THE LIVING WORLD
Unit Eight. The Living Environment
37. Behavior and the Environment
37.12. Altruism and Group Living
Altruism—the performance of an action that benefits another individual at a cost to the actor—occurs in many guises in the animal world. In many bird species, for example, parents are assisted in raising their young by other birds, which are called helpers at the nest. In species of both mammals and birds, individuals that spy a predator will give an alarm call, alerting other members of their group, even though such an act would seem to call the predator’s attention to the caller. Finally, lionesses with cubs will allow all cubs in the pride to nurse, including cubs of other females.
The existence of altruism has long perplexed evolutionary biologists. If altruism imposes a cost to an individual, how could an allele for altruism be favored by natural selection? One would expect such alleles to be at a disadvantage, and thus their frequency in the gene pool should decrease through time.
A number of explanations have been put forward to explain the evolution of altruism. One suggestion often heard on television documentaries is that such traits evolve for the good of the species. The problem with such explanations is that natural selection operates on individuals within species, not on species themselves. Thus, natural selection will not favor alleles that lead an individual to act in ways that benefit others at a cost to itself; it is even possible for traits to evolve that are detrimental to the species as a whole, as long as they benefit the individual.
In some cases, selection can operate on groups of individuals, but this is rare. For example, if an allele for supercannibalism evolved within a population, individuals with that allele would be favored, as they would have more to eat; however, the group might eventually eat itself to extinction, and the allele would be removed from the species. In certain circumstances, such group selection can occur, but the conditions for it to occur are rarely met in nature. In most cases, consequently, the “good of the species” cannot explain the evolution of altruistic traits.
Another possibility is that seemingly altruistic acts aren’t altruistic after all. For example, helpers at the nest are often young and gain valuable parenting experience by assisting established breeders. Moreover, by hanging around an area, such individuals may inherit the territory when the established breeders die. Similarly, alarm callers (figure 37.16) may actually benefit by causing other animals to panic. In the ensuing confusion, the caller may be able to slip off undetected. Detailed field studies in recent years have demonstrated that some acts truly are altruistic, but others are not as they seemed.

Figure 37.16. An altruistic act—or is it?
A meerkat sentinel on duty. Meerkats, Suricata suricata, are a species of highly social mongoose living in the semiarid sands of the Kalahari Desert in southern Africa. This meerkat is taking its turn to act as a lookout for predators. Under the security of its vigilance, the other members of the group can focus their attention on foraging. The sentinel puts his own life at risk when he gives an alarm, an apparent example of altruistic behavior.
Reciprocity
Robert Trivers, now of Rutgers University, proposed that individuals may form “partnerships” in which mutual exchanges of altruistic acts occur because it benefits both participants to do so. In the evolution of such reciprocal altruism, “cheaters” (nonreciprocators) are discriminated against and are cut off from receiving future aid. According to Trivers, if the altruistic act is relatively inexpensive, the small benefit a cheater receives by not reciprocating is far outweighed by the potential cost of not receiving future aid. Under these conditions, cheating should not occur.
For example, vampire bats roost in hollow trees in groups of 8 to 12 individuals. Because these bats have a high metabolic rate, individuals that have not fed recently may die. Bats that have found a host imbibe a great deal of blood; giving up a small amount presents no great energy cost to the donor, and it can keep a roostmate from starvation. Vampire bats tend to share blood with past reciprocators. If an individual fails to give blood to a bat from which it had received blood in the past, it will be excluded from future bloodsharing.
Kin Selection
The most influential explanation for the origin of altruism was presented by William D. Hamilton in 1964. It is perhaps best introduced by quoting a passing remark made in a pub in 1932 by the great population geneticist J. B. S. Haldane. Haldane said that he would willingly lay down his life for two brothers or eight first cousins. Evolutionary speaking, Haldane’s statement makes sense, because for each allele Haldane received from his parents, his brothers each had a 50% chance of receiving the same allele. Consequently, it is statistically expected that two of his brothers would pass on as many of Haldane’s particular combination of alleles to the next generation as Haldane himself would. Similarly, Haldane and a first cousin would share an eighth of their alleles. Their parents, which are siblings, would each share half their alleles, and each of their children would receive half of these, of which half on the average would be in common: 1/2 x 1/2 x 1/2 =1/8. Eight first cousins would therefore pass on as many of those alleles to the next generation as Haldane himself would. Hamilton saw Haldane’s point clearly: Natural selection will favor any strategy that increases the net flow of an individual’s alleles to the next generation.
Hamilton showed that by directing aid toward kin, or close genetic relatives, an altruist may increase the reproductive success of its relatives enough to compensate for the reduction in its own fitness. Because the altruist’s behavior increases the propagation of its own alleles in relatives, it will be favored by natural selection. Selection that favors altruism directed toward relatives is called kin selection. Although the behaviors being favored are cooperative, the genes are actually “behaving selfishly,” because they encourage the organisms to support copies of themselves in other individuals. In other words, if an individual has a dominant allele that causes altruism, any action that increases the fitness of relatives and thus increases the frequency of this allele in future generations will be favored, even if that action is detrimental to the particular individual taking the action.
Examples of Kin Selection
Hamilton’s kin selection model predicts that altruism is likely to be directed toward close relatives. The more closely related two individuals are, the greater the potential genetic payoff.
Many examples of kin selection are known from the animal world. Belding’s ground squirrels give alarm calls when they spot a predator such as a coyote or a badger. Such predators may attack a calling squirrel, so giving a signal places the caller at risk. The social unit of a ground squirrel colony consists of a female and her daughters, sisters, aunts, and nieces. When they mature, males disperse long distances from where they are born; thus, adult males in the colony are not genetically related to the females. By marking all squirrels in a colony with an individual dye pattern on their fur and by recording which individuals gave calls and the social circumstances of their calling, researchers found that females who have relatives living nearby are more likely to give alarm calls than females with no kin nearby. Males tend to call much less frequently, as would be expected because they are not related to most colony members.
Another example of kin selection comes from a bird called the white-fronted bee-eater, which lives along rivers in Africa in colonies of 100 to 200 birds. In contrast to the ground squirrels, it is the males that usually remain in the colony in which they were born, and the females that disperse to join new colonies. Many bee-eaters do not raise their own offspring, but rather help others. Many of these birds are relatively young, but helpers also include older birds whose nesting attempts have failed. The presence of a single helper, on average, doubles the number of offspring that survive. Two lines of evidence support the idea that kin selection is important in determining helping behavior in this species. First, helpers are normally males, which are usually related to other birds in the colony, and not females, which are not related. Second, when birds have the choice of helping different parents, they almost invariably choose the parents to which they are most closely related. The graph in figure 37.17 compares the probability of a bird helping in a nest (on the y axis) and the relationship of the helper (on the x axis). The more closely related the helper (toward the right side of the graph), the higher the probability that it will be a helper in the nest.
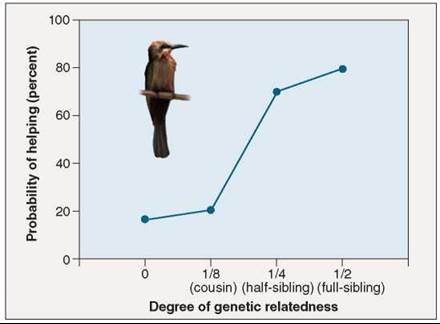
Figure 37.17. Kin selection is common among vertebrates.
In the white-fronted bee-eaters (Merops bullockoides), nonbreeding individuals will help raise the offspring of others. Most helpers are close relatives, and the probability that a bird will help another increases with genetic relatedness.
Key Learning Outcome 37.12. Many factors could be responsible for the evolution of altruistic behaviors. Individuals may benefit directly if altruistic acts are reciprocated; also kin selection explains how alleles for altruism can increase in frequency if altruistic acts are directed toward relatives.