MCAT Biology Review
Chapter 7: The Cardiovascular System
7.3 Physiology of the Cardiovascular System
The cardiovascular system transports many compounds, including gases, nutrients, and waste products, to and from the body’s tissues through the red blood cells and plasma. Furthermore, it serves an important role in immunity through specialized cells, such as leukocytes, which help the body fight localized or systemic pathogens. Capillaries within the body can dilate and constrict to maintain proper body temperature. In addition, the circulatory system mediates the formation of blood clots to repair damaged vessels. These functions reflect the important jobs of the cardiovascular system, which include maintenance of blood pressure, gas and solute exchange, coagulation, and thermoregulation.
BLOOD PRESSURE
Before we can even discuss gas and solute exchange, it is important to recognize that, for the circulatory system to serve its predominant functions, blood pressure must be kept sufficiently high to propel blood forward. Blood pressure, therefore, provides healthcare professionals with information regarding the effectiveness of the circulatory system. In addition, high blood pressure, or hypertension, is a pathological state that may result in damage to the blood vessels and organs. Blood pressure is a measure of the force per unit area exerted on the wall of the blood vessels and is measured with a sphygmomanometer. Sphygmomanometers measure the gauge pressure in the systemic circulation, which is the pressure above and beyond atmospheric pressure (760 mmHg at sea level), as discussed in Chapter 4 of MCAT Physics and Math Review. Blood pressure is expressed as a ratio of the systolic (ventricular contraction) to diastolic (ventricular relaxation) pressures. Pressure gradually drops from the arterial to venous circulation, with the largest drop occurring across the arterioles, as shown in Figure 7.8. Normal blood pressure is considered to be between 90/60 and 120/80.
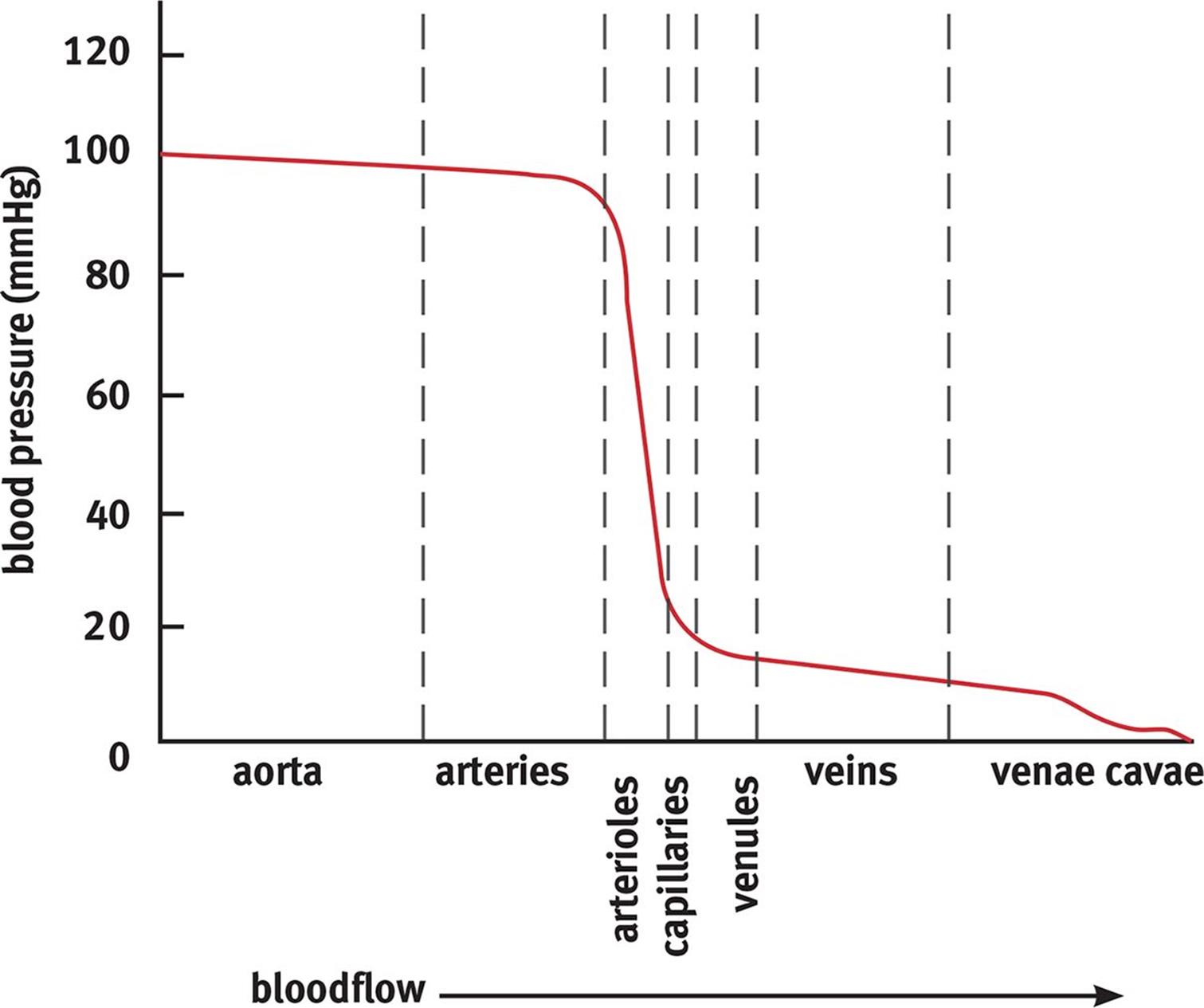 Figure 7.8. Mean Arterial Pressure at Different Locations in the Cardiovascular System
Figure 7.8. Mean Arterial Pressure at Different Locations in the Cardiovascular System
KEY CONCEPT
The largest drop in blood pressure occurs across the arterioles. This is necessary because the capillaries are thin-walled and unable to withstand the pressure of the arterial side of the vasculature.
An analogy can be drawn between circulation and an electric circuit. Much like an electromotive force (voltage) drives a current through a given electrical resistance, the pressure gradient across the circulatory system drives cardiac output through a given vascular resistance. This analogy is an important one to remember because the equations of electric circuits can be applied to the cardiovascular system. For example, Ohm’s law (V = IR) can be translated into the following equation in circulation:
ΔP = CO × TPR
Equation 7.2
where ΔP is the pressure differential across the circulation, CO is the cardiac output, and TPR is the total peripheral (vascular) resistance.
It is also important to note that arterioles and capillaries act much like resistors in a circuit. When electricity travels through a wire, the wire itself provides an intrinsic level of resistance that limits the flow of electricity through it. Resistance is based on three factors: resistivity, length, and cross-sectional area. Resistivity has no obvious correlate in physiology, but the other two factors certainly do. The longer a blood vessel is, the more resistance it offers. The larger the cross-sectional area of a blood vessel, the less resistance it offers. In addition, arteries are highly muscular and are able to expand and contract as needed to change vascular resistance and maintain blood pressure. Arterioles can also contract to limit the amount of blood entering a given capillary bed (much like increasing resistance will decrease current flow to a given branch in a circuit). Finally, with the exception of the three portal systems, all systemic capillary beds are in parallel with each other. Therefore, opening capillary beds will decrease vascular resistance (like adding another resistor in parallel) and, assuming the body can compensate, increase cardiac output.
REAL WORLD
Vital signs include temperature, heart rate, respiratory rate, and blood pressure. They provide a quick snapshot of a person’s condition at a given moment. High blood pressure may indicate clinical hypertension, anxiety, catecholamine-producing tumors, or stimulant use, while low blood pressure may indicate hemorrhage, a heart attack, sepsis (severe bloodstream infection), or neurological damage. These are very broad differential diagnoses; however, vital signs combined with a quick clinical history provide a way for clinicians to identify the patients that require immediate intervention.
Blood pressure is regulated using baroreceptors in the walls of the vasculature. Baroreceptors are specialized neurons that detect changes in the mechanical forces on the walls of the vessel. When the blood pressure is too low, they can stimulate the sympathetic nervous system, which causes vasoconstriction, thereby increasing the blood pressure. In addition, chemoreceptors can sense when the osmolarity of the blood is too high, which could indicate dehydration. This promotes the release of antidiuretic hormone (ADH or vasopressin), a peptide hormone made in the hypothalamus but stored in the posterior pituitary, which increases the reabsorption of water, thereby increasing blood volume and pressure (while also diluting the blood). Low perfusion to the juxtaglomerular cells of the kidney stimulates aldosterone release through the renin–angiotensin–aldosterone system; aldosterone increases the reabsorption of sodium (and, by extension, water), thereby increasing the blood volume and pressure.
So, what if blood pressure is too high? Neurologically, sympathetic impulses could decrease, permitting relaxation of the vasculature with a concurrent drop in blood pressure. Within the heart, specialized atrial cells are able to secrete a hormone called atrial natriuretic peptide (ANP). This hormone aids in the loss of salt within the nephron, acting as a natural diuretic with loss of fluid. Interestingly, ANP is a fairly weak diuretic. Some fluid is lost, but it is often not enough to counter the effects of a high-salt diet on blood pressure. Indeed, the human body has many different ways to raise blood pressure, but very few ways to lower it.
GAS AND SOLUTE EXCHANGE
Blood pressure ensures sufficient forward flow of blood through the system. However, what happens when the blood reaches the capillaries? Here, oxygen and nutrients diffuse out of the blood into tissues, while waste products like carbon dioxide, hydrogen ions, urea, and ammonia diffuse into the blood. In addition, hormones are secreted into the capillaries, travel with the circulation, and diffuse into their target tissue. Ions and fluid must also be returned to the blood to ensure that no area becomes too swollen with fluid. Regardless of the substance being exchanged, there is one fundamental concept to be considered in this process: concentration gradients. In each case, one side of the capillary wall has a higher concentration of a given substance than the other. This allows for movement of gases and solutes by diffusion.
Oxygen
Oxygen is carried primarily by hemoglobin in the blood. Hemoglobin is a protein composed of four cooperative subunits, each of which has a prosthetic heme group that binds to an oxygen molecule. The binding of oxygen occurs at the heme group’s central iron atom, which can undergo changes in its oxidation state. The binding or releasing of oxygen to or from the iron atom in the heme group is an oxidation–reduction reaction. It is also important to note that some oxygen does diffuse into the blood and dissolve into the plasma, but this amount is negligible compared to the quantity of oxygen bound to hemoglobin. The level of oxygen in the blood is often measured as the partial pressure of O2 within the blood, or PaO2. A normal PaO2 is approximately 70–100 mmHg. However, this measurement is inconvenient because it involves taking a sample of blood from an artery. Oxygen saturation—that is, the percentage of hemoglobin molecules carrying oxygen—is easily measured using a finger probe. Most healthy people have an oxygen saturation above 97%.
In the lungs, oxygen diffuses into the alveolar capillaries. As the first oxygen binds to a heme group, it induces a conformational shift in the shape of hemoglobin from taut to relaxed. This shift increases hemoglobin’s affinity for oxygen, making it easier for subsequent molecules of oxygen to bind to the remaining three unoccupied heme groups. As other heme groups acquire an oxygen molecule, the affinity continues to increase, thus creating a positive feedback-like (spiraling forward) mechanism. Once all of the hemoglobin subunits are bound to oxygen, the removal of one molecule of oxygen will induce a conformational shift, decreasing the overall affinity for oxygen, and making it easier for the other molecules of oxygen to leave the heme groups. This is again a spiraling forward process; as oxygen molecules leave hemoglobin, it becomes progressively easier for more oxygen to be removed. This phenomenon is a form of allosteric regulation referred to as cooperative binding and results in the classic sigmoidal (S-shaped) oxyhemoglobin dissociation curve shown in Figure 7.9.
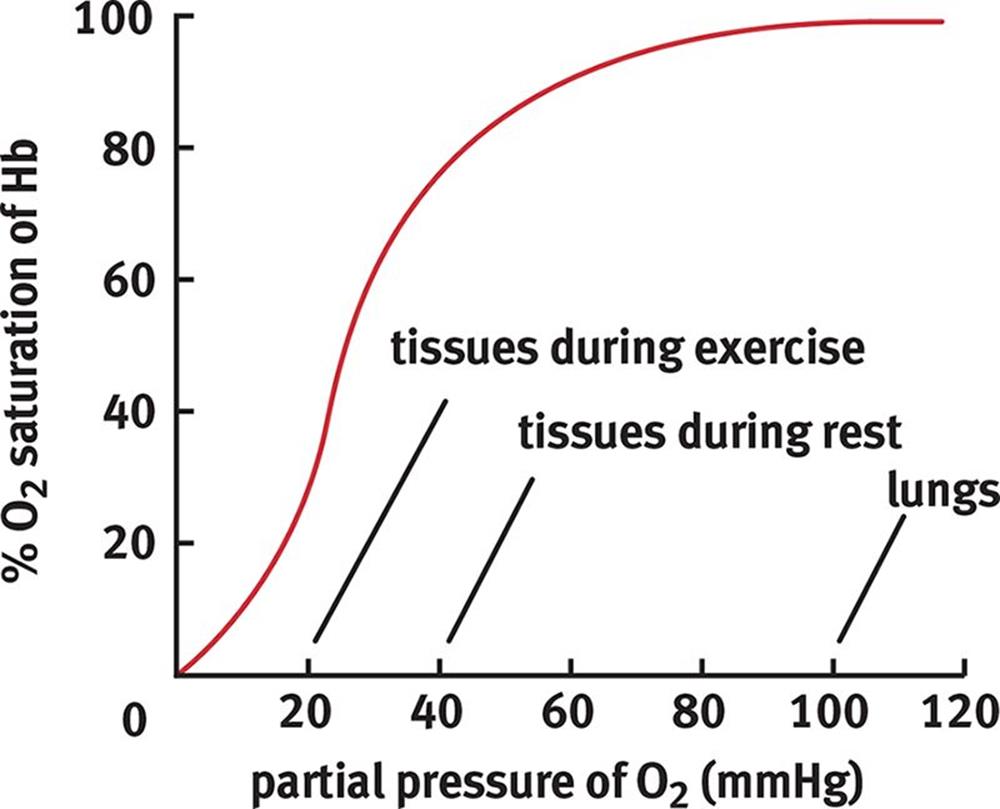 Figure 7.9. Oxyhemoglobin Dissociation Curve
Figure 7.9. Oxyhemoglobin Dissociation Curve
Before looking at shifts in this curve, let’s make sure we understand what everything means. According to the curve, the blood is 100% saturated in the lungs, at a partial pressure of 100 mmHg O2. The tissues are at a lower partial pressure of oxygen, around 40 mmHg during rest; at this lower partial pressure, the hemoglobin is approximately 80% saturated. Therefore, about 100 – 80 = 20% of the oxygen has been released from the hemoglobin. Where did this oxygen go? Into the tissues, of course.
During exercise, the partial pressure of oxygen in the tissues is even lower—around 20 mmHg. At this lower partial pressure, the hemoglobin is approximately 30% saturated. Therefore, about 100 – 30 = 70% of the oxygen has been released to the tissues. In reality, unloading of oxygen is also facilitated by shifts in the hemoglobin curve that occur during exercise, as described later.
Carbon Dioxide
Delivering oxygen to tissues is only part of the job of transporting respiratory gases; removing carbon dioxide gas (CO2), the primary waste product of cellular respiration, is also important. Carbon dioxide gas, like oxygen gas, is nonpolar and therefore has low solubility in the aqueous plasma; only a small percentage of the total CO2 being transported in the blood to the lungs will be dissolved in the plasma. Carbon dioxide can be carried by hemoglobin, but hemoglobin has a much lower affinity for carbon dioxide than for oxygen. The vast majority of CO2 exists in the blood as the bicarbonate ion (HCO−3). When CO2 enters a red blood cell, it encounters the enzyme carbonic anhydrase, which catalyzes the combination reaction between carbon dioxide and water to form carbonic acid (H2CO3). Carbonic acid, a weak acid, will dissociate into a proton and the bicarbonate anion. The hydrogen ion (proton) and bicarbonate ion both have high solubilities in water, making them a more effective method of transporting metabolic waste products to the lungs for excretion. Upon reaching the alveolar capillaries in the lungs, the same reactions that led to the formation of the proton and bicarbonate anion can be reversed, allowing us to breathe out carbon dioxide:
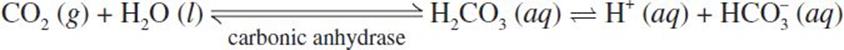
This chemical reaction is important, not only because it provides an effective means of ridding the body’s tissues of carbon dioxide gas, but also because the concentration of free protons in the blood affects pH; the pH, in turn, can have allosteric effects on the oxyhemoglobin dissociation curve. Increased carbon dioxide production will cause a right shift in the bicarbonate buffer equation, resulting in decreased pH (increased [H+]). These protons can bind to hemoglobin, reducing hemoglobin’s affinity for oxygen. This decreased affinity can be seen in the oxyhemoglobin curve as a shift to the right; this is known as the Bohr effect. Note that the triggers for this right shift (increased PaCO2, increased [H+], decreased pH) are often associated with oxygen demand; higher rates of cellular metabolism result in increased carbon dioxide production and accumulation of lactic acid, both of which decrease pH. This decreased affinity allows more oxygen to be unloaded at the tissues, as shown in Figure 7.10. Looking at the red and green lines, we see that hemoglobin is nearly 100% saturated in the lungs (at a partial pressure of 100 mmHg O2) for both lines. However, the green line is significantly lower than the red one when we reach a partial pressure of 20 mmHg O2, around that of exercising muscle. Therefore, the right shift results in a greater unloading of oxygen into the tissues.
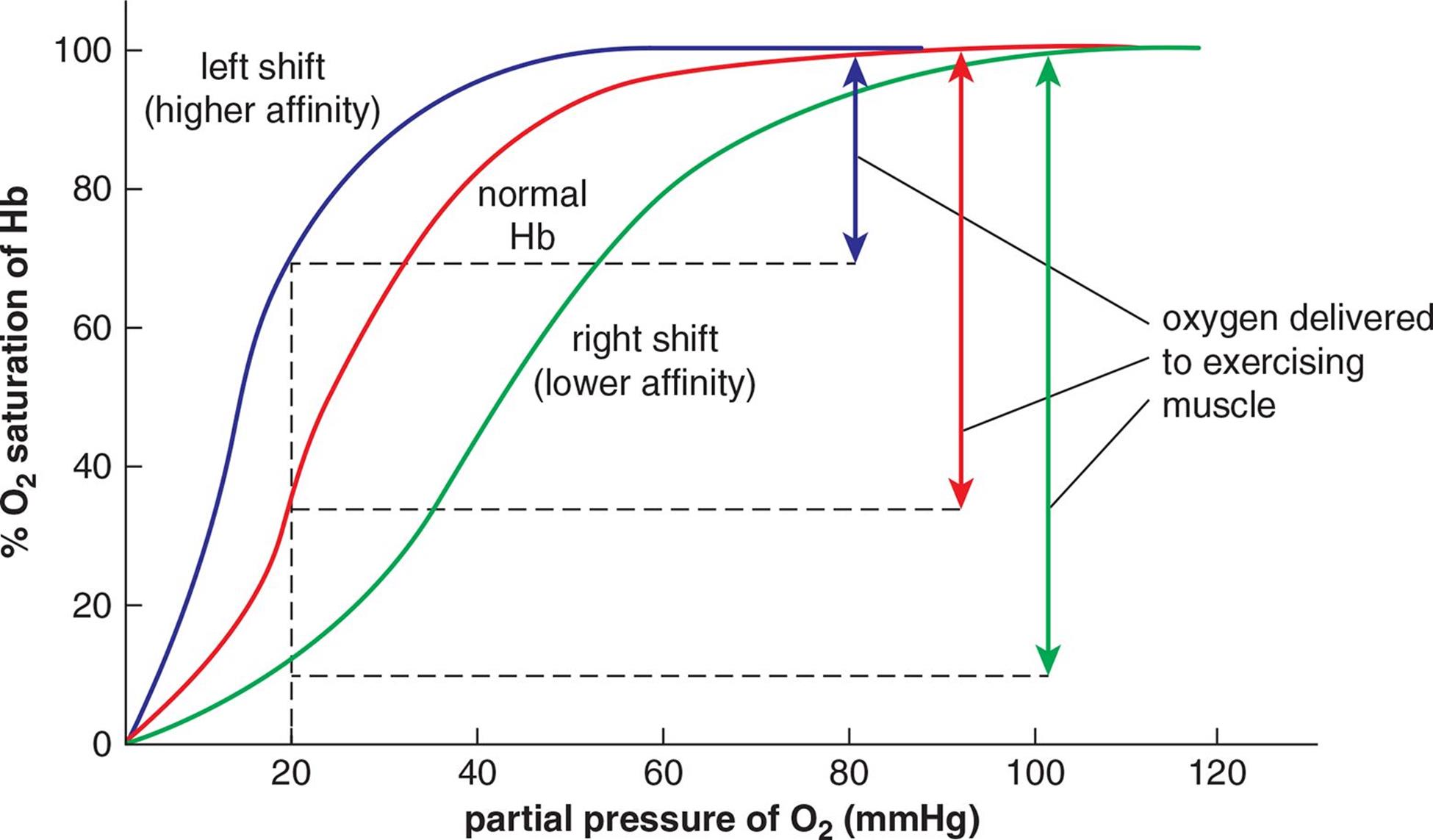 Figure 7.10. Shifts in the Oxyhemoglobin Dissociation Curve
Figure 7.10. Shifts in the Oxyhemoglobin Dissociation Curve
MNEMONIC
Causes of a right shift of the oxyhemoglobin curve: Exercise is the right thing to do
The following occur during exercise:
· Increased PaCO2
· Increased [H+] (decreased pH)
· Increased temperature
Other causes of a right shift in the oxyhemoglobin curve include increased temperature and increased 2,3-bisphosphoglycerate (2,3-BPG), a side product of glycolysis in red blood cells.
A left shift, like the blue line in Figure 7.10, may occur due to decreased PaCO2, decreased [H+], increased pH, decreased temperature, and decreased 2,3-BPG. In addition, fetal hemoglobin (HbF) has a higher affinity for oxygen than adult hemoglobin (HbA). This should make sense because fetal red blood cells must literally pull oxygen off of maternal hemoglobin and onto fetal hemoglobin.
KEY CONCEPT
Fetal hemoglobin has a higher affinity for oxygen than adult hemoglobin, and therefore has a left-shifted curve compared to adult hemoglobin.
The bicarbonate buffer system is also important because it links the respiratory and renal systems. Disturbances in either of these systems can lead to changes in the pH of the blood. For example, if an individual hyperventilates, excess CO2 will be blown off, shifting the bicarbonate buffer system to the left and decreasing the concentration of protons. This leads to an increase in pH, or what is known as respiratory alkalosis. The kidney can compensate for this change by increasing excretion of bicarbonate, which brings the pH back to normal. In contrast, in renal tubular acidosis type I, the kidney is unable to excrete acid effectively. This leads to a buildup of protons in the blood (metabolic acidosis), which causes the buffer system to shift to the left. The excess CO2 formed in the process can be exhaled, and the person may increase respiratory rate to compensate, bringing the pH back to normal.
Nutrients, Waste, and Hormones
In addition to respiratory gases, blood also carries nutrients, waste products, and hormones to the appropriate location for use or disposal. As discussed earlier, concentration gradients guide much of the movement of these substances to and from the tissues.
Carbohydrates and amino acids are absorbed into the capillaries of the small intestine and enter the systemic circulation via the hepatic portal system. Fats are absorbed into lacteals in the small intestine, bypassing the hepatic portal circulation to enter systemic circulation via the thoracic duct. Once in the bloodstream, fats are packaged in lipoproteins, which are water-soluble. The absorption of nutrients is covered more extensively in Chapter 9 of MCAT Biology Review.
Wastes, such as carbon dioxide, ammonia, and urea, enter the bloodstream by traveling down their respective concentration gradients from the tissues to the capillaries. The blood eventually travels to the kidney, where these waste products are filtered or secreted for elimination from the body.
Hormones enter the circulation in or near the organ where the hormone is produced. This usually occurs by exocytosis, allowing for secretion of hormones into the bloodstream. Certain hormones are carried by proteins in the blood and are released under specific conditions. Once hormones reach their target tissues, they can activate cell-surface receptors (peptide hormones) or diffuse into the cell to activate intracellular or intranuclear receptors (steroid hormones).
Fluid Balance
In the bloodstream, two pressure gradients are essential for maintaining a proper balance of fluid volume and solute concentrations between the blood and the interstitium (the cells surrounding the blood vessels). These are the opposing but related hydrostatic and osmotic (oncotic) pressures.
Hydrostatic pressure is the force per unit area that the blood exerts against the vessel walls. This is generated by the contraction of the heart and the elasticity of the arteries, and can be measured upstream in the large arteries as blood pressure. Hydrostatic pressure pushes fluid out of the bloodstream and into the interstitium through the capillary walls, which are somewhat leaky by design. Osmotic pressure, on the other hand, is the “sucking” pressure generated by solutes as they attempt to draw water into the bloodstream. Because most of this osmotic pressure is attributable to plasma proteins, it is usually called oncotic pressure.
At the arteriole end of a capillary bed, hydrostatic pressure (pushing fluid out) is much larger than oncotic pressure (drawing fluid in), and there is a net efflux of water from the circulation, as shown in Figure 7.11. As fluid moves out of the vessels, the hydrostatic pressure drops significantly, but the osmotic pressure stays about the same. Therefore, at the venule end of the capillary bed, hydrostatic pressure (pushing fluid out) has dropped below oncotic pressure (drawing fluid in), and there is a net influx of water back into the circulation.
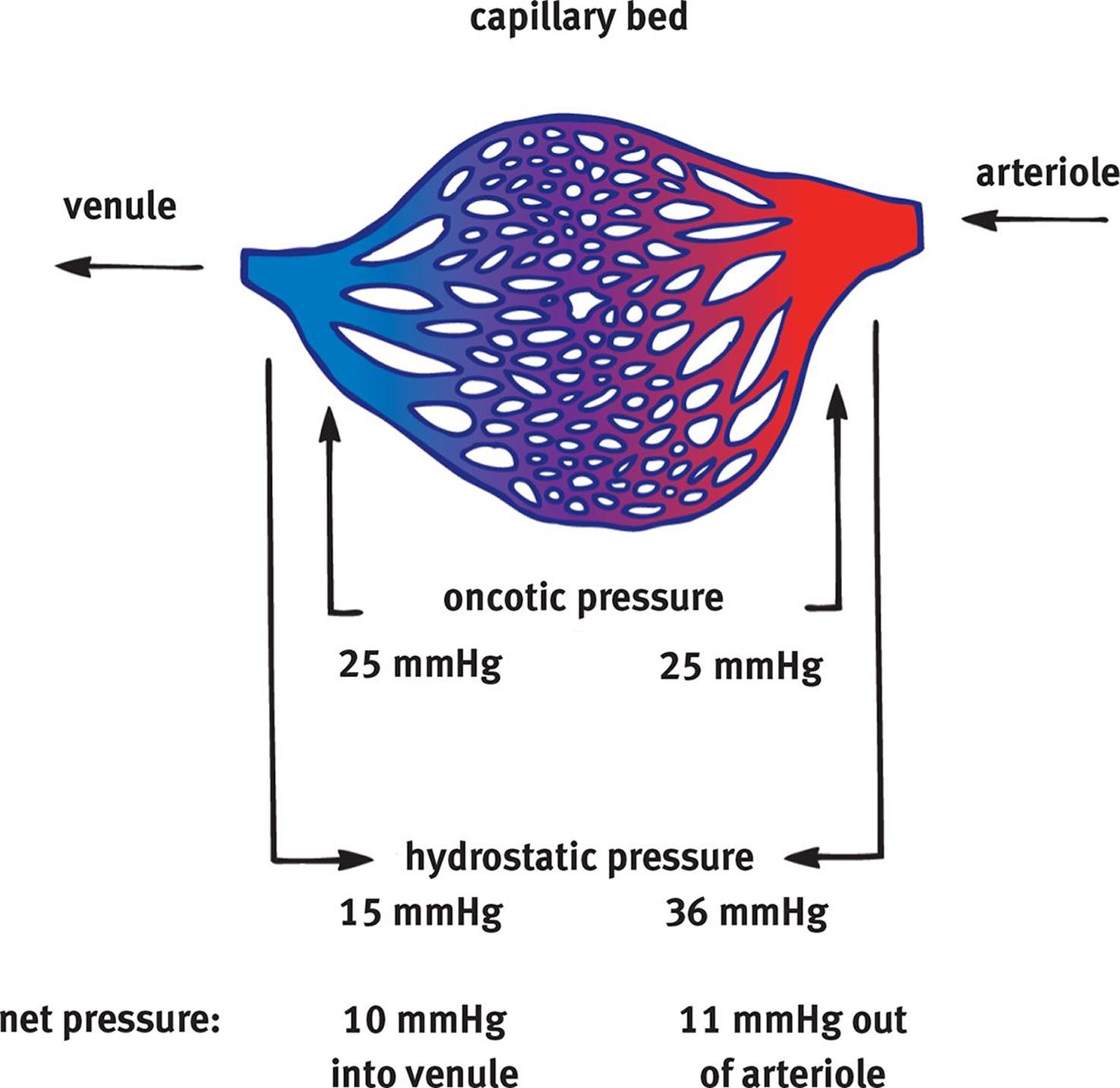 Figure 7.11. Starling Forces at a Capillary Bed
Figure 7.11. Starling Forces at a Capillary Bed
KEY CONCEPT
Hydrostatic pressure pushes fluid out of vessels and is dependent on blood pressure driven by the heart and the elastic arteries. Osmotic pressure pulls fluid back into the vessels and is dependent on the number of particles dissolved in the plasma; most are proteins, so we often refer to this as oncotic pressure.
The balance of these opposing pressures, also called Starling forces, is essential for maintaining the proper fluid volumes and solute concentrations inside and outside the vasculature. Imbalance of these pressures can result in too much or too little fluid in the tissues. For example, accumulation of excess fluid in the interstitium results in a condition called edema. We should note that some interstitial fluid is also taken up by the lymphatic system. Lymphatic fluid (lymph) is returned to the central circulatory system by way of a channel called the thoracic duct. Blockage of lymph nodes by infection or surgery can also result in edema. Although you do not need to learn or memorize the Starling equation, which quantifies the net filtration rate between two fluid compartments, you should understand that the movement of solutes and fluid at the capillary level is governed by pressure differentials, just like the movement of carbon dioxide and oxygen in the lungs.
COAGULATION
We have now covered most of the functions of red blood cells and plasma. We’ve briefly touched on white blood cells, which we’ll explore more extensively in Chapter 8 of MCAT Biology Review. This leaves us with platelets, which protect the vascular system in the event of damage by forming a clot. Clots are composed of both coagulation factors (proteins) and platelets, and they prevent (or at least minimize) blood loss. When the endothelium of a blood vessel is damaged, it exposes the underlying connective tissue, which contains collagen and a protein called tissue factor. When platelets come into contact with exposed collagen, they sense this as evidence of injury. In response, they release their contents and begin to aggregate, or clump together. Simultaneously, coagulation factors, most of which are secreted by the liver, sense tissue factor and initiate a complex activation cascade. While the details of the coagulation cascade are beyond the scope of the MCAT, it is important to know that the endpoint of the cascade is the activation of prothrombin to form thrombin by thromboplastin. Thrombin can then convert fibrinogen intofibrin. Fibrin ultimately forms small fibers that aggregate into a woven structure, like a net, that captures red blood cells and other platelets, forming a stable clot over the area of damage, as shown in Figure 7.12. A clot that forms on a surface vessel that has been cut is called a scab.
REAL WORLD
Certain genetic diseases, such as hemophilia, cause malfunctions in the cascade of clotting reactions and increase the risk of life-threatening blood loss from even relatively minor injuries. Hemophilia A is the most common and, as a sex-linked trait, is far more common in males than females.
Ultimately, the clot will have to be broken down. This task is accomplished predominantly by plasmin, which is generated from plasminogen.
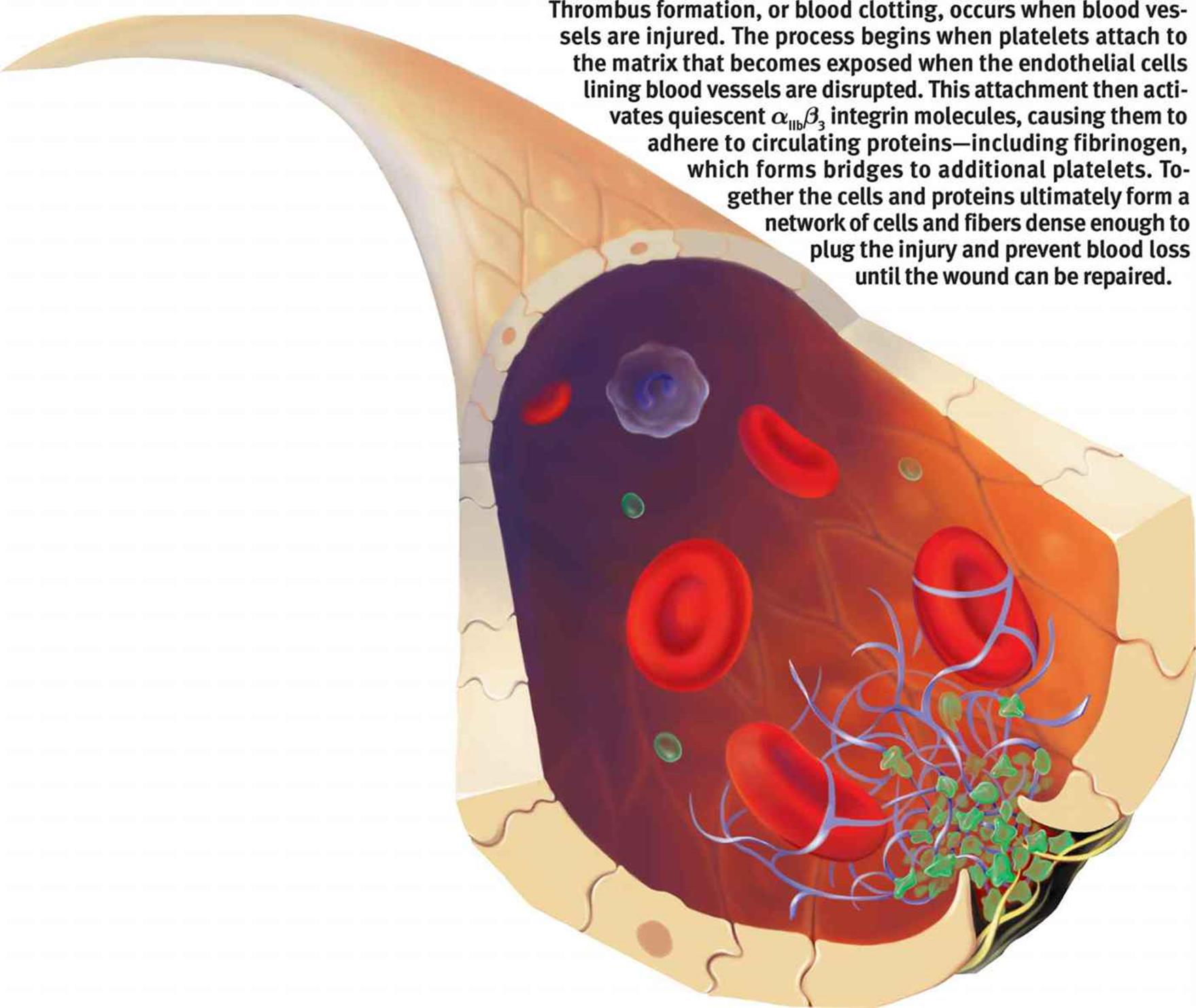 Figure 7.12. Thrombus (Clot) Formation
Figure 7.12. Thrombus (Clot) Formation
MCAT Concept Check 7.3:
Before you move on, assess your understanding of the material with these questions.
1. In bacterial sepsis (overwhelming bloodstream infection), a number of capillary beds throughout the body open simultaneously. What effect would this have on the blood pressure? Besides the risk of infection, why might sepsis be dangerous for the heart?
2. What is the chemical equation for the bicarbonate buffer system? What enzyme catalyzes this reaction?
3. Where should you look on the oxyhemoglobin dissociation curve to determine the amount of oxygen that has been delivered to tissues?
4. What can cause a right shift of the oxyhemoglobin dissociation curve? What can cause a left shift?
· Right shift:
· Left shift:
5. Exposure of which subendothelial compounds start the coagulation cascade? What protein helps stabilize the clot?
· Starts the cascade:
· Stabilizes the clot: