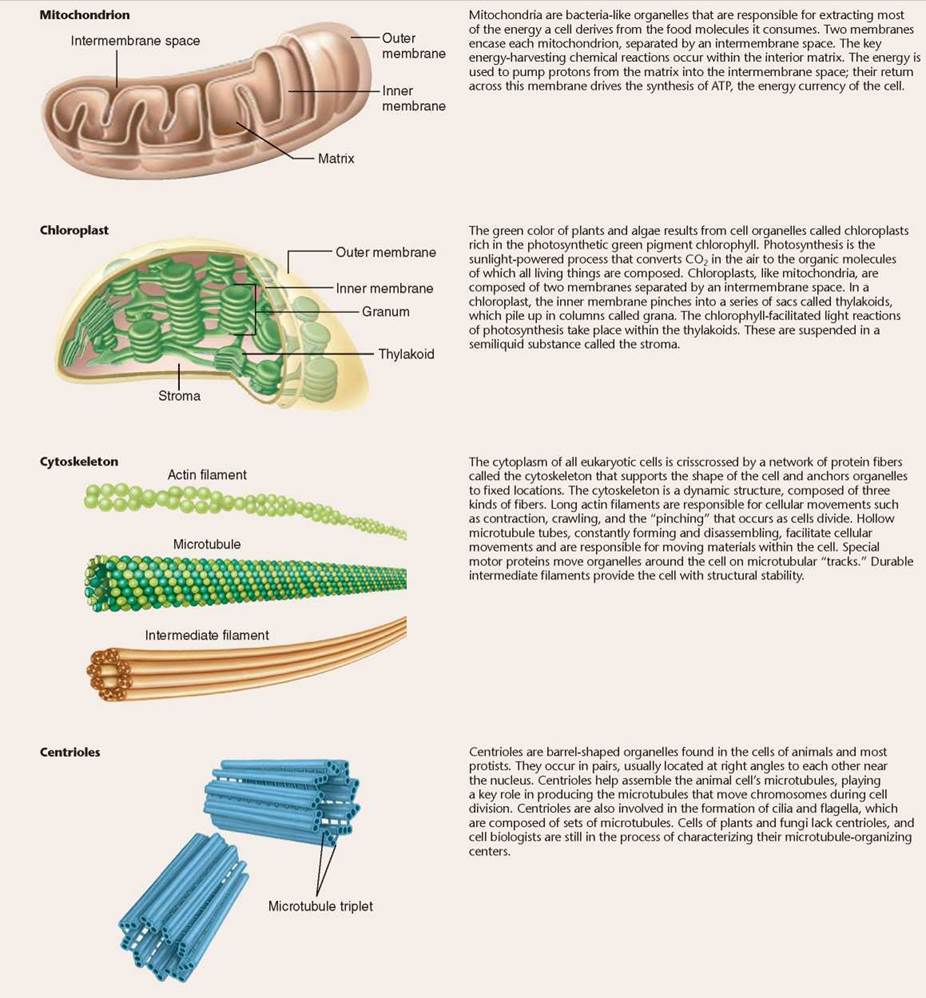
Swimming with Flagella and Cilia. Some eukaryotic cells contain flagella (singular, flagellum), fine, long, threadlike organelles protruding from the cell surface. The cutaway view in figure 4.15 shows how a flagellum arises from a microtubular structure called a basal body, with groups of microtubules arranged in rows of three, as you can see in the cross-sectional view. Some of these microtubules extend up into the flagellum, which consists of a circle of nine microtubule pairs surrounding two central ones (again seen in the cross-sectional view). This 9 + 2 arrangement is a fundamental feature of eukaryotes and apparently evolved early in their history. Even in cells that lack flagella, derived structures with the same 9 + 2 arrangement often occur, like in the sensory hairs of the human ear. In humans, we find a single long flagellum on each sperm cell that propels the cell in a swimming motion. If flagella are numerous and organized in dense rows, they are called cilia. Cilia do not differ from flagella in their structure, but cilia are usually short. The Paramecium in figure 4.16 is covered with cilia, giving it a furry appearance. In humans, dense mats of cilia project from cells that line our breathing tube, the trachea, to move mucus and dust particles out of the respiratory tract into the throat (where we can expel these unneeded contaminants by spitting or swallowing). Eukaryotic flagella serve a similar function as flagella found in prokaryotes, discussed in section 4.3, but they are very different structurally.
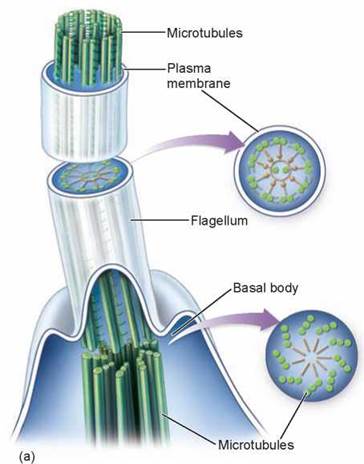
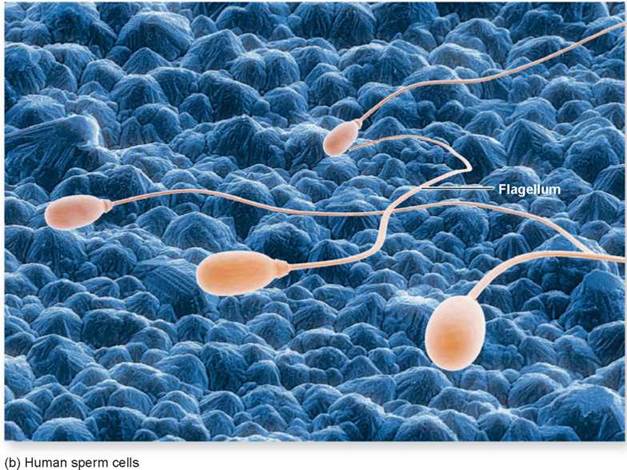
Figure 4.15. Flagella.
(a) A eukaryotic flagellum springs directly from a basal body and is composed of a ring of nine pairs of microtubules with two microtubules in its core. (b) Human sperm cells have one flagellum.
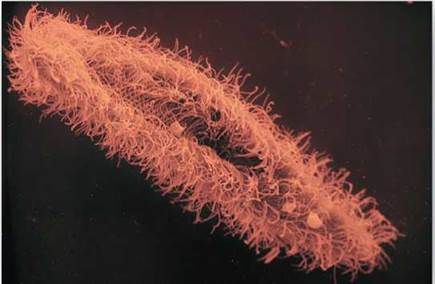
Figure 4.16. Cilia.
The surface of this Paramecium is covered with a dense forest of cilia.
Moving Materials Within the Cell
All eukaryotic cells must move materials from one place to another in the cytoplasm. Most cells use the endomembrane system as an intracellular highway. As you saw on page 82, the Golgi complex packages materials into vesicles that come from the channels of the endoplasmic reticulum to the far reaches of the cell. However, this highway is only effective over short distances. When a cell has to transport materials through long extensions like the axon of a nerve cell, the endomembrane system highways are too slow. For these situations, eukaryotic cells have developed high-speed locomotives that run along microtubular tracks. Lysosomes move along such microtubular tracks to reach a food vacuole, and mitochondria travel down them to the far-away tips of long axons.
For long-distance intracellular transport, four components are required, illustrated in figure 4.17: a vesicle or organelle (the light tan structure) that is to be transported; a motor molecule, in this case dynein, that provides the energy-driven motion; connector molecules that connect the vesicle to the motor molecule (in the figure, they are the dynactin complex and other associated proteins); and microtubules (the green tube) on which the vesicle will ride like a train on a rail, pulled by a locomotive. As nature’s tiniest motors, these motor proteins literally pull the transport vesicles along the microtubular tracks.
How does such a tiny motor work? A motor protein uses ATP to power its movement. Scientists have proposed that they use a type of stepping action. Also, the direction of movement is different for different motor proteins. The dynein motor protein shown in figure 4.17 moves inward toward the cell’s center, dragging the vesicle with it as it travels along toward the “-” end of a microtubule. Another motor protein, kinesin, directs movement in the opposite direction, toward the “+” end of a microtubule, which is toward the periphery of the cell. The destination of a particular transport vesicle and its contents is thus determined by the nature of the linking protein embedded within the vesicle’s membrane. Like possessing a ticket to one of two destinations, if a vesicle links to kinesin, it moves outward; if it links to dynein, it moves inward.
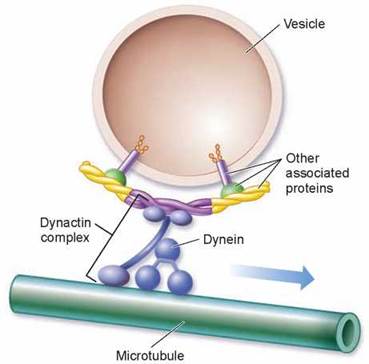
Figure 4.17. Molecular motors.
Vesicles that are transported within cells are attached with connector molecules, such as the dynactin complex shown here, to motor molecules, like dynein, which move along microtubules.
Key Learning Outcome 4.8. The cytoskeleton is a latticework of protein fibers that determines a cell's shape and anchors organelles to particular locations within the cytoplasm. Cells can move by changing their shape and move substances around the cell using molecular motors.