Lippincott’s Illustrated Reviews: Biochemistr, Sixth Edition (2014)
UNIT III: Lipid Metabolism
Chapter 18. Cholesterol, Lipoprotein, and Steroid Metabolism
I. OVERVIEW
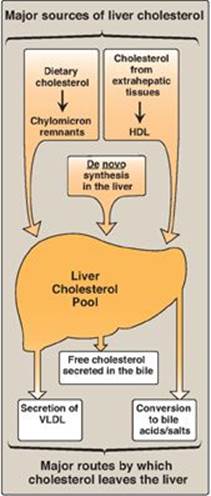
Cholesterol, the characteristic steroid alcohol of animal tissues, performs a number of essential functions in the body. For example, cholesterol is a structural component of all cell membranes, modulating their fluidity, and, in specialized tissues, cholesterol is a precursor of bile acids, steroid hormones, and vitamin D. It is, therefore, critically important that the cells of the body be assured an appropriate supply of cholesterol. To meet this need, a complex series of transport, biosynthetic, and regulatory mechanisms has evolved. The liver plays a central role in the regulation of the body’s cholesterol homeostasis. For example, cholesterol enters the liver’s cholesterol pool from a number of sources including dietary cholesterol as well as that synthesized de novo by extrahepatic tissues and by the liver itself. Cholesterol is eliminated from the liver as unmodified cholesterol in the bile, or it can be converted to bile salts that are secreted into the intestinal lumen. It can also serve as a component of plasma lipoproteins that carry lipids to the peripheral tissues. In humans, the balance between cholesterol influx and efflux is not precise, resulting in a gradual deposition of cholesterol in the tissues, particularly in the endothelial linings of blood vessels. This is a potentially life-threatening occurrence when the lipid deposition leads to plaque formation, causing the narrowing of blood vessels (atherosclerosis) and increased risk of cardio-, cerebro-, and peripheral vascular disease. Figure 18.1 summarizes the major sources of liver cholesterol and the routes by which cholesterol leaves the liver.
Figure 18.1 Sources of liver cholesterol (influx) and routes by which cholesterol leaves the liver (efflux). HDL = highdensity lipoprotein; VLDL = very-lowdensity lipoprotein.
II. STRUCTURE OF CHOLESTEROL
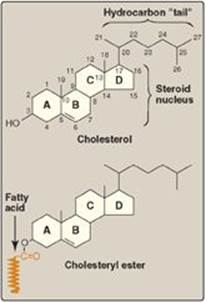
Cholesterol is a very hydrophobic compound. It consists of four fused hydrocarbon rings (A–D) called the “steroid nucleus,” and it has an eight-carbon, branched hydrocarbon chain attached to carbon 17 of the D ring. Ring A has a hydroxyl group at carbon 3, and ring B has a double bond between carbon 5 and carbon 6 (Figure 18.2).
A. Sterols
Steroids with eight to ten carbon atoms in the side chain at carbon 17 and a hydroxyl group at carbon 3 are classified as sterols. Cholesterol is the major sterol in animal tissues. It arises from de novo synthesis and absorption of dietary cholesterol. [Note: Intestinal uptake of cholesterol is mediated, at least in part, by the protein Niemann-Pick C1-like 1 protein (NPC1-L1), the target of the drug ezetimibe that reduces absorption of dietary cholesterol (see p. 176). Plant sterols (phytosterols), such as β-sitosterol, are poorly absorbed by humans (5% absorbed as compared to 40% for cholesterol). After entering the enterocytes, they are actively transported back into the intestinal lumen. Defects in the transporter result in the rare condition of sitosterolemia. Because some cholesterol is transported back as well, plant sterols reduce the absorption of dietary cholesterol. Daily ingestion of plant sterol esters supplied, for example, in spreads or juices, is one of a number of dietary strategies to reduce plasma cholesterol levels (see p. 363).]
Figure 18.2 Structure of cholesterol and its ester.
B. Cholesteryl esters
Most plasma cholesterol is in an esterified form (with a fatty acid attached at carbon 3, as shown in Figure 18.2), which makes the structure even more hydrophobic than free (unesterified) cholesterol. Cholesteryl esters are not found in membranes and are normally present only in low levels in most cells. Because of their hydrophobicity, cholesterol and its esters must be transported in association with protein as a component of a lipoprotein particle (see p. 227) or be solubilized by phospholipids and bile salts in the bile (see p. 226).
III. SYNTHESIS OF CHOLESTEROL
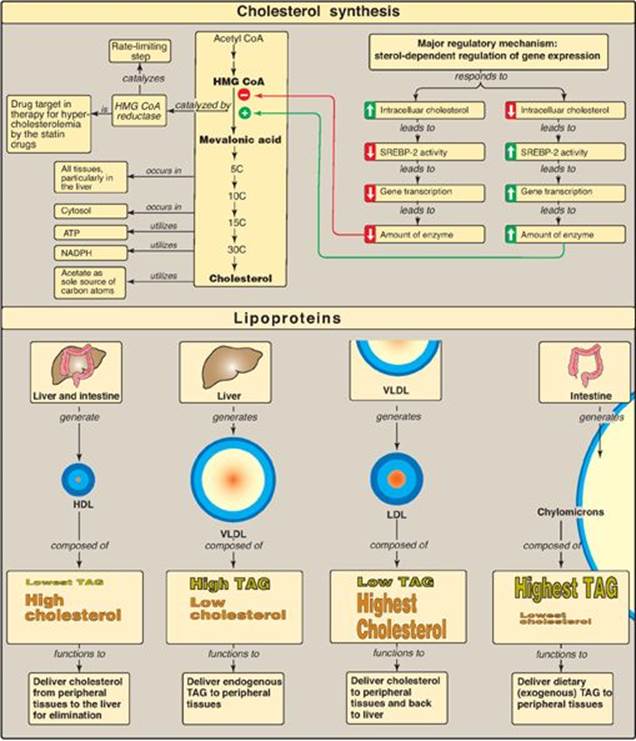
Cholesterol is synthesized by virtually all tissues in humans, although liver, intestine, adrenal cortex, and reproductive tissues, including ovaries, testes, and placenta, make the largest contributions to the body’s cholesterol pool. As with fatty acids, all the carbon atoms in cholesterol are provided by acetyl coenzyme A (CoA), and nicotinamide adenine dinucleotide phosphate (NADPH) provides the reducing equivalents. The pathway is endergonic, being driven by hydrolysis of the high-energy thioester bond of acetyl CoA and the terminal phosphate bond of adenosine triphosphate (ATP). Synthesis requires enzymes in both the cytosol and the membrane of the smooth endoplasmic reticulum (ER). The pathway is responsive to changes in cholesterol concentration, and regulatory mechanisms exist to balance the rate of cholesterol synthesis within the body against the rate of cholesterol excretion. An imbalance in this regulation can lead to an elevation in circulating levels of plasma cholesterol, with the potential for vascular disease.
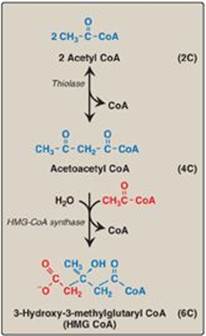
Figure 18.3 Synthesis of HMG CoA. CoA = coenzyme A.
A. Synthesis of 3-hydroxy-3-methylglutaryl coenzyme A
The first two reactions in the cholesterol synthetic pathway are similar to those in the pathway that produces ketone bodies (see Figure 16.22, p. 196). They result in the production of 3-hydroxy-3-methylglutaryl CoA ([HMG CoA] Figure 18.3). First, two acetyl CoA molecules condense to form acetoacetyl CoA. Next, a third molecule of acetyl CoA is added by HMG CoA synthase, producing HMG CoA, a six-carbon compound. [Note: Liver parenchymal cells contain two isoenzymes of the synthase. The cytosolic enzyme participates in cholesterol synthesis, whereas the mitochondrial enzyme functions in the pathway for ketone body synthesis.]
B. Synthesis of mevalonate
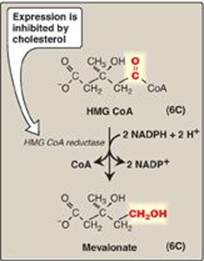
The next step, the reduction of HMG CoA to mevalonate, is catalyzed by HMG CoA reductase and is the rate-limiting and key regulated step in cholesterol synthesis. It occurs in the cytosol, uses two molecules of NADPH as the reducing agent, and releases CoA, making the reaction irreversible (Figure 18.4). [Note: HMG CoA reductase is an integral membrane protein of the ER, with its catalytic domain projecting into the cytosol.] Regulation of reductase activity is discussed below.
Figure 18.4 Synthesis of mevalonate. HMG CoA = hydroxymethylglutaryl coenzyme A; NADP(H) = nicotinamide adenine dinucleotide phosphate.
C. Synthesis of cholesterol
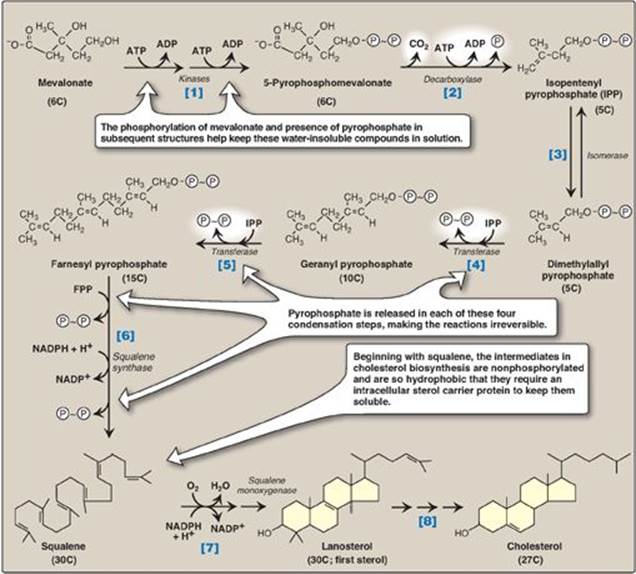
The reactions and enzymes involved in the synthesis of cholesterol from mevalonate are illustrated in Figure 18.5. [Note: The numbers shown in brackets below correspond to numbered reactions shown in this figure.]
[1] Mevalonate is converted to 5-pyrophosphomevalonate in two steps, each of which transfers a phosphate group from ATP.
[2] A five-carbon isoprene unit, isopentenyl pyrophosphate (IPP), is formed by the decarboxylation of 5-pyrophosphomevalonate. The reaction requires ATP. [Note: IPP is the precursor of a family of molecules with diverse functions, the isoprenoids. Cholesterol is a sterol isoprenoid. Nonsterol isoprenoids include dolichol (see p. 167) and ubiquinone, or coenzyme Q (see p. 75).]
[3] IPP is isomerized to 3,3-dimethylallyl pyrophosphate (DPP).
[4] IPP and DPP condense to form ten-carbon geranyl pyrophosphate (GPP).
[5] A second molecule of IPP then condenses with GPP to form 15-carbon farnesyl pyrophosphate (FPP). [Note: Covalent attachment of farnesyl to proteins, a process known as “prenylation,” is one mechanism for anchoring proteins (such as ras) to plasma membranes.]
[6] Two molecules of FPP combine, releasing pyrophosphate, and are reduced, forming the 30-carbon compound squalene. [Note: Squalene is formed from six isoprenoid units. Because three ATP are hydrolyzed per mevalonate residue converted to IPP, a total of 18 ATP are required to make the polyisoprenoid squalene.]
[7] Squalene is converted to the sterol lanosterol by a sequence of reactions catalyzed by ER-associated enzymes that use molecular oxygen and NADPH. The hydroxylation of linear squalene triggers the cyclization of the structure to lanosterol.
[8] The conversion of lanosterol to cholesterol is a multistep, ER-associated process involving shortening of the side-chain, oxidative removal of methyl groups, reduction of double bonds, and migration of a double bond. Smith-Lemli-Opitz syndrome (SLOS), an autosomal-recessive disorder of cholesterol biosynthesis, is caused by a partial deficiency in 7-dehydrocholesterol-7-reductase, the enzyme that reduces the double bond in 7-dehydrocholesterol (7-DHC), thereby converting it to cholesterol. SLOS is one of several multisystem, embryonic malformation syndromes associated with impaired cholesterol synthesis. [Note: 7-DHC is converted to vitamin D3 in the skin (see p. 386).]
Figure 18.5 Synthesis of cholesterol from mevalonate. ADP = adenosine diphosphate; ![]() = phosphate;
= phosphate; ![]() ~
~![]() = pyrophosphate; NADP(H) = nicotinamide adenine dinucleotide phosphate.
= pyrophosphate; NADP(H) = nicotinamide adenine dinucleotide phosphate.
D. Regulation of cholesterol synthesis
HMG CoA reductase is the major control point for cholesterol biosynthesis and is subject to different kinds of metabolic control.
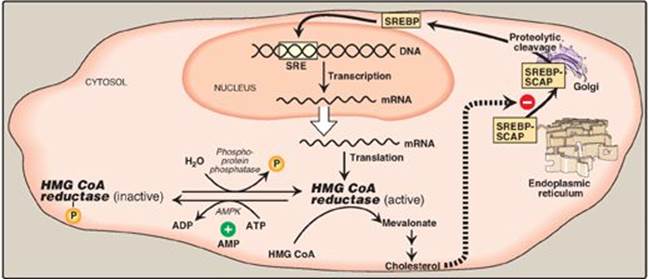
1. Sterol-dependent regulation of gene expression: Expression of the gene for HMG CoA reductase is controlled by the transcription factor, SREBP-2 (sterol regulatory element–binding protein-2) that binds DNA at the cis-acting sterol regulatory element (SRE) upstream of the reductase gene. SREBP-2 is an integral protein of the ER membrane, and associates with a second ER membrane protein, SCAP (SREBP cleavage–activating protein). When sterol levels in the cell are low, the SREBP–SCAP complex moves from the ER to the Golgi. In the Golgi membrane, SREBP-2 is sequentially acted upon by two proteases, which generate a soluble fragment that enters the nucleus, binds the SRE, and functions as a transcription factor. This results in increased synthesis of HMG CoA reductase and, therefore, increased cholesterol synthesis (Figure 18.6). If sterols are abundant, however, they bind SCAP at its sterol-sensing domain and induce the binding of SCAP to yet other ER membrane proteins, the insigs (insulin-induced gene [products]). This results in the retention of the SCAP–SREBP complex in the ER, thereby preventing the activation of SREBP-2, and leading to downregulation of cholesterol synthesis. [Note: SREBP-1 upregulates expression of enzymes involved in fatty acid synthesis in response to insulin (see p. 184).]
2. Sterol-accelerated enzyme degradation: The reductase itself is a sterol-sensing integral protein of the ER membrane. When sterol levels in the cell are high, the enzyme binds to insig proteins. Binding leads to ubiquitination and proteasomal degradation of the reductase (see p. 247).
3. Sterol-independent phosphorylation/dephosphorylation: HMG CoA reductase activity is controlled covalently through the actions of adenosine monophosphate (AMP)-activated protein kinase ([AMPK] see p. 183) and a phosphoprotein phosphatase (see Figure 18.6). The phosphorylated form of the enzyme is inactive, whereas the dephosphorylated form is active. [Note: Because AMPK is activated by AMP, cholesterol synthesis, like fatty acid synthesis, is decreased when ATP availability is decreased.]
4. Hormonal regulation: The amount of HMG CoA reductase is controlled hormonally. An increase in insulin and thyroxine favors upregulation of the expression of the gene for the reductase. Glucagon and the glucocorticoids have the opposite effect.
Figure 18.6 Regulation of hydroxymethylglutaryl coenzyme A (HMG CoA) reductase. SRE = sterol regulatory element; SREBP = sterol regulatory element-binding protein; SCAP = SREBP cleavage-activating protein; AMPK = adenosine monophosphate-activated protein kinase; ADP = adenosine diphosphate; ![]() = phosphate; mRNA = messenger RNA.
= phosphate; mRNA = messenger RNA.
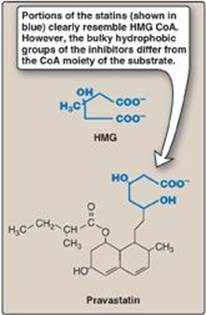
5. Inhibition by drugs: The statin drugs (atorvastatin, fluvastatin, lovastatin, pravastatin, rosuvastatin, and simvastatin) are structural analogs of HMG CoA, and are (or are metabolized to) reversible, competitive inhibitors of HMG CoA reductase (Figure 18.7). They are used to decrease plasma cholesterol levels in patients with hypercholesterolemia.
IV. DEGRADATION OF CHOLESTEROL
The ring structure of cholesterol cannot be metabolized to CO2 and H2O in humans. Rather, the intact sterol nucleus is eliminated from the body by conversion to bile acids and bile salts, a small percentage of which is excreted in the feces, and by secretion of cholesterol into the bile, which transports it to the intestine for elimination. Some of the cholesterol in the intestine is modified by bacteria before excretion. The primary compounds made are the isomers coprostanol and cholestanol, which are reduced derivatives of cholesterol. Together with cholesterol, these compounds make up the bulk of neutral fecal sterols.
Figure 18.7 Structural similarity of hydroxymethylglutaric acid (HMG) and pravastatin, a clinically useful cholesterol-lowering drug of the “statin” family. CoA = coenzyme A.
V. BILE ACIDS AND BILE SALTS
Bile consists of a watery mixture of organic and inorganic compounds. Phosphatidylcholine, or lecithin (see p. 202), and conjugated bile salts are quantitatively the most important organic components of bile. Bile can either pass directly from the liver, where it is synthesized into the duodenum through the common bile duct, or be stored in the gallbladder when not immediately needed for digestion.
A. Structure of the bile acids
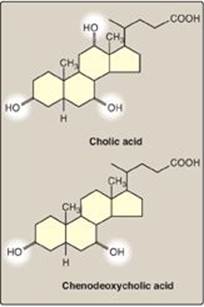
The bile acids contain 24 carbons, with two or three hydroxyl groups and a side chain that terminates in a carboxyl group. The carboxyl group has a pKa of about 6. In the duodenum (pH approximately 6), this group will be protonated in half of the molecules (the bile acids) and deprotonated in the rest (the bile salts). The terms “bile acid” and “bile salt” are frequently used interchangeably, however. Both forms have hydroxyl groups that are α in orientation (they lie “below” the plane of the rings) and the methyl groups that are β (they lie “above” the plane of the rings). Therefore, the molecules have both a polar and a nonpolar face and can act as emulsifying agents in the intestine, helping prepare dietary triacylglycerol and other complex lipids for degradation by pancreatic digestive enzymes.
Figure 18.8 Bile acids. [Note: The ionized forms are bile salts.]
B. Synthesis of bile acids
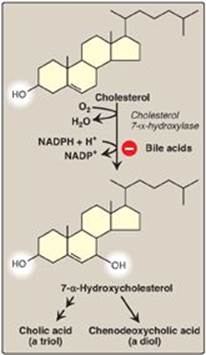
Bile acids are synthesized in the liver by a multistep, multiorganelle pathway in which hydroxyl groups are inserted at specific positions on the steroid structure; the double bond of the cholesterol B ring is reduced; and the hydrocarbon chain is shortened by three carbons, introducing a carboxyl group at the end of the chain. The most common resulting compounds, cholic acid (a triol) and chenodeoxycholic acid (a diol), as shown in Figure 18.8, are called “primary” bile acids. [Note: The rate-limiting step in bile acid synthesis is the introduction of a hydroxyl group at carbon 7 of the steroid nucleus by 7-α-hydroxylase, an ER-associated cytochrome P450 monooxygenasefound only in liver. Expression of the enzyme is downregulated by bile acids (Figure 18.9)]
C. Synthesis of conjugated bile acids
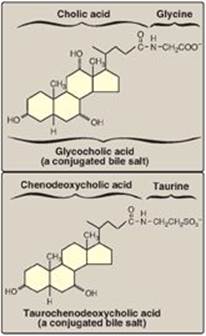
Before the bile acids leave the liver, they are conjugated to a molecule of either glycine or taurine (an end product of cysteine metabolism) by an amide bond between the carboxyl group of the bile acid and the amino group of the added compound. These new structures include glycocholic and glycochenodeoxycholic acids and taurocholic and taurochenodeoxycholic acids (Figure 18.10). The ratio of glycine to taurine forms in the bile is approximately 3:1. Addition of glycine or taurine results in the presence of a carboxyl group with a lower pKa (from glycine) or a sulfonate group (from taurine), both of which are fully ionized (negatively charged) at the alkaline pH of bile. The conjugated, ionized bile salts are more effective detergents than the unconjugated ones because of their enhanced amphipathic nature. Therefore, only the conjugated forms are found in the bile. Individuals with genetic deficiencies in the conversion of cholesterol to bile acids are treated with exogenously supplied chenodeoxycholic acid.
Bile salts provide the only significant mechanism for cholesterol excretion, both as a metabolic product of cholesterol and as a solubilizer of cholesterol in bile.
Figure 18.9 Synthesis of the bile acids, cholic acid and chenodeoxycholic acid, from cholesterol.
D. Action of intestinal flora on bile salts
Bacteria in the intestine can deconjugate (remove glycine and taurine) bile salts. They can also remove the hydroxyl group at carbon 7, producing “secondary” bile salts such as deoxycholic acid from cholic acid and lithocholic acid from chenodeoxycholic acid (Figure 18.11).
Figure 18.10 Conjugated bile salts. Note “cholic” in the names.
E. Enterohepatic circulation
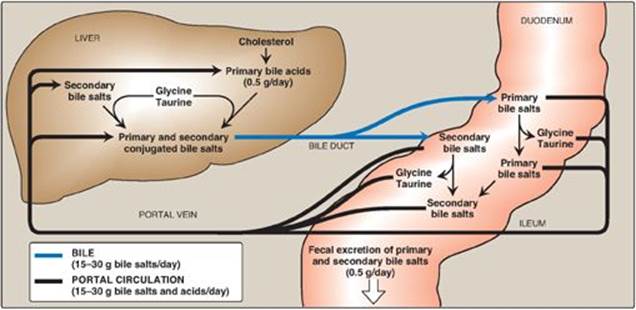
Bile salts secreted into the intestine are efficiently reabsorbed (greater than 95%) and reused. The liver actively secretes bile salts into the bile. In the intestine, they are reabsorbed in the terminal ileum via a Na+-bile salt cotransporter and returned to the blood via a separate transport system. [Note: Lithocolic acid is only poorly absorbed.] They are efficiently taken up from blood by the hepatocytes via an isoform of the cotransporter and reused. [Note: Albumin binds bile salts noncovalently and transports them through the blood as was seen with fatty acids (see p. 181).] The continuous process of secretion of bile salts into the bile, their passage through the duodenum where some are deconjugated then dehydroxylated to secondary bile salts, their uptake in the ileum, and their subsequent return to the liver as a mixture of primary and secondary forms is termed the enterohepatic circulation (see Figure 18.11). Between 15 and 30 g of bile salts are secreted from the liver into the duodenum each day, yet only about 0.5 g (less than 3%) is lost daily in the feces. Approximately 0.5 g/day is synthesized from cholesterol in the liver to replace the amount lost. Bile acid sequestrants, such as cholestyramine, bind bile salts in the gut; prevent their reabsorption; and, so, promote their excretion. They are used in the treatment of hypercholesterolemia because the removal of bile salts relieves the inhibition on bile acid synthesis in the liver, thereby diverting additional cholesterol into that pathway. [Note: Dietary fiber also binds bile salts and increases their excretion (see p. 365).]
Figure 18.11 Enterohepatic circulation of bile salts. [Note: Primary forms are converted to secondary forms by dehydroxylation.]
F. Bile salt deficiency: cholelithiasis
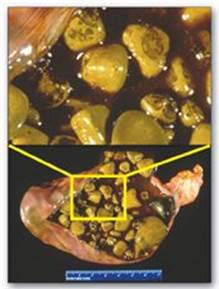
The movement of cholesterol from the liver into the bile must be accompanied by the simultaneous secretion of phospholipid and bile salts. If this dual process is disrupted and more cholesterol is present than can be solubilized by the bile salts and phosphatidylcholine present, the cholesterol may precipitate in the gallbladder, leading to cholesterol gallstone disease, or cholelithiasis (Figure 18.12). This disorder is typically caused by a decrease of bile acids in the bile. Cholelithiasis also may result from increased secretion of cholesterol into bile, as seen with the use of fibrates (for example, gemfibrozil) to reduce cholesterol (and triacylglycerol) in the blood. Laparoscopic cholecystectomy (surgical removal of the gallbladder through a small incision) is currently the treatment of choice. However, for patients who are unable to undergo surgery, oral administration of chenodeoxycholic acid to supplement the body’s supply of bile acids results in a gradual (months to years) dissolution of the gallstones. [Note: Cholesterol stones account for over 85% of cases of cholelithiasis, with bilirubin and mixed stones accounting for the rest].
Figure 18.12 Gallbladder with gallstones.
VI. PLASMA LIPOPROTEINS
The plasma lipoproteins are spherical macromolecular complexes of lipids and specific proteins (apolipoproteins). The lipoprotein particles include chylomicrons, very-low-density lipoproteins (VLDLs), low-density lipoproteins (LDLs), and high-density lipoproteins (HDLs). They differ in lipid and protein composition, size, density (Figure 18.13), and site of origin. [Note: Because lipoprotein particles constantly interchange lipids and apolipoproteins with each other, the actual apolipoprotein and lipid content of each class of particles is somewhat variable.] Lipoproteins function both to keep their component lipids soluble as they transport them in the plasma and to provide an efficient mechanism for transporting their lipid contents to (and from) the tissues. In humans, the transport system is less perfect than in other animals and, as a result, humans experience a gradual deposition of lipid (especially cholesterol) in tissues. This is a potentially life-threatening occurrence when the lipid deposition contributes to plaque formation, causing the narrowing of blood vessels (atherosclerosis).
A. Composition of plasma lipoproteins
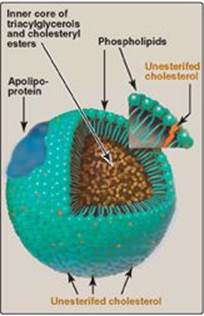
Lipoproteins are composed of a neutral lipid core (containing triacylglycerol [TAG] and cholesteryl esters) surrounded by a shell of amphipathic apolipoproteins, phospholipid, and unesterified (free) cholesterol (Figure 18.14). These amphipathic compounds are oriented so that their polar portions are exposed on the surface of the lipoprotein, thereby rendering the particle soluble in aqueous solution. The TAG and cholesterol carried by the lipoproteins are obtained either from the diet (exogenous source) or from de novo synthesis (endogenous source). [Note: The cholesterol (C) content of plasma lipoproteins is now routinely measured in fasting blood. Total C = LDL – C + HDL – C + VLDL – C, where VLDL – C is calculated by dividing TAG by 5 because TAG represent 20% of the volume of VLDL. The goal value for total cholesterol is less than 200 mg/dl.]
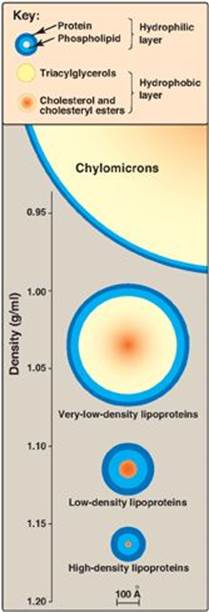
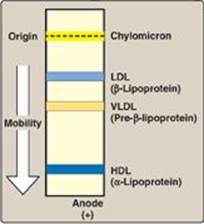
1. Size and density of lipoprotein particles: Chylomicrons are the lipoprotein particles lowest in density and largest in size and that contain the highest percentage of lipid (as TAG) and the lowest percentage of protein. VLDLs and LDLs are successively denser, having higher ratios of protein to lipid. HDL particles are the smallest and densest. Plasma lipoproteins can be separated on the basis of their electrophoretic mobility, as shown in Figure 18.15, or on the basis of their density by ultracentrifugation.
2. Apolipoproteins: The apolipoproteins associated with lipoprotein particles have a number of diverse functions, such as providing recognition sites for cell-surface receptors and serving as activators or coenzymes for enzymes involved in lipoprotein metabolism. Some of the apolipoproteins are required as essential structural components of the particles and cannot be removed (in fact, the particles cannot be produced without them), whereas others are transferred freely between lipoproteins. Apolipoproteins are divided by structure and function into several major classes, denoted by letters, with each class having subclasses (for example, apolipoprotein [apo] C-I, apo C-II, and apo C-III). [Note: Functions of all of the apolipoproteins are not yet known.]
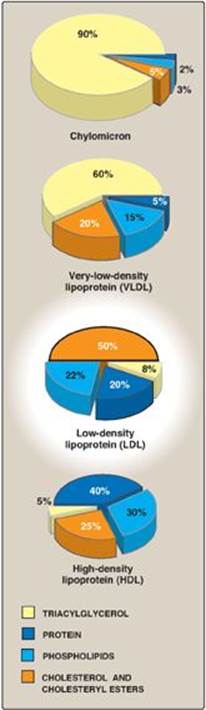
Figure 18.13 Approximate size and density of serum lipoproteins. Each family of lipoproteins exhibits a range of sizes and densities, and this figure shows typical values. The width of the rings approximates the amount of each component. [Note: Although cholesterol and its esters are shown as one component in the center of each particle, physically, cholesterol is a surface component, whereas cholesteryl esters are located in the interior of the lipoproteins.]
B. Metabolism of chylomicrons
Chylomicrons are assembled in intestinal mucosal cells and carry dietary (exogenous) TAG, cholesterol, fat-soluble vitamins, and cholesteryl esters to the peripheral tissues (Figure 18.16). [Note: TAGs account for close to 90% of the lipids in a chylomicron.]
1. Synthesis of apolipoproteins: Apo B-48 is unique to chylomicrons. Its synthesis begins on the rough ER, and it is glycosylated as it moves through the ER and Golgi. [Note: Apo B-48 is so named because it constitutes the N-terminal 48% of the protein encoded by the gene for apo B. Apo B-100, which is synthesized by the liver and found in VLDL and LDL, represents the entire protein encoded by this gene. Posttranscriptional editing (see p. 457) of a cytosine to a uracil in intestinal apo B-100 messenger RNA (mRNA) creates a nonsense (stop) codon (see p. 433), allowing translation of only 48% of the mRNA.]
2. Assembly of chylomicrons: The enzymes involved in TAG, cholesterol, and phospholipid synthesis are located in the smooth ER. Assembly of the apolipoproteins and lipid into chylomicrons requires microsomal triglyceride transfer protein ([MTP], see p. 178), which loads apo B-48 with lipid. This occurs before transition from the ER to the Golgi, where the particles are packaged in secretory vesicles. These fuse with the plasma membrane releasing the lipoproteins, which then enter the lymphatic system and, ultimately, the blood. [Note: Chylomicrons leave the lymphatic system via the thoracic duct that empties into the left subclavian vein.]
Figure 18.14 Structure of a typical lipoprotein particle.
3. Modification of nascent chylomicron particles: The particle released by the intestinal mucosal cell is called a “nascent” chylomicron because it is functionally incomplete. When it reaches the plasma, the particle is rapidly modified, receiving apolipoproteins E (which is recognized by hepatic receptors) and C. The latter includes apo C-II, which is necessary for the activation of lipoprotein lipase (LPL), the enzyme that degrades the TAG contained in the chylomicron (see below). The source of these apolipoproteins is circulating HDL (see Figure 18.16).
4. Degradation of triacylglycerol by lipoprotein lipase: LPL is an extracellular enzyme that is anchored by heparan sulfate to the capillary walls of most tissues, but predominantly those of adipose tissue and cardiac and skeletal muscle. Adult liver does not have this enzyme. [Note: A hepatic lipase is found on the surface of endothelial cells of the liver. It plays some role in TAG degradation in chylomirons and VLDL and is particularly important in HDL metabolism (see p. 236).] LPL, activated by apo C-II on circulating lipoprotein particles, hydrolyzes the TAG contained in these particles to yield fatty acids and glycerol. The fatty acids are stored (by the adipose) or used for energy (by the muscle). If they are not immediately taken up by a cell, the long-chain fatty acids are transported by serum albumin until their uptake does occur. The glycerol is used by the liver, for example, in lipid synthesis or gluconeogenesis. [Note: Patients with a deficiency of LPL or apo C-II (type 1 hyperlipoproteinemia, or familial LPL-deficiency) show a dramatic accumulation (1,000 mg/dl or greater) of chylomicron-TAG in the plasma (hypertriacylglycerolemia) even in the fasted state. These individuals are at increased risk for acute pancreatitis.]
Figure 18.15 Electrophoretic mobility of plasma lipoproteins. The order of low-density lipoprotein (LDL) and very-low-density lipoprotein (VLDL) is reversed if ultracentrifugation is used as the separation technique. HDL = highdensity lipoprotein.
5. Regulation of lipoprotein lipase activity: LPL is synthesized by adipose tissue and by cardiac and skeletal muscle. Expression of the tissue-specific isozymes is regulated by nutritional state and hormonal level. For example, in the fed state (elevated insulin levels), LPL synthesis is increased in adipose but decreased in muscle tissue. Fasting (decreased insulin) favors LPL synthesis in muscle. [Note: The highest concentration of LPL is in cardiac muscle, reflecting the use of fatty acids to provide much of the energy needed for cardiac function.]
Figure 18.16 Metabolism of chylomicrons. Apo B-48, apo C-II, and apo E are apolipoproteins found as specific components of plasma lipoproteins. The lipoprotein particles are not drawn to scale (see Figure 18.13 for details of their size and density). TAG = triacylglycerol; C = cholesterol; CE = cholesteryl ester; HDL = high-density lipoprotein particle.
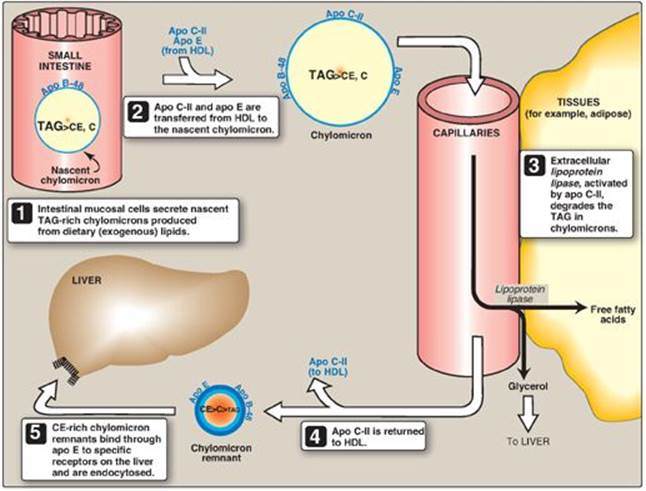
6. Formation of chylomicron remnants: As the chylomicron circulates, and more than 90% of the TAG in its core is degraded by LPL, the particle decreases in size and increases in density. In addition, the C apolipoproteins (but not apo E) are returned to HDL. The remaining particle, called a “remnant,” is rapidly removed from the circulation by the liver, whose cell membranes contain lipoprotein receptors that recognize apo E (see Figure 18.16). Chylomicron remnants bind to these receptors and are taken into the hepatocytes by endocytosis. The endocytosed vesicle then fuses with a lysosome, and the apolipoproteins, cholesteryl esters, and other components of the remnant are hydrolytically degraded, releasing amino acids, free cholesterol, and fatty acids. The receptor is recycled. (A more detailed discussion of the mechanism of receptor-mediated endocytosis is illustrated for LDL in Figure 18.20.)
C. Metabolism of very-low-density lipoproteins
VLDLs are produced in the liver (Figure 18.17). They are composed predominantly of endogenous TAG (approximately 60%), and their function is to carry this lipid from the liver (site of synthesis) to the peripheral tissues. There, the TAG is degraded by LPL, as discussed for chylomicrons (see p. 228). [Note: Nonalcoholic “fatty liver” (hepatic steatosis) occurs in conditions in which there is an imbalance between hepatic TAG synthesis and the secretion of VLDL. Such conditions include obesity and type 2 diabetes mellitus.]
Figure 18.17 Metabolism of very-low-density lipoprotein (VLDL) and low-density lipoprotein (LDL). Apo B-100, apo C-II, and apo E are apolipoproteins found as specific components of plasma lipoprotein particles. The lipoproteins are not drawn to scale (see Figure 18.13 for details of their size and density). TAG = triacylglycerol; HDL = high-density lipoprotein particle; IDLs = intermediate-density lipoprotein particles; C = cholesterol; CE = cholesteryl ester.
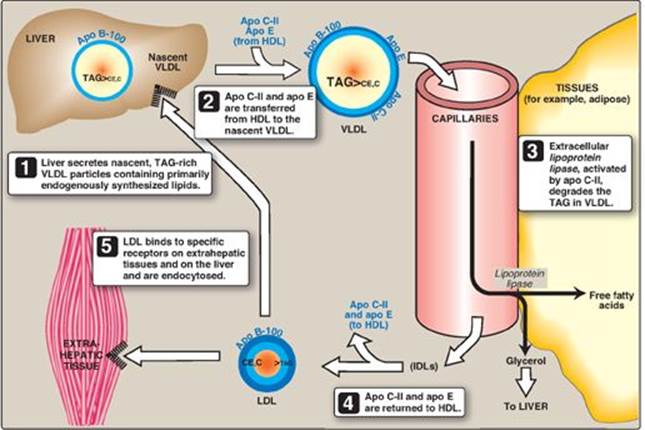
1. Release from liver: VLDLs are secreted directly into the blood by the liver as nascent particles containing apo B-100. They must obtain apo C-II and apo E from circulating HDL (see Figure 18.17). As with chylomicrons, apo C-II is required for activation of LPL. [Note: Abetalipoproteinemia is a rare hypolipoproteinemia caused by a defect in MTP, leading to an inability to load apo B with lipid. As a consequence, few VLDLs or chylomicrons are formed, and TAGs accumulate in the liver and intestine.]
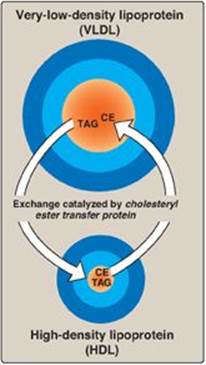
2. Modification in the circulation: As VLDLs pass through the circulation, TAG is degraded by LPL, causing the VLDLs to decrease in size and become denser. Surface components, including the C and E apolipoproteins, are returned to HDL, but the particles retain apo B-100. Additionally, some TAGs are transferred from VLDL to HDL in an exchange reaction that concomitantly transfers cholesteryl esters from HDL to VLDL. This exchange is accomplished by cholesteryl ester transfer protein (CETP) as shown in Figure 18.18.
3. Conversion to low-density lipoproteins: With these modifications, the VLDL is converted in the plasma to LDL. Intermediate-sized particles, the intermediate-density lipoproteins (IDLs), or VLDL remnants, are observed during this transition. IDLs can also be taken up by liver cells through receptor-mediated endocytosis that uses apo E as the ligand. [Note: Apo E is normally present in three isoforms, E-2, (the least common), E-3 (the most common), and E-4. Apo E-2 binds poorly to receptors, and patients who are homozygotic for apo E-2 are deficient in the clearance of chylomicron remnants and IDLs. These individuals have familial type III hyperlipoproteinemia (familial dysbetalipoproteinemia, or broad beta disease), with hypercholesterolemia and premature atherosclerosis. Not as yet well understood is the fact that the E-4 isoform confers increased susceptibility to and decreased age of onset of the late-onset form of Alzheimer disease. The effect is dose dependent, with homozygotes being at greatest risk. Estimates of the risk vary.]
Figure 18.18 Transfer of cholesteryl ester (CE) from HDL to VLDL in exchange for triacylglycerol (TAG).
D. Metabolism of low-density lipoproteins
LDL particles contain much less TAG than their VLDL predecessors and have a high concentration of cholesterol and cholesteryl esters (Figure 18.19).
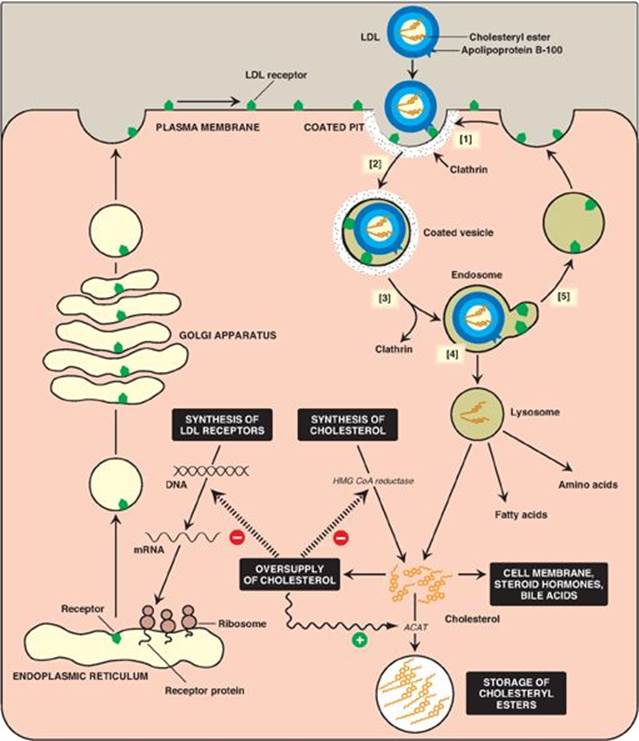
1. Receptor-mediated endocytosis: The primary function of LDL particles is to provide cholesterol to the peripheral tissues (or return it to the liver). They do so by binding to cell surface membrane LDL receptors that recognize apo B-100 (but not apo B-48). Because these LDL receptors can also bind apo E, they are known as apo B-100/apo E receptors. A summary of the uptake and degradation of LDL particles is presented in Figure 18.20. [Note: The numbers in brackets below refer to corresponding numbers on that figure.] A similar mechanism of receptor-mediated endocytosis is used for the cellular uptake and degradation of chylomicron remnants and IDLs by the liver.
[1] LDL receptors are negatively charged glycoproteins that are clustered in pits on cell membranes. The cytosolic side of the pit is coated with the protein clathrin, which stabilizes the pit.
[2] After binding, the LDL–receptor complex is taken in by endocytosis. [Note: A deficiency of functional LDL receptors causes a significant elevation in plasma LDL-C. Patients with such deficiencies have type II hyperlipidemia (familial hypercholesterolemia, or FH) and premature atherosclerosis. Autosomal-dominant hypercholesterolemia can also be caused by increased activity of a protease, proprotein convertase subtilisin/kexin type 9 (PCSK9), which promotes degradation of the receptor, and by defects in apo B-100 that reduce its binding to the receptor.]
[3] The vesicle containing LDL loses its clathrin coat and fuses with other similar vesicles, forming larger vesicles called endosomes.
[4] The pH of the endosome falls (due to the proton-pumping activity of endosomal ATPase), which allows separation of the LDL from its receptor. The receptors then migrate to one side of the endosome, whereas the LDLs stay free within the lumen of the vesicle. [Note: This structure is called CURL, the compartment for uncoupling of receptor and ligand.]
[5] The receptors can be recycled, whereas the lipoprotein remnants in the vesicle are transferred to lysosomes and degraded by lysosomal acid hydrolases, releasing free cholesterol, amino acids, fatty acids, and phospholipids. These compounds can be reutilized by the cell. [Note: Storage diseases caused by rare autosomal-recessive deficiencies in the ability to hydrolyze lysosomal cholesteryl esters (late-onset Wolman disease), or to transport free cholesterol out of the lysosome (Niemann-Pick disease, Type C) have been identified.]
Figure 18.19 Composition of the plasma lipoproteins. Note the high concentration of cholesterol and cholesteryl esters in LDL.
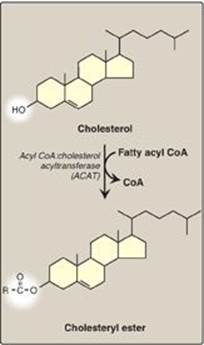
2. Effect of endocytosed cholesterol on cellular cholesterol homeostasis: The chylomicron remnant-, IDL-, and LDL-derived cholesterol affects cellular cholesterol content in several ways (see Figure 18.20). First, expression of the gene for HMG CoA reductase is inhibited by high cholesterol, as a result of which, de novo cholesterol synthesis decreases. Additionally, degradation of the reductase is accelerated. Second, synthesis of new LDL receptor protein is reduced by decreasing the expression of the LDL receptor gene, thus limiting further entry of LDL cholesterol into cells. [Note: Regulation of the LDL receptor gene involves a SRE and SREBP-2, as was seen in the regulation of the gene for HMG CoA reductase (see p. 222). This allows coordinate regulation of the expression of these proteins.] Third, if the cholesterol is not required immediately for some structural or synthetic purpose, it is esterified by acyl CoA:cholesterol acyltransferase (ACAT). ACAT transfers a fatty acid from a fatty acyl CoA to cholesterol, producing a cholesteryl ester that can be stored in the cell (Figure 18.21). The activity of ACAT is enhanced in the presence of increased intracellular cholesterol.
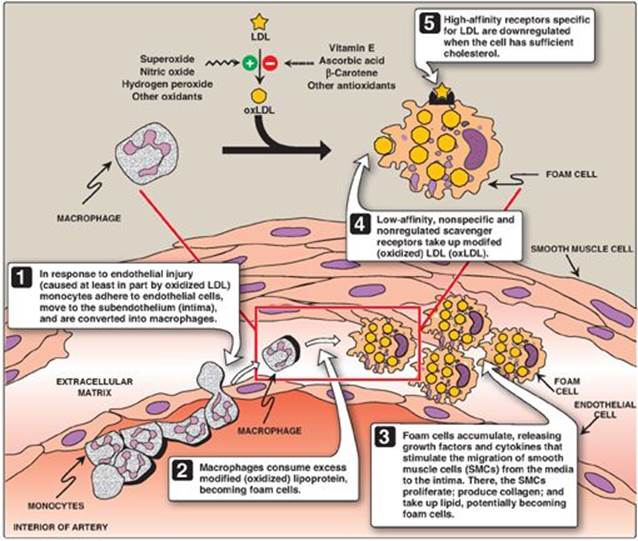
3. Uptake of chemically modified LDL by macrophage scavenger receptors: In addition to the highly specific and regulated receptor-mediated pathway for LDL uptake described above, macrophages possess high levels of scavenger receptor activity. These receptors, known as scavenger receptor class A (SR-A), can bind a broad range of ligands and mediate the endocytosis of chemically modified LDL in which the lipid components or apo B have been oxidized. Unlike the LDL receptor, the scavenger receptor is not downregulated in response to increased intracellular cholesterol. Cholesteryl esters accumulate in macrophages and cause their transformation into “foam” cells, which participate in the formation of atherosclerotic plaque (Figure 18.22).
Figure 18.20 Cellular uptake and degradation of low-density lipoprotein (LDL). [Note: Oversupply of cholesterol accelerates the degradation of HMG CoA reductase. It also decreases synthesis of the reductase by preventing expression of its gene as seen with the LDL receptor.] ACAT = acyl CoA:cholesterol acyltransferase; HMG CoA = hydroxymethylglutaryl coenzyme A; mRNA = messenger RNA.
E. Metabolism of high-density lipoproteins
HDLs comprise a heterogeneous family of lipoproteins with a complex metabolism that is not yet completely understood. HDL particles are formed in blood by the addition of lipid to apo A-1, an apolipoprotein made by the liver and intestine and secreted into blood. [Note: HDLs are also formed within the liver and intestine.] Apo A-1 accounts for about 70% of the apolipoproteins in HDL. HDLs perform a number of important functions, including the following.
1. Apolipoprotein supply: HDL particles serve as a circulating reservoir of apo C-II (the apolipoprotein that is transferred to VLDL and chylomicrons and is an activator of LPL) and apo E (the apolipoprotein required for the receptor-mediated endocytosis of IDLs and chylomicron remnants).
2. Uptake of unesterified cholesterol: Nascent HDLs are disc-shaped particles containing primarily phospholipid (largely phosphatidylcholine) and apolipoproteins A, C, and E. They take up cholesterol from nonhepatic (peripheral) tissues and return it to the liver as cholesteryl esters (Figure 18.23). [Note: HDL particles are excellent acceptors of unesterified cholesterol as a result of their high concentration of phospholipids, which are important solubilizers of cholesterol.]
Figure 18.21 Synthesis of intracellular cholesteryl ester by ACAT. [Note: Lecithin:cholesterol acyl transferase (LCAT) is the extracellular enzyme that esterifies cholesterol using phosphatidylcholine (lecithin) as the source of the fatty acid.] CoA = coenzyme A.
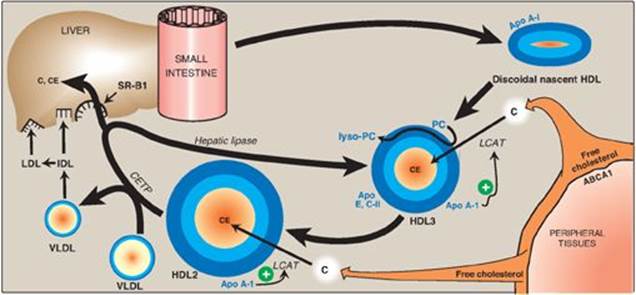
3. Esterification of cholesterol: When cholesterol is taken up by HDL, it is immediately esterified by the plasma enzyme lecithin:cholesterol acyltransferase (LCAT, also known as PCAT, in which “P” stands for phosphatidylcholine, the source of the fatty acid). This enzyme is synthesized and secreted by the liver. LCAT binds to nascent HDL, and is activated by apo A-I. LCAT transfers the fatty acid from carbon 2 of phosphatidylcholine to cholesterol. This produces a hydrophobic cholesteryl ester, which is sequestered in the core of the HDL, and lysophosphatidylcholine, which binds to albumin. [Note: Esterification maintains the cholesterol concentration gradient, allowing continued efflux of cholesterol to HDL.] As the discoidal nascent HDL accumulates cholesteryl esters, it first becomes a spherical, relatively cholesteryl ester–poor HDL3 and, eventually, a cholesteryl ester–rich HDL2 particle that carries these esters to the liver. CETP (see p. 231) moves some of the cholesteryl esters from HDL to VLDL in exchange for TAG, relieving product inhibition of LCAT. Because VLDLs are catabolized to LDL, the cholesteryl esters transferred by CETP are ultimately taken up by the liver (see p. 236).
Figure 18.22 Role of oxidized lipoproteins in plaque formation in an arterial wall. LDL = low-density lipoprotein.
4. Reverse cholesterol transport: The selective transfer of cholesterol from peripheral cells to HDL, from HDL to the liver for bile acid synthesis or disposal via the bile, and to steroidogenic cells for hormone synthesis, is a key component of cholesterol homeostasis. This process of reverse cholesterol transport is, in part, the basis for the inverse relationship seen between plasma HDL concentration and atherosclerosis and for HDL’s designation as the “good” cholesterol carrier. [Note: Exercise and estrogen raise HDL levels.] Reverse cholesterol transport involves efflux of cholesterol from peripheral cells to HDL, esterification of the cholesterol by LCAT, binding of the cholesteryl ester–rich HDL (HDL2) to liver (and steroidogenic cells), the selective transfer of the cholesteryl esters into these cells, and the release of lipid-depleted HDL (HDL3). The efflux of cholesterol from peripheral cells is mediated, at least in part, by the transport protein ABCA1. [Note: Tangier disease is a very rare deficiency of ABCA1 and is characterized by the virtual absence of HDL particles due to degradation of lipid-poor apo A-1.] The uptake of cholesteryl esters by the liver is mediated by a cell-surface receptor, SR-B1 (scavenger receptor class B type 1) that binds HDL (see p. 234 for SR-A receptors). The HDL particle itself is not taken up. Instead, there is selective uptake of the cholesteryl ester from the HDL particle. [Note: Hepatic lipase, with its ability to degrade both TAG and phospholipids, also participates in the conversion of HDL2 to HDL3.]
Figure 18.23 Metabolism of high-density lipoprotein (HDL) particles. Apo = apolipoprotein; ABCA1 = transport protein; C = cholesterol; CE = cholesteryl ester; LCAT = lecithin:cholesterol acyltransferase; VLDL = very-low-density lipoprotein; IDL = intermediate-density lipoprotein; LDL = low-density lipoprotein; CETP = cholesteryl ester transfer protein; SR-B1 = scavenger receptor B1.
ABCA1 is an ATP-binding cassette (ABC) protein. ABC proteins use energy from ATP hydrolysis to transport materials, including lipids, in and out of cells and across intracellular compartments. In addition to Tangier disease, defects in specific ABC proteins result in sitosterolemia, X-linked adrenoleukodystrophy, respiratory distress syndrome due to decreased surfactant secretion, and cystic fibrosis.
F. Role of lipoprotein (a) in heart disease
Lipoprotein (a), or Lp(a), is a particle that, when present in large quantities in the plasma, is associated with an increased risk of coronary heart disease. Lp(a) is nearly identical in structure to an LDL particle. Its distinguishing feature is the presence of an additional apolipoprotein molecule, apo(a), that is covalently linked at a single site to apo B-100. Circulating levels of Lp(a) are determined primarily by genetics. However, factors such as diet may play some role, as trans fatty acids have been shown to increase Lp(a), whereas ω-3 fatty acids decrease it. [Note: Apo(a) is structurally homologous to plasminogen, the precursor of a blood protease whose target is fibrin, the main protein component of blood clots (See Chapter 34 online). It is hypothesized that elevated Lp(a) slows the breakdown of blood clots that trigger heart attacks because it competes with plasminogen for binding to fibrin. The physiologic function of Lp(a) in unknown. Niacin reduces Lp(a), as well as LDL-cholesterol and TAGs, and raises HDL.]
VII. STEROID HORMONES
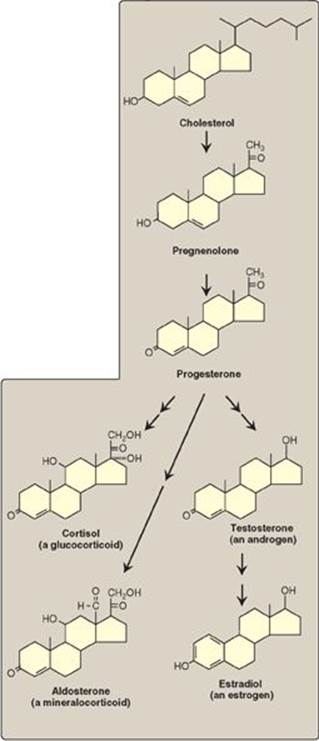
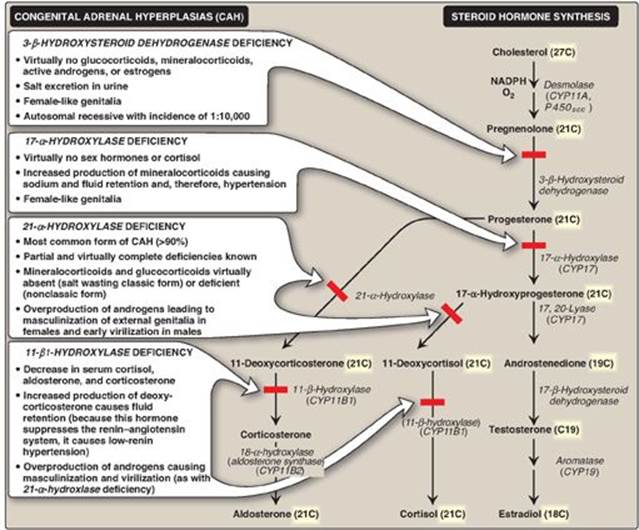
Cholesterol is the precursor of all classes of steroid hormones: glucocorticoids (for example, cortisol), mineralocorticoids (for example, aldosterone), and the sex hormones (that is, androgens, estrogens, and progestins) as shown in Figure 18.24). [Note: Glucocorticoids and mineralocorticoids are collectively called corticosteroids.] Synthesis and secretion occur in the adrenal cortex (cortisol, aldosterone, and androgens), ovaries and placenta (estrogens and progestins), and testes (testosterone). Steroid hormones are transported by the blood from their sites of synthesis to their target organs. Because of their hydrophobicity, they must be complexed with a plasma protein. Plasma albumin can act as a nonspecific carrier and does carry aldosterone. However, specific steroid-carrier plasma proteins bind the steroid hormones more tightly than does albumin (for example, corticosteroid-binding globulin, or transcortin, is responsible for transporting cortisol). A number of genetic diseases are caused by deficiencies in specific steps in the biosynthesis of steroid hormones. Some representative diseases are described in Figure 18.25.
Figure 18.24 Key steroid hormones.
A. Synthesis of steroid hormones
Synthesis involves shortening the hydrocarbon chain of cholesterol and hydroxylation of the steroid nucleus. The initial and rate-limiting reaction converts cholesterol to the 21-carbon pregnenolone. It is catalyzed by the cholesterol side-chain cleavage enzyme (desmolase, P450scc), a cytochrome P450 (CYP) mixed-function oxidase of the inner mitochondrial membrane (see p. 149). NADPH and O2 are required for the reaction. The cholesterol substrate can be newly synthesized, taken up from lipoproteins, or released by an esterase from cholesteryl esters stored in the cytosol of steroidogenic tissues. The cholesterol moves to the outer mitochondrial membrane. An important control point is the subsequent movement from the outer to the inner mitochondrial membrane. This process is mediated by StAR (steroidogenic acute regulatory) protein. Pregnenolone is the parent compound for all steroid hormones (see Figure 18.25). It is oxidized and then isomerized to progesterone, which is further modified to the other steroid hormones by hydroxylation reactions that occur in the ER and mitochondria. Like desmolase, the enzymes primarily are CYP proteins. A defect in the activity or amount of an enzyme in this pathway can lead to a deficiency in the synthesis of hormones beyond the affected step and to an excess in the hormones or metabolites before that step. Because all members of the pathway have potent biologic activity, serious metabolic imbalances occur with enzyme deficiencies (see Figure 18.25). Collectively these disorders are known as the congenital adrenal hyperplasias. [Note: Addison disease, due to autoimmune destruction of the adrenal cortex, is characterized by adrenocortical insufficiency.]
Figure 18.25 Steroid hormone synthesis and associated diseases. [Note: 3-β-Hydroxysteroid dehydrogenase, CYP17, and CYP11B2 are bifunctional enzymes. Synthesis of testosterone and the estrogens from cholesterol occurs primarily outside of the adrenal gland.] NADPH = nicotinamide adenine dinucleotide phosphate; CYP = cytochrome P450.
B. Secretion of adrenal cortical steroid hormones
Steroid hormones are secreted on demand from their tissues of origin in response to hormonal signals. The corticosteroids and androgens are made in different regions of the adrenal cortex and are secreted into blood in response to different signals.
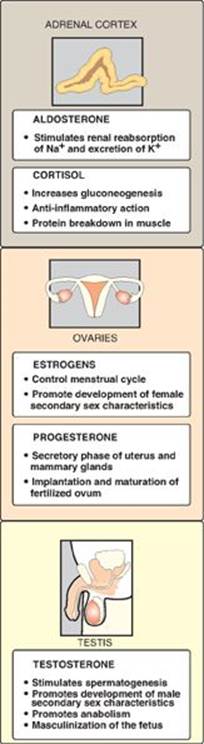
1. Cortisol: Its production in the middle layer (zona fasciculata) of the adrenal cortex is controlled by the hypothalamus, to which the pituitary gland is attached (Figure 18.26). In response to severe stress (for example, infection), corticotropin-releasing hormone (CRH), produced by the hypothalamus, travels through capillaries to the anterior lobe of the pituitary, where it induces the production and secretion of adrenocorticotropic hormone (ACTH). The polypeptide ACTH, the “stress hormone,” stimulates the adrenal cortex to synthesize and secrete the glucocorticoid cortisol. Cortisol allows the body to respond to stress through its effects on intermediary metabolism (for example, increased gluconeogenesis) and the inflammatory and immune responses. As cortisol levels rise, the release of CRH and ACTH is inhibited. [Note: ACTH binds to a membrane G protein–coupled receptor, resulting in cyclic AMP (cAMP) production and activation of protein kinase A (PKA) (see p. 94). PKA phosphorylates and activates both the esterase that converts cholesteryl ester to free cholesterol and StAR protein.]
2. Aldosterone: Its production in the outer layer (zona glomerulosa) of the adrenal cortex is induced by a decrease in the plasma Na+/K+ ratio, and by the hormone angiotensin II. Angiotensin II (an octapeptide) is produced from angiotensin I (a decapeptide) by angiotensin-converting enzyme (ACE), an enzyme found predominantly in the lungs but also distributed widely in the body. [Note: Angiotensin I is produced in the blood by cleavage of an inactive precursor, angiotensinogen, secreted by the liver. Cleavage is accomplished by the enzyme renin, made and secreted by the kidney.] Angiotensin II binds to cell-surface receptors. However, in contrast to ACTH, its effects are mediated through the phosphatidylinositol 4,5-bisphosphate pathway (see p. 205) and not by cAMP. Aldosterone’s primary effect is on the kidney tubules, where it stimulates sodium and water uptake and potassium excretion (Figure 18.27). [Note: An effect of aldosterone is an increase in blood pressure. Competitive inhibitors of ACE are used to treat renin-dependent hypertension.]
Figure 18.26 Pituitary hormone stimulation of steroid hormone synthesis and secretion.
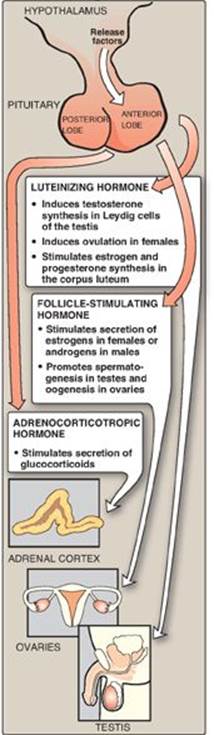
3. Androgens: Both the inner (zona reticularis) and middle layers of the adrenal cortex produce androgens, primarily dehydroepiandrosterone and androstenedione. Although adrenal androgens themselves are weak, they are converted in peripheral tissues to testosterone, a stronger androgen, and to estrogens.
Estrogens are derived from androstenedione and testosterone by aromatase (CYP19). Aromatase inhibitors are used in the treatment of estrogen-responsive breast cancer in postmenopausal women.
C. Secretion of steroid hormones from gonads
The testes and ovaries synthesize hormones necessary for sexual differentiation and reproduction. A single hypothalamic-releasing factor, gonadotropin-releasing hormone, stimulates the anterior pituitary to release the glycoproteins luteinizing hormone (LH) and follicle-stimulating hormone (FSH). Like ACTH, LH and FSH bind to surface receptors and cause an increase in cAMP. LH stimulates the testes to produce testosterone and the ovaries to produce estrogens and progesterone (see Figure 18.27). FSH regulates the growth of ovarian follicles and stimulates testicular spermatogenesis.
D. Mechanism of steroid hormone action
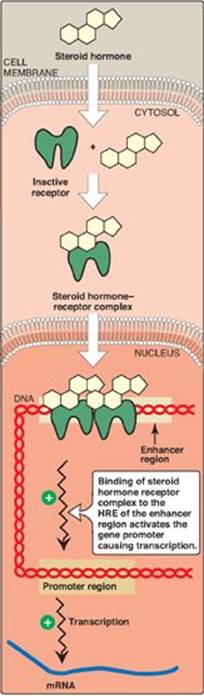
Each steroid hormone diffuses across the plasma membrane of its target cell and binds to a specific cytosolic or nuclear receptor. These receptor–ligand complexes accumulate in the nucleus, dimerize, and bind to specific regulatory DNA sequences (hormone-response elements [HREs]) in association with coactivator proteins, thereby causing promoter activation and increased transcription of targeted genes (Figure 18.28). An HRE is found in the promoter or an enhancer element (see p. 424) for genes that respond to a specific steroid hormone, thus insuring coordinated regulation of these genes. Hormone–receptor complexes can also inhibit transcription in association with corepressors. [Note: The binding of a hormone to its receptor causes a conformational change in the receptor that uncovers its DNA-binding domain, allowing the complex to interact through a zinc-finger motif with the appropriate sequence on the DNA. Receptors for the steroid hormones, plus those for thyroid hormone, retinoic acid (see p. 382), and 1,25-dihydroxycholecalciferol (vitamin D, see p. 386), are members of a “superfamily” of structurally related gene regulators that function in a similar way.]
Figure 18.27 Actions of steroid hormones.
E. Further metabolism of steroid hormones
Steroid hormones are generally converted into inactive metabolic excretion products in the liver. Reactions include reduction of unsaturated bonds and the introduction of additional hydroxyl groups. The resulting structures are made more soluble by conjugation with glucuronic acid or sulfate (from 3ʹ-phosphoadenosyl-5ʹ-phosphosulfate, see p. 162). Approximately 20%–30% of these metabolites are secreted into the bile and then excreted in the feces, whereas the remainder are released into the blood and filtered from the plasma in the kidney, passing into the urine. These conjugated metabolites are fairly water soluble and do not need protein carriers.
VIII. CHAPTER SUMMARY
Cholesterol is a hydrophobic compound, with a single hydroxyl group located at carbon 3 of the A ring, to which a fatty acid can be attached, producing an even more hydrophobic cholesteryl ester. Cholesterol is synthesized by virtually all human tissues, although primarily by liver, intestine, adrenal cortex, and reproductive tissues (Figure 18.29). All the carbon atoms in cholesterol are provided by acetyl coenzyme A (CoA), and nicotinamide adenine dinucleotide phosphate provides the reducing equivalents. The pathway is driven by hydrolysis of the high-energy thioester bond of acetyl CoA and the terminal phosphate bond of adenosine triphosphate. Cholesterol is synthesized in the cytoplasm. The rate-limiting and regulated step in cholesterol synthesis is catalyzed by the smooth endoplasmic reticulum–membrane protein, hydroxymethylglutaryl coenzyme A (HMG CoA) reductase, which produces mevalonate from HMG CoA. The enzyme is regulated by a number of mechanisms: 1) Expression of the gene for the reductase is activated when cholesterol levels are low, via the transcription factor, sterol regulatory element–binding protein-2 (SREBP-2), bound to a sterol regulatory element (SRE), resulting in increased enzyme and, therefore, more cholesterol, synthesis; 2) degradation of the reductase is accelerated when cholesterol levels are high; 3) reductase activity is controlled by adenosine monophosphate (AMP)–activated protein kinase ([AMPK], which phosphorylates and inactivates the reductase) and an insulin-activated protein phosphatase (which dephosphorylates and activates it); and 4) expression of the gene for the reductase is upregulated by insulin and downregulated by glucagon. Statins are competitive inhibitors of HMG CoA reductase. These drugs are used to decrease plasma cholesterol in patients with hypercholesterolemia. The ring structure of cholesterol cannot be degraded in humans.
Cholesterol can be eliminated from the body either by conversion to bile salts or by secretion into the bile. Bile salts and phosphatidylcholine are quantitatively the most important organic components of bile. The rate-limiting step in bile acid synthesis is catalyzed by cholesterol-7-gα-hydroxylase, which is inhibited by bile acids. Before the bile acids leave the liver, they are conjugated to a molecule of either glycine or taurine, producing the conjugated bile salts glycocholic or taurocholic acid and glycochenodeoxycholic or taurochenodeoxycholic acid. Bile salts (deprotonated) are more amphipathic than bile acids (protonated) and, therefore, are more effective emulsifiers of dietary fat. In the intestine, bacteria can remove the glycine and taurine and can remove a hydroxyl group from the steroid nucleus, producing the secondary bile salts, deoxycholic and lithocholic acids. More than 95% of the bile salts are efficiently reabsorbed in the intestinal ileum by a sodium–bile salt cotransporter, returned to the blood, and carried by albumin back to the liver where they are taken up by the hepatic form of the cotransporter and reused (enterohepatic circulation, which bile acid sequestrants reduce). If more cholesterol enters the bile than can be solubilized by the available bile salts and phosphatidylcholine, cholesterol gallstone disease (cholelithiasis) can occur.
The plasma lipoproteins include chylomicrons, very-low-density lipoproteins (VLDLs), low-density lipoproteins (LDLs), and high-density lipoproteins (HDLs). They function to keep lipids (primarily triacyl-glycerol[TAG] and cholesteryl esters) soluble as they transport them between tissues. Lipoproteins are composed of a neutral lipid (TAG, cholesteryl esters, or both) core surrounded by a shell of amphipathic apolipoproteins, phospholipid, and unesterified cholesterol. Chylomicrons are assembled in intestinal mucosal cells from dietary lipids (primarily TAG). Each nascent chylomicron particle has one molecule of apolipoprotein (apo) B-48. They are released from the cells into the lymphatic system and travel to the blood, where they receive apo C-II and apo E from HDLs. Apo C-II activates endothelial lipoprotein lipase (LPL), which degrades the TAG in chylomicrons to fatty acids and glycerol. The fatty acids that are released are stored (in the adipose) or used for energy (by the muscle). The glycerol is metabolized by the liver. Patients with a deficiency of LPL or apo C-IIshow a dramatic accumulation of chylomicrons in the plasma (type I hyperlipoproteinemia, or familial LPL deficiency) even if fasted. After most of the TAG is removed, apo C-II is returned to the HDL, and the chylomicron remnant, carrying most of the dietary cholesterol, binds to a receptor on the liver that recognizes apo E. The particle is endocytosed, and its contents degraded by lysosomal enzymes. Defective uptake of these remnants causes type III hyperlipoproteinemia. Nascent VLDLs are produced in the liver and are composed predominantly of TAG. They contain a single molecule of apo B-100. Like nascent chylomicrons, VLDLs receive apo C-II and apo E from HDL in the plasma. The function of VLDL is to carry hepatic TAG to the peripheral tissues where LPL degrades the lipid. Additionally, the VLDL particle receives cholesteryl esters from HDL in exchange for TAG. This process is accomplished by cholesteryl ester transfer protein (CETP). Eventually, VLDL in the plasma is converted to LDL, a much smaller, denser particle. Apo C-II and apo E are returned to HDL, but the LDL retains apo B-100, which is recognized by receptors on peripheral tissues and the liver. LDLs undergo receptor-mediated endocytosis, and their contents are degraded in the lysosomes. A deficiency of functional LDL receptors causes type II hyperlipoproteinemia (familial hypercholesterolemia). The endocytosed cholesterol decreases synthesis of HMG CoA reductase (and of LDL receptors) through prevention of SREBP-2 binding to the SRE. Some of it can also be esterified by acyl CoA:cholesterol acyltransferase (ACAT) and stored. HDL are created by lipidation of apo A-1 synthesized in the liver and intestine. They have a number of functions, including: 1) serving as a circulating reservoir of apo C-II and apo E for chylomicrons and VLDL; 2) removing unesterified cholesterol from from peripheral tissues via ABCA1and esterifying it using lecithin:cholesterol acyl transferase(LCAT), a liver-synthesized plasma enzyme that is activated by apo A-1; and 3) delivering these cholesteryl esters to the liver (reverse cholesterol transport) for uptake via scavenger receptor-B1(SR-B1).
Cholesterol is the precursor of all classes of steroid hormones, which include glucocorticoids, mineralocorticoids, and the sex hormones (androgens, estrogens, and progestins). Synthesis, using primarily cytochrome P450 mixed-function oxidases, occurs in the adrenal cortex (cortisol, aldosterone, and androgens), ovaries and placenta (estrogens and progestins), and testes (testosterone). The initial and rate-limiting step is the conversion of cholesterol to pregnenolone by the side-chain cleavage enzyme P450scc. Deficiencies in synthesis Iead to congenital adrenal hyperplasia. Each steroid hormone diffuses across the plasma membrane of its target cell and binds to a specific cytosolic or nuclear receptor. These receptor–ligand complexes accumulate in the nucleus, dimerize, and bind to specific regulatory DNA sequences (hormone-response elements) in association with coactivator proteins, thereby causing promoter activation and increased transcription of targeted genes. In association with corepressors, transcription is decreased.
Figure 18.28 Activation of transcription by interaction of steroid hormone-receptor complex with hormone response element (HRE). The receptor contains domains that bind the hormone, DNA, and proteins that relax the DNA. mRNA = messenger RNA.
Study Questions
Choose the ONE best answer.
18.1 Mice were genetically engineered to contain hydroxymethylglutaryl coenzyme A reductase in which serine 871, a phosphorylation site, was replaced by alanine. Which of the following statements concerning the modified form of the enzyme is most likely to be correct?
A. The enzyme is nonresponsive to adenosine triphosphate depletion.
B. The enzyme is nonresponsive to statin drugs.
C. The enzyme is nonresponsive to the sterol response element–sterol response element–binding protein system.
D. The enzyme is unable to be degraded by the ubiquitin–proteasome system.
Correct answer = A. The reductase is regulated by covalent phosphorylation and dephosphorylation. Depletion of adenosine triphosphate results in a rise in adenosine monophosphate (AMP), which activates AMP kinase (AMPK), thereby phosphorylating and inactivating the enzyme. In the absence of the serine, a common phosphorylation site, the enzyme cannot be phosphorylated by AMPK. The enzyme is also regulated physiologically through changes in transcription and degradation and pharmacologically by statin drugs (competitive inhibitors), but none of these depends on serine phosphorylation.
Figure 18.29 Concept map for cholesterol and the lipoproteins. HMG CoA = hydroxymethylglutaryl coenzyme A; SREBP = sterol regulatory element-binding protein; HDL = high-density lipoprotein; VLDL = very-low-density lipoprotein; LDL = lowdensity lipoprotein; TAG = triacylglycerol; NADP(H) = nicotinamide adenine dinucleotide phosphate.
18.2 Calculate the amount of cholesterol in the low-density lipoproteins in an individual whose fasting blood gave the following lipid-panel test results: total cholesterol = 300 mg/dl, high-density lipoprotein cholesterol = 25 mg/dl, triglycerides = 150 mg/dl.
A. 55 mg/dl
B. 95 mg/dl
C. 125 mg/dl
D. 245 mg/dl
Correct answer = D. The total cholesterol in the blood of a fasted individual is equal to the sum of the cholesterol in low-density lipoproteins plus the cholesterol in high-density lipoproteins plus the cholesterol in very-low-density lipoproteins (VLDLs). This last term is calculated by dividing the triacylglycerol value by 5 because cholesterol accounts for about 1/5 of the volume of VLDL in fasted blood.
For Questions 18.3 and 18.4:
A young girl with a history of severe abdominal pain was taken to her local hospital at 5 a.m. in severe distress. Blood was drawn, and the plasma appeared milky, with the triacylglycerol level in excess of 2,000 mg/dl (normal = 4–150 mg/dl). The patient was placed on a diet extremely limited in fat but supplemented with medium-chain triglycerides.
Correct answer = A. The milky appearance of her blood was a result of triacylglycerol-rich chylomicrons. Because 5 a.m. is presumably several hours after her evening meal, the patient must have difficulty degrading these lipoprotein particles. Intermediate-, low-, and high-density lipoproteins contain primarily cholesteryl esters, and, if one or more of these particles was elevated, it would cause hypercholesterolemia. Very-low-density lipoproteins do not cause the described “milky appearance” in plasma.
18.3 Which of the following lipoprotein particles are most likely responsible for the appearance of the patient’s plasma?
A. Chylomicrons
B. High-density lipoproteins
C. Intermediate-density lipoproteins
D. Low-density lipoproteins
E. Very-low-density lipoproteins
Correct answer = C. The triacylglycerol (TAG) in chylomicrons is degraded by endothelial lipoprotein lipase, which requires apo C-II as a coenzyme. Deficiency of the enzyme or coenzyme results in decreased ability to degrade chylomicrons to their remnants, which get cleared by the liver. Apo A-I is the coenzyme for lecithin:cholesterol acyltransferase; apo B-48 is the ligand for the hepatic receptor that binds chylomicron remnants; cholesteryl ester transfer protein catalyzes the cholesteryl ester–TAG exchange between high-density and very-low-density lipoproteins (VLDLs); and microsomal triglyceride transfer protein is involved in the formation, not degradation, of chylomicrons (and VLDLs).
18.4 Which one of the following proteins is most likely to be deficient in this patient?
A. Apo A-I
B. Apo B-48
C. Apo C-II
D. Cholesteryl ester transfer protein
E. Microsomal triglyceride transfer protein
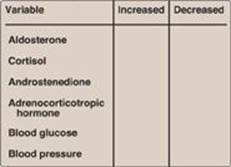
18.5 Complete the table below for an individual with classic 21-α-hydroxylase deficiency relative to a normal individual.
How might the results be changed if this individual were deficient in 17-α-hydroxylase, rather than 21-α-hydroxylase?
21-α-Hydroxylase deficiency causes mineralocorticoids (aldosterone) and glucocorticoids (cortisol) to be virtually absent. Because aldosterone increases blood pressure, and cortisol increases blood glucose, their deficiencies result in a decrease in blood pressure and blood glucose, respectively. Cortisol normally feeds back to inhibit adrenocorticotropic hormone (ACTH) release by the pituitary, and, so, its absence results in an elevation in ACTH. The loss of 21-α-hydroxylase pushes progesterone and pregnenolone to androgen synthesis, therefore, causes androstenedione levels to rise.
With 17-α-hydroxylase deficiency, sex hormone synthesis would be inhibited. Mineralocorticoid production would be increased, leading to hypertension.