MCAT Biology Review
Chapter 3: Embryogenesis and Development
3.2 Mechanisms of Development
As described earlier, cells undergo determinate cleavage to commit to a particular cell line, and inducers can be used to communicate from one group of differentiating cells to another. In this section, we take a more specific look at the biochemical properties of these developmental mechanisms, as well as a few selected others.
CELL SPECIALIZATION
An adult human being is composed of approximately 37 trillion cells. These cells are organized into tissues, which form organs within organ systems. In order to create an organism as complex as a human being, each cell must perform a specialized function. In addition, the cells in an organ must be organized such that the organ can function properly. For example, the pancreas must create both digestive enzymes (trypsin, carboxypeptidases A and B, pancreatic lipase, and others) and endocrine hormones (insulin, glucagon, and somatostatin). The cells that synthesize digestive enzymes must be located such that the cell products can enter ducts to ultimately empty into the duodenum. Likewise, the cells that synthesize endocrine hormones must be located near a blood vessel to put their products into systemic circulation. In order to accomplish this, the cell must be determined and then differentiated.
Determination
As described earlier, determination is the commitment of a cell to having a particular function in the future. Prior to determination, the cell can become any cell type. After determination, the cell is committed to a specific lineage. There are multiple pathways by which determination may occur. If during cleavage, the existing mRNA and protein in the parent cell has been asymmetrically distributed between the daughter cells, the presence of specific mRNA and protein molecules may result in determination. Determination may also occur due to secretion of specific molecules from nearby cells. These molecules, also called morphogens, may cause nearby cells to follow a particular developmental pathway. Determination is a commitment to a particular cell type, but note that the cell has not yet actually produced the products it needs to carry out the functions of that cell type—that is the goal of differentiation.
Differentiation
After a cell’s fate has been determined, the cell must begin to undertake changes that cause the cell to develop into the determined cell type. This includes changing the structure, function, and biochemistry of the cell to match the cell type through differentiation.
KEY CONCEPT
When a cell is determined, it is committed to a particular cell lineage. When the cell differentiates, it assumes the structure, function, and biochemistry of that cell type.
Cells that have not yet differentiated, or which give rise to other cells that will differentiate, are known as stem cells. Stem cells exist in embryonic tissues as well as adult tissues. The tissues a particular stem cell can differentiate into are determined by its potency. Cells with the greatest potency are called totipotent and include embryonic stem cells; totipotent cells can ultimately differentiate into any cell type, either in the fetus or in the placental structures. After the 16-cell stage, the cells of the morula begin to differentiate into two groups: the inner cell mass and the trophoblast cells. After a few more cycles of cell division, these totipotent cells start to differentiate into the three germ cell layers. At this stage, the cells are said to be pluripotent; these cells can differentiate into any cell type except for those found in the placental structures. Finally, as the cells continue to become more specialized, they are said to be multipotent. Multipotent stem cells can differentiate into multiple types of cells within a particular group. For example, hematopoietic stem cells are cells that are capable of differentiating into all of the cells found in blood, including the various types of white blood cells, red blood cells, and platelets—but not into skin cells, neurons, or muscle cells. While we use all of these different terms to describe potency, it is important to recognize that potency is a spectrum, not a series of strict definitions. Also, note that stem cells exist not only in embryos, but also in adults—we have stem cells that give rise to skin, blood, and the epithelial lining of the digestive tract, among others.
KEY CONCEPT
Stem cells are able to differentiate into different cell types. The potency of the stem cell determines how many different cell types a stem cell can become. As cells become more differentiated, the potency of the cell gets more narrow (from totipotent to pluripotent to multipotent).
Over the last few decades, stem cell research has been a hotly contested issue. While harvesting of embryonic stem cells, highlighted in Figure 3.8, ultimately results in destruction of the embryo, it is thought that these cells could be used to regenerate human tissues, including the spinal cord (following injury) and cardiac myocytes (following a heart attack). There are also immunologic concerns, as transplantation of stem cells of a different genetic makeup would evoke an immune response, resulting in rejection. In addition, pluripotent cells may not always differentiate into the desired tissue, once implanted, and may even become cancerous.
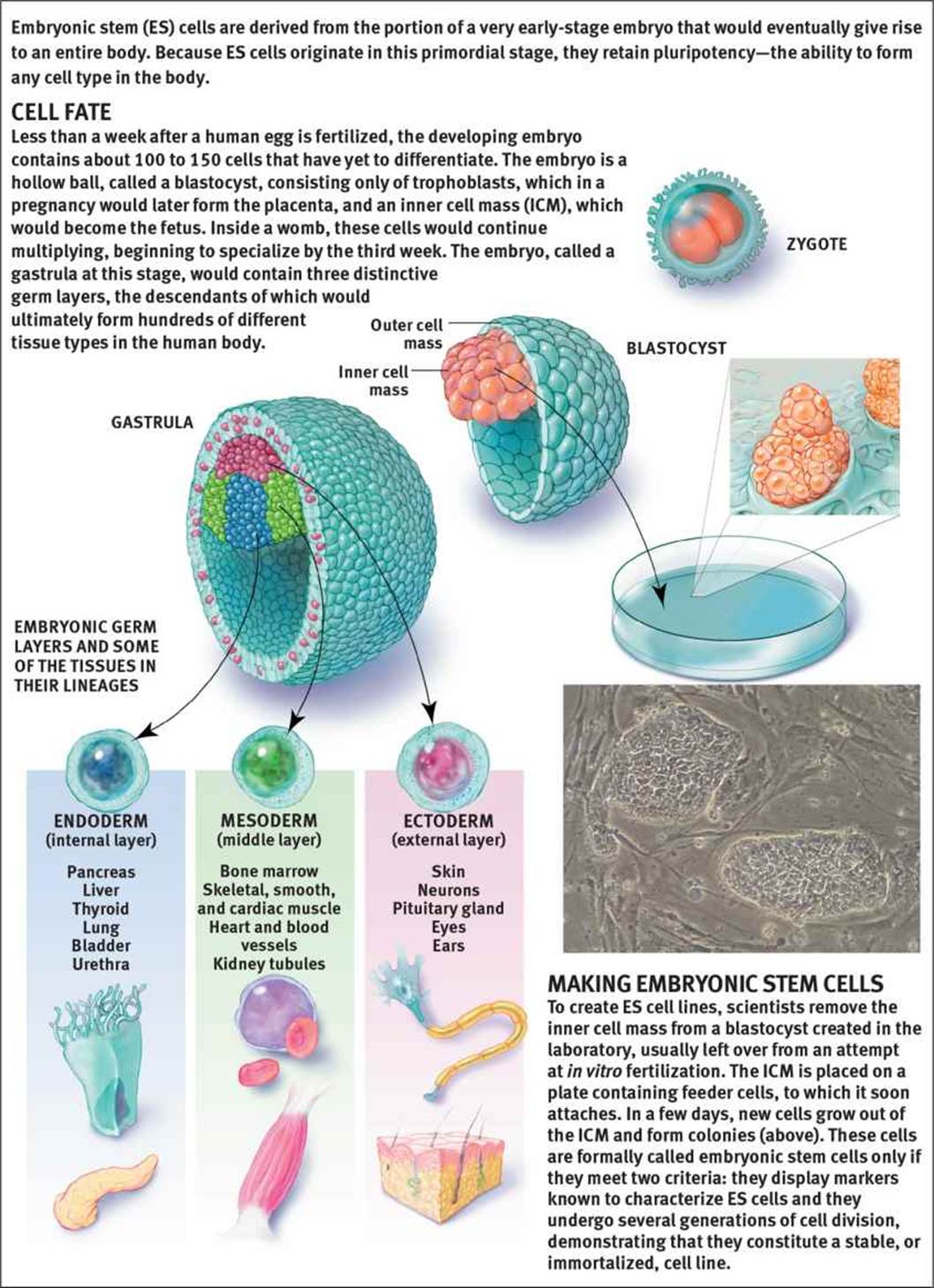 Figure 3.8. Embryonic Stem Cells
Figure 3.8. Embryonic Stem Cells
In order to address this controversy, many researchers have begun investigating adult stem cells. At best, these cells are multipotent, able to differentiate into only a few different cell types. Researchers may take adult stem cells and use various transcription factors to increase potency in these cells. One of the potential advantages of this is that a stem cell can be taken from the skin of a patient, induced to become a different tissue type, and then implanted into that same patient. This offers reduced risk of rejection of foreign tissue. However, it is challenging to induce differentiation into the correct cell type, and most organs have a complex structure that depends on a number of different cell types, each of which requires different signals. Research is ongoing and holds promise, despite limited success so far.
BRIDGE
Stem cells are a cornerstone of biotechnology. Gene studies can be performed by introducing altered embryonic stem cells that contain transgenes into mice. Stem cells lacking a particular gene can be used to create knockout mice. These processes are discussed in Chapter 6 ofMCAT Biochemistry Review.
CELL–CELL COMMUNICATION
The determination and differentiation of a cell depends on the location of the cell itself as well as the identity of the surrounding cells. The developing cell receives signals from organizing cells around it and may also secrete its own signals. As discussed previously, surrounding tissues induce a developing cell to become a particular cell type via inducers; the term inducer may also refer to the cell secreting the signal. The cell that is induced is called a responder (responsive cell); to be induced, a responder must be competent, or able to respond to the inducing signal.
Cell–cell communication can occur via autocrine, paracrine, juxtacrine, or endocrine signals. Autocrine signals act on the same cell that secreted the signal in the first place. Paracrine signals act on cells in the local area. Juxtacrine signals do not usually involve diffusion, but rather feature a cell directly stimulating receptors of the adjacent cell. Finally, endocrine signals involve secreted hormones that travel through the bloodstream to a distant target tissue.
Inducers
Inducers are commonly growth factors, which are peptides that promote differentiation and mitosis in certain tissues. Most growth factors only function on specific cell types or in certain areas, as determined by the competence of these cells. In this way, certain growth factors can code for particular tissues. For example, Pax6 is expressed in the ectoderm of the head, but in no other location. Therefore, as the optic vesicle approaches the overlying ectoderm while producing this factor, development of the lens of the eye is induced in the overlying ectoderm. Interestingly, induction is not always a one-way pathway. To that end, differentiation of the lens then triggers the optic vesicle to form the optic cup, which ultimately becomes the retina. This is known as reciprocal development. Most tissues will be exposed to multiple inducers during the course of development.
REAL WORLD
In development of the eyes, lateral outpocketings from the brain (optic vesicles) grow out and touch the overlying ectoderm. The optic vesicle induces the ectoderm to form the lens placode. The lens placode in turn induces the optic vesicle to create the optic cup. The optic cup then induces the lens placode to develop into the cornea and lens. Experiments with frog embryos show that if this ectoderm is subsequently transplanted to the trunk (after the optic vesicles have grown out), a lens will develop in the trunk. If, however, the ectoderm is transplanted before the outgrowth of the optic vesicles, it will not.
One of the main methods of signaling occurs via the use of gradients. Morphogens, or molecules that cause determination of cells, diffuse through the organism. Locations closer to the origin of the morphogen will be exposed to higher concentrations, while areas further away will have less exposure. Multiple morphogens are secreted simultaneously, resulting in unique combinations of morphogen exposure throughout the organism, which can thereby induce the differentiation of specific cell types. Some common morphogens include transforming growth factor beta (TGF-β), sonic hedgehog (Shh), and epidermal growth factor (EGF).
CELL MIGRATION, CELL DEATH, AND REGENERATION
Induction and differentiation allow for the creation of different types of cells; however, these cells are not always in the right location to carry out their function. Further, the sculpting of various anatomic structures requires not only differentiation, but death of some cells. Certain organs also have the ability to recreate injured or surgically removed portions of tissue.
Cell Migration
Cells must be able to disconnect from adjacent structures and migrate to their anatomically correct location. For example, the anterior pituitary gland originates from a segment of oral ectoderm and must migrate from the top of the mouth to its final location just below the hypothalamus. Neural crest cells also undergo extensive migration. These cells form at the edge of the neural folds during neurulation and then migrate throughout the body to form many different structures, including the sensory ganglia, autonomic ganglia, adrenal medulla, and Schwann cells, as well as specific cell types in other tissues, such as calcitonin-producing cells of the thyroid, melanocytes in the skin, and others.
Cell Death
Apoptosis, or programmed cell death, occurs at various times in development. For example, the fingers are originally webbed during development of the hand. The cells of the webbing later undergo apoptosis, resulting in separation of each individual finger and toe. Apoptosis may occur via apoptotic signals or preprogramming.
During the process of apoptosis, the cell undergoes changes in morphology and divides into many self-contained pieces called apoptotic blebs, which can then be digested by other cells, as shown in Figure 3.9. This allows for recycling of materials. Because the blebs are contained by a membrane, this also prevents the release of potentially harmful substances into the extracellular environment. This is different from necrosis, which is a process of cell death in which a cell dies as a result of injury. In necrosis, internal substances can be leaked, causing irritation of nearby tissues, or even an immune response.
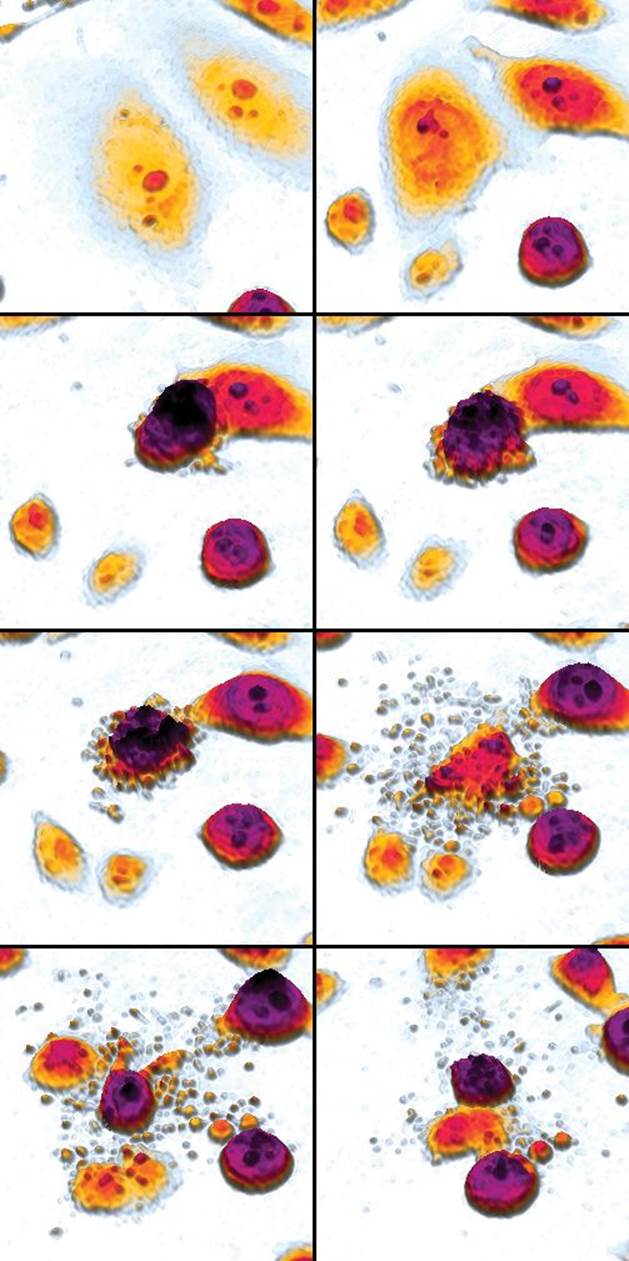 Figure 3.9. Apoptosis An apoptotic cell disintegrates into blebs, which can be absorbed and digested by other cells.
Figure 3.9. Apoptosis An apoptotic cell disintegrates into blebs, which can be absorbed and digested by other cells.
Regeneration
Regenerative capacity, or the ability of an organism to regrow certain parts of the body, varies from species to species. Some species, such as salamanders and newts, have an enhanced capacity to regenerate because they retain extensive clusters of stem cells within their bodies. When regeneration is required, these stem cells can then migrate to the necessary part of the body to initiate regrowth. These species are said to undergo complete regeneration, in that the lost or damaged tissues are replaced with identical tissues. In contrast, incomplete regeneration implies that the newly formed tissue is not identical in structure or function to the tissue that has been injured or lost.
Humans typically exhibit incomplete regeneration in response to injury. However, in humans, regenerative capacity varies by the tissue type. Liver tissue has a high regenerative capacity, often able to undergo extensive regeneration following injury or loss. For example, living donors are often able to donate up to 50 percent of their liver tissue because their own livers will regenerate the missing portion. Unfortunately, the heart has little, if any, regenerative capacity, and scarring often results following an injury such as a heart attack. The kidneys have moderate regenerative capacity and are able to repair nephrons after injury to the tubules; however, this regenerative capacity is easily overwhelmed, and kidney failure may result.
SENESCENCE AND AGING
As organisms age, changes occur in both molecular and cellular structure. This results in disruption of metabolism and, eventually, death of the organism. Senescence, or biological aging, occurs as these changes accumulate and can occur at the cellular and organismal level. At the cellular level, senescence results in the failure of cells to divide, normally after approximately 50 divisions in vitro. Research has demonstrated that this may be due to shortened telomeres, or the ends of chromosomes. Telomeres prevent the loss of genetic information from the ends of chromosomes and help prevent the DNA from unraveling—their high concentration of guanine and cytosine enables telomeres to “knot off” the end of the chromosome. Telomeres are difficult to replicate, however, so they shorten during each round of DNA synthesis. Eventually, the telomeres become too short, and the cell is no longer able to replicate. Some cells, including germ cells, fetal cells, and tumor cells, express an enzyme known as telomerase. This enzyme is a reverse transcriptase that is able to synthesize the ends of chromosomes, preventing senescence. Telomerase allows for cells to divide indefinitely and may play a role in the survival of cancer cells.
At the organismal level, senescence represents changes in the body’s ability to respond to a changing environment. Aging is complex and often involves not only cellular senescence but also the accumulation of chemical and environmental insults over time.
MCAT Concept Check 3.2:
Before you move on, assess your understanding of the material with these questions.
1. What is the difference between determination and differentiation?
· Determination:
· Differentiation:
2. What are the three types of potency? What lineages can a cell of each type differentiate into?
|
Type of Potency |
Cell Lineages |
3. What are the four types of cell–cell communication?
·
·
·
·
4. What is the difference between apoptosis and necrosis?
· Apoptosis:
· Necrosis: