MCAT Biology Review
Chapter 12: Genetics and Evolution
12.4 Evolution
The development of evolutionary thought has a relatively short history; the first theories suggesting that new species may arise from older ones were proposed in the 19th century. Significant alterations to these initial theories have been made since then.
NATURAL SELECTION
Natural selection, sometimes phrased as survival of the fittest, is the theory that certain characteristics or traits possessed by individuals within a species may help those individuals to have greater reproductive success, thus passing on those traits to offspring. This theory was originally proposed by Charles Darwin in his 1859 publication On the Origin of Species. His theory was built on several basic tenets including:
· Organisms produce offspring, few of which survive to reproductive maturity.
· Chance variations within individuals in a population may be heritable. If these variations give an organism a slight survival advantage, the variation is termed favorable.
· Individuals with a greater preponderance of these favorable variations are more likely to survive to reproductive age and produce offspring; the overall result will be in an increase in these traits in future generations. This level of reproductive success is termed fitness, and an organism’s fitness is directly related to the relative genetic contribution of this individual to the next generation.
Darwin’s theory was ultimately proven to be correct in many ways, although not completely. In the 20th century, the field of modern genetics led to the development of the currently accepted theory.
KEY CONCEPT
Evolution is not equivalent to natural selection. The MCAT likes to test your ability to understand that natural selection is simply a mechanism for evolution. Natural selection is, however, equivalent to survival of the fittest.
Modern Theories
The modern synthesis model, sometimes called neo-Darwinism, adds knowledge of genetic inheritance and changes in the gene pool to Darwin’s original theory. Once scientists showed that inheritance occurs through the passing of genes from parent to child, and that genes ultimately change due to mutation or recombination, Darwin’s theory was updated to its current form: when mutation or recombination results in a change that is favorable to the organism’s reproductive success, that change is more likely to pass on to the next generation; the opposite is also true. This process is termed differential reproduction. Over time, those traits passed on by the more successful organisms will become pervasive in the gene pool. Because it is the gene pool that changes over time, it is important to note that it is populations that evolve, not individuals.
Also germane to the modernization of Darwin’s theory is a shift in scope to focus on inclusive fitness over the fitness of an individual organism. Inclusive fitness is a measure of an organism’s success in the population. This is based on the number of offspring, success in supporting offspring, and the ability of the offspring to then support others. Early descriptions of evolutionary success, like those of Darwin, were based solely on the number of viable offspring of an organism. However, contemporary theories take into account the benefits of certain behaviors on the population at large. For example, the existence of altruism could be supported by the observation that close relatives of an individual will share many of the same genes; thus, promoting the reproduction and survival of related or similar individuals can also lead to genetic success. Other species show examples of inclusive fitness by protecting the offspring of the group at large. By sacrificing themselves to protect the young, these organisms ensure the passing of genes to future generations. Inclusive fitness therefore promotes the idea that altruistic behavior can improve the fitness and success of a species as a whole.
One final theory to consider was proposed as a result of research into the fossil record. Upon examination, it was discovered that little evolution would occur within a lineage of related lifeforms for long periods of time, followed by a massive explosion of evolutionary change. Niles Eldridge and Stephen Jay Gould proposed the theory of punctuated equilibrium to explain this in 1972. In contrast to Darwin’s theory, punctuated equilibrium suggests that changes in some species occur in rapid bursts rather than evenly over time.
Modes of Natural Selection
Natural selection may occur as stabilizing selection, directional selection, or disruptive selection, as shown in Figure 12.11.
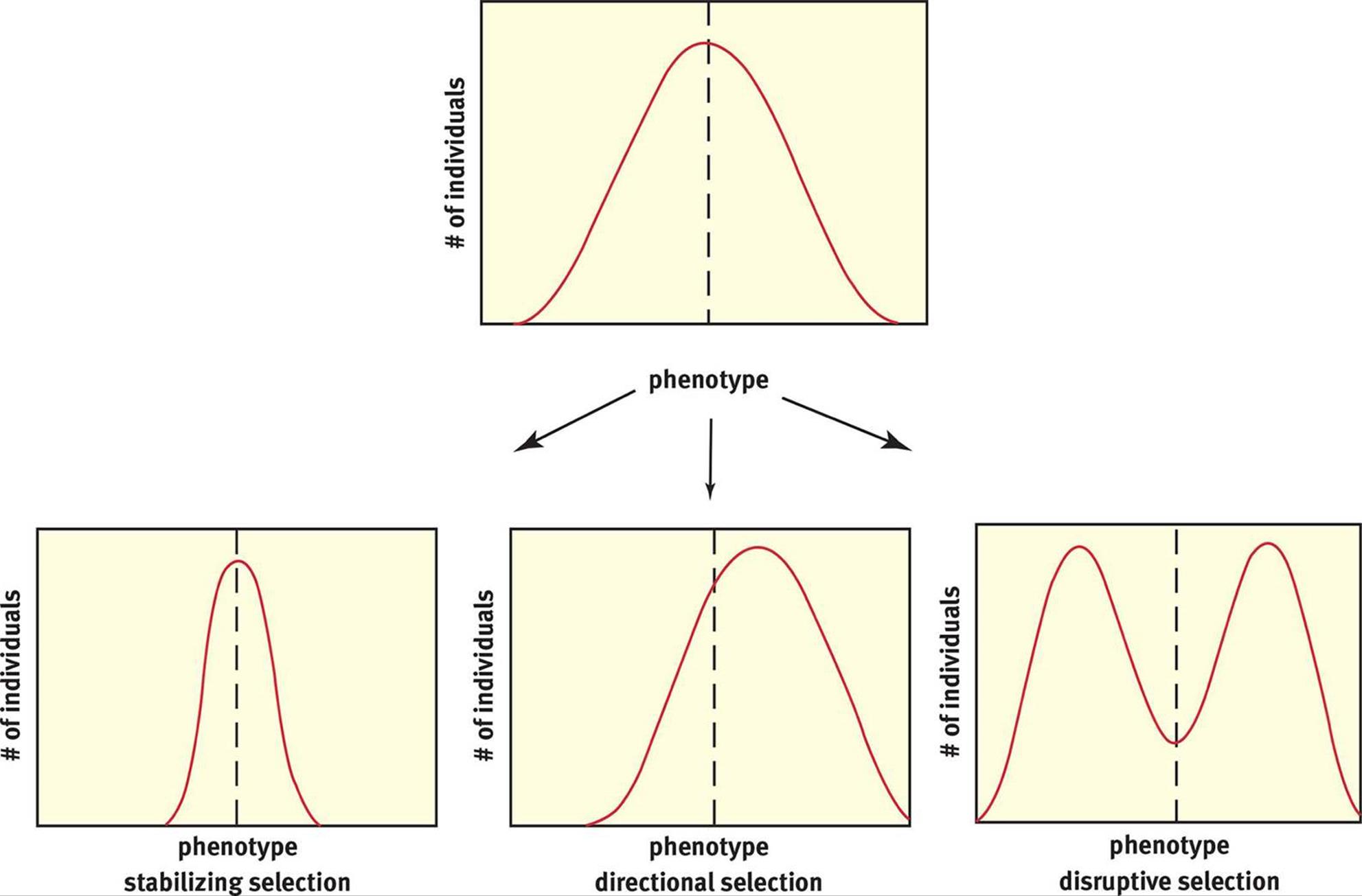 Figure 12.11. Modes of Natural Selection
Figure 12.11. Modes of Natural Selection
Stabilizing selection keeps phenotypes within a specific range by selecting against extremes. For instance, human birth weight is maintained within a narrow band by stabilizing selection. Fetuses that weigh too little may not be healthy enough to survive, and fetuses that weigh too much can experience trauma during delivery through the relatively narrow birth canal. In addition, the larger the fetus, the more resources it requires from the mother. For all of these reasons, it is advantageous to keep birth weights within a narrow range.
Adaptive pressure can lead to the emergence and dominance of an initially extreme phenotype through directional selection. For example, if we have a heterogeneous plate of bacteria, very few may have resistance to antibiotics. If the plate is then treated with ampicillin (an antibiotic), only those colonies that exhibit resistance to this antibiotic will survive. A new standard phenotype emerges as a result of differential survivorship. Natural selection is the history of differential survivorship over time. The emergence of mosquitoes resistant to dichlorodiphenyltrichloroethane(DDT), a type of pesticide, is attributed to directional selection.
In disruptive selection, two extreme phenotypes are selected over the norm. When Darwin studied finches on the Galapagos Islands, he noted that although there were many species, all the species arguably had a common ancestor, given their similar appearances. However, when he compared beak sizes, they were all either large or small, as shown in Figure 12.12. No animals exhibited the intermediate phenotype of medium-size beaks. Darwin hypothesized that the sizes of seeds on the island (the finches’ food) led to this effect. Seeds were either quite large or fairly small, requiring a large or small beak, respectively. Thus, if the original ancestor had a medium-size beak, over time, the animals with slightly larger or smaller beaks would be selected. Adaptive radiation is a related concept that describes the rapid rise of a number of different species from a common ancestor. The benefit of adaptive radiation is that it allows the various species to occupy different niches (specific ways of life, including habitat location and utilization of resources), decreasing competition for limited resources.
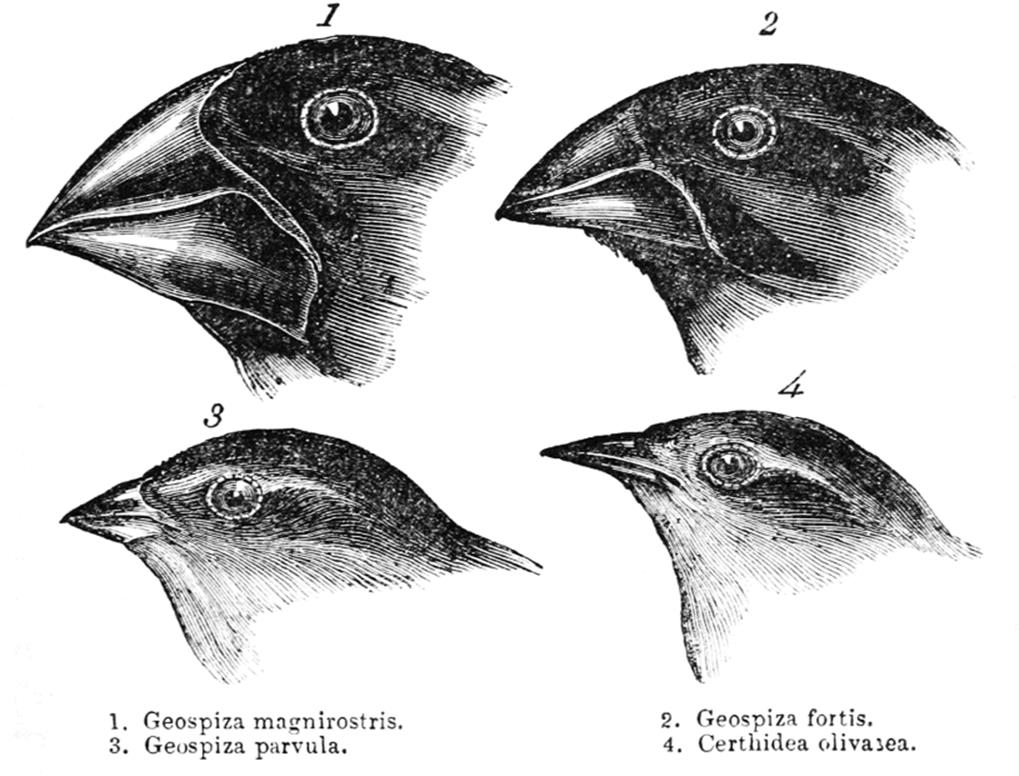 Figure 12.12. Darwin’s Finches Image drawn by Darwin in 1845.
Figure 12.12. Darwin’s Finches Image drawn by Darwin in 1845.
SPECIATION
A species is defined as the largest group of organisms capable of breeding to form fertile offspring; the formation of a new species through evolution is called speciation. If we took two populations from the same species and separated them geographically for a long period of time, different evolutionary pressures would lead to different adaptive selections. If enough time passed, the changes would be sufficient to lead to isolation, in which the progeny of these populations could no longer freely interbreed. We would now consider the two groups separate species. Reproductive isolation may occur either prezygotically or postzygotically. Prezygotic mechanisms prevent formation of the zygote completely; postzygotic mechanisms allow for gamete fusion but yield either nonviable or sterile offspring. Examples of prezygotic mechanisms include temporal isolation (breeding at different times), ecological isolation (living in different niches within the same territory), behavioral isolation (a lack of attraction between members of the two species due to differences in pheromones, courtship displays, and so on), reproductive isolation (incompatibility of reproductive anatomy), or gametic isolation (intercourse can occur, but fertilization cannot). Postzygotic mechanisms include hybrid inviability (formation of a zygote that cannot develop to term), hybrid sterility (forming hybrid offspring that cannot reproduce), and hybrid breakdown (forming first-generation hybrid offspring that are viable and fertile, but second-generation hybrid offspring that are inviable or infertile). As described earlier in this chapter, mules are an example of postzygotic hybrid sterility. Although a horse and donkey can produce a viable mule, the mule will be sterile and thus unable to contribute to a self-perpetuating mule lineage.
Patterns of Evolution
When we look at similarities between two species, we must be careful to determine whether those similarities are due to sharing a common ancestor or sharing a common environment with the same evolutionary pressures. When analyzing species this way, three patterns of evolution emerge: divergent evolution, parallel evolution, and convergent evolution, as shown in Figure 12.13.
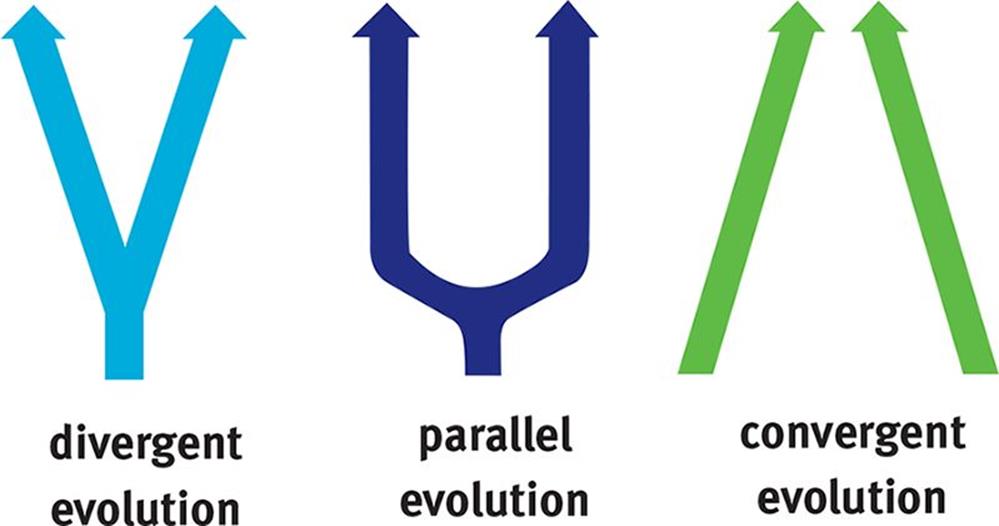 Figure 12.13. Patterns of Evolution
Figure 12.13. Patterns of Evolution
Divergent evolution refers to the independent development of dissimilar characteristics in two or more lineages sharing a common ancestor. For example, seals and cats are both mammals in the order Carnivora, yet they differ markedly in general appearance. These two species live in very different environments and adapted to different selection pressures while evolving.
Parallel evolution refers to the process whereby related species evolve in similar ways for a long period of time in response to analogous environmental selection pressures.
Convergent evolution refers to the independent development of similar characteristics in two or more lineages not sharing a recent common ancestor. For example, fish and dolphins have come to resemble one another physically, although they belong to different classes of vertebrates. They evolved certain similar features in adapting to the conditions of aquatic life.
MEASURING EVOLUTIONARY TIME
Evolution is slow process, featuring changes in the environment and subsequent changes in genotypes and phenotypes of a population over time. The rate of evolution is measured by the rate of change of a genotype over a period of time and is related to the severity of the evolutionary pressures put on the species. In other words, if a species is already perfectly suited to its habitat and there are no changes to the conditions in which it lives, the rate of evolution will be exceedingly slow—although there will still be some small base rate of genetic mutation. On the other hand, if an organism lives in a rapidly changing environment, the rate of evolution will be greater, as selection for and against certain traits will be actively occurring within that population.
By comparing DNA sequences between different species, scientists can quantify the degree of similarity between two organisms. For example, chimpanzees share over 95 percent of their genome with humans, whereas mice share only about 70 percent. As species become more taxonomically distant, the proportion of shared genome will decrease. Molecular evolutionists correlate the degree of genomic similarity with the amount of time since two species split off from the same common ancestor; the more similar the genomes, the more recently the two species separated from each other. This is sometimes called the molecular clock model.
MCAT Concept Check 12.4:
Before you move on, assess your understanding of the material with these questions.
1. What are the key tenets of each of the following theories of evolution?
· Natural selection:
· Modern synthesis model:
· Inclusive fitness:
· Punctuated equilibrium:
2. What are the three patterns of selection? What changes would each create to the population phenotype?
|
Pattern of Selection |
Change to Population Phenotype |
3. What are the three patterns of evolution between species? What is the outcome of each one?
|
Pattern of Evolution |
Outcome |
4. What is the biological definition of a species?