THE LIVING WORLD
Unit two. The Living Cell
7. How Cells Harvest Energy from Food
7.3. Harvesting Electrons from Chemical Bonds
The first step of oxidative respiration in the mitochondrion is the oxidation of the three-carbon molecule called pyruvate, which is the end product of glycolysis. The cell harvests pyruvate’s considerable energy in two steps: first, by oxidizing pyruvate to form acetyl-CoA, and then by oxidizing acetyl-CoA in the Krebs cycle.
Step One: Producing Acetvl-CoA
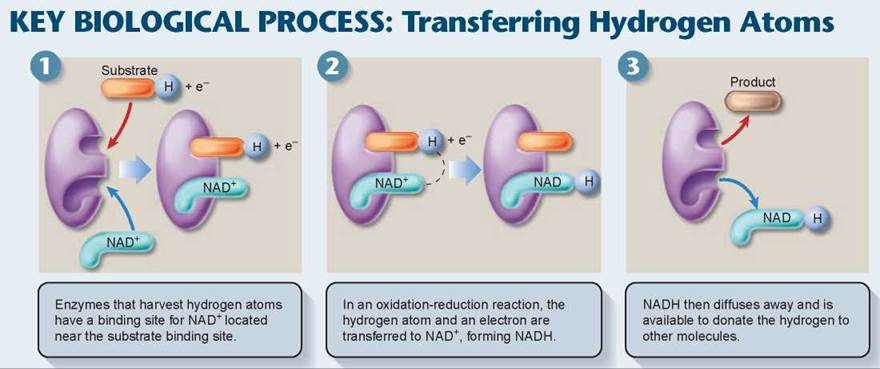
Pyruvate is oxidized in a single reaction that cleaves off one of pyruvate’s three carbons. This carbon then departs as part of a CO2 molecule, shown in figure 7.5 coming off the pathway with the green arrow. Pyruvate dehydrogenase, the complex of enzymes that removes CO2 from pyruvate, is one of the largest enzymes known. It contains 60 subunits! In the course of the reaction, a hydrogen and electrons are removed from pyruvate and donated to NAD+ to form NADH. The Key Biological Process illustration below shows how an enzyme catalyzes this reaction, bringing the substrate (pyruvate) into proximity with NAD+. Cells use NAD+ to carry hydrogen atoms and energetic electrons from one molecule to another. NAD+ oxidizes energy-rich molecules by acquiring their hydrogens (this proceeds 1 → 2 → 3 in the figure) and then reduces other molecules by giving the hydrogens to them (this proceeds 3 → 2 → 1). Now focus again on figure 7.5. The two-carbon fragment (called an acetyl group) that remains after removing CO2 from pyruvate is joined to a cofactor called coenzyme A (CoA) by pyruvate dehydrogenase, forming a compound known as acetyl-CoA. If the cell has plentiful supplies of ATP, acetyl-CoA is funneled into fat synthesis, with its energetic electrons preserved for later needs. If the cell needs ATP now, the fragment is directed instead into ATP production through the Krebs cycle.
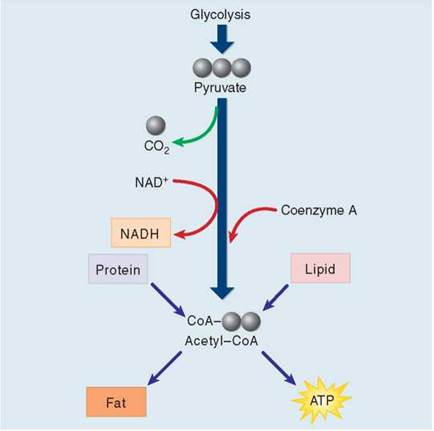
Figure 7.5. Producing acetyl-CoA.
Pyruvate, the three-carbon product of glycolysis, is oxidized to the two-carbon molecule acetyl-CoA, in the process losing one carbon atom as CO2 and an electron (donated to NAD+ to form NADH). Almost all the molecules you use as foodstuffs are converted to acetyl-CoA; the acetyl-CoA is then channeled into fat synthesis or into ATP production, depending on your body's needs.
A Closer Look
Metabolic Efficiency and the Length of Food Chains
In the earth's ecosystems, the organisms that carry out photosynthesis are often consumed as food by other organisms. We call these "organism-eaters” heterotrophs. Humans are heterotrophs, as no human photosynthesizes.
It is thought that the first heterotrophs were ancient bacteria living in a world where photosynthesis had not yet introduced much oxygen into the oceans or atmosphere. The only mechanism they possessed to harvest chemical energy from their food was glycolysis. Neither oxygen-generating photosynthesis nor the oxidative stage of cellular respiration had evolved yet. It has been estimated that a heterotroph limited to glycolysis, as these ancient bacteria were, captures only 3.5% of the energy in the food it consumes.
Hence, if such a heterotroph preserves 3.5% of the energy in the photosynthesizers it consumes, then any other heterotrophs that consume the first heterotroph will capture through glycolysis 3.5% of the energy in it, or 0.12% of the energy available in the original photosynthetic organisms. A very large base of photosynthesizers would thus be needed to support a small number of heterotrophs.
When organisms became able to extract energy from organic molecules by oxidative cellular respiration, which we discuss on the next page, this constraint became far less severe, because the efficiency of oxidative respiration is estimated to be about 32%. This increased efficiency results in the transmission of much more energy from one trophic level to another than does glycolysis. (A trophic level is a step in the movement of energy through an ecosystem.) The efficiency of oxidative cellular respiration has made possible the evolution of food chains, in which photosynthesizers are consumed by heterotrophs, which are consumed by other heterotrophs, and so on. You will read more about food chains in chapter 36.
Even with this very efficient oxidative metabolism, approximately two-thirds of the available energy is lost at each trophic level, and that puts a limit on how long a food chain can be. Most food chains, like the East African grassland ecosystem illustrated here, involve only three or rarely four trophic levels. Too much energy is lost at each transfer to allow chains to be much longer than that.
For example, it would be impossible for a large human population to subsist by eating lions captured from the grasslands of East Africa; the amount of grass available there would not support enough zebras and other herbivores to maintain the number of lions needed to feed the human population. Thus, the ecological complexity of our world is fixed in a fundamental way by the chemistry of oxidative cellular respiration.
Photosynthesizers. The grass under this yellow fever tree grows actively during the hot, rainy season, capturing the energy of the sun and storing it in molecules of glucose, which are then converted into starch and stored in the grass.

Herbivores. These zebras consume the grass and transfer some of its stored energy into their own bodies.

Carnivores. The lion feeds on zebras and other animals, capturing part of their stored energy and storing it in its own body.

Scavengers. This hyena and the vultures occupy the same stage in the food chain as the lion. They also consume the body of the dead zebra, after it has been abandoned by the lion.

Refuse utilizers. These butterflies, mostly Precis octavia, are feeding on the material left in the hyena's dung after the food the hyena consumed had passed through its digestive tract.

A food chain in the savannas, or open grasslands, of East Africa.
At each of these levels in the food chain, only about a third or less of the energy present is used by the recipient.
Step Two: The Krebs Cycle
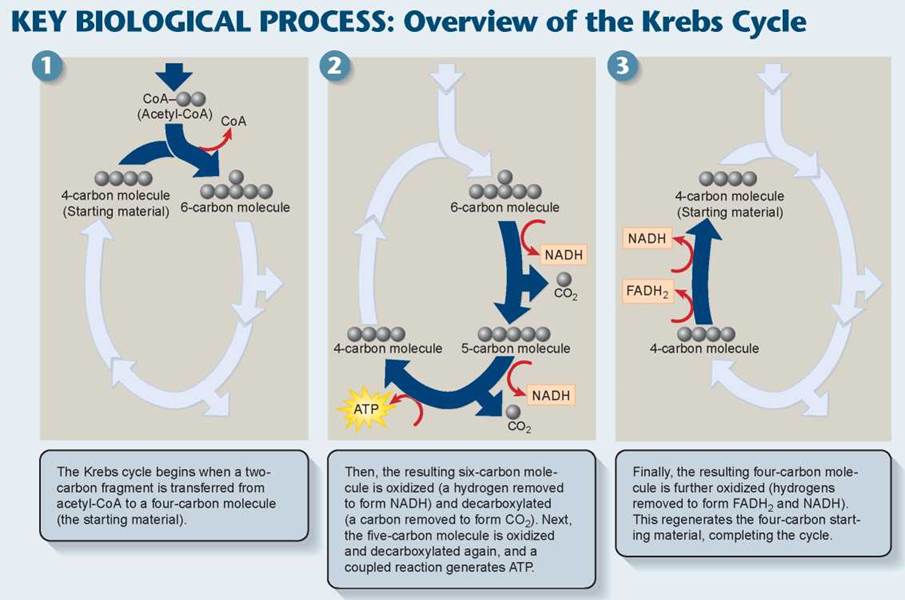
The next stage in oxidative respiration is called the Krebs cycle, named after the man who discovered it. The Krebs cycle (not to be confused with the Calvin cycle in photosynthesis) takes place within the mitochondrion. While a complex process, its nine reactions can be broken down into three stages, as indicated by the overview presented in the Key Biological Process illustration below:
Stage 1. Acetyl-CoA joins the cycle, binding to a four-carbon molecule and producing a six-carbon molecule.
Stage 2. Two carbons are removed as CO2, their electrons donated to NAD+, and a four-carbon molecule is left. A molecule of ATP is also produced.
Stage 3. More electrons are extracted, forming NADH and FADH2; the four-carbon starting material is regenerated.
To examine the Krebs cycle in more detail, follow along the series of individual reactions illustrated in figure 7.6. The cycle starts when the two-carbon acetyl-CoA fragment produced from pyruvate is stuck onto a four-carbon sugar called oxaloacetate. Then, in rapid-fire order, a series of eight additional reactions occur (steps 2 through 9). When it is all over, two carbon atoms have been expelled as CO2, one ATP molecule has been made in a coupled reaction, eight more energetic electrons have been harvested and taken away as NADH or on other carriers, such as FADH2, which serves the same function as NADH, and we are left with the same four-carbon sugar we started with. The process of reactions is a cycle—that is, a circle of reactions. In each turn of the cycle, a new acetyl group replaces the two CO2 molecules lost, and more electrons are extracted. Note that a single glucose molecule produces two turns of the cycle, one for each of the two pyruvate molecules generated by glycolysis.
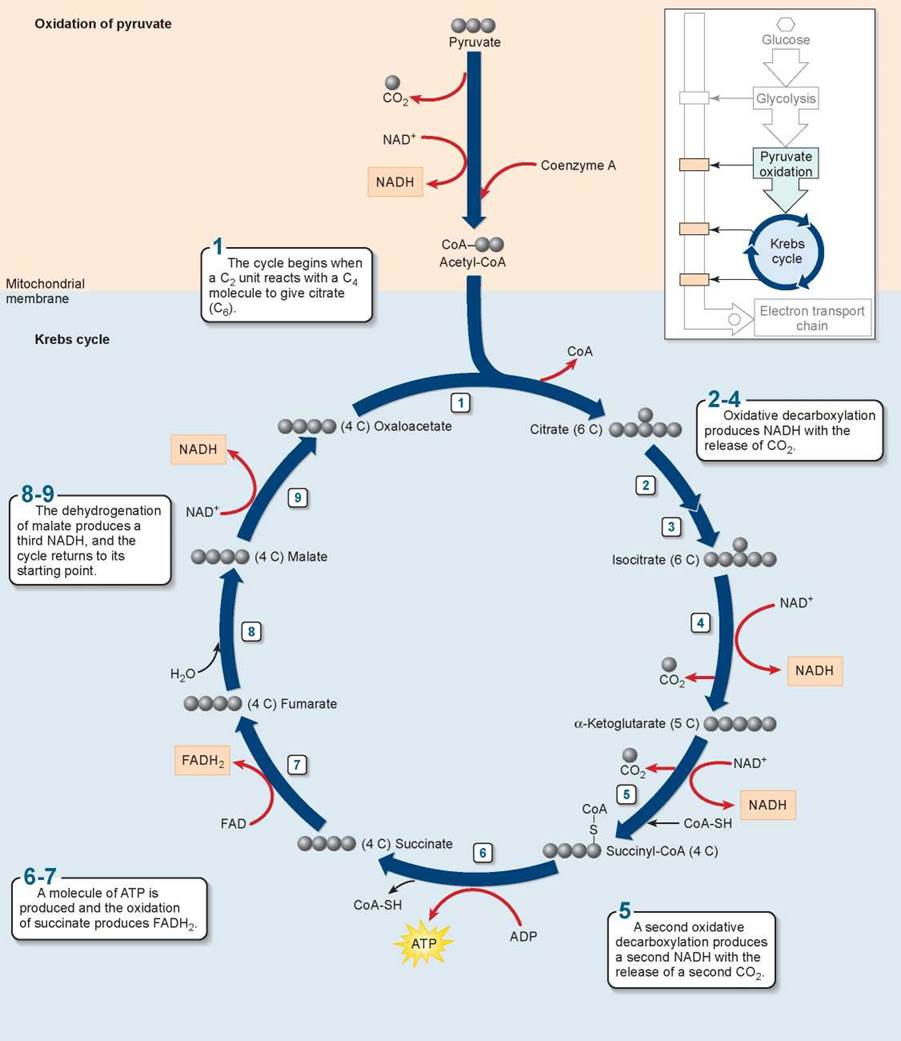
Figure 7.6. The Krebs cycle.
This series of nine enzyme-catalyzed reactions takes place within the mitochondrion.
In the process of cellular respiration, glucose is entirely consumed. The six-carbon glucose molecule is first cleaved into a pair of three-carbon pyruvate molecules during glycolysis. One of the carbons of each pyruvate is then lost as CO2 in the conversion of pyruvate to acetyl-CoA, and the other two carbons are lost as CO2 during the oxidations of the Krebs cycle. All that is left to mark the passing of the glucose molecule into six CO2 molecules is its energy, preserved in four ATP molecules and electrons carried by 10 NADH and two FADH2 carriers.
Key Learning Outcome 7.3. The end product of glycolysis, pyruvate, is oxidized to the two-carbon acetyl-CoA, yielding a pair of electrons plus CO2. Acetyl-CoA then enters the Krebs cycle, yielding ATP, many energized electrons, and two CO2 molecules.