Harper’s Illustrated Biochemistry, 29th Edition (2012)
SECTION II. Bioenergetics & the Metabolism of Carbohydrates & Lipids
Chapter 15. Lipids of Physiologic Significance
Kathleen M. Botham, PhD, DSc & Peter A. Mayes, PhD, DSc
OBJECTIVES
After studying this chapter, you should be able to:
![]() Define simple and complex lipids and identify the lipid classes in each group.
Define simple and complex lipids and identify the lipid classes in each group.
![]() Indicate the structure of saturated and unsaturated fatty acids, explain how the chain length and degree of unsaturation influence their melting point, give examples, and explain the nomenclature.
Indicate the structure of saturated and unsaturated fatty acids, explain how the chain length and degree of unsaturation influence their melting point, give examples, and explain the nomenclature.
![]() Understand the difference between cis and trans carbon-carbon double bonds.
Understand the difference between cis and trans carbon-carbon double bonds.
![]() Describe how eicosanoids are formed by modification of the structure of unsaturated fatty acids; identify the various eicosanoid classes and indicate their functions.
Describe how eicosanoids are formed by modification of the structure of unsaturated fatty acids; identify the various eicosanoid classes and indicate their functions.
![]() Outline the general structure of triacylglycerols and indicate their function.
Outline the general structure of triacylglycerols and indicate their function.
![]() Outline the general structure of phospholipids and glycosphingolipids and indicate the functions of the different classes.
Outline the general structure of phospholipids and glycosphingolipids and indicate the functions of the different classes.
![]() Appreciate the importance of cholesterol as the precursor of many biologically important steroids, including steroid hormones, bile acids, and vitamins D.
Appreciate the importance of cholesterol as the precursor of many biologically important steroids, including steroid hormones, bile acids, and vitamins D.
![]() Recognize the cyclic nucleus common to all steroids and explain the difference between the “chair” and “boat” forms of the six-carbon rings and that the rings may be either cis or trans in relation to each other, making many stereoisomers possible.
Recognize the cyclic nucleus common to all steroids and explain the difference between the “chair” and “boat” forms of the six-carbon rings and that the rings may be either cis or trans in relation to each other, making many stereoisomers possible.
![]() Explain why free radicals are damaging to tissues and identify the three stages in the chain reaction of lipid peroxidation that produces them continuously.
Explain why free radicals are damaging to tissues and identify the three stages in the chain reaction of lipid peroxidation that produces them continuously.
![]() Understand how antioxidants protect lipids from peroxidation by either inhibiting chain initiation or breaking the chain and give physiological and nonphysiological examples.
Understand how antioxidants protect lipids from peroxidation by either inhibiting chain initiation or breaking the chain and give physiological and nonphysiological examples.
![]() Understand that many lipid molecules are amphipathic, having both hydrophobic and hydrophilic groups in their structure, and explain how this influences their behavior in an aqueous environment and enables certain classes, including phospholipids, sphingolipids, and cholesterol, to form the basic structure of biologic membranes.
Understand that many lipid molecules are amphipathic, having both hydrophobic and hydrophilic groups in their structure, and explain how this influences their behavior in an aqueous environment and enables certain classes, including phospholipids, sphingolipids, and cholesterol, to form the basic structure of biologic membranes.
BIOMEDICAL IMPORTANCE
The lipids are a heterogeneous group of compounds, including fats, oils, steroids, waxes, and related compounds, that are related more by their physical than by their chemical properties. They have the common property of being (1) relatively insoluble in water and (2) soluble in nonpolar solvents such as ether and chloroform. They are important dietary constituents not only because of their high energy value, but also because fat-soluble vitamins and essential fatty acids are contained in the fat of natural foods. Fat is stored in adipose tissue, where it also serves as a thermal insulator in the subcutaneous tissues and around certain organs. Nonpolar lipids act as electrical insulators, allowing rapid propagation of depolarization waves along myelinated nerves. Combinations of lipid and protein (lipoproteins) serve as the means of transporting lipids in the blood. Lipids have essential roles in nutrition and health and knowledge of lipid biochemistry is necessary for the understanding of many important biomedical conditions, including obesity, diabetes mellitus, and atherosclerosis.
LIPIDS ARE CLASSIFIED AS SIMPLE OR COMPLEX
1. Simple lipids: Esters of fatty acids with various alcohols.
a. Fats: Esters of fatty acids with glycerol. Oils are fats in the liquid state.
b. Waxes: Esters of fatty acids with higher molecular weight monohydric alcohols.
2. Complex lipids: Esters of fatty acids containing groups in addition to an alcohol and a fatty acid.
a. Phospholipids: Lipids containing, in addition to fatty acids and an alcohol, a phosphoric acid residue. They frequently have nitrogen-containing bases and other substituents, for example, in glycerophospholipids the alcohol is glycerol and in sphingophospholipids the alcohol is sphingosine.
b. Glycolipids (glycosphingolipids): Lipids containing a fatty acid, sphingosine, and carbohydrate.
c. Other complex lipids: Lipids such as sulfolipids and amino lipids. Lipoproteins may also be placed in this category.
3. Precursor and derived lipids: These include fatty acids, glycerol, steroids, other alcohols, fatty aldehydes, ketone bodies (Chapter 22), hydrocarbons, lipid-soluble vitamins, and hormones.
Because they are uncharged, acylglycerols (glycerides), cholesterol, and cholesteryl esters are termed neutral lipids.
FATTY ACIDS ARE ALIPHATIC CARBOXYLIC ACIDS
Fatty acids occur in the body mainly as esters in natural fats and oils, but are found in the unesterified form as free fatty acids, a transport form in the plasma. Fatty acids that occur in natural fats usually contain an even number of carbon atoms. The chain may be saturated (containing no double bonds) or unsaturated (containing one or more double bonds) (Figure 15–1).
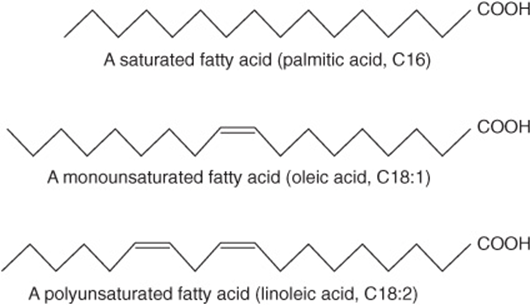
FIGURE 15–1 Fatty acids.
Fatty Acids Are Named after Corresponding Hydrocarbons
The most frequently used systematic nomenclature names the fatty acid after the hydrocarbon with the same number and arrangement of carbon atoms, with -oic being substituted for the final -e (Genevan system). Thus, saturated acids end in -anoic, for example, octanoic acid, and unsaturated acids with double bonds end in -enoic, for example, octadecenoic acid (oleic acid).
Carbon atoms are numbered from the carboxyl carbon (carbon no. 1). The carbon atoms adjacent to the carboxyl carbon (nos. 2, 3, and 4) are also known as the α, β, and γ carbons, respectively, and the terminal methyl carbon is known as the ω- or n-carbon.
Various conventions use Δ for indicating the number and position of the double bonds (Figure 15–2); for example, Δ9 indicates a double bond between carbons 9 and 10 of the fatty acid; ω9 indicates a double bond on the ninth carbon counting from the ω-carbon. In animals, additional double bonds are introduced only between the existing double bond (eg, ω9, ω6, or ω3) and the carboxyl carbon, leading to three series of fatty acids known as the ω9, ω6, and ω3 families, respectively.
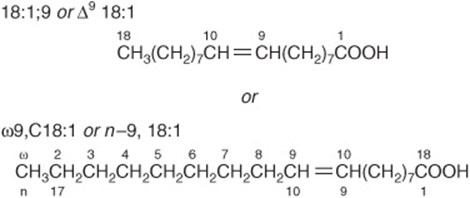
FIGURE 15–2 Oleic acid. n - 9 is equivalent to ω9.
Saturated Fatty Acids Contain No Double Bonds
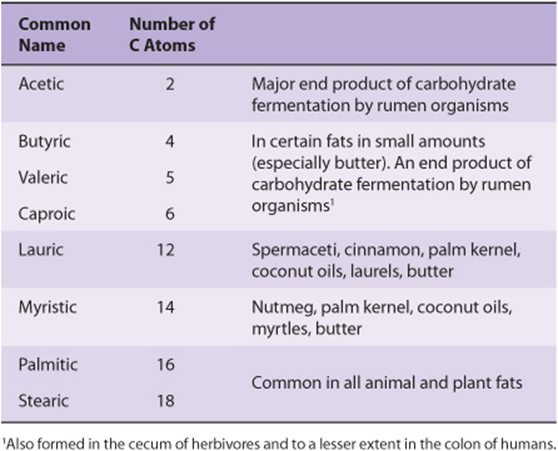
Saturated fatty acids may be envisaged as based on acetic acid (CH3—COOH) as the first member of the series in which—CH2—is progressively added between the terminal CH3—and—COOH groups. Examples are shown in Table 15-1. Other higher members of the series are known to occur, particularly in waxes. A few branched-chain fatty acids have also been isolated from both plant and animal sources.
TABLE 15–1 Saturated Fatty Acids
Unsaturated Fatty Acids Contain One or More Double Bonds
Unsaturated fatty acids (Figure 15–1, Table 15-2) may be further subdivided as follows:
TABLE 15–2 Unsaturated Fatty Acids of Physiologic and Nutritional Significance
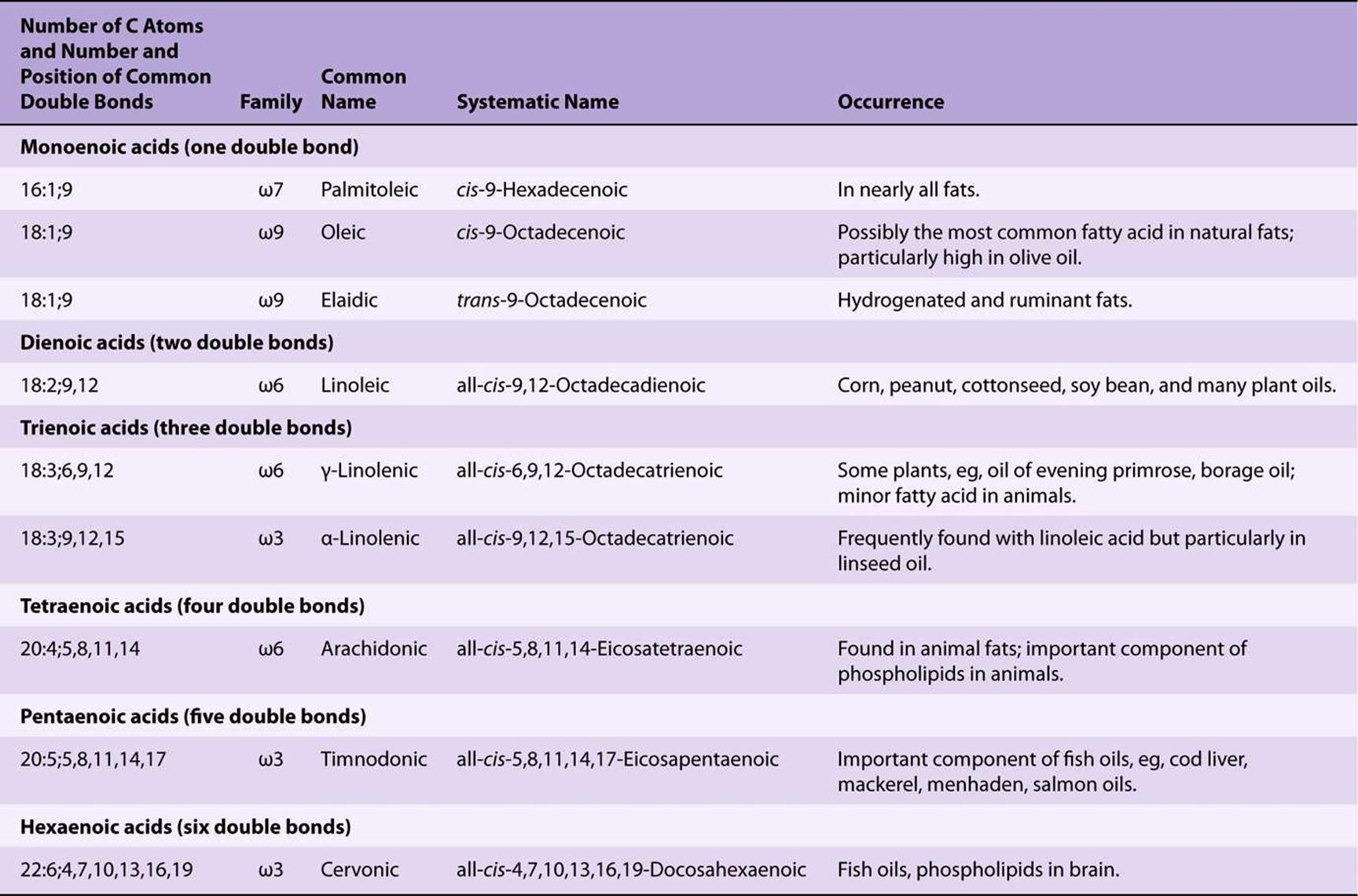
1. Monounsaturated (monoethenoid, monoenoic) acids, containing one double bond.
2. Polyunsaturated (polyethenoid, polyenoic) acids, containing two or more double bonds.
3. Eicosanoids: These compounds, derived from eicosa (20-carbon) polyenoic fatty acids, comprise the prostanoids, leukotrienes (LTs), and lipoxins (LXs). Prostanoids include prostaglandins (PGs), prostacyclins (PGIs), and thromboxanes (TXs).
Prostaglandins exist in virtually every mammalian tissue, acting as local hormones; they have important physiologic and pharmacologic activities. They are synthesized in vivo by cyclization of the center of the carbon chain of 20-carbon (eicosanoic) polyunsaturated fatty acids (eg, arachidonic acid) to form a cyclopentane ring (Figure 15–3). A related series of compounds, the thromboxanes, have the cyclopentane ring interrupted with an oxygen atom (oxane ring) (Figure 15–4). Three different eicosanoic fatty acids give rise to three groups of eicosanoids characterized by the number of double bonds in the side chains, for example, PG1, PG2, and PG3. Different substituent groups attached to the rings give rise to series of prostaglandins and thromboxanes, labeled A, B, etc—for example, the “E” type of prostaglandin (as in PGE2) has a keto group in position 9, whereas the “F” type has a hydroxyl group in this position. The leukotrienes and lipoxins (Figure 15–5) are a third group of eicosanoid derivatives formed via the lipoxygenase pathway (Figure 23–11). They are characterized by the presence of three or four conjugated double bonds, respectively. Leukotrienes cause bronchoconstriction as well as being potent proinflammatory agents, and play a part in asthma.
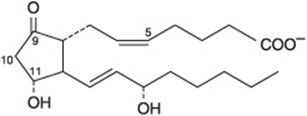
FIGURE 15–3 Prostaglandin E2 (PGE2).
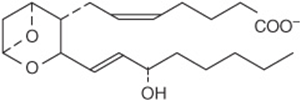
FIGURE 15–4 Thromboxane A2 (TXA2).
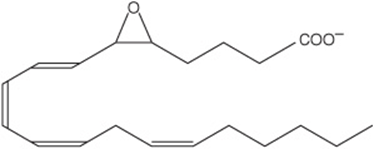
FIGURE 15–5 Leukotriene A4 (LTA4).
Most Naturally Occurring Unsaturated Fatty Acids Have cis Double Bonds
The carbon chains of saturated fatty acids form a zigzag pattern when extended at low temperatures (Figure 15–1). At higher temperatures, some bonds rotate, causing chain shortening, which explains why biomembranes become thinner with increases in temperature. A type of geometric isomerism occurs in unsaturated fatty acids, depending on the orientation of atoms or groups around the axes of double bonds, which do not allow rotation. If the acyl chains are on the same side of the bond, it is cis-, as in oleic acid; if on opposite sides, it is trans-, as in elaidic acid, the trans isomer of oleic acid (Figure 15–6). Double bonds in naturally occurring unsaturated long-chain fatty acids are nearly all in the cis configuration, the molecules being “bent” 120° at the double bond. Thus, oleic acid has an L shape, whereas elaidic acid remains “straight.” Increase in the number of cis double bonds in a fatty acid leads to a variety of possible spatial configurations of the molecule—for example, arachidonic acid, with four cis double bonds, is bent into a U shape (Figure 15–7). This has profound significance for molecular packing in cell membranes and on the positions occupied by fatty acids in more complex molecules such as phospholipids. Trans double bonds alter these spatial relationships. Trans fatty acids are present in certain foods, arising as a by-product of the saturation of fatty acids during hydrogenation, or “hardening,” of natural oils in the manufacture of margarine. An additional small contribution comes from the ingestion of ruminant fat that contains trans fatty acids arising from the action of microorganisms in the rumen. Consumption of trans fatty acids is now known to be detrimental to health and is associated with increased risk of diseases including cardiovascular disease and diabetes mellitus. This has led to improved technology to produce soft margarine low in trans fatty acids or containing none at all.
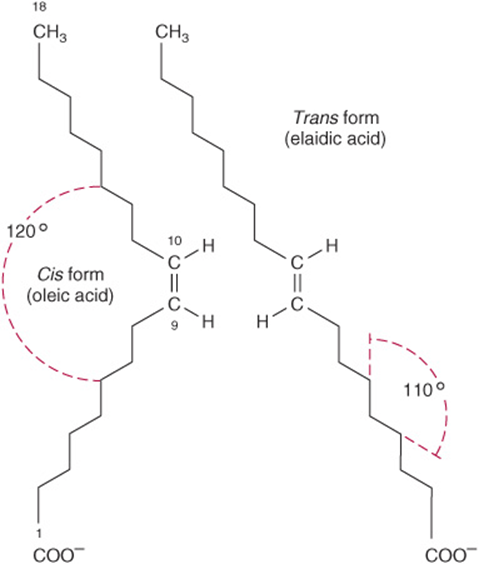
FIGURE 15–6 Geometric isomerism of Δ9, 18:1 fatty acids (oleic and elaidic acids).
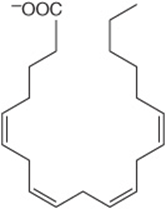
FIGURE 15–7 Arachidonic acid.
Physical and Physiologic Properties of Fatty Acids Reflect Chain Length and Degree of Unsaturation
The melting points of even-numbered carbon fatty acids increase with chain length and decrease according to unsaturation. A triacylglycerol containing three saturated fatty acids of 12 carbons or more is solid at body temperature, whereas if the fatty acid residues are 18:2, it is liquid to below 0°C. In practice, natural acylglycerols contain a mixture of fatty acids tailored to suit their functional roles. The membrane lipids, which must be fluid at all environmental temperatures, are more unsaturated than storage lipids. Lipids in tissues that are subject to cooling, for example, in hibernators or in the extremities of animals, are more unsaturated.
TRIACYLGLYCEROLS (TRIGLYCERIDES)* ARE THE MAIN STORAGE FORMS OF FATTY ACIDS
The triacylglycerols (Figure 15–8) are esters of the trihydric alcohol glycerol and fatty acids. Mono- and diacylglycerols, wherein one or two fatty acids are esterified with glycerol, are also found in the tissues. These are of particular significance in the synthesis and hydrolysis of triacylglycerols.
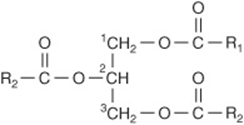
FIGURE 15–8 Triacylglycerol.
Carbons 1 & 3 of Glycerol Are Not Identical
To number the carbon atoms of glycerol unambiguously, the -sn (stereochemical numbering) system is used. It is important to realize that carbons 1 and 3 of glycerol are not identical when viewed in three dimensions (shown as a projection formula in Figure 15–9). Enzymes readily distinguish between them and are nearly always specific for one or the other carbon; for example, glycerol is always phosphorylated on sn-3 by glycerol kinase to give glycerol 3-phosphate and not glycerol 1-phosphate.
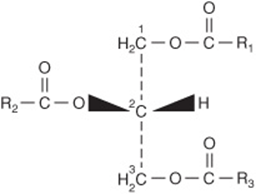
FIGURE 15–9 Triacyl-sn-glycerol.
PHOSPHOLIPIDS ARE THE MAIN LIPID CONSTITUENTS OF MEMBRANES
Phospholipids may be regarded as derivatives of phosphatidic acid (Figure 15–10), in which the phosphate is esterified with the—OH of a suitable alcohol. Phosphatidic acid is important as an intermediate in the synthesis of triacylglycerols as well as phosphoglycerols but is not found in any great quantity in tissues.
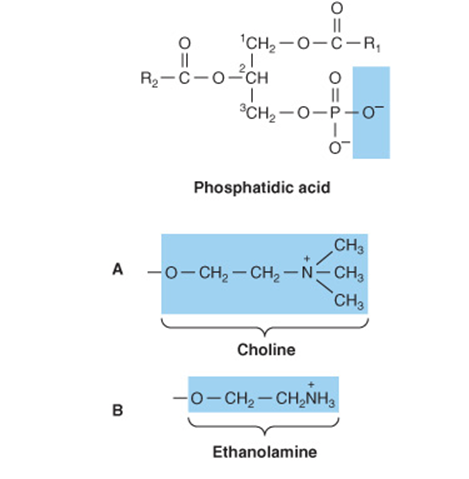
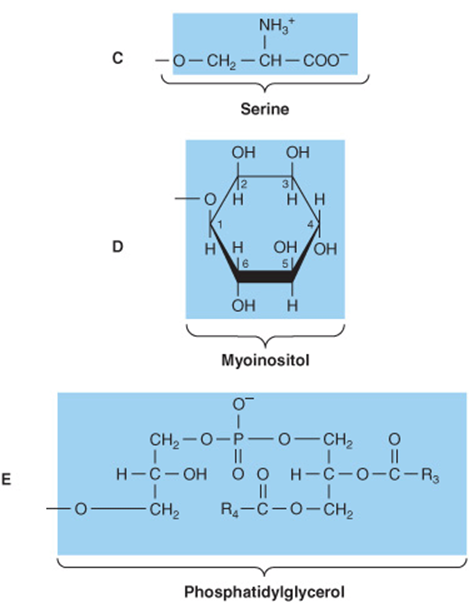
FIGURE 15–10 Phosphatidic acid and its derivatives. The O- shown shaded in phosphatidic acid is substituted by the substituents shown to form in (A) 3-phosphatidylcholine, (B) 3-phosphatidylethanolamine, (C) 3-phosphatidylserine, (D) 3-phosphatidylinositol, and (E) cardiolipin (diphosphatidylglycerol).
Phosphatidylcholines (Lecithins) Occur in Cell Membranes
Phosphoacylglycerols containing choline (Figure 15–10) are the most abundant phospholipids of the cell membrane and represent a large proportion of the body’s store of choline. Choline is important in nervous transmission, as acetylcholine, and as a store of labile methyl groups. Dipalmitoyl lecithin is a very effective surface-active agent and a major constituent of the surfactant preventing adherence, due to surface tension, of the inner surfaces of the lungs. Its absence from the lungs of premature infants causes respiratory distress syndrome. Most phospholipids have a saturated acyl radical in the sn-1 position but an unsaturated radical in the sn-2 position of glycerol.
Phosphatidylethanolamine (cephalin) and phos-phatidylserine (found in most tissues) are also found in cell membranes and differ from phosphatidylcholine only in that ethanolamine or serine, respectively, replaces choline (Figure 15–10). Phosphatidylserine also plays a role in apop-tosis (programmed cell death).
Phosphatidylinositol Is a Precursor of Second Messengers
The inositol is present in phosphatidylinositol as the stereoisomer, myoinositol (Figure 15–10). Phosphatidylinositol 4,5-bisphosphate is an important constituent of cell membrane phospholipids; upon stimulation by a suitable hormone agonist, it is cleaved into diacylglycerol and inositol trisphosphate, both of which act as internal signals or second messengers.
Cardiolipin Is a Major Lipid of Mitochondrial Membranes
Phosphatidic acid is a precursor of phosphatidylglycerol, which in turn gives rise to cardiolipin (Figure 15–10). This phospholipid is found only in mitochondria and is essential for the mitochondrial function. Decreased cardiolipin levels or alterations in its structure or metabolism cause mitochondrial dysfunction in aging and in pathological conditions including heart failure, hypothyroidism, and Barth syndrome (cardioskeletal myopathy).
Lysophospholipids Are Intermediates in the Metabolism of Phosphoglycerols
These are phosphoacylglycerols containing only one acyl radical, for example, lysophosphatidylcholine (lysolecithin) (Figure 15–11), important in the metabolism and interconversion of phospholipids. It is also found in oxidized lipoproteins and has been implicated in some of their effects in promoting atherosclerosis.
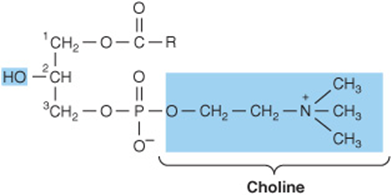
FIGURE 15–11 Lysophosphatidylcholine (lysolecithin).
Plasmalogens Occur in Brain & Muscle
These compounds constitute as much as 10% of the phospholipids of brain and muscle. Structurally, the plasmalogens resemble phosphatidylethanolamine but possess an ether link on the sn-1 carbon instead of the ester link found in acylglycerols. Typically, the alkyl radical is an unsaturated alcohol (Figure 15–12). In some instances, choline, serine, or inositol may be substituted for ethanolamine.
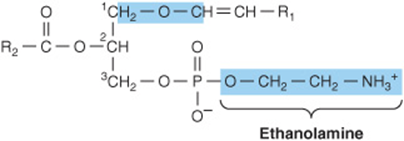
FIGURE 15–12 Plasmalogen.
Sphingomyelins Are Found in the Nervous System
Sphingomyelins are found in large quantities in brain and nerve tissue. On hydrolysis, the sphingomyelins yield a fatty acid, phosphoric acid, choline, and a complex amino alcohol, sphingosine (Figure 15–13). No glycerol is present. The combination of sphingosine plus fatty acid is known as ceramide, a structure also found in the glycosphingolipids (see next Section below).
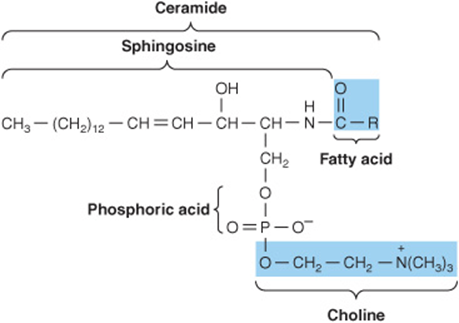
FIGURE 15–13 A sphingomyelin.
GLYCOLIPIDS (GLYCOSPHINGOLIPIDS) ARE IMPORTANT IN NERVE TISSUES & IN THE CELL MEMBRANE
Glycolipids are widely distributed in every tissue of the body, particularly in nervous tissue such as brain. They occur particularly in the outer leaflet of the plasma membrane, where they contribute to cell surface carbohydrates.
The major glycolipids found in animal tissues are glycosphingolipids. They contain ceramide and one or more sugars. Galactosylceramide is a major glycosphingolipid of brain and other nervous tissue, found in relatively low amounts elsewhere. It contains a number of characteristic C24 fatty acids, for example, cerebronic acid.
Galactosylceramide (Figure 15–14) can be converted to sulfogalactosylceramide (sulfatide), present in high amounts in myelin. Glucosylceramide is the predominant simple glycosphingolipid of extraneural tissues, also occurring in the brain in small amounts. Gangliosides are complex glycosphingolipids derived from glucosylceramide that contain in addition one or more molecules of a sialic acid. Neuraminic acid (NeuAc; see Chapter 14) is the principal sialic acid found in human tissues. Gangliosides are also present in nervous tissues in high concentration. They appear to have receptor and other functions. The simplest ganglioside found in tissues is GM3, which contains ceramide, one molecule of glucose, one molecule of galactose, and one molecule of NeuAc. In the shorthand nomenclature used, G represents ganglioside; M is a monosialo-containing species; and the subscript 3 is a number assigned on the basis of chromatographic migration. GM1 (Figure 15–15), a more complex ganglioside derived from GM3, is of considerable biologic interest, as it is known to be the receptor in human intestine for cholera toxin. Other gangliosides can contain anywhere from one to five molecules of sialic acid, giving rise to di-, trisialogangliosides, etc.
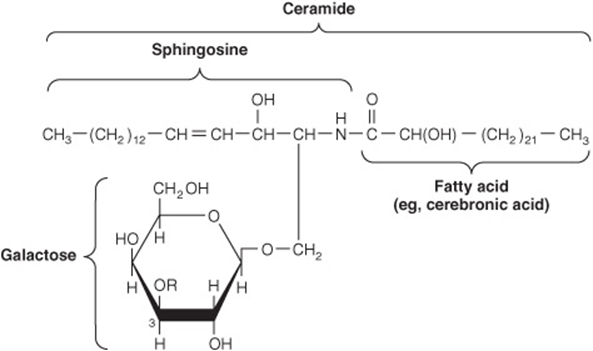
FIGURE 15–14 Structure of galactosylceramide (galactocerebroside, ![]() ), and sulfogalactosylceramide (a sulfatide,
), and sulfogalactosylceramide (a sulfatide, ![]() .)
.)
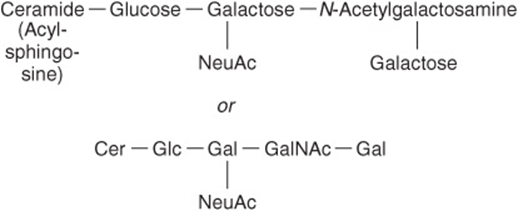
FIGURE 15–15 GM1 ganglioside, a monosialoganglioside, the receptor in human intestine for cholera toxin.
STEROIDS PLAY MANY PHYSIOLOGICALLY IMPORTANT ROLES
Cholesterol is probably the best-known steroid because of its association with atherosclerosis and heart disease. However, biochemically it is also of significance because it is the precursor of a large number of equally important steroids that include the bile acids, adrenocortical hormones, sex hormones, D vitamins, cardiac glycosides, sitosterols of the plant kingdom, and some alkaloids.
All steroids have a similar cyclic nucleus resembling phenanthrene (rings A, B, and C) to which a cyclopentane ring (D) is attached. The carbon positions on the steroid nucleus are numbered as shown in Figure 15–16. It is important to realize that in structural formulas of steroids, a simple hexagonal ring denotes a completely saturated six-carbon ring with all valences satisfied by hydrogen bonds unless shown otherwise; that is, it is not a benzene ring. All double bonds are shown as such. Methyl side chains are shown as single bonds unattached at the farther (methyl) end. These occur typically at positions 10 and 13 (constituting C atoms 19 and 18). A side chain at position 17 is usual (as in cholesterol). If the compound has one or more hydroxyl groups and no carbonyl or carboxyl groups, it is a sterol, and the name terminates in -ol.
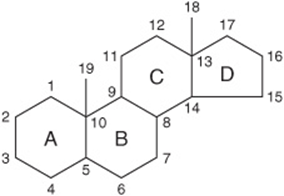
FIGURE 15–16 The steroid nucleus.
Because of Asymmetry in the Steroid Molecule, Many Stereoisomers Are Possible
Each of the six-carbon rings of the steroid nucleus is capable of existing in the three-dimensional conformation either of a “chair” or a “boat” (Figure 15–17). In naturally occurring steroids, virtually all the rings are in the “chair” form, which is the more stable conformation. With respect to each other, the rings can be either cis or trans (Figure 15–18). The junction between the A and B rings can be cis or trans in naturally occurring steroids. That between B and C is trans, as is usually the C/D junction. Bonds attaching substituent groups above the plane of the rings (β bonds) are shown with bold solid lines, whereas those bonds attaching groups below (α bonds) are indicated with broken lines. The A ring of a 5α steroid is always trans to the B ring, whereas it is cis in a 5β steroid. The methyl groups attached to C10 and C13 are invariably in the β configuration.
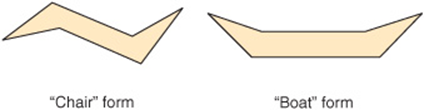
FIGURE 15–17 Conformations of stereoisomers of the steroid nucleus.
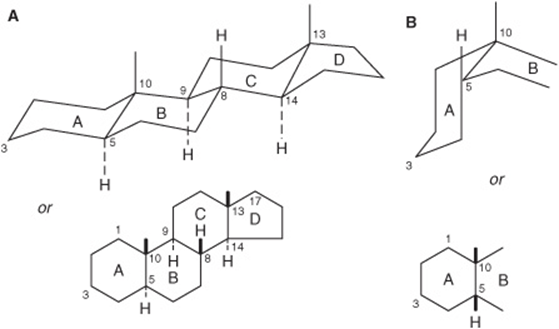
FIGURE 15–18 Generalized steroid nucleus, showing (A) an all-trans configuration between adjacent rings and (B) a cis configuration between rings A and B.
Cholesterol Is a Significant Constituent of Many Tissues
Cholesterol (Figure 15–19) is widely distributed in all cells of the body but particularly in nervous tissue. It is a major constituent of the plasma membrane and of plasma lipoproteins. It is often found as cholesteryl ester, where the hydroxyl group on position 3 is esterified with a long-chain fatty acid. It occurs in animals but not in plants or bacteria.
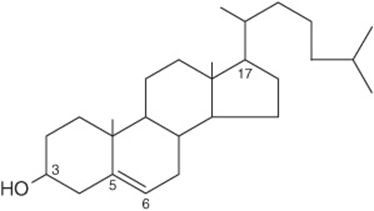
FIGURE 15–19 Cholesterol, 3-hydroxy-5,6-cholestene.
Ergosterol Is a Precursor of Vitamin D
Ergosterol occurs in plants and yeast and is important as a precursor of vitamin D (Figure 15–20). When irradiated with ultraviolet light, ring B is opened to form vitamin D2 in a process similar to the one that forms vitamin D3from 7-dehydrocholesterol in the skin (Figure 44–3).
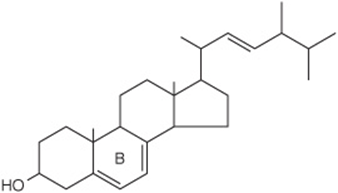
FIGURE 15–20 Ergosterol.
Polyprenoids Share the Same Parent Compound as Cholesterol
Although not steroids, polyprenoids are related because they are synthesized, like cholesterol (Figure 26–2), from five-carbon isoprene units (Figure 15–21). They include ubiquinone (Chapter 13), which participates in the respiratory chain in mitochondria, and the long-chain alcohol dolichol (Figure 15–22), which takes part in glycoprotein synthesis by transferring carbohydrate residues to asparagine residues of the polypeptide (Chapter 47). Plant-derived isoprenoid compounds include rubber, camphor, the fat-soluble vitamins A, D, E, and K, and β-carotene (provitamin A).
![]()
FIGURE 15–21 Isoprene unit.
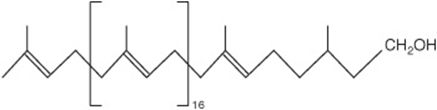
FIGURE 15–22 Dolichol—a C95 alcohol.
LIPID PEROXIDATION IS A SOURCE OF FREE RADICALS
Peroxidation (auto-oxidation) of lipids exposed to oxygen is responsible not only for deterioration of foods (rancidity), but also for damage to tissues in vivo, where it may be a cause of cancer, inflammatory diseases, atherosclerosis, and aging. The deleterious effects are considered to be caused by free radicals (ROO•, RO•, OH•) produced during peroxide formation from fatty acids containing methylene-interrupted double bonds, that is, those found in the naturally occurring polyunsaturated fatty acids (Figure 15–23). Lipid peroxidation is a chain reaction providing a continuous supply of free radicals that initiate further peroxidation and thus has potentially devastating effects. The whole process can be depicted as follows:
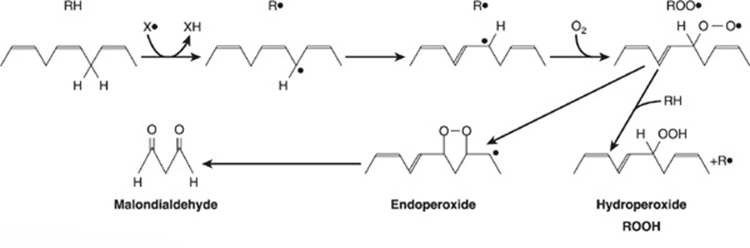
FIGURE 15–23 Lipid peroxidation. The reaction is initiated by an existing free radical (X•), by light, or by metal ions. Malondialdehyde is only formed by fatty acids with three or more double bonds and is used as a measure of lipid peroxidation together with ethane from the terminal two carbons of ω3 fatty acids and pentane from the terminal five carbons of ω6 fatty acids.
1. Initiation:
![]()
2. Propagation:
![]()
3. Termination:

To control and reduce lipid peroxidation, both humans in their activities and nature invoke the use of antioxidants. Propyl gallate, butylated hydroxyanisole (BHA), and butylated hydroxytoluene (BHT) are antioxidants used as food additives. Naturally occurring antioxidants include vitamin E (tocopherol), which is lipid soluble, and urate and vitamin C, which are water soluble. Beta-carotene is an antioxidant at low PO2. Antioxidants fall into two classes: (1) preventive antioxidants, which reduce the rate of chain initiation and (2) chain-breaking antioxidants, which interfere with chain propagation. Preventive antioxidants include catalase and other peroxidases such as glutathione peroxidase (Figure 21–3) that react with ROOH; selenium, which is an essential component of glutathione peroxidase and regulates its activity, and chelators of metal ions such as EDTA (ethylenediaminetetraacetate) and DTPA (diethylenetriaminepentaacetate). In vivo, the principal chain-breaking antioxidants are superoxide dismutase, which acts in the aqueous phase to trap superoxide free radicals ![]() urate, and vitamin E, which acts in the lipid phase to trap ROO• radicals (Figure 44–6).
urate, and vitamin E, which acts in the lipid phase to trap ROO• radicals (Figure 44–6).
Peroxidation is also catalyzed in vivo by heme compounds and by lipoxygenases found in platelets and leukocytes. Other products of auto-oxidation or enzymic oxidation of physiologic significance include oxysterols (formed from cholesterol) and isoprostanes (formed from the peroxidation of polyunsaturated fatty acids such as arachidonic acid).
AMPHIPATHICLIPIDS SELF-ORIENT AT OIL: WATER INTERFACES
They Form Membranes, Micelles, Liposomes, & Emulsions
In general, lipids are insoluble in water since they contain a predominance of nonpolar (hydrocarbon) groups. However, fatty acids, phospholipids, sphingolipids, bile salts, and, to a lesser extent, cholesterol contain polar groups. Therefore, a part of the molecule is hydrophobic, or water insoluble; and a part is hydrophilic, or water soluble. Such molecules are described as amphipathic (Figure 15–24). They become oriented at oil-water interfaces with the polar group in the water phase and the nonpolar group in the oil phase. A bilayer of such amphipathic lipids is the basic structure in biologic membranes (Chapter 40). When a critical concentration of these lipids is present in an aqueous medium, they form micelles. Liposomes may be formed by sonicating an amphipathic lipid in an aqueous medium. They consist of spheres of lipid bilayers that enclose part of the aqueous medium. Aggregations of bile salts into micelles and liposomes and the formation of mixed micelles with the products of fat digestion are important in facilitating absorption of lipids from the intestine. Liposomes are of potential clinical use—particularly when combined with tissue-specific antibodies—as carriers of drugs in the circulation, targeted to specific organs, for example, in cancer therapy. In addition, they are used for gene transfer into vascular cells and as carriers for topical and transdermal delivery of drugs and cosmetics. Emulsions are much larger particles, formed usually by nonpolar lipids in an aqueous medium. These are stabilized by emulsifying agents such as amphipathic lipids (eg, lecithin), which form a surface layer separating the main bulk of the nonpolar material from the aqueous phase (Figure 15–24).
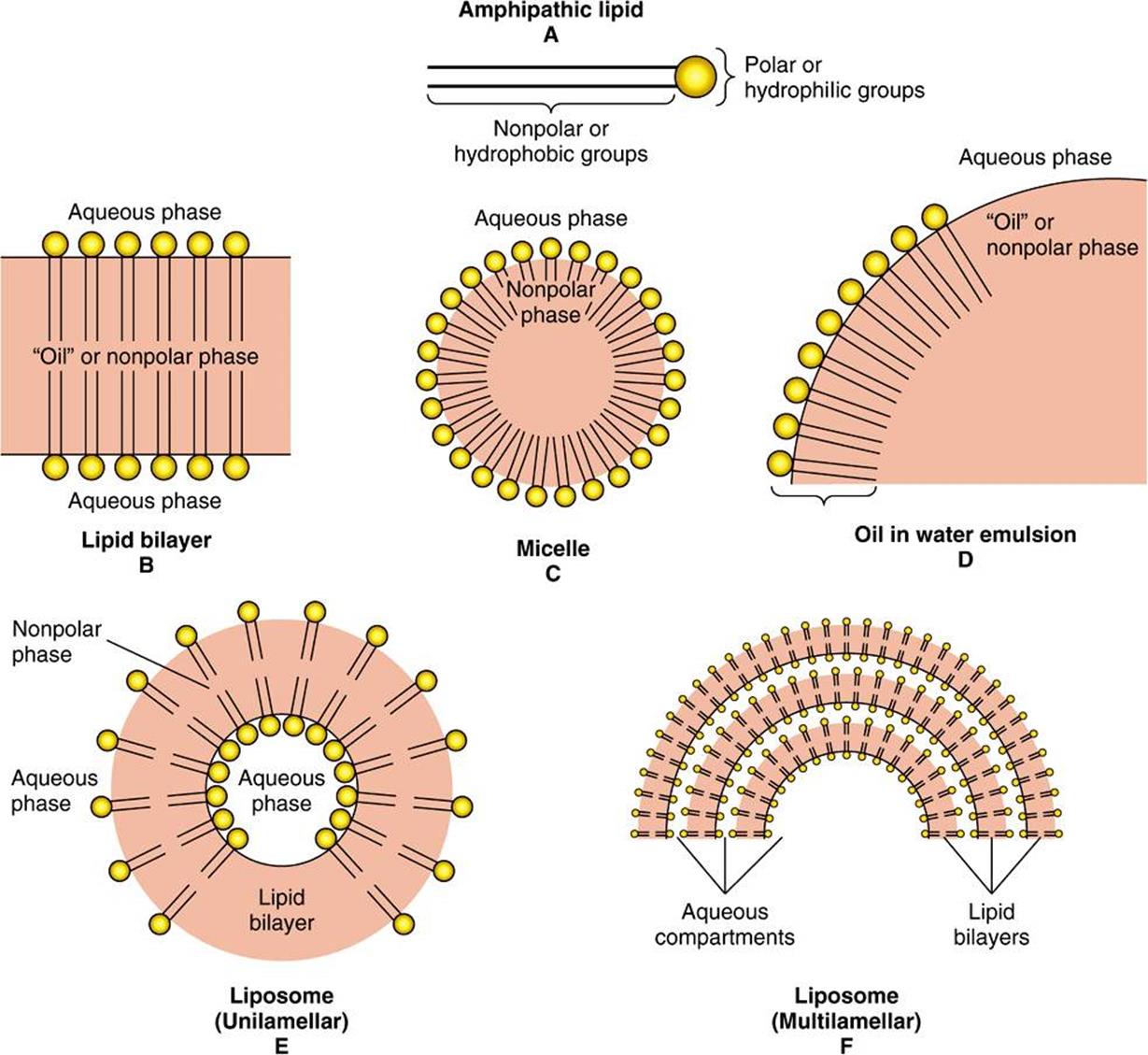
FIGURE 15–24 Formation of lipid membranes, micelles, emulsions, and liposomes from amphipathic lipids, for example, phospholipids.
SUMMARY
![]() Lipids have the common property of being relatively insoluble in water (hydrophobic) but soluble in nonpolar solvents. Amphipathic lipids also contain one or more polar groups, making them suitable as constituents of membranes at lipid-water interfaces.
Lipids have the common property of being relatively insoluble in water (hydrophobic) but soluble in nonpolar solvents. Amphipathic lipids also contain one or more polar groups, making them suitable as constituents of membranes at lipid-water interfaces.
![]() The lipids of major physiologic significance are fatty acids and their esters, together with cholesterol and other steroids.
The lipids of major physiologic significance are fatty acids and their esters, together with cholesterol and other steroids.
![]() Long-chain fatty acids may be saturated, monounsaturated, or polyunsaturated, according to the number of double bonds present. Their fluidity decreases with chain length and increases according to degree of unsaturation.
Long-chain fatty acids may be saturated, monounsaturated, or polyunsaturated, according to the number of double bonds present. Their fluidity decreases with chain length and increases according to degree of unsaturation.
![]() Eicosanoids are formed from 20-carbon polyunsaturated fatty acids and make up an important group of physiologically and pharmacologically active compounds known as prostaglandins, thromboxanes, leukotrienes, and lipoxins.
Eicosanoids are formed from 20-carbon polyunsaturated fatty acids and make up an important group of physiologically and pharmacologically active compounds known as prostaglandins, thromboxanes, leukotrienes, and lipoxins.
![]() The esters of glycerol are quantitatively the most significant lipids, represented by triacylglycerol (“fat”), a major constituent of some lipoprotein classes and the storage form of lipid in adipose tissue. Phosphoacylglycerols are amphipathic lipids and have important roles—as major constituents of membranes and the outer layer of lipoproteins, as surfactant in the lung, as precursors of second messengers, and as constituents of nervous tissue.
The esters of glycerol are quantitatively the most significant lipids, represented by triacylglycerol (“fat”), a major constituent of some lipoprotein classes and the storage form of lipid in adipose tissue. Phosphoacylglycerols are amphipathic lipids and have important roles—as major constituents of membranes and the outer layer of lipoproteins, as surfactant in the lung, as precursors of second messengers, and as constituents of nervous tissue.
![]() Glycolipids are also important constituents of nervous tissue such as brain and the outer leaflet of the cell membrane, where they contribute to the carbohydrates on the cell surface.
Glycolipids are also important constituents of nervous tissue such as brain and the outer leaflet of the cell membrane, where they contribute to the carbohydrates on the cell surface.
![]() Cholesterol, an amphipathic lipid, is an important component of membranes. It is the parent molecule from which all other steroids in the body, including major hormones such as the adrenocortical and sex hormones, D vitamins, and bile acids, are synthesized.
Cholesterol, an amphipathic lipid, is an important component of membranes. It is the parent molecule from which all other steroids in the body, including major hormones such as the adrenocortical and sex hormones, D vitamins, and bile acids, are synthesized.
![]() Peroxidation of lipids containing polyunsaturated fatty acids leads to generation of free radicals that damage tissues and cause disease.
Peroxidation of lipids containing polyunsaturated fatty acids leads to generation of free radicals that damage tissues and cause disease.
REFERENCES
Benzie IFF: Lipid peroxidation: a review of causes, consequences, measurement and dietary influences. Int J Food Sci Nutr 1996;47:233.
Christie WW: Lipid Analysis, 3rd ed. The Oily Press, 2003.
Dowhan W, Bodanov H, Mileykovskaya E: Functional roles of lipids in membranes. In: Biochemistry of Lipids, Lipoproteins and Membranes, 5th ed. Vance DE, Vance JE (editors). Elsevier, 2008:1-37.
Gunstone FD, Harwood JL, Dijkstra AJ: The Lipid Handbook with CD-Rom. CRC Press, 2007.
Gurr MI, Harwood JL, Frayn K: Lipid Biochemistry. Blackwell Publishing, 2002.
*According to the standardized terminology of the International Union of Pure and Applied Chemistry and the International Union of Biochemistry, the monoglycerides, diglycerides, and triglycerides should be designated monoacylglycerols, diacylglycerols, and triacylglycerols, respectively. However, the older terminology is still widely used, particularly in clinical medicine.