Lippincott’s Illustrated Reviews: Biochemistr, Sixth Edition (2014)
UNIT III: Lipid Metabolism
Chapter 15. Dietary Lipid Metabolism
I. OVERVIEW
Lipids are a heterogeneous group of water-insoluble (hydrophobic) organic molecules (Figure 15.1). Because of their insolubility in aqueous solutions, body lipids are generally found compartmentalized, as in the case of membrane-associated lipids or droplets of triacylglycerol in adipocytes, or transported in plasma in association with protein, as in lipoprotein particles (see p. 227), or on albumin. Lipids are a major source of energy for the body, and they also provide the hydrophobic barrier that permits partitioning of the aqueous contents of cells and subcellular structures. Lipids serve additional functions in the body (for example, some fat-soluble vitamins have regulatory or coenzyme functions, and the prostaglandins and steroid hormones play major roles in the control of the body’s homeostasis). Not surprisingly, deficiencies or imbalances of lipid metabolism can lead to some of the major clinical problems encountered by physicians, such as atherosclerosis, diabetes, and obesity.
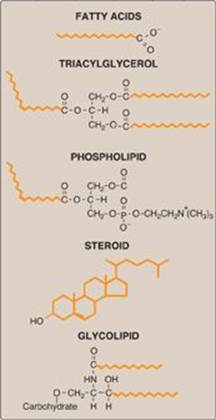
Figure 15.1 Structures of some common classes of lipids. Hydrophobic portions of the molecules are shown in orange.
II. DIGESTION, ABSORPTION, SECRETION, AND UTILIZATION OF DIETARY LIPIDS
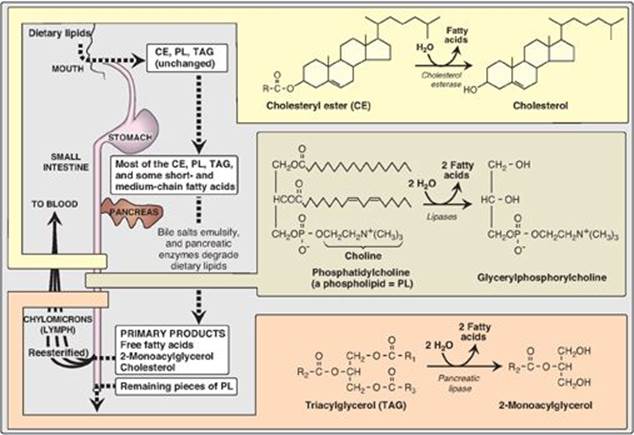
The average daily intake of lipids by U.S. adults is about 81 g, of which more than 90% is normally triacylglycerol ([TAG], formerly called triglyceride). The remainder of the dietary lipids consists primarily of cholesterol, cholesteryl esters, phospholipids, and unesterified (“free”) fatty acids. The digestion of dietary lipids is summarized in Figure 15.2.
A. Processing of dietary lipid in the stomach
The digestion of lipids begins in the stomach, catalyzed by a lipase (lingual lipase) that originates from glands at the back of the tongue. TAG molecules, particularly those containing fatty acids of short- or medium-chain length (fewer than 12 carbons such as are found in milk fat), are the primary target of this enzyme. These same TAGs are also degraded by a separate gastric lipase, secreted by the gastric mucosa. Both enzymes are relatively acid stable, with pH optimums of pH 4 to pH 6. These “acid lipases” play a particularly important role in lipid digestion in neonates, for whom milk fat is the primary source of calories. They also become important digestive enzymes in individuals with pancreatic insufficiency such as those with cystic fibrosis (CF). Lingual and gastric lipases aid these patients in degrading TAG molecules (especially those with short- to medium-chain length fatty acids) despite a near or complete absence of pancreatic lipase (see below).
Figure 15.2 Overview of lipid digestion.
1. Cystic fibrosis: CF is the most common lethal genetic disease in Caucasians of Northern European ancestry and has a prevalence of about 1:3,300 births in the United States. CF is an autosomal recessive disorder caused by mutations to the gene for the CF transmembrane conductance regulator (CFTR) protein that functions as a chloride channel on epithelium in the pancreas, lungs, testes, and sweat gands. Defective CFTR results in decreased secretion of chloride and increased uptake of sodium and water. In the pancreas, the depletion of water on the cell surface results in thickened secretions that clog the pancreatic ducts, preventing pancreatic enzymes from reaching the intestine, thereby leading to pancreatic insufficiency. Treatment includes replacement of these enzymes and supplementation with fat-soluble vitamins. [Note: CF also causes chronic lung infections with progressive pulmonary disease and male infertility.]
B. Emulsification of dietary lipid in the small intestine
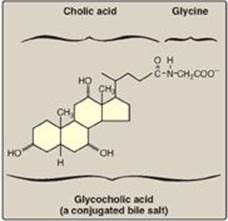
The critical process of emulsification of dietary lipids occurs in the duodenum. Emulsification increases the surface area of the hydrophobic lipid droplets so that the digestive enzymes, which work at the interface of the droplet and the surrounding aqueous solution, can act effectively. Emulsification is accomplished by two complementary mechanisms, namely, use of the detergent properties of the conjugated bile salts and mechanical mixing due to peristalsis. Bile salts, made in the liver and stored in the gallbladder, are amphipathic derivatives of cholesterol (see p. 224). Conjugated bile salts consist of a hydroxylated sterol ring structure with a side chain to which a molecule of glycine or taurine is covalently attached by an amide linkage (Figure 15.3). These emulsifying agents interact with the dietary lipid particles and the aqueous duodenal contents, thereby stabilizing the particles as they become smaller from peristalsis and preventing them from coalescing. A more complete discussion of bile salt metabolism is given on p. 225.
Figure 15.3 Structure of glycocholic acid.
C. Degradation of dietary lipids by pancreatic enzymes
The dietary TAG, cholesteryl esters, and phospholipids are enzymically degraded (“digested”) by pancreatic enzymes, whose secretion is hormonally controlled.
1. Triacylglycerol degradation: TAG molecules are too large to be taken up efficiently by the mucosal cells of the intestinal villi. They are, therefore, acted upon by an esterase, pancreatic lipase, which preferentially removes the fatty acids at carbons 1 and 3. The primary products of hydrolysis are, thus, a mixture of 2-monoacylglycerol and free fatty acids (see Figure 15.2). [Note: This enzyme is found in high concentrations in pancreatic secretions (2%–3% of the total protein present), and it is highly efficient catalytically, thus insuring that only severe pancreatic deficiency, such as that seen in CF, results in significant malabsorption of fat.] A second protein, colipase, also secreted by the pancreas, binds the lipase at a ratio of 1:1 and anchors it at the lipid–aqueous interface. Colipase restores activity to lipase in the presence of inhibitory substances like bile salts that bind the micelles. [Note: Colipase is secreted as the zymogen, procolipase, which is activated in the intestine by trypsin.] Orlistat, an antiobesity drug, inhibits gastric and pancreatic lipases, thereby decreasing fat absorption, resulting in weight loss.
2. Cholesteryl ester degradation: Most dietary cholesterol is present in the free (nonesterified) form, with 10%–15% present in the esterified form. Cholesteryl esters are hydrolyzed by pancreatic cholesteryl ester hydrolase(cholesterol esterase), which produces cholesterol plus free fatty acids (see Figure 15.2). Activity of this enzyme is greatly increased in the presence of bile salts.
3. Phospholipid degradation: Pancreatic juice is rich in the proenzyme of phospholipase A2 that, like procolipase, is activated by trypsin and, like cholesteryl ester hydrolase, requires bile salts for optimum activity. Phospholipase A2 removes one fatty acid from carbon 2 of a phospholipid, leaving a lysophospholipid. For example, phosphatidylcholine (the predominant phospholipid of digestion) becomes lysophosphatidylcholine. The remaining fatty acid at carbon 1 can be removed by lysophospholipase, leaving a glycerylphosphoryl base (for example, glycerylphosphorylcholine, see Figure 15.2) that may be excreted in the feces, further degraded, or absorbed.
Figure 15.4 Hormonal control of lipid digestion in the small intestine.
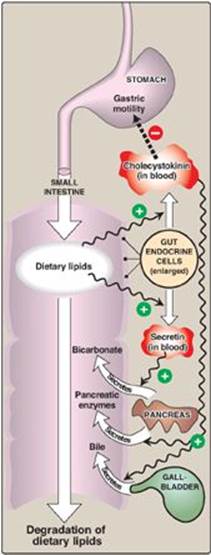
4. Control of lipid digestion: Pancreatic secretion of the hydrolytic enzymes that degrade dietary lipids in the small intestine is hormonally controlled (Figure 15.4). Cells in the mucosa of the lower duodenum and jejunum produce a small peptide hormone, cholecystokinin (CCK), in response to the presence of lipids and partially digested proteins entering these regions of the upper small intestine. CCK acts on the gallbladder (causing it to contract and release bile, a mixture of bile salts, phospholipids, and free cholesterol) and on the exocrine cells of the pancreas (causing them to release digestive enzymes). It also decreases gastric motility, resulting in a slower release of gastric contents into the small intestine (see p. 353). Other intestinal cells produce another small peptide hormone, secretin, in response to the low pH of the chyme entering the intestine. Secretin causes the pancreas to release a solution rich in bicarbonate that helps neutralize the pH of the intestinal contents, bringing them to the appropriate pH for digestive activity by pancreatic enzymes.
D. Absorption of lipids by intestinal mucosal cells, or enterocytes
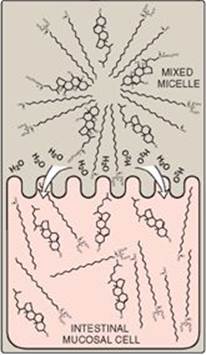
Free fatty acids, free cholesterol, and 2-monoacylglycerol are the primary products of lipid digestion in the jejunum. These, plus bile salts and fat-soluble vitamins (A, D, E, and K), form mixed micelles (that is, disc-shaped clusters of a mixture of amphipathic lipids that coalesce with their hydrophobic groups on the inside and their hydrophilic groups on the outside). Mixed micelles are, therefore, soluble in the aqueous environment of the intestinal lumen (Figure 15.5). These particles approach the primary site of lipid absorption, the brush border membrane of the enterocytes (mucosal cell). This membrane is separated from the liquid contents of the intestinal lumen by an unstirred water layer that mixes poorly with the bulk fluid. The hydrophilic surface of the micelles facilitates the transport of the hydrophobic lipids through the unstirred water layer to the brush border membrane where they are absorbed. Bile salts are absorbed in the terminal ileum, with less than 5% being lost in the feces. [Note: Relative to other dietary lipids, cholesterol is only poorly absorbed by the enterocytes. Drug therapy (for example, with ezetimibe) can further reduce cholesterol absorption in the small intestine.] Short- and medium-chain length fatty acids are water soluble and, thus, do not require the assistance of mixed micelles for absorption by the intestinal mucosa.
E. Resynthesis of triacylglycerols and cholesteryl esters
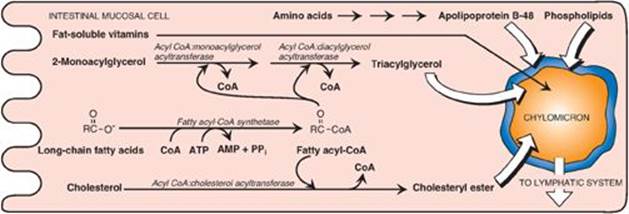
The mixture of lipids absorbed by the enterocytes migrates to the endoplasmic reticulum where biosynthesis of complex lipids takes place. The long-chain length fatty acids are first converted into their activated form by fatty acyl-coenzyme A (CoA) synthetase (thiokinase) as shown in Figure 15.6. Using the fatty acyl CoA derivatives, the 2-monoacylglycerols absorbed by the enterocytes are converted to TAGs through sequential reacylations by two acyltransferases, acyl CoA:monoacylglycerol acyltransferase and acyl CoA:diacylglycerol acyltransferase. Lysophospholipids are reacylated to form phospholipids by a family of acyltransferases, and cholesterol is esterified with a fatty acid primarily by acyl CoA:cholesterol acyltransferase (see p. 232). [Note: Virtually all long-chain fatty acids entering the enterocytes are used in this fashion to form TAGs, phospholipids, and cholesteryl esters. Short- and medium-chain length fatty acids are not converted to their CoA derivatives and are not reesterified to 2-monoacylglycerol. Instead, they are released into the portal circulation, where they are carried by serum albumin to the liver.]
F. Lipid malabsorption
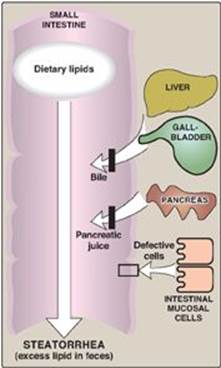
Lipid malabsorption, resulting in increased lipid (including the fat-soluble vitamins and essential fatty acids, see p. 182) in the feces, a condition known as steatorrhea, can be caused by disturbances in lipid digestion and/or absorption (Figure 15.7). Such disturbances can result from several conditions, including CF (causing poor digestion) and short bowel syndrome (causing decreased absorption).
The ability of short- and medium-chain length fatty acids to be taken up by enterocytes without the aid of mixed micelles has made them important in dietary therapy for individuals with malabsorption disorders.
Figure 15.5 Absorption of lipids contained in a mixed micelle by an intestinal mucosal cell. [Note: The micelle itself is not taken up.]
G. Secretion of lipids from enterocytes
The newly resynthesized TAGs and cholesteryl esters are very hydrophobic and aggregate in an aqueous environment. It is, therefore, necessary that they be packaged as particles of lipid droplets surrounded by a thin layer composed of phospholipids, unesterified cholesterol, and a molecule of the protein apolipoprotein B-48 (see p. 228). This layer stabilizes the particle and increases its solubility, thereby preventing multiple particles from coalescing. [Note: Microsomal triglyceride transfer protein is essential for the assembly of these (and other) TAG-rich apolipoprotein B–containing particles in the endoplasmic reticulum.] The lipoprotein particles are released by exocytosis from enterocytes into the lacteals (lymphatic vessels originating in the villi of the small intestine). The presence of these particles in the lymph after a lipid-rich meal gives it a milky appearance. This lymph is called chyle (as opposed to chyme, the name given to the semifluid mass of partially digested food that passes from the stomach to the duodenum), and the particles are named chylomicrons. Chylomicrons follow the lymphatic system to the thoracic duct and are then conveyed to the left subclavian vein, where they enter the blood. The steps in the production of chylomicrons are summarized in Figure 15.6. [Note: Once released into blood, chylomicrons pick up apolipoproteins E and C-II.] (For a more detailed description of chylomicron structure and metabolism, see p. 228.)
Figure 15.6 Assembly and secretion of chylomicrons by intestinal mucosal cells. [Note: Short- and medium-chain length fatty acids do not require incorporation into micelles or chylomicrons and directly enter into the blood.] CoA = coenzyme A; AMP = adenosine monophosphate; PPi = pyrophosphate.
H. Use of dietary lipids by the tissues
TAG contained in chylomicrons is broken down primarily in the capillaries of skeletal and cardiac muscle and adipose tissues. TAG in chylomicrons is degraded to free fatty acids and glycerol by lipoprotein lipase (LPL). This enzyme is synthesized primarily by adipocytes and muscle cells. It is secreted and becomes associated with the luminal surface of endothelial cells in the capillary beds of the peripheral tissues. [Note: Familial LPL deficiency (type I hyperlipoproteinemia) is a rare, autosomal recessive disorder caused by a deficiency of LPL or its coenzyme apolipoprotein C-II (see p. 228). The result is fasting chylomicronemia and hypertriacylglycerolemia.]
1. Fate of free fatty acids: The free fatty acids derived from the hydrolysis of TAG may either directly enter adjacent muscle cells or adipocytes or be transported in the blood in association with serum albumin until they are taken up by cells. [Note: Serum albumin is a large glycoprotein secreted by the liver. It transports a number of primarily hydrophobic compounds in the circulation, including free fatty acids and some drugs.] Most cells can oxidize fatty acids to produce energy (see p. 190). Adipocytes can also reesterify free fatty acids to produce TAG molecules, which are stored until the fatty acids are needed by the body (see p. 188).
2. Fate of glycerol: Glycerol released from TAG is taken up from the blood and phosphorylated by hepatic glycerol kinase to produce glycerol 3-phosphate, which can enter either glycolysis or gluconeogenesis by oxidation to dihydroxyacetone phosphate (see p. 190).
Figure 15.7 Possible causes of steatorrhea.
3. Fate of the remaining chylomicron components: After most of the TAG has been removed, the chylomicron remnants (which contain cholesteryl esters, phospholipids, apolipoproteins, fat-soluble vitamins, and a small amount of TAG) bind to receptors on the liver (apolipoprotein E is the ligand; see p. 230) and are endocytosed. The intracellular remnants are hydrolyzed to their component parts. Cholesterol and the nitrogenous bases of phospholipids (for example, choline) can be recycled by the body. [Note: If removal of remnants by the liver is decreased due to impaired binding to their receptor, they accumulate in the plasma. This is seen in the rare type III hyperlipoproteinemia (also called familial dysbetalipoproteinemia, see p. 231).]
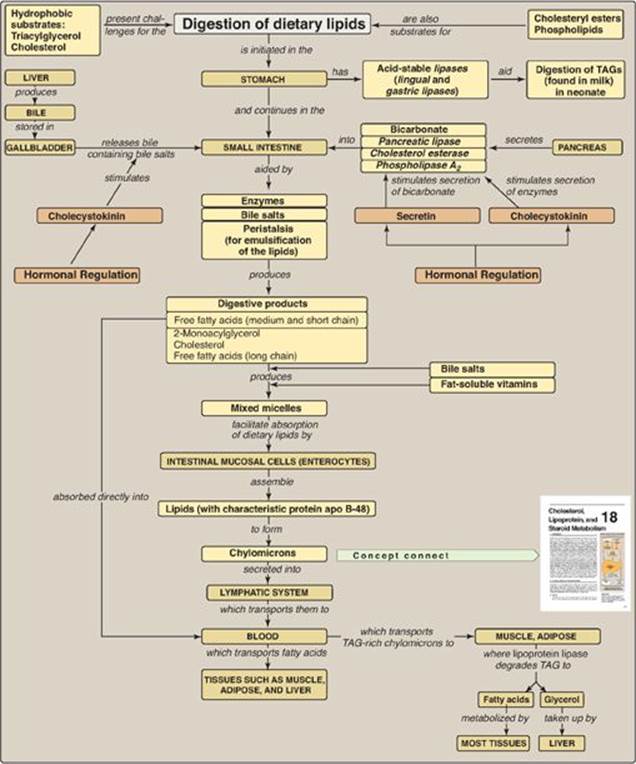
Figure 15.8 Key concept map for metabolism of dietary lipids. apo = apolipoprotein; TAGs = triacylglycerols.
III. CHAPTER SUMMARY
The digestion of dietary lipids begins in the stomach and continues in the small intestine (Figure 15.8). The hydrophobic nature of lipids requires that the dietary lipids, particularly those that contain long-chain length fatty acids (LCFAs), be emulsified for efficient degradation. Triacylglycerols (TAG) obtained from milk contain short- to medium-chain length fatty acids that can be degraded in the stomach by the acid lipases (lingual lipaseand gastric lipase). Cholesteryl esters (CEs), phospholipids (PLs), and TAG containing LCFAs are degraded in the small intestine by enzymes secreted by the pancreas. The most important of these enzymes are pancreatic lipase, phospholipase A2, and cholesterol esterase. The dietary lipids are emulsified in the small intestine using peristaltic action and bile salts, which serve as detergents. The primary products resulting from enzymatic degradation of dietary lipid are 2-monoacylglycerol, unesterified cholesterol, and free fatty acids. These compounds, plus the fat-soluble vitamins, form mixed micelles that facilitate the absorption of dietary lipids by intestinal mucosal cells (enterocytes). These cells resynthesize TAG, CE, and PL using LCFAs and also synthesize protein (apolipoprotein B-48), all of which are then assembled with the fat-soluble vitamins into lipoprotein particles called chylomicrons. Short- and medium-chain fatty acids enter blood directly. Chylomicrons are released into the lymph, which carries them to the blood, where their lipid core is degraded by lipoprotein lipase(with apolipoprotein C-II as the coenzyme) in muscle and adipose tissues. Thus, dietary lipids are made available to the peripheral tissues. Problems with fat absorption cause steatorrhea. A deficiency in the ability to degrade chylomicron components, or remove their remnants after TAG has been removed, results in accumulation of these particles in blood.
Study Questions
Choose the ONE best answer.
15.1 Which one of the following statements about the digestion of lipids is correct?
A. Large lipid droplets are emulsified (have their surface area increased) in the mouth through the act of chewing (mastication).
B. The enzyme colipase facilitates the binding of bile salts to mixed micelles, maximizing the activity of pancreatic lipase.
C. The peptide hormone secretin causes the gallbladder to contract and release bile.
D. Patients with cystic fibrosis have difficulties with digestion because their thickened pancreatic secretions are less able to reach the small intestine, the primary site of lipid digestion.
E. Formation of triacylglycerol-rich chylomicrons is independent of protein synthesis in the intestinal mucosa.
Correct answer = D. Patients with cystic fibrosis, a genetic disease due to a deficiency of a functional chloride transporter, have thickened secretions that impede the flow of pancreatic enzymes into the duodenum. Emulsification occurs through peristalsis, which provides mechanical mixing, and bile salts that function as detergents. Colipase restores activity to pancreatic lipase in the presence of inhibitory bile salts that bind the micelles. Cholecystokinin is the hormone that causes contraction of the gallbladder and release of stored bile, and secretin causes release of bicarbonate. Chylomicron formation requires synthesis of apolipoprotein B-48.
15.2 Which one of the following statements about the absorption of lipids from the intestine is correct?
A. Dietary triacylglycerol must be completely hydrolyzed to free fatty acids and glycerol before absorption.
B. The triacylglycerol carried by chylomicrons is degraded by lipoprotein lipase to fatty acids that are taken up by muscle and adipose tissues and glycerol that is taken up by the liver.
C. Fatty acids that contain fewer than 12 carbon atoms are absorbed and enter the circulation primarily via the lymphatic system.
D. Deficiencies in the ability to absorb fat result in excessive amounts of chylomicrons in the blood.
Correct answer = B. The triacylglycerols (TAGs) in chylomicrons are degraded to fatty acids and glycerol by lipoprotein lipase on the endothelial surface of capillaries in muscle and adipose, thus providing a source of fatty acids to these tissues for degradation or storage and providing glycerol for hepatic metabolism. In the duodenum, TAG are degraded to one 2-monoacyl-glycerol + two free fatty acids that get absorbed. Medium- and short-chain fatty acids enter directly into blood (not lymph), and they neither require micelles nor get packaged into chylomicrons. Because chylomicrons contain dietary lipids that were digested and absorbed, a defect in fat absorption would result in decreased production of chylomicrons.