MCAT Biology Review
Chapter 7: The Cardiovascular System
Introduction
As late as the 19th century, physicians adhered to the principle of health known as humoralism. This theory was developed by Greek and Roman physicians and philosophers and adopted by Islamic physicians; it remained dominant in medical thought and practice until it was ultimately displaced by modern medical research in the 1800s. The humoral theory holds that the human body is composed of four fluids or substances called humors—black bile, yellow bile, phlegm, and blood. In the state of health, these four humors are in balance, but excess or deficiency of any one of them would cause illness, disease, and even maladaptive personality characteristics. Over the course of a lifetime, the levels of each of the four humors would rise and fall in accordance with diet and activity, resulting in maladies reflective of the imbalance. Treatments were intended to restore this balance.
Perhaps one of the most well-known treatments associated with humoralism is the practice of bloodletting. Because many diseases were associated with an excess of blood, physicians would rely on the withdrawal of significant amounts of blood from their patients to restore balance to the four humors. Methods for bloodletting were many, and some were dramatic, including drawing blood from major veins in the arm or neck and puncturing arteries. Devices known as scarificators were developed to cut through to the superficial vessels. Most famously, leeches were used, especially in the early 19th century, to draw out the excess blood. In fact, in the early decades of the 1800s, hundreds of millions of leeches were used by European physicians; in the 1830s, France alone imported about 40 million leeches per year for medical treatments.
Now, while the humoral theory has been completely discredited by modern medical research, some practices associated with humoralism are still being used, albeit based on very different medical understanding and for different purposes. For example, new research has shown that medicinal leeches can be used effectively in microsurgery, where they help prevent blood coagulation, and in reconstructive surgery, where they help stimulate circulation to the reattached organ.
The cardiovascular system is one of the most commonly tested on the MCAT. It serves a variety of functions, including the movement of respiratory gases, nutrients, and wastes. We will review the structures and functional anatomy of the cardiovascular system and then discuss blood and its functional components. We’ll also trace the pathways created by the electrically excitable cells that initiate and spread contractions through the heart. A quick recap of genetics and inheritance will help us to understand the functional consequences of ABO and Rh antigens. In addition, the binding capability of oxygen and carbon dioxide to hemoglobin will be discussed in detail.
7.1 Anatomy of the Cardiovascular System
The cardiovascular system consists of a muscular, four-chambered heart, blood vessels, and blood, as shown in Figure 7.1. The heart acts as a pump, circulating blood through the vasculature. The vasculature consists of arteries, capillaries, and veins. After blood travels through veins, it is returned to the right side of the heart where it is pumped to the lungs to be reoxygenated. Then, the oxygenated blood returns to the left side of the heart where it is recirculated to the rest of the body.
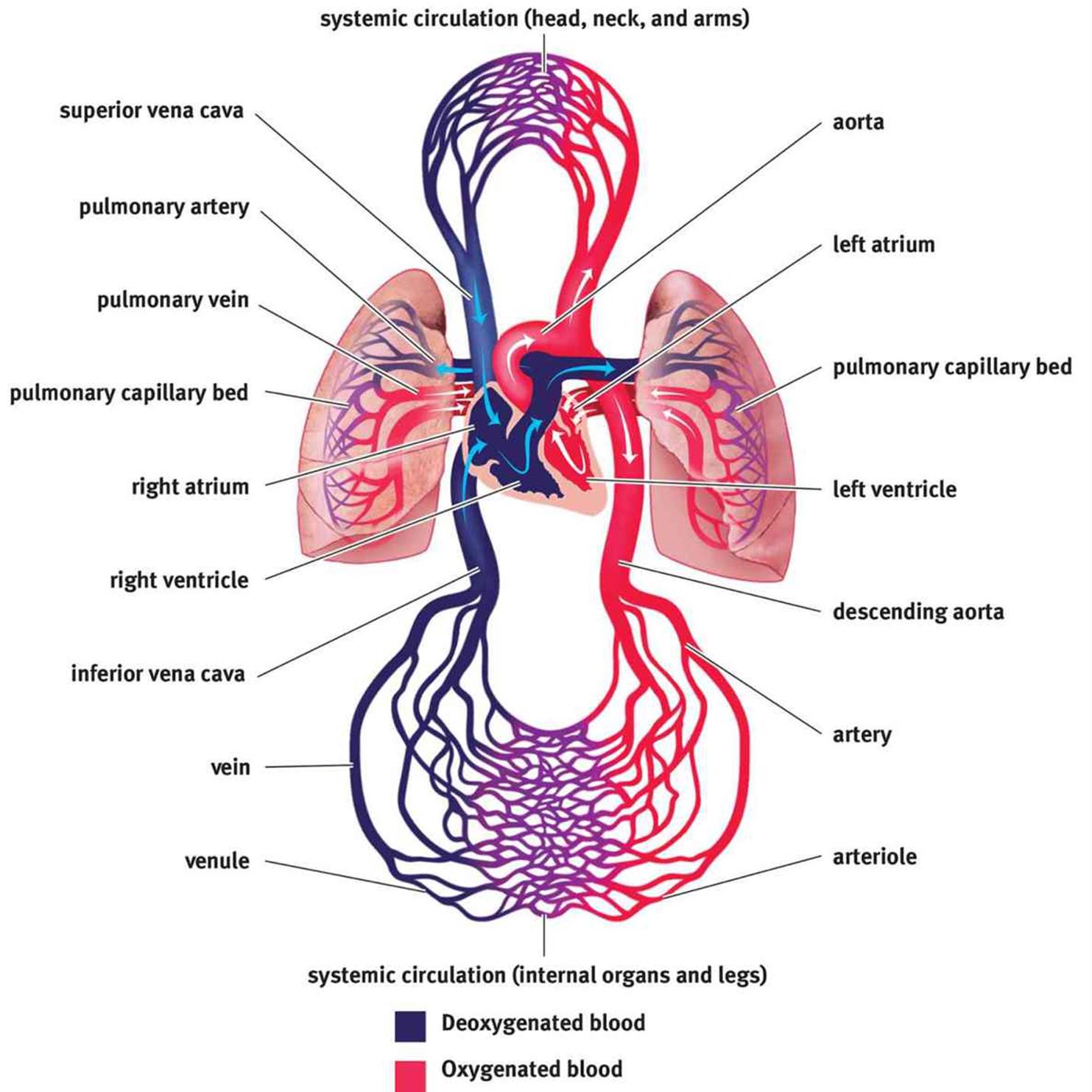 Figure 7.1. Anatomy of the Cardiovascular System
Figure 7.1. Anatomy of the Cardiovascular System
THE HEART
The heart is a four-chambered structure composed predominantly of cardiac muscle. We often speak of the heart as a pump, supporting a single circulatory system. However, the heart is actually composed of two pumps supporting two different circulations in series with one another. The right side of the heart accepts deoxygenated blood returning from the body and moves it to the lungs by way of the pulmonary arteries; this constitutes the first pump (pulmonary circulation). The second pump is the left side of the heart, which receives oxygenated blood from the lungs by way of the pulmonary veins and forces it out to the body through the aorta (systemic circulation).
Each side of the heart consists of an atrium and a ventricle. The atria are thin-walled structures where blood is received from either the venae cavae (deoxygenated blood entering the right heart) or the pulmonary veins (oxygenated blood entering the left heart). The atria contract to push blood into the ventricles. After the ventricles fill, they contract to send blood to the lungs and the systemic circulation. Note that the ventricles are far more muscular than the atria, allowing for more powerful contractions necessary to push blood through the rest of the cardiovascular system.
The atria are separated from the ventricles by the atrioventricular valves. Likewise, the ventricles are separated from the vasculature by the semilunar valves. These valves allow the pump to create the pressure within the ventricles necessary to propel the blood forward within the circulation, while also preventing backflow of blood. The valve between the right atrium and the right ventricle is known as the tricuspid valve (three leaflets), while the valve between the left atrium and the left ventricle is known as the mitral or bicuspid valve (two leaflets). The valve that separates the right ventricle from the pulmonary circulation is known as the pulmonary valve, while the valve that separates the left ventricle from the aorta is known as the aortic valve. Both semilunar valves have three leaflets.
MNEMONIC
· Atrioventricular valves: LAB RAT
· Left Atrium = Bicuspid
· Right Atrium = Tricuspid
One of the central themes in biology is something that we’ve touched on already in previous chapters: structure and function are related. The right and left sides of the heart are two different pumps, with the right heart supporting the circulation to the lungs, while the left heart pumps blood into the systemic circulation. Blood leaving the left heart must travel a considerable distance so that blood pressure can be maintained as far away as the feet. Thus, the left heart is actually more muscular than the right heart. In fact, if the right heart were as muscular as the left and pumped blood as forcefully, this would cause damage to the lungs.
Electric Conduction of the Heart
The coordinated, rhythmic contraction of cardiac muscle originates in an electrical impulse generated by and traveling through a pathway formed by four electrically excitable structures, as shown in Figure 7.2. This commonly tested pathway consists of, in order, the sinoatrial (SA) node, the atrioventricular (AV) node, the bundle of His (AV bundle) and its branches, and the Purkinje fibers. Impulse initiation occurs at the SA node, which generates 60–100 signals per minute without requiring any neural input. This small collection of cells is located in the wall of the right atrium. As the depolarization wave spreads from the SA node, it causes the two atria to contract simultaneously. While most ventricular filling is passive (that is, blood moves from the atria to the ventricles based solely on ventricular relaxation), atrial systole (contraction) results in an increase in atrial pressure that forces a little more blood into the ventricles. This additional volume of blood is called the atrial kick and accounts for about 5–30 percent of cardiac output. Next, the signal reaches the AV node, which sits at the junction of the atria and ventricles. The signal is delayed here to allow for the ventricles to fill completely before they contract. The signal then travels down the bundle of His and its branches, embedded in the interventricular septum (wall), and to the Purkinje fibers, which distribute the electrical signal through the ventricular muscle. The muscle cells are connected by intercalated discs, which contain many gap junctions directly connecting the cytoplasm of adjacent cells, thereby allowing for coordinated ventricular contraction.
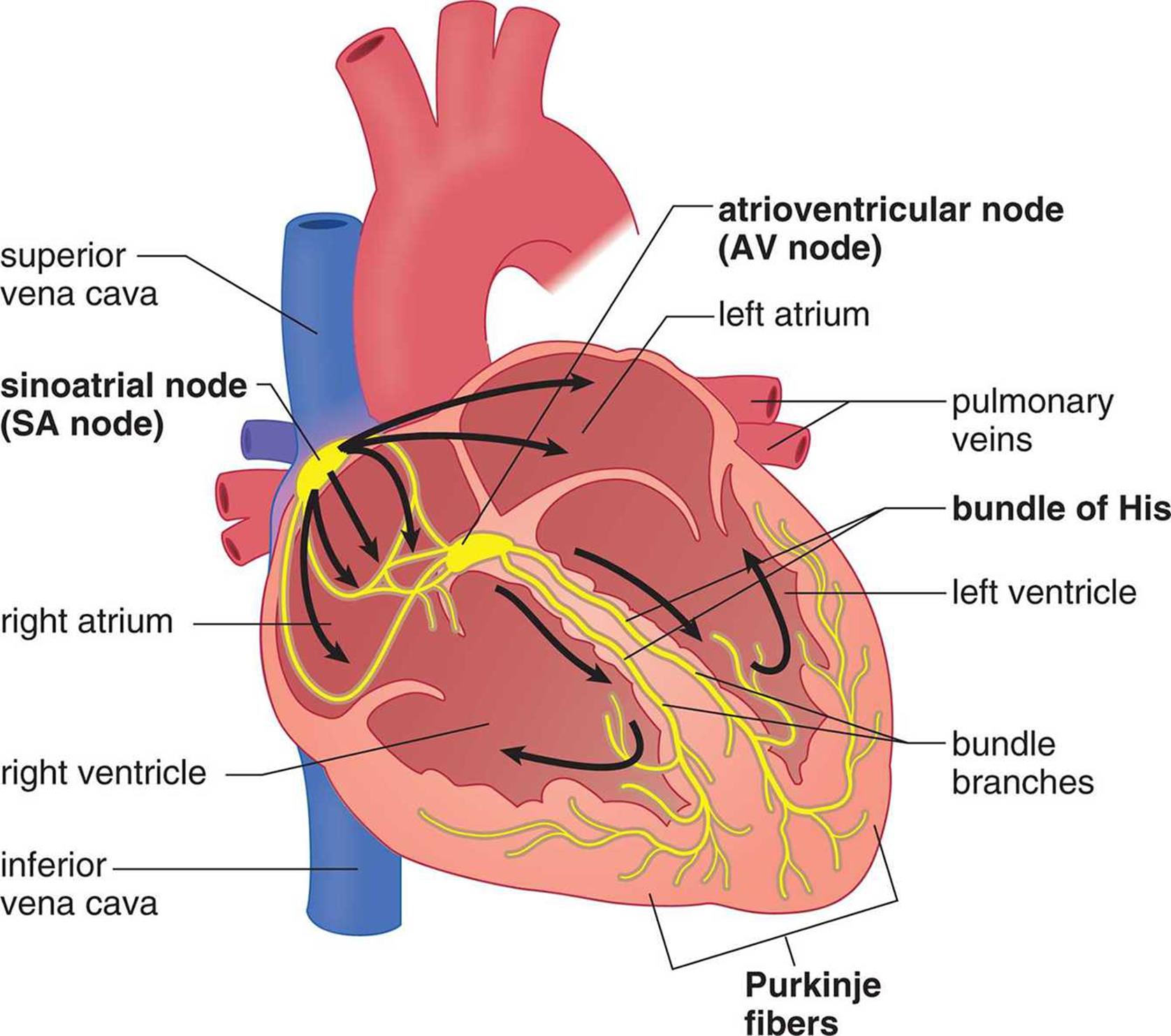 Figure 7.2. Electrical Conduction System of the Heart Electrical impulses travel from the SA node to the AV node, through the bundle of His, and finally to the Purkinje fibers.
Figure 7.2. Electrical Conduction System of the Heart Electrical impulses travel from the SA node to the AV node, through the bundle of His, and finally to the Purkinje fibers.
KEY CONCEPT
Cardiac muscle has myogenic activity, meaning that it can contract without any neural input. The SA node generates about 60–100 beats per minute, even if all innervation to the heart is cut. The neural input to the heart is important in speeding up and slowing the rate of contraction, but not generating it in the first place.
The SA node has an intrinsic rhythm of 60–100 signals per minute, so the normal human heart rate is 60–100 beats per minute. Highly conditioned athletes may have heart rates significantly lower than 60, in the range of 40–50 beats per minute. Stress, exercise, excitement, surprise, or danger can cause the heart rate to rise significantly above 100.
REAL WORLD
The heart’s electrical impulses can be detected on the body’s surface by placing electrodes on the skin on opposite sides of the heart. A recording of these currents is called an electrocardiogram (ECG or EKG; the K, by the way, reflects the German spelling). Electrocardiograms are incredibly powerful tools for assessing the status of a patient’s heart. A normal EKG is shown below.
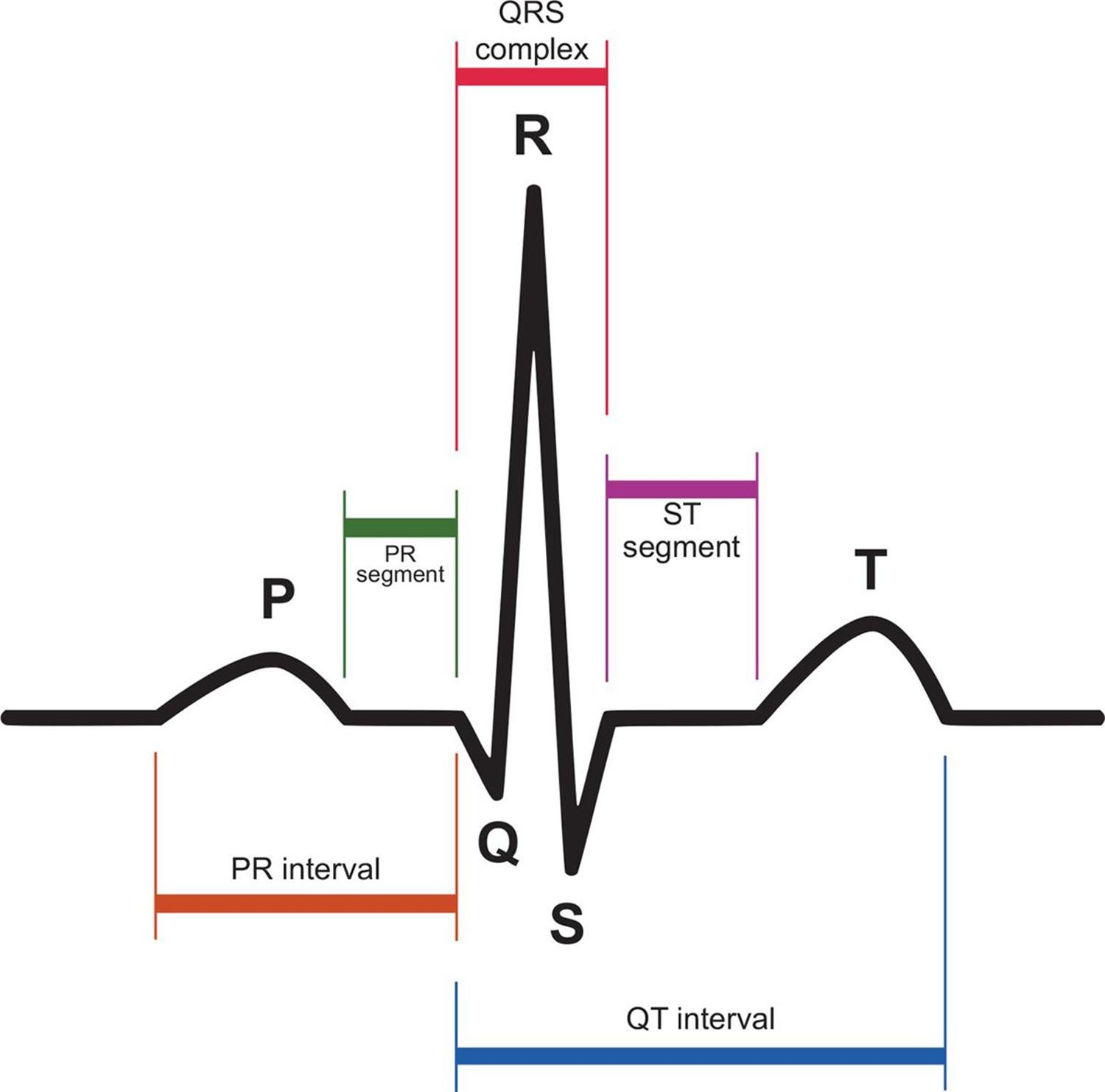
Depolarization precedes cardiac muscle contraction, so the electrical spikes of the EKG occur just before a cardiac contractile event. The P-wave occurs immediately before the atria contract, and the QRS complex occurs just before the ventricles contract. The T-wave represents ventricular repolarization.
The circulatory system is under autonomic control. The autonomic division, which consists of the sympathetic (“fight-or-flight”) and parasympathetic (“rest-and-digest”) branches, controls the heart and affects the vasculature. Sympathetic signals speed up the heart rate and increase the contractility of cardiac muscle, while parasympathetic signals, provided by the vagus nerve, slow it down.
Contraction
The heart is a muscle that must contract in order to move blood. Each heartbeat is composed of two phases, known as systole and diastole. During systole, ventricular contraction and closure of the AV valves occurs and blood is pumped out of the ventricles. During diastole, the heart is relaxed, the semilunar valves are closed, and blood from the atria fills the ventricles. Contraction of the ventricular muscle generates a higher pressure during systole, whereas their relaxation during diastole causes the pressure to decrease. The elasticity of the walls of the large arteries, which stretch to receive the volume of blood from the heart, allows the vessels to maintain sufficient pressure while the ventricular muscles are relaxed. In fact, if it weren’t for the elasticity of the large arteries, your diastolic blood pressure would plummet to zero. The normal events of one heartbeat, including pressures in the left atrium, left ventricle, and aorta; left ventricular volume; normal and pathologic heart sounds; and an EKG are shown in Figure 7.3.
A measure to be aware of is cardiac output, or the total blood volume pumped by a ventricle in a minute. Does it matter which ventricle one chooses? As mentioned previously, the two pumps are connected in series, so the volumes of blood passing through each side must be the same, much like the electrical current between two resistors in series must be the same. Cardiac output (CO) is the product of heart rate (HR, beats per minute) and stroke volume (SV, volume of blood pumped per beat):
CO = HR × SV
Equation 7.1
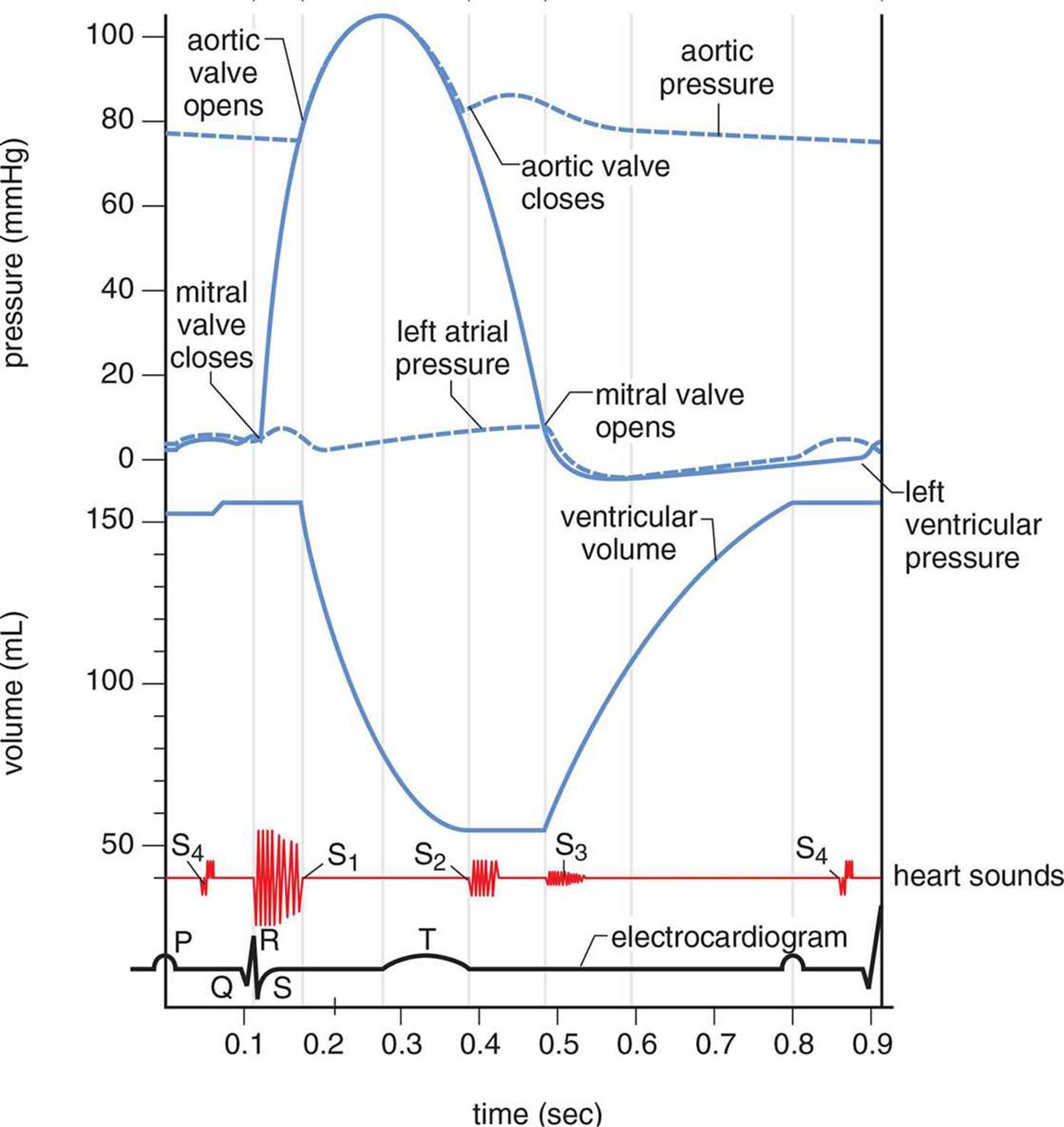 Figure 7.3. The Cardiac Cycle The MCAT will not expect a thorough understanding of every detail of this diagram; it is more important to see how the changes in valves (open or closed), pressures, and volumes are related.
Figure 7.3. The Cardiac Cycle The MCAT will not expect a thorough understanding of every detail of this diagram; it is more important to see how the changes in valves (open or closed), pressures, and volumes are related.
For humans, cardiac output is about 5 liters per minute. During periods of exercise or rest, the autonomic nervous system will increase (sympathetic) or decrease (parasympathetic) cardiac output, respectively.
REAL WORLD
A heart attack, or myocardial infarction, is caused by a lack of bloodflow through the coronary arteries, which results in decreased oxygen delivery to the cardiac muscle itself. The demand on the heart requires too much ATP for anaerobic respiration, so the muscle tissue begins to die. A person suffering a heart attack is often given a β-blocker, which blocks the sympathetic stimulation of the heart, resulting in lower heart rate and lower contractility. With a β-blocker, the heart does not work as hard, so its oxygen demand is diminished, which helps to prevent further damage to cardiac tissue.
REAL WORLD
The rhythmic impulses we hear when we listen to someone’s heart with a stethoscope (lub dub) are referred to as the heart sounds. The first sound, S1, is produced when the two AV valves close at the start of systole to prevent backflow into the atria. The second sound, S2, is produced when the two semilunar valves close at the end of systole to prevent backflow into the ventricles. Extra heart sounds (S3 and S4) can result from stiffness of the heart muscle or high blood pressure. Heart murmurs, which may be so loud as to be audible without a stethoscope, may arise when the valves malfunction and become either narrow and stiff or wide and floppy, resulting in abnormal flow patterns across the valve.
REAL WORLD
There is a limit to how fast the heart can beat and still pump blood effectively. Because the heart fills with blood when it is relaxing (diastole), the faster it beats, the less time there is for blood to enter the heart during relaxation. Thus, a faster heartbeat means diminishing returns in terms of the amount of blood supplied to the body. A dangerous condition called ventricular tachycardia (often abbreviated v-tach) describes rates upward of 200 beats per minute. The heart in v-tach cannot properly fill with blood and, paradoxically, stops pumping blood despite its fast rate. Systemic pressures drop precipitously. Death will result unless the heart is forced out of this abnormal rhythm.
THE VASCULATURE
In order to deliver blood to the entire body, the circulatory system utilizes vessels of different sizes. The three major types of vessels are arteries, veins, and capillaries. Blood travels away from the heart in arteries, the largest of which in the systemic circulation is the aorta. Major arteries, such as the coronary, common carotid, and renal arteries, divide the bloodflow from the aorta toward different peripheral tissues. Arteries branch into arterioles, which ultimately lead to capillaries that perfuse the tissues. On the venous side of a capillary network, the capillaries join together into venules, which join to form veins. Venous blood empties into the superior and inferior venae cavae for entry into the right side of the heart. All blood vessels are lined with endothelial cells. This special type of cell helps to maintain the vessel by releasing chemicals that aid in vasodilation and vasoconstriction. In addition, endothelial cells can allow white blood cells to pass through the vessel wall and into the tissues during an inflammatory response. Finally, endothelial cells release certain chemicals when damaged that are necessary in the formation of blood clots to repair the vessel and stop bleeding.
MNEMONIC
Arteries carry blood away from the heart.
Cross-sections of the different blood vessels are shown in Figure 7.4. Don’t worry about the names of the layers; simply be able to recognize that the same types of cells comprise the different vessels and that arteries have much more smooth muscle than veins.
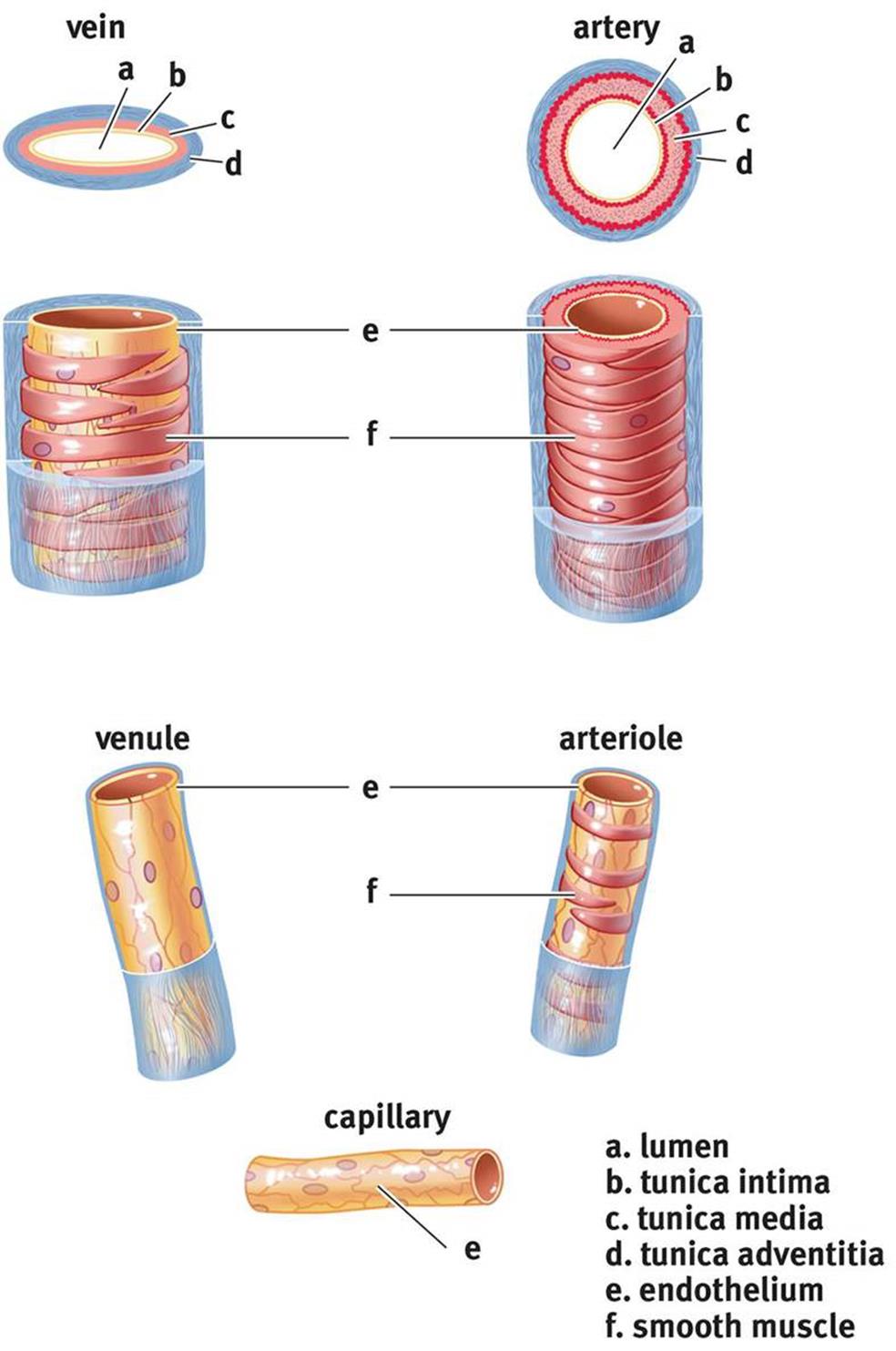 Figure 7.4. Structure of Blood Vessels
Figure 7.4. Structure of Blood Vessels
Arteries
Arteries move blood away from the heart to the lungs and other parts of the body. Most arteries contain oxygenated blood; in fact, only the pulmonary arteries and umbilical arteries contain deoxygenated blood. Smaller, muscular arteries are known as arterioles.
Arteries are highly muscular and elastic, creating tremendous resistance to the flow of blood. This is one of the reasons why the left heart must generate much higher pressures: to overcome the resistance caused by systemic arteries. After arteries are filled with blood, the elastic recoil from their walls maintains a high pressure and forces blood forward.
Capillaries
Capillaries are vessels with a single endothelial cell layer and are so small that red blood cells must pass through the capillaries in a single-file line. The thin wall of the capillary allows easy diffusion of gases (O2 and CO2), nutrients (most notably, glucose), and wastes (ammonia and urea, among others). Capillaries are therefore the interface for communication of the circulatory system with the tissues. Remember, too, that blood also carries hormones, so capillaries allow endocrine signals to arrive at their target tissues.
Capillaries can be quite delicate. When capillaries are damaged, blood can leave the capillaries and enter the interstitial space. If this occurs in a closed space, it results in a bruise.
Veins
Veins are thin-walled, inelastic vessels that transport blood to the heart. Except for the pulmonary and umbilical veins, all veins carry deoxygenated blood. Venules are smaller venous structures that connect capillaries to the larger veins of the body.
The smaller amount of smooth muscle in the walls of veins gives them less recoil than arteries. Furthermore, veins are able to stretch to accommodate larger quantities of blood. Indeed, three-fourths of our total blood volume may be in venous circulation at any one time. Note that, even though the volume of arterial blood is normally much less than the volume of venous blood, the total volume passing through either side of the heart per time (cardiac output) is the same.
Given that the heart is located in the chest, bloodflow in most veins is upward from the lower body back to the heart, against gravity. In the inferior vena cava, this translates into a large amount of blood in a vertical column. The pressure at the bottom of this venous column in the large veins of the legs can be quite high. In fact, it can exceed systolic pressure (120 mmHg), going as high as 200 mmHg or more. Thus, veins must have structures to push the blood forward and prevent backflow. Larger veins contain valves; as blood flows forward in the veins, the valves open. When blood tries to move backward, the valves will slam shut. Failure of the venous valves can result in the formation of varicose veins, which are distended where blood has pooled. Pregnant women are especially susceptible to the formation of varicose veins due to an increase in the total blood volume during pregnancy and compression of the inferior vena cava by the fetus.
REAL WORLD
Blood clots may form in the deep veins of the legs as a result of injury, inactivity (blood stasis), or a hypercoagulable state (a tendency for the blood to clot excessively). The clots may dislodge and travel through the right atrium and right ventricle, out the pulmonary artery, and into the lungs. Such clots, called pulmonary emboli (or, more specifically, thromboemboli), block segments of the pulmonary arteries and produce rapid, labored breathing and chest pain. Death may occur if the thromboemboli are large.
In addition to high pressure in the lower extremities, the small amount of smooth muscle also creates a challenge for propelling blood forward. Thus, the veins must rely on an external force to generate the pressure to push blood toward the heart. Most veins are surrounded by skeletal muscles, which squeeze the veins as the muscles contract, forcing the blood up against gravity in much the same way that squeezing the bottom of a tube of toothpaste causes the contents to be expelled through the top of the tube. This is why sitting motionless for long periods of time, such as in a cramped middle seat on a long transoceanic flight or after surgery, can increase the risk of blood clot formation in the veins of the legs and pelvis. Blood pools in the lower extremities, and sluggish blood coagulates more easily. A clot in the deep veins of the leg is called a deep vein thrombosis (DVT). This clot may become dislodged and travel through the right heart to the lungs, where it can cause a life-threatening condition called a pulmonary embolus.
REAL WORLD
Many patients may be immobile following a surgical procedure or during a long hospital stay. This causes a threat of DVT and pulmonary emboli, which are very undesirable complications in someone who is already sick. Thus, in hospitals, special wraps are placed on the legs that contract rhythmically in order to prevent pooling of blood and the formation of blood clots. In addition, many patients receive medications like heparin or warfarin to prevent the formation of clots.
CIRCULATION
Circulation is, by definition, circular. Let’s return to the anatomy of the heart and vasculature, shown in Figure 7.1 earlier to determine the flow of blood through the body. A close-up of bloodflow through the heart is also shown in Figure 7.5. Here, we begin with the return of blood to the right atrium. Blood returns to the heart from the body via the venae cavae, which are divided into the superior vena cava (SVC) and the inferior vena cava (IVC). The superior vena cava returns blood from the portions of the body above the heart, while the inferior vena cava returns blood from portions of the body below the heart. Deoxygenated blood enters the right atrium, travels through the tricuspid valve, and enters the right ventricle. On contraction, the blood from the right ventricle passes through the pulmonary valve and enters the pulmonary arteries, where it travels to the lungs in continuously smaller vessels. Once the blood reaches the capillaries that line the alveoli, it participates in gas exchange, with carbon dioxide leaving the blood and oxygen entering the blood. The blood then travels into pulmonary venules and into the pulmonary veins, which carry the blood to the left side of the heart. Oxygenated blood enters the left atrium, travels through the mitral valve, and enters the left ventricle. On contraction, the blood from the left ventricle passes through the aortic valve and enters the aorta. From the aorta, blood enters arteries, then arterioles, and then capillaries. After gas and nutrient exchange occurs at the capillaries, the blood enters the venules, which lead to the larger veins. The veins then empty into either the SVC or IVC for return to the right side of the heart.
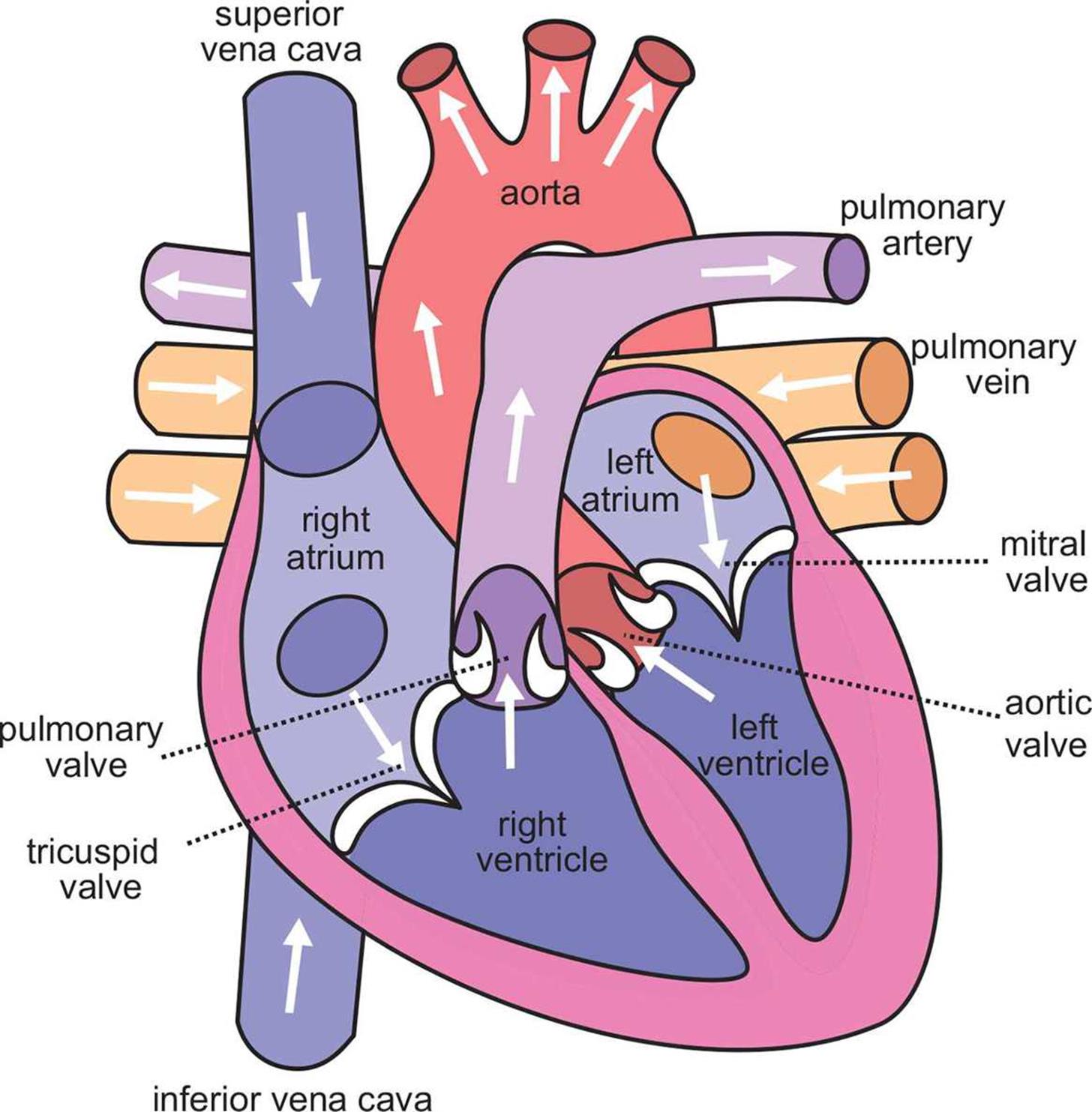 Figure 7.5. Bloodflow through the Heart
Figure 7.5. Bloodflow through the Heart
Written in shorthand, the pathway appears like this:

REAL WORLD
While bicuspid valve is an acceptable alternative name for mitral valve, it is rarely used in medicine. Most practitioners will refer to valves by one-letter abbreviations (M for mitral). The name mitral refers to a miter, which is the name for the hat the pope and bishops wear in the Catholic faith. The two large leaflets of the mitral valve somewhat resemble this headdress.
In most cases, blood will pass through only one capillary bed before returning to the heart. However, there are three portal systems in the body, in which blood will pass through two capillary beds in series before returning to the heart. In the hepatic portal system, blood leaving capillary beds in the walls of the gut passes through the hepatic portal vein before reaching the capillary beds in the liver. In the hypophyseal portal system, blood leaving capillary beds in the hypothalamus travels to a capillary bed in the anterior pituitary to allow for paracrine secretion of releasing hormones. In the renal portal system, blood leaving the glomerulus travels through an efferent arteriole before surrounding the nephron in a capillary network called the vasa recta.
MCAT Concept Check 7.1:
Before you move on, assess your understanding of the material with these questions.
1. Starting from entering the heart from the venae cavae, what are the four chambers through which blood passes in the heart? Which valve prevents backflow into each chamber?
|
Heart Chamber |
Valve that Prevents Backflow |
2. Starting with the site of impulse initiation, what are the structures in the conduction system of the heart?
3. Compare and contrast arteries, capillaries, and veins:
|
Vessel |
Carries Blood Which Direction? |
Relative Wall Thickness |
Smooth Muscle Present? |
Contains Valves? |
|
Artery |
||||
|
Capillary |
||||
|
Vein |
4. Why does the right side of the heart contain less cardiac muscle than the left side?
5. If all autonomic input to the heart were cut, what would happen?