Lippincott’s Illustrated Reviews: Biochemistr, Sixth Edition (2014)
UNIT IV: Nitrogen Metabolism
Chapter 21. Conversion of Amino Acids to Specialized Products
I. OVERVIEW
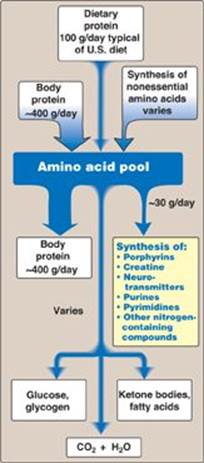
In addition to serving as building blocks for proteins, amino acids are precursors of many nitrogen-containing compounds that have important physiologic functions (Figure 21.1). These molecules include porphyrins, neurotransmitters, hormones, purines, and pyrimidines.
II. PORPHYRIN METABOLISM
Porphyrins are cyclic compounds that readily bind metal ions, usually ferrous (Fe2+) or ferric (Fe3+) iron. The most prevalent metalloporphyrin in humans is heme, which consists of one Fe2+ coordinated in the center of the tetrapyrrole ring of protoporphyrin IX (see p. 280). Heme is the prosthetic group for hemoglobin, myoglobin, the cytochromes, the cytochrome P450 (CYP) monooxygenase system, catalase, nitric oxide synthase, and peroxidase. These hemeproteins are rapidly synthesized and degraded. For example, 6–7 g of hemoglobin are synthesized each day to replace heme lost through the normal turnover of erythrocytes. The simultaneous synthesis and degradation of the associated porphyrins and recycling of the bound iron ions is coordinated with the turnover of hemeproteins.
Figure 21.1 Amino acids as precursors of nitrogen-containing compounds.
A. Structure of porphyrins
Porphyrins are cyclic molecules formed by the linkage of four pyrrole rings through methenyl bridges (Figure 21.2). Three structural features of these molecules are relevant to understanding their medical significance.
1. Side chains: Different porphyrins vary in the nature of the side chains that are attached to each of the four pyrrole rings. Uroporphyrin contains acetate (–CH2–COO–) and propionate (–CH2–CH2–COO–) side chains; coproporphyrin contains methyl (–CH3) and propionate groups; and protoporphyrin IX (and heme) contains vinyl (–CH=CH2), methyl, and propionate groups.
Figure 21.2 Structures of uroporphyrin I and uroporphyrin III.

2. Distribution of side chains: The side chains of porphyrins can be ordered around the tetrapyrrole nucleus in four different ways, designated by Roman numerals I to IV. Only type III porphyrins, which contain an asymmetric substitution on ring D (see Figure 21.2), are physiologically important in humans. [Note: Protoporphyrin IX is a member of the type III series.]
3. Porphyrinogens: These porphyrin precursors (for example, uroporphyrinogen) exist in a chemically reduced, colorless form and serve as intermediates between porphobilinogen and the oxidized, colored protoporphyrins in heme biosynthesis.
B. Biosynthesis of heme
The major sites of heme biosynthesis are the liver, which synthesizes a number of heme proteins (particularly the CYP proteins), and the erythrocyte-producing cells of the bone marrow, which are active in hemoglobin synthesis. [Note: Over 85% of all heme synthesis occurs in erythroid tissue.] In the liver, the rate of heme synthesis is highly variable, responding to alterations in the cellular heme pool caused by fluctuating demands for heme proteins. In contrast, heme synthesis in erythroid cells is relatively constant and is matched to the rate of globin synthesis. The initial reaction and the last three steps in the formation of porphyrins occur in mitochondria, whereas the intermediate steps of the biosynthetic pathway occur in the cytosol (see Figure 21.8). [Note: Mature red blood cells (RBCs) lack mitochondria and are unable to synthesize heme.]
Figure 21.3 Pathway of porphyrin synthesis: Formation of porphobilinogen. ALAS = δ-aminolevulinic acid synthase; CoA = coenzyme A; PLP = pyridoxal phosphate. (Continued in Figures 21.4 and 21.5.)
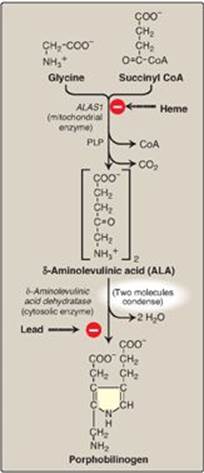
1. Formation of δ-aminolevulinic acid: All the carbon and nitrogen atoms of the porphyrin molecule are provided by glycine (a nonessential amino acid) and succinyl coenzyme A (a citric acid cycle intermediate) that condense to form δ-aminolevulinic acid (ALA) in a reaction catalyzed by ALA synthase ([ALAS] Figure 21.3) This reaction requires pyridoxal phosphate (PLP) as a coenzyme and is the committed and rate-limiting step in porphyrin biosynthesis. [Note: There are two isoforms of ALAS, 1 and 2, each produced by different genes and controlled by different mechanisms. ALAS1 is found in all tissues, whereas ALAS2 is erythroid specific. Loss-of-function mutations in ALAS2 result in X-linked sideroblastic anemia.]
a. Effect of heme (hemin): When porphyrin production exceeds the availability of the apoproteins that require it, heme accumulates and is converted to hemin by the oxidation of Fe2+ to Fe3+. Hemin decreases the amount (and thereby the activity) of ALAS1 by repressing transcription of its gene, increasing degradation of its messenger RNA, and decreasing import of the enzyme into mitochondria. [Note: In erythroid cells, ALAS2 is controlled by the availability of intracellular iron.]
b. Effect of drugs: Administration of any of a large number of drugs results in a significant increase in hepatic ALAS1 activity. These drugs are metabolized by the microsomal CYP monooxygenase system, a hemeprotein oxidase system found in the liver (see p. 149). In response to these drugs, the synthesis of CYP proteins increases, leading to an enhanced consumption of heme, a component of these proteins. This, in turn, causes a decrease in the concentration of heme in liver cells. The lower intracellular heme concentration leads to an increase in the synthesis of ALAS1 and prompts a corresponding increase in the synthesis of ALA.
2. Formation of porphobilinogen: The condensation of two molecules of ALA to form porphobilinogen by zinc-containing ALA dehydratase (porphobilinogen synthase) is extremely sensitive to inhibition by heavy metal ions (for example, lead) that replace the zinc (see Figure 21.3). This inhibition is, in part, responsible for the elevation in ALA and the anemia seen in lead poisoning.
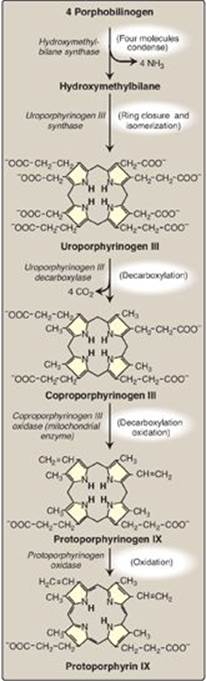
3. Formation of uroporphyrinogen: The condensation of four porphobilinogens produces the linear tetrapyrrole hydroxymethylbilane, which is cyclized and isomerized by uroporphyrinogen III synthase to produce the asymmetric uroporphyrinogen III. This cyclic tetrapyrrole undergoes decarboxylation of its acetate groups, generating coproporphyrinogen III (Figure 21.4). These reactions occur in the cytosol.
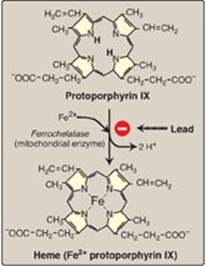
4. Formation of heme: Coproporphyrinogen III enters the mitochondrion, and two propionate side chains are decarboxylated by coproporphyrinogen III oxidase to vinyl groups generating protoporphyrinogen IX, which is oxidized to protoporphyrin IX. The introduction of iron (as Fe2+) into protoporphyrin IX can occur spontaneously, but the rate is enhanced by ferrochelatase, an enzyme that, like ALA dehydratase, is inhibited by lead (Figure 21.5).
Figure 21.4 Pathway of porphyrin synthesis: formation of protoporphyrin IX. (Continued from Figure 21.3.) [Note: Deficiency in uroporphyrinogen III synthase prevents isomerization, resulting in production of type I porphyrins.]
C. Porphyrias
Porphyrias are rare, inherited (or occasionally acquired) defects in heme synthesis, resulting in the accumulation and increased excretion of porphyrins or porphyrin precursors (see Figure 21.8). [Note: With few exceptions, porphyrias are inherited as autosomal-dominant disorders.] Each porphyria results in the accumulation of a unique pattern of intermediates caused by the deficiency of an enzyme in the heme synthetic pathway. [Note: “Porphyria” refers to the red-blue color caused by pigment-like porphyrins in the urine of some patients with defects in heme synthesis.]
1. Clinical manifestations: The porphyrias are classified as erythropoietic or hepatic, depending on whether the enzyme deficiency occurs in the erythropoietic cells of the bone marrow or in the liver. Hepatic porphyrias can be further classified as chronic or acute. In general, individuals with an enzyme defect prior to the synthesis of the tetrapyrroles manifest abdominal and neuropsychiatric signs, whereas those with enzyme defects leading to the accumulation of tetrapyrrole intermediates show photosensitivity (that is, their skin itches and burns [pruritus] when exposed to visible light). [Note: Photosenstivity is a result of the oxidation of colorless porphyrinogens to colored porphyrins, which are photosensitizing molecules thought to participate in the formation of superoxide radicals from oxygen. These reactive oxygen species can oxidatively damage membranes and cause the release of destructive enzymes from lysosomes.]
Figure 21.5 Pathway of porphyrin synthesis: formation of heme. (Continued from Figures 21.3 and 21.4.) Fe2+ = ferrous iron.
a. Chronic hepatic porphyria: Porphyria cutanea tarda, the most common porphyria, is a chronic disease of the liver. The disease is associated with a deficiency in uroporphyrinogen decarboxylase, but clinical expression of the enzyme deficiency is influenced by various factors, such as hepatic iron overload, exposure to sunlight, alcohol ingestion, estrogen therapy, and the presence of hepatitis B or C or HIV infections. Clinical onset is typically during the fourth or fifth decade of life. Porphyrin accumulation leads to cutaneous symptoms (Figure 21.6) as well as urine that is red to brown in natural light (Figure 21.7) and pink to red in fluorescent light.
Figure 21.6 Skin eruptions in a patient with porphyria cutanea tarda.
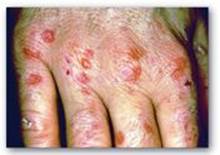
b. Acute hepatic porphyrias: Acute hepatic porphyrias (ALA dehydratase deficiency porphyria, acute intermittent porphyria, hereditary coproporphyria, and variegate porphyria) are characterized by acute attacks of gastrointestinal (GI), neuropsychiatric, and motor symptoms that may be accompanied by photosensitivity. Porphyrias leading to accumulation of ALA and porphobilinogen, such as acute intermittent porphyria, cause abdominal pain and neuropsychiatric disturbances, ranging from anxiety to delirium. Symptoms of the acute hepatic porphyrias are often precipitated by use of drugs, such as barbiturates and ethanol, which induce the synthesis of the heme-containing CYP microsomal drug oxidation system. This further decreases the amount of available heme, which, in turn, promotes increased synthesis of ALAS1.
c. Erythropoietic porphyrias: The chronic erythropoietic porphyrias (congenital erythropoietic porphyria and erythropoietic protoporphyria) are characterized by skin rashes and blisters that appear in early childhood. [Note: Patients with erythropoietic protoporphyria are also at risk for hepatobiliary disease.]
2. Increased δ-aminolevulinic acid synthase activity: One common feature of the porphyrias is a decreased synthesis of heme. In the liver, heme normally functions as a repressor of the gene for ALAS1. Therefore, the absence of this end product results in an increase in the synthesis of ALAS1 (derepression). This causes an increased synthesis of intermediates that occur prior to the genetic block. The accumulation of these toxic intermediates is the major pathophysiology of the porphyrias.
Figure 21.7 Urine from a patient with porphyria cutanea tarda (right) and from a patient with normal porphyrin excretion (left).

3. Treatment: During acute porphyria attacks, patients require medical support, particularly treatment for pain and vomiting. The severity of acute symptoms of the porphyrias can be diminished by intravenous injection of hemin and glucose, which decreases the synthesis of ALAS1. Protection from sunlight, ingestion of β-carotene (a free-radical scavenger), and phlebotomy are helpful in porphyrias with photosensitivity.
Figure 21.8 Summary of heme synthesis. 1Also referred to as porphobilinogen synthase. 2Also referred to as porphobilinogen deaminase. [Note: Deficiencies in ALA synthase-1 (ALAS1) are unknown. Deficiencies in ALAS2 result in an anemia, not a porphyria.]
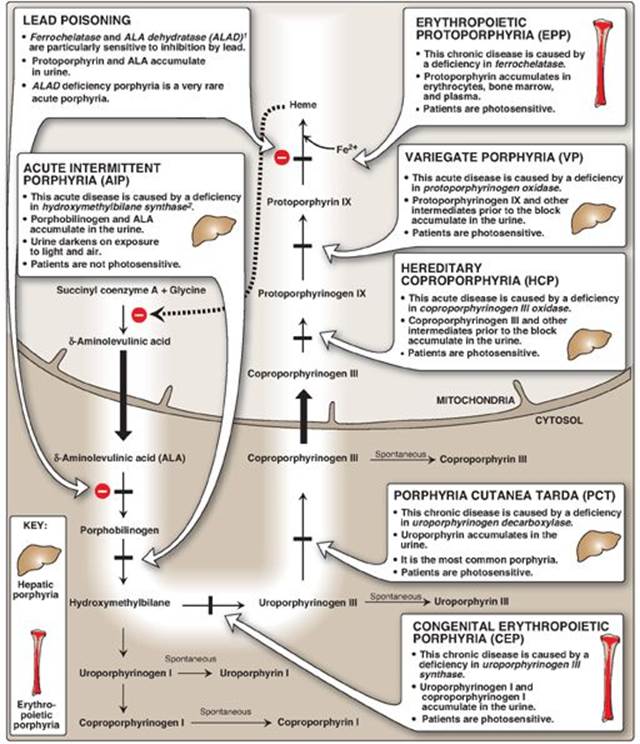
D. Degradation of heme
After approximately 120 days in the circulation, red blood cells are taken up and degraded by the reticuloendothelial system, particularly in the liver and spleen (Figure 21.9). Approximately 85% of heme destined for degradation comes from senescent RBCs. The remainder is from the degradation of heme proteins other than hemoglobin.
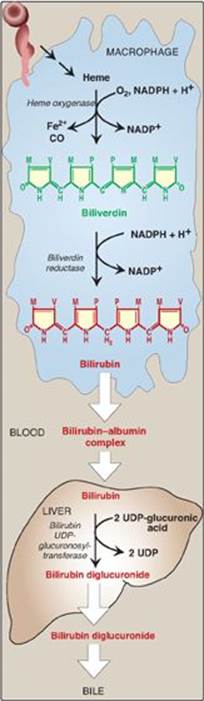
1. Formation of bilirubin: The first step in the degradation of heme is catalyzed by the microsomal heme oxygenase system of the reticuloendothelial cells. In the presence of nicotinamide adenine dinucleotide phosphate and O2, the enzyme catalyzes three successive oxygenations that result in opening of the porphyrin ring (converting cyclic heme to linear biliverdin), production of carbon monoxide (CO), and release of Fe2+ (see Figure 21.9). [Note: The CO has biologic function, acting as a signaling molecule and anti-inflammatory.] Biliverdin, a green pigment, is reduced, forming the red-orange bilirubin. Bilirubin and its derivatives are collectively termed bile pigments. [Note: The changing colors of a bruise reflect the varying pattern of intermediates that occurs during heme degradation.]
Bilirubin, unique to mammals, appears to function at low levels as an antioxidant. In this role, it is oxidized to biliverdin, which is then reduced by biliverdin reductase, regenerating bilirubin.
2. Uptake of bilirubin by the liver: Bilirubin is only slightly soluble in plasma and, therefore, is transported to the liver by binding noncovalently to albumin. [Note: Certain anionic drugs, such as salicylates and sulfonamides, can displace bilirubin from albumin, permitting bilirubin to enter the central nervous system (CNS). This causes the potential for neural damage in infants (see p. 285).] Bilirubin dissociates from the carrier albumin molecule; enters a hepatocyte via facilitated diffusion; and binds to intracellular proteins, particularly the protein ligandin.
3. Formation of bilirubin diglucuronide: In the hepatocyte, the solubility of bilirubin is increased by the addition of two molecules of glucuronic acid, producing bilirubin diglucuronide. [Note: This process is referred to as conjugation.] The reaction is catalyzed by microsomal bilirubin UDP-glucuronosyltransferase (bilirubin UGT) using uridine diphosphate (UDP)-glucuronic acid as the glucuronate donor. [Note: Varying degrees of deficiency of bilirubin UGT result in Crigler-Najjar I and II and Gilbert syndrome, with Crigler-Najjar I being the most severe deficiency.]
Figure 21.9 Formation of bilirubin from heme and its conversion to bilirubin diglucuronide. UDP = uridine diphosphate; CO = carbon monoxide; NADP(H) = nicotinamide adenine dinucleotide phosphate.
4. Secretion of bilirubin into bile: Bilirubin diglucuronide (conjugated bilirubin [CB]) is actively transported against a concentration gradient into the bile canaliculi and then into the bile. This energy-dependent, rate-limiting step is susceptible to impairment in liver disease. [Note: A rare deficiency in the protein required for transport of CB out of the liver results in Dubin-Johnson syndrome.] Unconjugated bilirubin (UCB) is normally not secreted into bile.
Figure 21.10 Catabolism of heme ![]() = bilirubin;
= bilirubin; ![]() = conjugated bilirubin;
= conjugated bilirubin; ![]() = urobilinogen;
= urobilinogen; ![]() = urobilin;
= urobilin; ![]() = stercobilin.
= stercobilin.
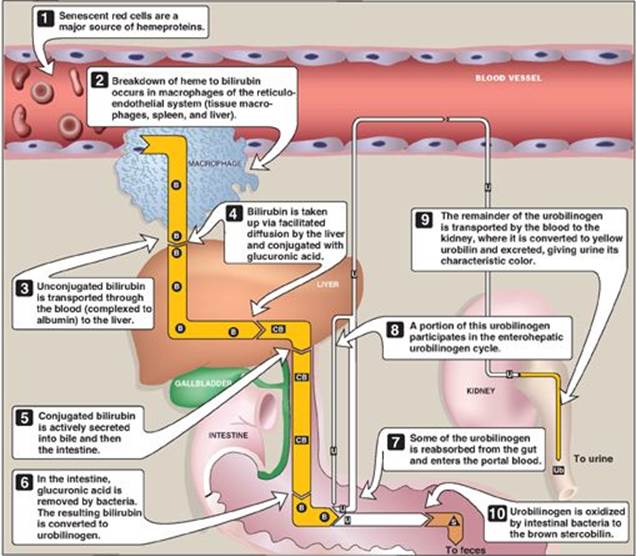
5. Formation of urobilins in the intestine: Bilirubin diglucuronide is hydrolyzed and reduced by bacteria in the gut to yield urobilinogen, a colorless compound. Most of the urobilinogen is oxidized by intestinal bacteria to stercobilin, which gives feces the characteristic brown color. However, some of the urobilinogen is reabsorbed from the gut and enters the portal blood. A portion of this urobilinogen participates in the enterohepatic urobilinogen cycle in which it is taken up by the liver and then resecreted into the bile. The remainder of the urobilinogen is transported by the blood to the kidney, where it is converted to yellow urobilin and excreted, giving urine its characteristic color. The metabolism of bilirubin is summarized in Figure 21.10.
E. Jaundice
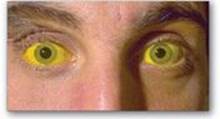
Jaundice (also called icterus) refers to the yellow color of skin, nail beds, and sclerae (whites of the eyes) caused by deposition of bilirubin, secondary to increased bilirubin levels in the blood (hyper-bilirubinemia) as shown in Figure 21.11. Although not a disease, jaundice is usually a symptom of an underlying disorder. [Note: Blood bilirubin levels are normally about 1 mg/dl. Jaundice is seen at 2–3 mg/dl.]
Figure 21.11 Jaundiced patient, with the sclerae of his eyes appearing yellow.
1. Types of jaundice: Jaundice can be classified into three major types described below. However, in clinical practice, jaundice is often more complex than indicated in this simple classification. For example, the accumulation of bilirubin may be a result of defects at more than one step in its metabolism.
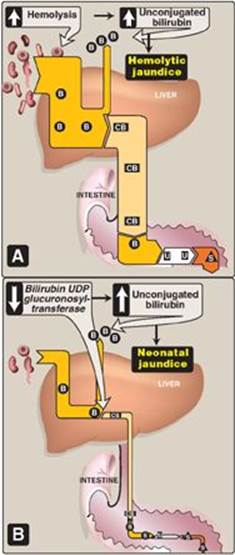
a. Hemolytic jaundice: The liver has the capacity to conjugate and excrete over 3,000 mg of bilirubin per day, whereas the normal production of bilirubin is only 300 mg/day. This excess capacity allows the liver to respond to increased heme degradation with a corresponding increase in conjugation and secretion of bilirubin diglucuronide. However, extensive hemolysis (for example, in patients with sickle cell anemia or pyruvate kinase or glucose 6-phosphate dehydrogenase deficiency) may produce bilirubin faster than it can be conjugated. UCB levels in the blood become elevated, causing jaundice (Figure 21.12A). [Note: With hemolysis, more CB is made and excreted into the bile, the amount of urobilinogen entering the enterohepatic circulation is increased, and urinary urobilinogen is increased.]
b. Hepatocellular jaundice: Damage to liver cells (for example, in patients with cirrhosis or hepatitis) can cause UCB levels in the blood to increase as a result of decreased conjugation. Urobilinogen is increased in the urine because hepatic damage decreases the enterohepatic circulation of this compound, allowing more to enter the blood, from which it is filtered into the urine. The urine consequently darkens, whereas stools may be a pale, clay color. Plasma levels of alanine and aspartate transaminases (AST and ALT, respectively; see p. 251) are elevated. [Note: If CB is made but is not efficiently secreted from the liver into bile (intrahepatic cholestasis), it can diffuse (“leak”) into the blood, causing a conjugated hyperbilirubinemia.]
Figure 21.12 Alterations in the metabolism of heme. A. Hemolytic jaundice. B. Neonatal jaundice. ![]() = conjugated bilirubin;
= conjugated bilirubin; ![]() = bilirubin;
= bilirubin; ![]() = urobilinogen;
= urobilinogen; ![]() = stercobilin; UDP = uridine diphosphate
= stercobilin; UDP = uridine diphosphate
c. Obstructive jaundice: In this instance, jaundice is not caused by overproduction of bilirubin or decreased conjugation but, instead, results from obstruction of the common bile duct (extrahepatic cholestasis). For example, the presence of a tumor or bile stones may block the duct, preventing passage of CB into the intestine. Patients with obstructive jaundice experience GI pain and nausea and produce stools that are a pale, clay color. The liver “regurgitates” CB into the blood (hyperbilirubinemia). The CB is eventually excreted in the urine (which darkens upon standing), and is referred to as “urinary bilirubin.” Urinary urobilinogen is absent.
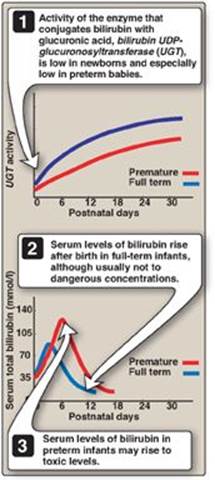
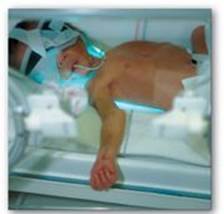
2. Jaundice in newborns: The majority of newborn infants (60% of full term and 80% of preterm) show a rise in UCB in the first postnatal week (and a transient, “physiologic” jaundice) because the activity of hepatic bilirubin UGT is low at birth (it reaches adult levels in about 4 weeks) as shown in Figures 21.12B and 21.13. Elevated UCB, in excess of the binding capacity of albumin (20–25 mg/dl), can diffuse into the basal ganglia, cause toxic encephalopathy (kernicterus) and a pathologic jaundice. Therefore, newborns with significantly elevated bilirubin levels are treated with blue fluorescent light (phototherapy), as shown in Figure 21.14, which converts bilirubin to more polar and, therefore, water-soluble isomers. These photoisomers can be excreted into the bile without conjugation to glucuronic acid. [Note: Because of solubility differences, only UCB crosses the blood brain barrier, and only CB appears in urine.]
3. Determination of bilirubin concentration: Bilirubin is commonly measured by the van den Bergh reaction, in which diazotized sulfanilic acid reacts with bilirubin to form red azodipyrroles that are measured colorimetrically. In aqueous solution, the water-soluble CB reacts rapidly with the reagent (within one minute), and is said to be “direct-reacting.” The UCB, which is much less soluble in aqueous solution, reacts more slowly. However, when the reaction is carried out in methanol, both CB and UCB are soluble and react with the reagent, providing the total bilirubin value. The “indirect-reacting” bilirubin, which corresponds to the UCB, is obtained by subtracting the direct-reacting bilirubin from the total bilirubin. [Note: In normal plasma, only about 4% of the total bilirubin is conjugated or direct-reacting, because most is secreted into bile.]
Figure 21.13 Neonatal jaundice. UDP = uridine diphosphate.
III. OTHER NITROGEN-CONTAINING COMPOUNDS
A. Catecholamines
Dopamine, norepinephrine, and epinephrine are biologically active (biogenic) amines that are collectively termed catecholamines. Dopamine and norepinephrine are synthesized in the brain and function as neurotransmitters. Norepinephrine is also synthesized in the adrenal medulla, as is epinephrine.
1. Function: Outside the CNS, norepinephrine and its methylated derivative, epinephrine, are hormone regulators of carbohydrate and lipid metabolism. Norepinephrine and epinephrine are released from storage vesicles in the adrenal medulla in response to fright, exercise, cold, and low levels of blood glucose. They increase the degradation of glycogen and triacylglycerol as well as increase blood pressure and the output of the heart. These effects are part of a coordinated response to prepare the individual for stress, and are often called the “fight-or-flight” reactions.
Figure 21.14 Phototherapy in neonatal jaudice.
2. Synthesis: The catecholamines are synthesized from tyrosine, as shown in Figure 21.15. Tyrosine is first hydroxylated by tyrosine hydroxylase to form 3,4-dihydroxyphenylalanine (DOPA) in a reaction analogous to that described for the hydroxylation of phenylalanine (see p. 270). The tetrahydrobiopterin (BH4)-requiring enzyme is abundant in the CNS, the sympathetic ganglia, and the adrenal medulla and is the rate-limiting step of the pathway. DOPA is decarboxylated in a reaction requiring PLP (see p. 378) to form dopamine, which is hydroxylated by dopamine β-hydroxylase to yield norepinephrine in a reaction that requires ascorbate (vitamin C) and copper. Epinephrine is formed from norepinephrine by an N-methylation reaction using S-adenosylmethionine (SAM) as the methyl donor (see p. 264).
Figure 21.15 Synthesis of catecholamines. PLP = pyridoxal phosphate; Cu2+ = copper.
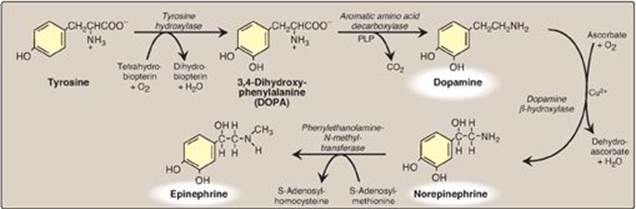
Parkinson disease, a neurodegenerative movement disorder, is due to insufficient dopamine production as a result of the idiopathic loss of dopamine-producing cells in the brain. Administration of L-DOPA (levodopa) is the most common treatment. Dopamine cannot cross the blood brain barrier.
3. Degradation: The catecholamines are inactivated by oxidative deamination catalyzed by monoamine oxidase (MAO) and by O-methylation carried out by catechol-O-methyltransferase (COMT) using SAM as the methyl donor (Figure 21.16). The two reactions can occur in either order. The aldehyde products of the MAO reaction are oxidized to the corresponding acids. The metabolic products of these reactions are excreted in the urine as vanillylmandelic acid (VMA) from epinephrine and norepinephrine and homovanillic acid from dopamine. [Note: VMA is increased with pheochromocytomas, rare tumors of the adrenal gland characterized by excessive production of catecholamines.]
Figure 21.16 Metabolism of the catecholamines by catechol-O-methyltranferase (COMT) and monoamine oxidase (MAO).
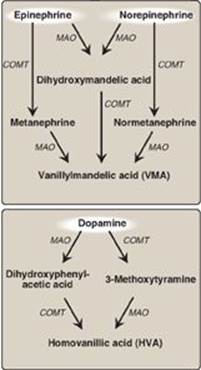
4. Monoamine oxidase inhibitors: MAO is found in neural and other tissues, such as the intestine and liver. In the neuron, this enzyme oxidatively deaminates and inactivates any excess neurotransmitter molecules (norepinephrine, dopamine, or serotonin) that may leak out of synaptic vesicles when the neuron is at rest. MAO inhibitors may irreversibly or reversibly inactivate the enzyme, permitting neurotransmitter molecules to escape degradation and, therefore, to both accumulate within the presynaptic neuron and to leak into the synaptic space. This causes activation of norepinephrine and serotonin receptors and may be responsible for the antidepressant action of these drugs.
B. Histamine
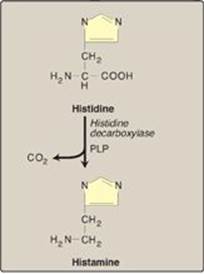
Histamine is a chemical messenger that mediates a wide range of cellular responses, including allergic and inflammatory reactions and gastric acid secretion. A powerful vasodilator, histamine is formed by decarboxylation of histidine in a reaction requiring PLP (Figure 21.17). It is secreted by mast cells as a result of allergic reactions or trauma. Histamine has no clinical applications, but agents that interfere with the action of histamine have important therapeutic applications.
Figure 21.17 Biosynthesis of histamine. PLP = pyridoxal phosphate.
C. Serotonin
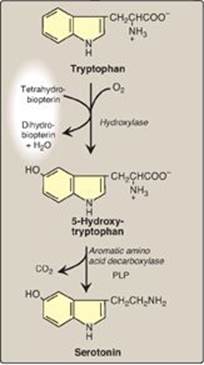
Serotonin, also called 5-hydroxytryptamine (5-HT), is synthesized and stored at several sites in the body (Figure 21.18). The largest amount by far is found in the intestinal mucosa. Smaller amounts occur in the CNS, where it functions as a neurotransmitter, and in platelets. Serotonin is synthesized from tryptophan, which is hydroxylated in a BH4-requiring reaction analogous to that catalyzed by phenylalanine hydroxylase. The product, 5-hydroxytryptophan, is decarboxylated to serotonin, which is degraded by MAO. Serotonin has multiple physiologic roles including pain perception, regulation of sleep, appetite, temperature, blood pressure, cognitive functions, and mood (causes a feeling of well-being). [Note: Selective serotonin reuptake inhibitors (SSRIs) maintain serotonin levels, thereby functioning as antidepressants.]
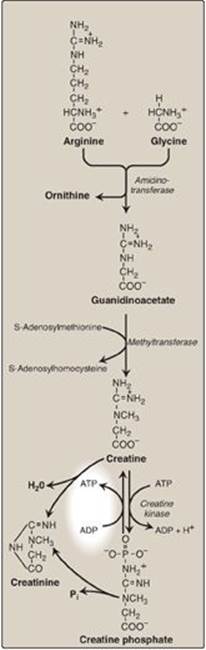
D. Creatine
Creatine phosphate (also called phosphocreatine), the phosphorylated derivative of creatine found in muscle, is a high-energy compound that provides a small but rapidly mobilized reserve of high-energy phosphates that can be reversibly transferred to adenosine diphosphate (Figure 21.19) to maintain the intracellular level of adenosine triphosphate (ATP) during the first few minutes of intense muscular contraction. [Note: The amount of creatine phosphate in the body is proportional to the muscle mass.]
1. Synthesis: Creatine is synthesized in liver and kidney tissue from glycine and the guanidino group of arginine, plus a methyl group from SAM (see Figure 21.19). Animal products are dietary sources. Creatine is reversibly phosphorylated to creatine phosphate by creatine kinase, using ATP as the phosphate donor. [Note: The presence of creatine kinase (MB isozyme) in the plasma is indicative of heart damage and is used in the diagnosis of myocardial infarction (see p. 65).]
Figure 21.18 Synthesis of serotonin. [Note: Serotonin is converted to melatonin in the pineal gland.] PLP = pyridoxal phosphate.
2. Degradation: Creatine and creatine phosphate spontaneously cyclize at a slow but constant rate to form creatinine, which is excreted in the urine. The amount excreted is proportional to the total creatine phosphate content of the body and, therefore, can be used to estimate muscle mass. When muscle mass decreases for any reason (for example, from paralysis or muscular dystrophy), the creatinine content of the urine falls. In addition, any rise in blood creatinine is a sensitive indicator of kidney malfunction, because creatinine normally is rapidly removed from the blood and excreted. A typical adult male excretes about 1–2 g of creatinine per day.
E. Melanin
Melanin is a pigment that occurs in several tissues, particularly the eye, hair, and skin. It is synthesized from tyrosine in melanocytes (pigment-forming cells) of the epidermis. It functions to protect underlying cells from the harmful effects of sunlight. [Note: A defect in melanin production results in oculocutaneous albinism, the most common type being due to defects in copper-containing tyrosinase (see p. 273).]
Figure 21.19 Synthesis of creatine. ADP = adenosine diphosphate; Pi = inorganic phosphate.
IV. CHAPTER SUMMARY
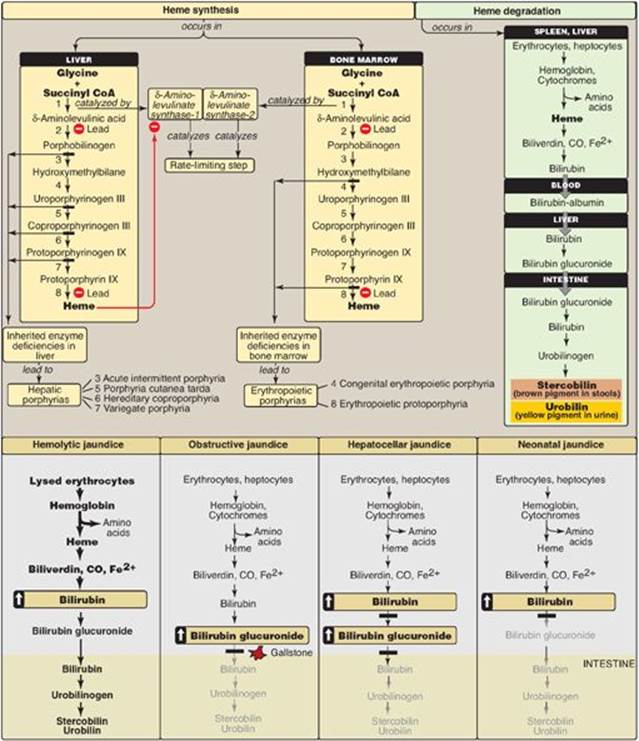
Amino acids are precursors of many nitrogen-containing compounds including porphyrins, which, in combination with ferrous (Fe2+) iron, form heme (Figure 21.20). The major sites of heme biosynthesis are the liver, which synthesizes a number of heme proteins (particularly cytochrome P450 enzymes), and the erythrocyte-producing cells of the bone marrow, which are active in hemoglobin synthesis. In the liver, the rate of heme synthesis is highly variable, responding to alterations in the cellular heme pool caused by fluctuating demands for hemeproteins. In contrast, heme synthesis in erythroid cells is relatively constant and is matched to the rate of globin synthesis. Porphyrin synthesis start with glycine and succinyl coenzyme A. The committed step in heme synthesis is the formation of δ-aminolevulinic acid (ALA). This reaction is catalyzed by ALA synthase-1 (ALAS1) in liver (inhibited by hemin, the oxidized form of heme that accumulates in the cell when heme is being underutilized) and ALAS2 in erythroid tissues (regulated by iron). Porphyrias are caused by inherited (primarily autosomal-dominant) or acquired defects in heme synthesis, resulting in the accumulation and increased excretion of porphyrins or porphyrin precursors. Enzymic defects early in the pathway cause abdominal pain and neuropsychiatric symptoms, whereas later defects cause photosensitivity. Degradation of hemeproteins occurs in the reticuloendothelial system, particularly in the liver and spleen. The first step in the degradation of heme is the production by heme oxygenase of the green pigment biliverdin, which is subsequently reduced to bilirubin. Bilirubin is transported to the liver, where its solubility is increased by the addition of two molecules of glucuronic acid. Bilirubin diglucuronide is transported into the bile canaliculi, where it is first hydrolyzed and reduced by bacteria in the gut to yield urobilinogen, then oxidized by intestinal bacteria to stercobilin. Jaundicerefers to the yellow color of the skin and sclerae that is caused by deposition of bilirubin, secondary to increased bilirubin levels in the blood. Three commonly encountered type of jaundice are hemolytic jaundice, obstructive jaundice, and hepatocellular jaundice. Other important N-containing compounds derived from amino acids include the catecholamines (dopamine, norepinephrine, and epinephrine), creatine, histamine, serotonin, and melanin.
Figure 21.20 Key concept map for heme metabolism. ![]() = Block in the pathway. [Note: Hepatocellular jaundice can be caused by decreased conjugation of bilirubin or decreased secretion of conjugated bilirubin into bile.] CoA = coenzyme A; CO = carbon monoxide.
= Block in the pathway. [Note: Hepatocellular jaundice can be caused by decreased conjugation of bilirubin or decreased secretion of conjugated bilirubin into bile.] CoA = coenzyme A; CO = carbon monoxide.
Study Questions
Choose the ONE best answer.
21.1 δ-Aminolevulinic acid synthase activity:
A. catalyzes the committed step in porphyrin biosynthesis.
B. is decreased by iron in erythrocytes.
C. is decreased in liver in individuals treated with certain drugs such as the barbiturate phenobarbital.
D. occurs in the cytosol.
E. requires biotin as a coenzyme.
Correct answer = A. δ-Aminolevulinic acid synthase is cytosolic and catalyzes the rate-limiting and regulated step of porphyrin synthesis. It requires pyridoxal phosphate as a coenzyme. Iron increases production of the erythroid isozyme. The hepatic isozyme is increased in patients treated with certain drugs.
21.2 A 50-year-old man presented with painful blisters on the backs of his hands. He was a golf instructor and indicated that the blisters had erupted shortly after the golfing season began. He did not have recent exposure to common skin irritants. He had partial complex seizure disorder that had begun about 3 years earlier after a head injury. The patient had been taking phenytoin (his only medication) since the onset of the seizure disorder. He admitted to an average weekly ethanol intake of about 18 12-oz cans of beer. The patient’s urine was reddish orange. Cultures obtained from skin lesions failed to grow organisms. A 24-hour urine collection showed elevated uroporphyrin (1,000 mg; normal, <27mg). The most likely diagnosis is:
A. acute intermittent porphyria.
B. congenital erythropoietic porphyria.
C. erythropoietic protoporphyria.
D. hereditary coproporphyria.
E. porphyria cutanea tarda.
Correct answer = E. The disease is associated with a deficiency in uroporphyrinogen decarboxylase, but clinical expression of the enzyme deficiency is influenced by hepatic injury caused by environmental (for example, ethanol) and infectious (for example, hepatitis B virus) agents. Exposure to sunlight can also be a precipitating factor. Clinical onset is typically during the fourth or fifth decade of life. Porphyrin accumulation leads to cutaneous symptoms and urine that is red to brown. Treatment of the patient’s seizure disorder with phenytoin caused increased synthesis of δ-aminolevulinic acid synthase and, therefore, of uroporphyrinogen, the substrate of the deficient enzyme. The laboratory and clinical findings are inconsistent with other porphyrias.
21.3 A patient presents with jaundice, abdominal pain, and nausea. Clinical laboratory studies give the following results:
![]()
What is the most likely cause of the jaundice?
A. Decreased hepatic conjugation of bilirubin
B. Decreased hepatic uptake of bilirubin
C. Decreased secretion of bile into the intestine
D. Increased hemolysis
Correct answer = C. The data are consistent with an obstructive jaundice in which a block in the common bile duct decreases the secretion of bile containing conjugated bilirubin (CB) into the intestine (stool will be pale in color). The liver “regurgitates” the CB into the blood (hyperbilirubinemia). The CB is excreted in the urine (which darkens) and is referred to as “urinary bilirubin.” Urinary urobilinogen is not present because its source is intestinal urobilinogen, which is low. The other choices do not match the data.
21.4 A 2-year-old child was brought to his pediatrician for evaluation of gastrointestinal problems. The parents report that the boy has been listless for the last few weeks. Lab tests reveal a microcytic, hypochromic anemia. Blood lead levels are elevated. Which of the enzymes listed below is most likely to have higher-than-normal activity in the liver of this child?
A. δ-Aminolevulinic acid synthase
B. Bilirubiun UDP-glucuronosyltransferase
C. Ferrochelatase
D. Heme oxygenase
E. Porphobilinogen synthase
Correct answer = A. This child has the acquired porphyria of lead poisoning. Lead inhibits δ-aminolevulinic acid dehydratase and, consequently, heme synthesis. The decrease in heme derepresses δ-aminolevulinic acid synthase-1 (the hepatic isozyme), resulting in an increase in its activity. The decrease in heme also results in decreased hemoglobin synthesis, and anemia is seen. Ferrochelatase is directly inhibited by lead. The other choices are enzymes of heme degradation.