MCAT Biology Review
Chapter 1: The Cell
1.5 Viruses and Subviral Particles
At the beginning of this chapter, we discussed the cell theory and noted that viruses do not fit the definition of living things because they are acellular. Viruses may be as small as 20 nm or as large as 300 nm. For reference, prokaryotes are 1–10 µm, and eukaryotes are about ten times larger. Unlike eukaryotic cells, viruses lack organelles and a nucleus.
VIRAL STRUCTURE
Viruses are composed of genetic material, a protein coat, and sometimes an envelope containing lipids. The genetic information may be circular or linear, single- or double-stranded, and composed of either DNA or RNA. The protein coat is known as a capsid. If an envelope is present, it will surround the capsid and is composed of phospholipids and virus-specific proteins. The envelope is very sensitive to heat, detergents, and desiccation; thus, enveloped viruses are easier to kill. On the other hand, viruses that do not have an envelope are more resistant to sterilization and are likely to persist on surfaces for an extended period of time.
Because viruses cannot reproduce independently, they are considered obligate intracellular parasites. Viruses must express and replicate genetic information within a host cell because they lack ribosomes to carry out protein synthesis. After hijacking a cell’s machinery, a virus will replicate and produce viral progeny, called virions, which can be released to infect additional cells.
Bacteriophages are viruses that specifically target bacteria. They do not actually enter bacteria; rather, they simply inject their genetic material, leaving the remaining structures outside the infected cell. In addition to a capsid, bacteriophages contain a tail sheath and tail fibers, as shown in Figure 1.15. The tail sheath can act like a syringe, injecting genetic material into a bacterium. The tail fibers help the bacteriophage to recognize and connect to the correct host cell.
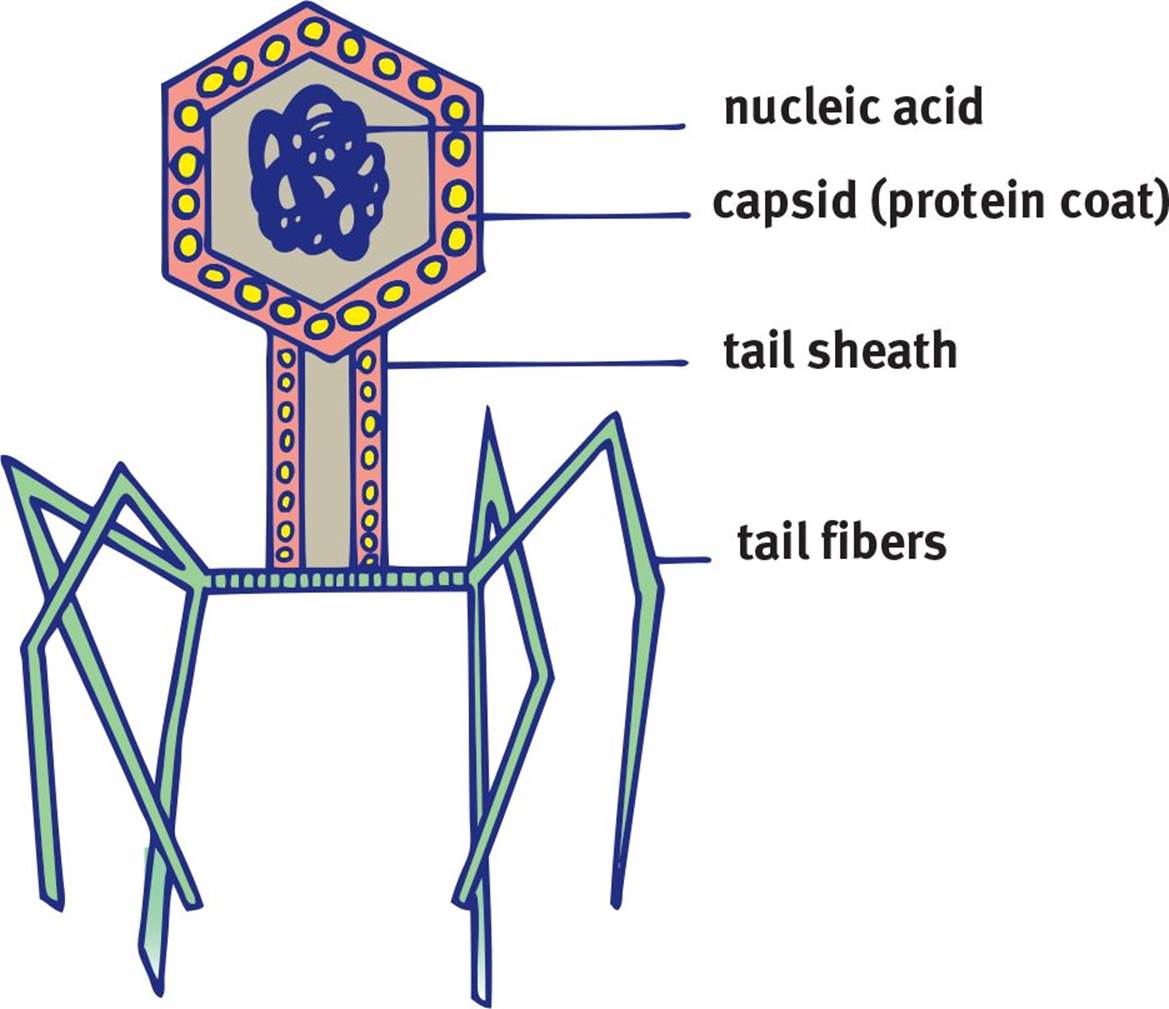 Figure 1.15. Structure of a Bacteriophage
Figure 1.15. Structure of a Bacteriophage
VIRAL GENOMES
Viral genomes come in a variety of shapes and sizes. Some are made of only a few genes, while others have several hundred. In addition, viral genomes may be made of either DNA or RNA, and may be single- or double-stranded in both cases.
Single-stranded RNA viruses may be positive sense or negative sense. Positive sense implies that the genome may be directly translated to functional proteins by the ribosomes of the host cell, just like mRNA. Negative-sense RNA viruses are a bit more complicated: these viruses require synthesis of an RNA strand complementary to the negative-sense RNA strand, which can then be used as a template for protein synthesis. Negative-sense RNA viruses must carry an RNA replicase in the virion to ensure that the complementary strand is synthesized.
Retroviruses are enveloped, single-stranded RNA viruses in the family Retroviridae; usually, the virion contains two identical RNA molecules. These viruses carry an enzyme known as reverse transcriptase, which synthesizes DNA from single-stranded RNA. The DNA then integrates into the host cell genome, where it is replicated and transcribed as if it were the host cell’s own DNA. This is a clever mechanism because the integration of the genetic material into the host cell genome allows for the cell to be infected indefinitely, and the only way to cure the infection is to kill the infected cell itself. The human immunodeficiency virus (HIV) is a retrovirus that utilizes this life cycle, which is one of the characteristics that make HIV so difficult to treat, as shown in Figure 1.16.
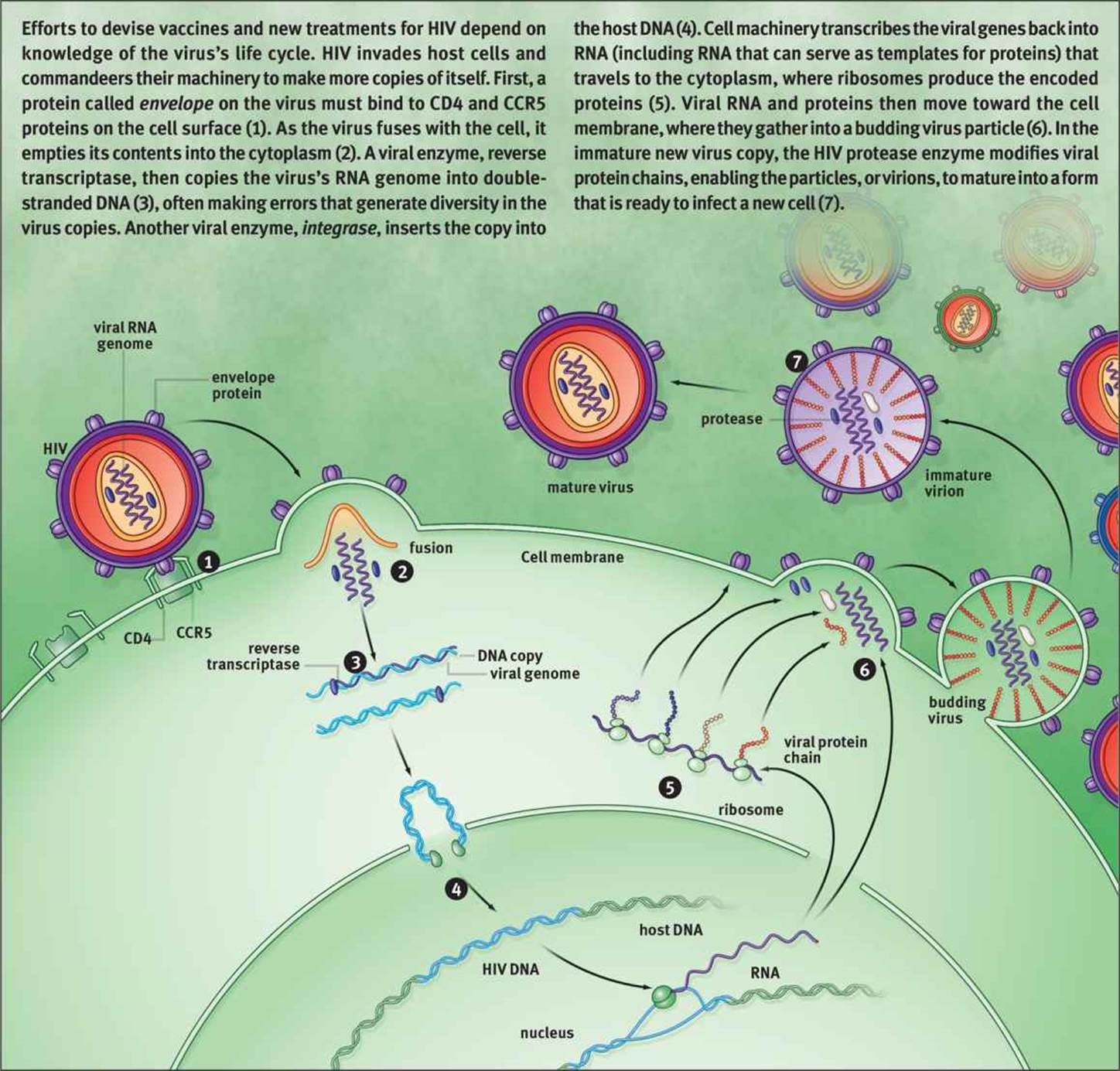 Figure 1.16. Life Cycle of the Human Immunodeficiency Virus (HIV)
Figure 1.16. Life Cycle of the Human Immunodeficiency Virus (HIV)
REAL WORLD
Both retroviruses and transduction are under investigation as methods of gene therapy. It is theorized that retroviruses and transduction methods can deliver functional versions of missing or altered genes, so that the correct proteins can be synthesized and certain disease states can be alleviated. Gene therapy is discussed in Chapter 6 of MCAT Biochemistry Review.
VIRAL LIFE CYCLE
As we have already discussed, viruses must infect a host cell and use the host cell’s machinery in order to reproduce. Here, we will discuss the viral life cycle in detail.
Infection
Viruses can only infect a specific set of cells. In order to infect a cell, the virus has to bind to specific receptors on the host cell. Without the proper receptors, a cell is essentially invisible to the virus. Once the virus binds the correct receptor, the virus and the cell are brought into close enough proximity to permit additional interactions. Enveloped viruses fuse with the plasma membrane of a cell, allowing the entry of the virion into the host cell. Sometimes a host cell may misinterpret the binding of a virus to the membrane as nutrients or other useful molecules and will actually bring the virus into the cytoplasm via endocytosis. As mentioned earlier, bacteriophages use tail fibers to anchor themselves to the cell membrane and then inject their viral genome into the host bacterium using the tail sheath. Some tail fibers even have enzymatic activity, allowing for both penetration of the cell wall and the formation of pores in the cell membrane.
REAL WORLD
HIV must bind to a receptor called CCR5 on white blood cells in order to infect them. Some people lack this receptor and are thus immune to HIV. There was recently a case in which an HIV-positive man with leukemia received a bone marrow transplant from a donor that lacked CCR5. Not only was his leukemia cured, but this transplant also resulted in remission of his HIV infection because the white blood cells from his newly acquired bone marrow were not susceptible to HIV infection.
Depending on the virus, different portions of the virion will be inserted into host cells. Enveloped viruses such as HIV fuse with the membrane and enter the cell intact, whereas bacteriophages only insert their genetic material, leaving their capsids outside the host cell.
Translation and Progeny Assembly
After infection, translation of viral genetic material must occur in order for the virus to reproduce. This requires translocation of the genetic material to the correct location in the cell. DNA viruses must go to the nucleus in order to be transcribed to mRNA. The mRNA then goes to the cytoplasm, where it is translated to proteins. Genetic material from positive-sense RNA viruses stays in the cytoplasm, where it is directly translated to protein by host cell ribosomes. Negative-sense RNA viruses require synthesis of a complementary RNA strand via RNA replicase, which can then be translated to form proteins. DNA formed through reverse transcription in retroviruses also travels to the nucleus, where it can be integrated into the host genome.
Using the ribosomes, tRNA, amino acids, and enzymes of the host cell, the viral RNA is translated into protein. Many of these proteins are structural capsid proteins and allow for creation of new virions in the cytoplasm in the host cell. Once the viral genome has been replicated, it can be packaged within the capsid. Note that the viral genome must be returned to its original form before packaging; for example, retroviruses must transcribe new copies of their single-stranded RNA from the DNA that entered the host genome. A single virus may create anywhere from hundreds to many thousands of new virions within a single host cell.
Progeny Release
Viral progeny may be released in multiple ways. First, the viral invasion may initiate cell death, which results in spilling of the viral progeny. Second, the host cell may lyse as a result of being filled with extremely large numbers of virions. Lysis is actually a disadvantage for the virus because the virus can no longer use the cell to carry out its life cycle. Finally, a virus can leave the cell by fusing with its plasma membrane as shown in Figure 1.17, a process known as extrusion. This process allows for survival of the host cell, and continued use of the host cell by the virus. A virus in this state is said to be in a productive cycle.
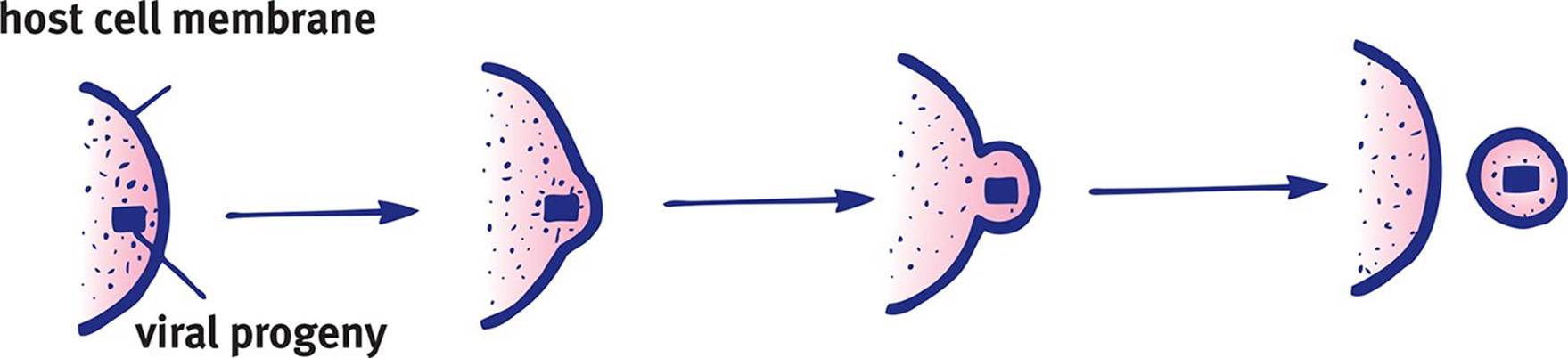 Figure 1.17. Viral Extrusion
Figure 1.17. Viral Extrusion
Lytic and Lysogenic Cycles
Depending on growth conditions and the specific virus, bacteriophages may enter a lytic or lysogenic life cycle. These two phases are similar to the lysis and productive cycle methods of progeny release discussed above.
During a lytic cycle, the bacteriophage makes maximal use of the cell’s machinery with little regard for the survival of the host cell. Once the host is swollen with new virions, the cell lyses, and other bacteria can be infected. Bacteria in the lytic phase are termed virulent.
In the event that the virus does not lyse the bacterium, it may integrate into the host genome as a provirus or prophage, beginning the lysogenic cycle. In this case, the virus will be replicated as the bacterium reproduces because it is now a part of the host’s genome. Although the virus may remain integrated into the host genome indefinitely, environmental factors (radiation, light, or chemicals) will cause the provirus to leave the genome and revert to a lytic cycle at some point. As mentioned earlier, trapping of segments of the bacterial genome can occur when the provirus leaves the genome, which allows transduction of genes from one bacterium to another. Although bacteriophages can kill a host bacterium, there may be some benefit to having them integrated in the lysogenic cycle. Infection with one strain of phage generally makes the bacterium less susceptible to superinfection (simultaneous infection) with other phages. Because the provirus is relatively innocuous, there may be some evolutionary advantage to this association. The lytic and lysogenic cycles are contrasted in Figure 1.18.
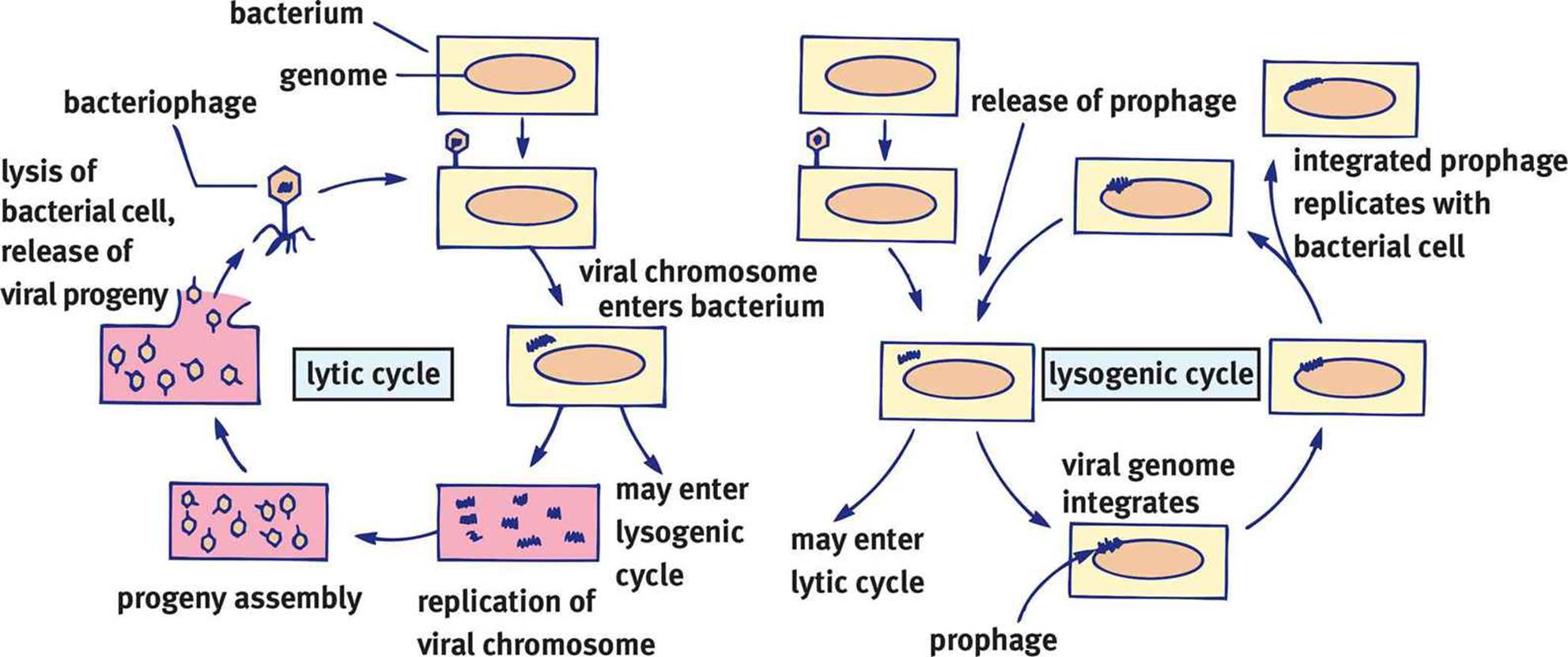 Figure 1.18. Lytic vs. Lysogenic Cycles of Bacteriophages
Figure 1.18. Lytic vs. Lysogenic Cycles of Bacteriophages
PRIONS AND VIROIDS
Prions and viroids are very small (subviral) particles that can cause disease under certain circumstances.
Prions
Prions are infectious proteins and are, thus, also nonliving things. Prions cause disease by triggering misfolding of other proteins, usually involving the conversion of a protein from an α-helical structure to a β-pleated sheet. This drastically reduces the solubility of the protein, as well as the ability of the cell to degrade the misfolded protein. Eventually, protein aggregates form, and function of the cell is reduced. Prions are known to cause bovine spongiform encephalopathy (mad cow disease), Creutzfeldt–Jakob disease, and familial fatal insomnia in humans.
Viroids
Viroids are small plant pathogens consisting of a very short circular single-stranded RNA. Viroids can bind to a large number of RNA sequences and will silence genes in the plant genome. This prevents synthesis of necessary proteins and can subsequently cause metabolic and structural derangements in the plant cell. Viroids are classically thought of as plant pathogens, but a few examples of human viroids do exist, including the hepatitis D virus (HDV). Alone, HDV is innocuous; however, when coinfected with hepatitis B virus (HBV), HDV is able to exert its silencing function on human hepatocytes.
MCAT Concept Check 1.5:
Before you move on, assess your understanding of the material with these questions.
1. Why are viruses considered obligate intracellular parasites?
2. Coronavirus, which causes the common cold, is described as an enveloped, single-stranded positive-sense RNA virus. What does this description indicate about the virus?
3. Briefly describe the pathway of retroviral nucleic acids from infection of a host cell to release of viral progeny:
4. What are the differences between the lytic and lysogenic cycles?
· Lytic cycle:
· Lysogenic cycle:
5. How do prions cause disease?