Harper’s Illustrated Biochemistry, 29th Edition (2012)
SECTION II. Bioenergetics & the Metabolism of Carbohydrates & Lipids
Chapter 24. Metabolism of Acylglycerols & Sphingolipids
Kathleen M. Botham, PhD, DSc & Peter A. Mayes, PhD, DSc
OBJECTIVES
After studying this chapter, you should be able to:
![]() Appreciate that the catabolism of triacylglycerols involves hydrolysis by a lipase to free fatty acids and glycerol and indicate the fate of these metabolites.
Appreciate that the catabolism of triacylglycerols involves hydrolysis by a lipase to free fatty acids and glycerol and indicate the fate of these metabolites.
![]() Understand that glycerol-3-phosphate is the substrate for the formation of both triacylglycerols and phosphoglycerols and that a branch point at phosphatidate leads to the synthesis of inositol phospholipids and cardiolipin via one branch and triacylglycerols and other phospholipids via the second branch.
Understand that glycerol-3-phosphate is the substrate for the formation of both triacylglycerols and phosphoglycerols and that a branch point at phosphatidate leads to the synthesis of inositol phospholipids and cardiolipin via one branch and triacylglycerols and other phospholipids via the second branch.
![]() Explain that plasmalogens and platelet activating factor (PAF) are formed by a complex pathway starting from dihydroxyacetone phosphate.
Explain that plasmalogens and platelet activating factor (PAF) are formed by a complex pathway starting from dihydroxyacetone phosphate.
![]() Illustrate the role of various phospholipases in the degradation and remodeling of phospholipids.
Illustrate the role of various phospholipases in the degradation and remodeling of phospholipids.
![]() Appreciate that ceramide is produced from the amino acid serine and is the precursor from which all sphingolipids are formed.
Appreciate that ceramide is produced from the amino acid serine and is the precursor from which all sphingolipids are formed.
![]() Indicate how sphingomyelin and glycosphingolipids are produced by reacting ceramide with phosphatidylcholine (with the release of diacylglycerol) or sugar residue(s), respectively.
Indicate how sphingomyelin and glycosphingolipids are produced by reacting ceramide with phosphatidylcholine (with the release of diacylglycerol) or sugar residue(s), respectively.
![]() Identify examples of disease processes caused by defects in phospholipid or sphingolipid synthesis or breakdown.
Identify examples of disease processes caused by defects in phospholipid or sphingolipid synthesis or breakdown.
BIOMEDICAL IMPORTANCE
Acylglycerols constitute the majority of lipids in the body. Triacylglycerols are the major lipids in fat deposits and in food, and their roles in lipid transport and storage and in various diseases such as obesity, diabetes, and hyperlipoproteinemia will be described in subsequent chapters. The amphipathic nature of phospholipids and sphingolipids makes them ideally suitable as the main lipid component of cell membranes. Phospholipids also take part in the metabolism of many other lipids. Some phospholipids have specialized functions; eg, dipalmitoyl lecithin is a major component of lung surfactant, which is lacking in respiratory distress syndrome of the newborn. Inositol phospholipids in the cell membrane act as precursors of hormone second messengers, and platelet-activating factor is an alkylphospholipid. Glycosphingolipids, containing sphingosine and sugar residues as well as fatty acid that are found in the outer leaflet of the plasma membrane with their oligosaccharide chains facing outward, form part of the glycocalyx of the cell surface and are important (1) in cell adhesion and cell recognition, (2) as receptors for bacterial toxins (eg, the toxin that causes cholera), and (3) as ABO blood group substances. A dozen or so glycolipid storage diseases have been described (eg, Gaucher’s disease and Tay-Sachs disease), each due to a genetic defect in the pathway for glycolipid degradation in the lysosomes.
HYDROLYSIS INITIATES CATABOLISM OF TRIACYLGLYCEROLS
Triacylglycerols must be hydrolyzed by a lipase to their constituent fatty acids and glycerol before further catabolism can proceed. Much of this hydrolysis (lipolysis) occurs in adipose tissue with release of free fatty acids into the plasma, where they are found combined with serum albumin (Figure 25–7). This is followed by free fatty acid uptake into tissues (including liver, heart, kidney, muscle, lung, testis, and adipose tissue, but not readily by brain), where they are oxidized or re-esterified. The utilization of glycerol depends upon whether such tissues have the enzyme glycerol kinase, which is found in significant amounts in liver, kidney, intestine, brown adipose tissue, and the lactating mammary gland.
TRIACYLGLYCEROLS & PHOSPHOGLYCEROLS ARE FORMED BY ACYLATION OF TRIOSE PHOSPHATES
The major pathways of triacylglycerol and phosphoglycerol biosynthesis are outlined in Figure 24–1. Important substances such as triacylglycerols, phosphatidylcholine, phosphatidylethanolamine, phosphatidylinositol, and cardiolipin, a constituent of mitochondrial membranes, are formed from glycerol-3-phosphate. Significant branch points in the pathway occur at the phosphatidate and diacylglycerol steps. Phosphoglycerols containing an ether link (—C—O—C—), the best known of which are plasmalogens and platelet-activating factor (PAF), are derived from dihydroxyacetone phosphate. Glycerol 3-phosphate and dihydroxyacetone phosphate are intermediates in glycolysis, making a very important connection between carbohydrate and lipid metabolism (see Chapter 16).
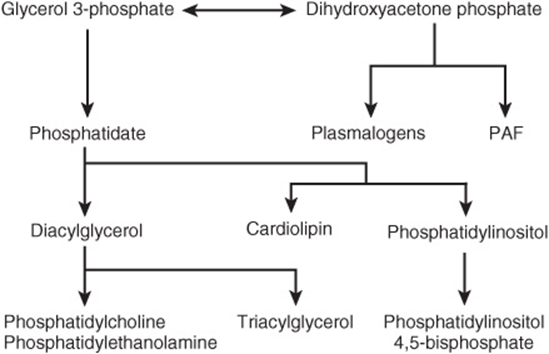
FIGURE 24–1 Overview of acylglycerol biosynthesis. (PAF, platelet-activating factor.)
Phosphatidate Is the Common Precursor in the Biosynthesis of Triacylglycerols, Many Phosphoglycerols, & Cardiolipin
Both glycerol and fatty acids must be activated by ATP before they can be incorporated into acylglycerols. Glycerol kinase catalyzes the activation of glycerol to sn-glycerol 3-phosphate. If the activity of this enzyme is absent or low, as in muscle or adipose tissue, most of the glycerol 3-phosphate is formed from dihydroxyacetone phosphate by glycerol-3-phosphate dehydrogenase (Figure 24–2).
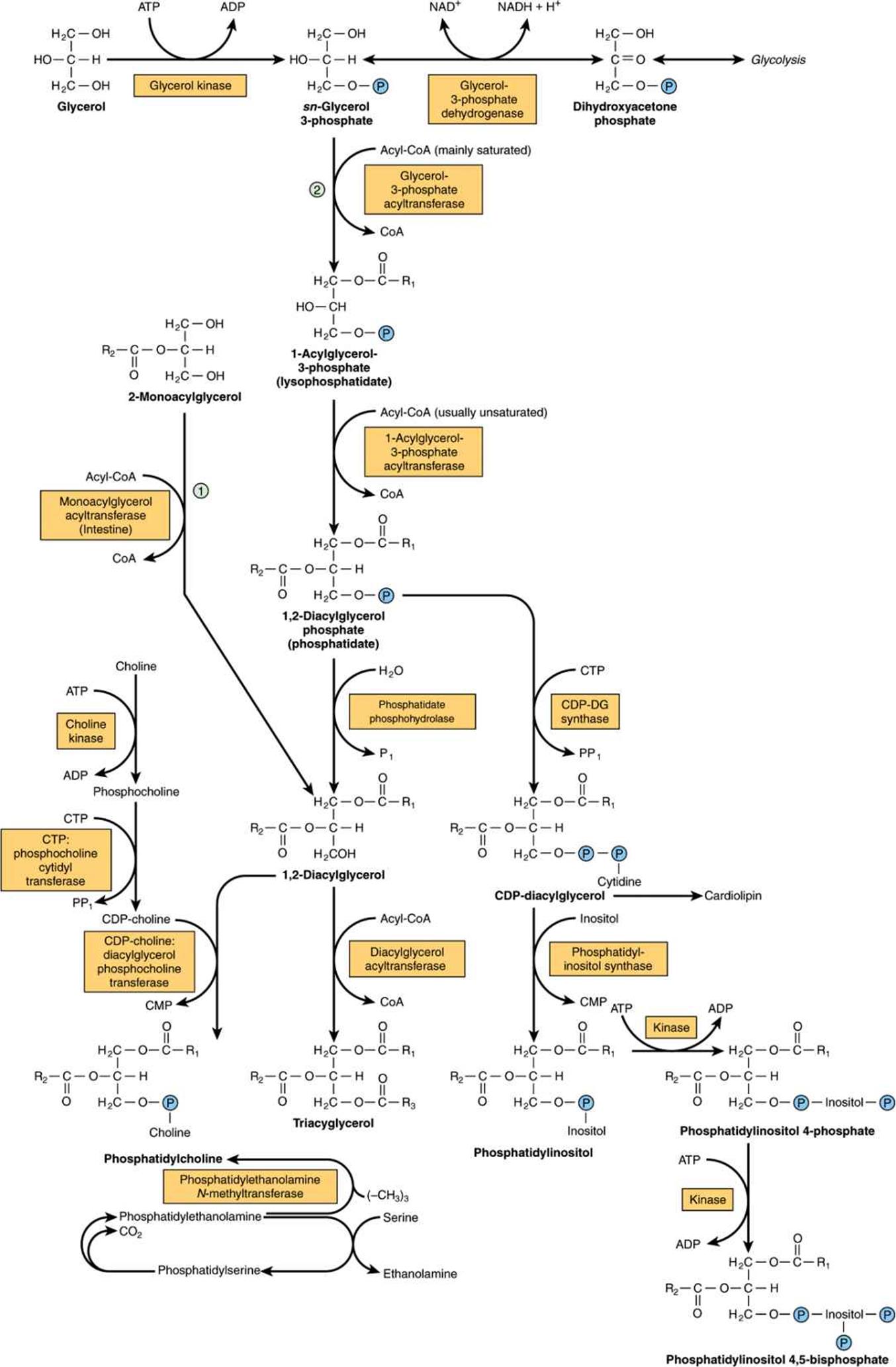
FIGURE 24–2 Biosynthesis of triacylglycerol and phospholipids. (![]() , Monoacylglycerol pathway;
, Monoacylglycerol pathway;![]() , glycerol phosphate pathway.) Phosphatidylethanolamine may be formed from ethanolamine by a pathway similar to that shown for the formation of phosphatidylcholine from choline.
, glycerol phosphate pathway.) Phosphatidylethanolamine may be formed from ethanolamine by a pathway similar to that shown for the formation of phosphatidylcholine from choline.
Biosynthesis of Triacylglycerols
Two molecules of acyl-CoA, formed by the activation of fatty acids by acyl-CoA synthetase (Chapter 22), combine with glycerol 3-phosphate to form phosphatidate (1,2-diacylglycerol phosphate). This takes place in two stages, catalyzed by glycerol-3-phosphate acyltransferase and 1-acylglycerol-3-phosphate acyltransferase. Phosphatidate is converted by phosphatidate phosphohydrolase and diacylglycerol acyltransferase (DGAT) to 1,2-diacylglycerol and then triacylglycerol. DGAT catalyzes the only step specific for triacylglycerol synthesis and is thought to be rate limiting in most circumstances. In intestinal mucosa, monoacylglycerol acyltransferase converts monoacylglycerol to 1,2-diacylglycerol in the monoacylglycerol pathway. Most of the activity of these enzymes resides in the endoplasmic reticulum, but some is found in mitochondria. Although phosphatidate phosphohydrolase protein is found mainly in the cytosol, the active form of the enzyme is membrane bound.
In the biosynthesis of phosphatidylcholine and phosphatidylethanolamine (Figure 24–2), choline or ethanolamine must first be activated by phosphorylation by ATP followed by linkage to CDP. The resulting CDP-choline or CDP-ethanolamine reacts with 1,2-diacylglycerol to form either phosphatidylcholine or phosphatidylethanolamine, respectively. Phosphatidylserine is formed from phosphatidylethanolamine directly by reaction with serine (Figure 24–2). Phosphatidylserine may re-form phosphatidylethanolamine by decarboxylation. An alternative pathway in liver enables phosphatidylethanolamine to give rise directly to phosphatidylcholine by progressive methylation of the ethanolamine residue. In spite of these sources of choline, it is considered to be an essential nutrient in many mammalian species, although this has not been established in humans.
The regulation of triacylglycerol, phosphatidylcholine, and phosphatidylethanolamine biosynthesis is driven by the availability of free fatty acids. Those that escape oxidation are preferentially converted to phospholipids, and when this requirement is satisfied they are used for triacylglycerol synthesis.
Cardiolipin (diphosphatidylglycerol; Figure 15–10) is a phospholipid present in mitochondria. It is formed from phosphatidylglycerol, which in turn is synthesized from CDP-diacylglycerol (Figure 24–2) and glycerol 3-phosphate according to the scheme shown in Figure 24–3. Cardiolipin, found in the inner membrane of mitochondria, has a key role in mitochondrial structure and function, and is also thought to be involved in programmed cell death (apoptosis).
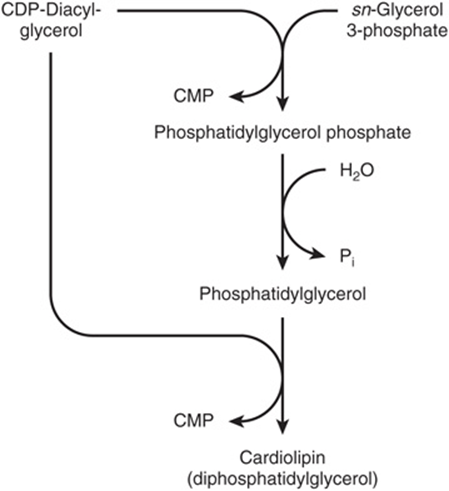
FIGURE 24–3 Biosynthesis of cardiolipin.
Biosynthesis of Glycerol Ether Phospholipids
This pathway is located in peroxisomes. Dihydroxyacetone phosphate is the precursor of the glycerol moiety of glycerol ether phospholipids (Figure 24–4). This compound combines with acyl-CoA to give 1-acyldihydroxyacetone phosphate. The ether link is formed in the next reaction, producing 1-alkyldihydroxyacetone phosphate, which is then converted to 1-alkylglycerol 3-phosphate. After further acylation in the 2 position, the resulting 1-alkyl-2-acylglycerol 3-phosphate (analogous to phosphatidate in Figure 24–2) is hydrolyzed to give the free glycerol derivative. Plasmalogens, which comprise much of the phospholipid in mitochondria, are formed by desaturation of the analogous 3-phosphoethanolamine derivative (Figure 24–4). Platelet-activating factor (PAF) (1-alkyl-2-acetyl-sn-glycerol-3-phosphocholine) is synthesized from the corresponding 3-phosphocholine derivative. It is formed by many blood cells and other tissues and aggregates platelets at concentrations as low as 10-11 mol/L. It also has hypotensive and ulcerogenic properties and is involved in a variety of biologic responses, including inflammation, chemotaxis, and protein phosphorylation.
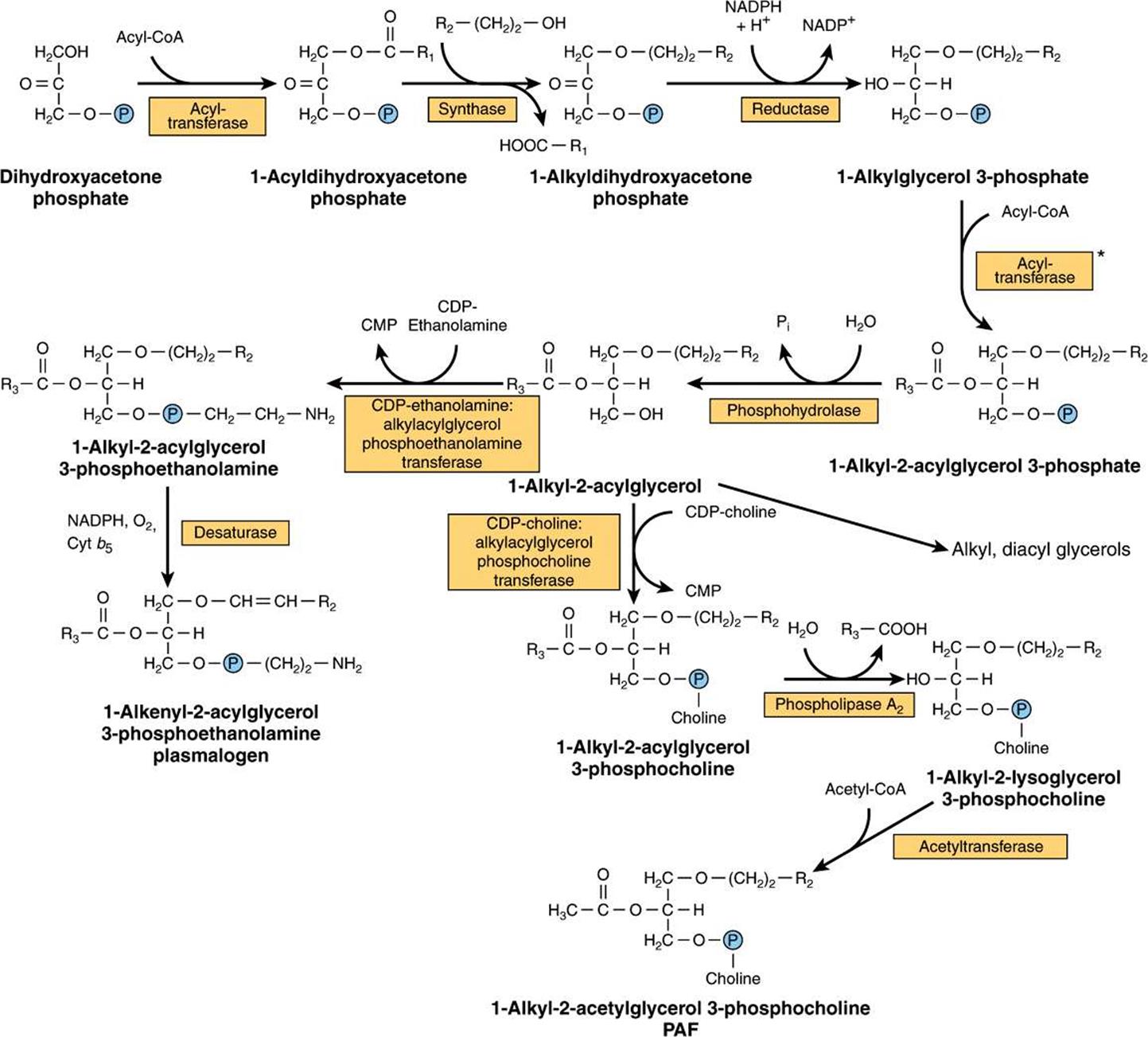
FIGURE 24–4 Biosynthesis of ether lipids, including plasmalogens, and platelet-activating factor (PAF). In the de novo pathway for PAF synthesis, acetyl-CoA is incorporated at stage*, avoiding the last two steps in the pathway shown here.
Phospholipases Allow Degradation & Remodeling of Phosphoglycerols
Although phospholipids are actively degraded, each portion of the molecule turns over at a different rate—eg, the turnover time of the phosphate group is different from that of the 1-acyl group. This is due to the presence of enzymes that allow partial degradation followed by resynthesis ( Figure 24–5s). Phospholipase A2 catalyzes the hydrolysis of glycerophospholipids to form a free fatty acid and lysophospholipid, which in turn may be reacylated by acyl-CoA in the presence of an acyltransferase. Alternatively, lysophospholipid (eg, lysolecithin) is attacked by lysophospholipase, forming the corresponding glyceryl phosphoryl base, which may then be split by a hydrolase liberating glycerol 3-phosphate plus base. Phospholipases A1, A2, B, C, and D attack the bonds indicated in Figure 24–6. Phospholipase A2 is found in pancreatic fluid and snake venom as well as in many types of cells; phospholipase C is one of the major toxins secreted by bacteria; and phospholipase D is known to be involved in mammalian signal transduction.
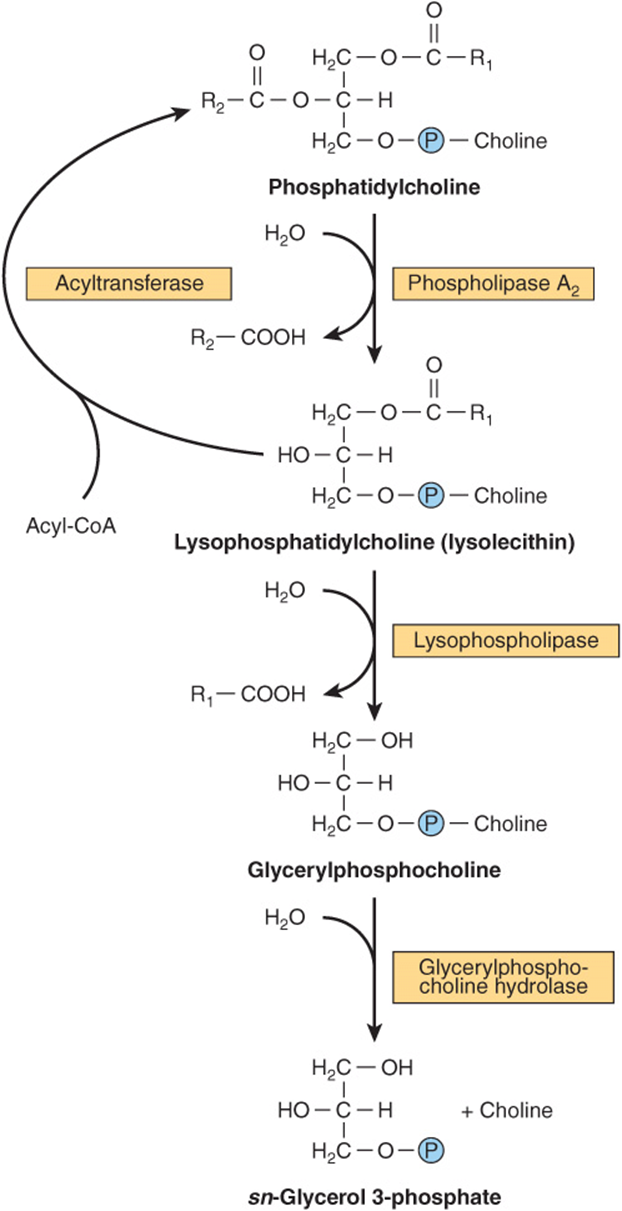
FIGURE 24–5 Metabolism of phosphatidylcholine (lecithin).
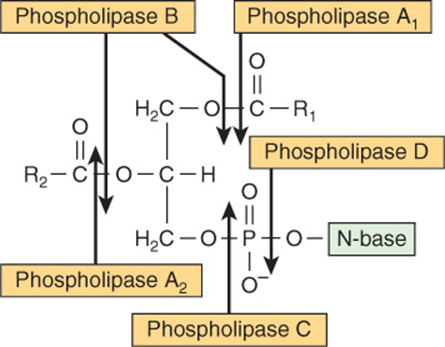
FIGURE 24–6 Sites of the hydrolytic activity of phospholipases on a phospholipid substrate.
Lysolecithin (lysophosphatidylcholine) may be formed by an alternative route that involves lecithin: cholesterol acyltransferase (LCAT). This enzyme, found in plasma, catalyzes the transfer of a fatty acid residue from the 2 position of lecithin to cholesterol to form cholesteryl ester and lysolecithin, and is considered to be responsible for much of the cholesteryl ester in plasma lipoproteins. Long-chain saturated fatty acids are found predominantly in the 1 position of phospholipids, whereas the polyunsaturated fatty acids (eg, the precursors of prostaglandins) are incorporated more frequently into the 2 position. The incorporation of fatty acids into lecithin occurs in three ways; by complete synthesis of the phospholipid; by transacylation between cholesteryl ester and lysolecithin; and by direct acylation of lysolecithin by acyl-CoA. Thus, a continuous exchange of the fatty acids is possible, particularly with regard to introducing essential fatty acids into phospholipid molecules.
ALL SPHINGOLIPIDS ARE FORMED FROM CERAMIDE
Ceramide is synthesized in the endoplasmic reticulum from the amino acid serine as shown in Figure 24–7. Ceramide is an important signaling molecule (second messenger) regulating pathways including programmed cell death (apoptosis), the cell cycle, and cell differentiation and senescence.
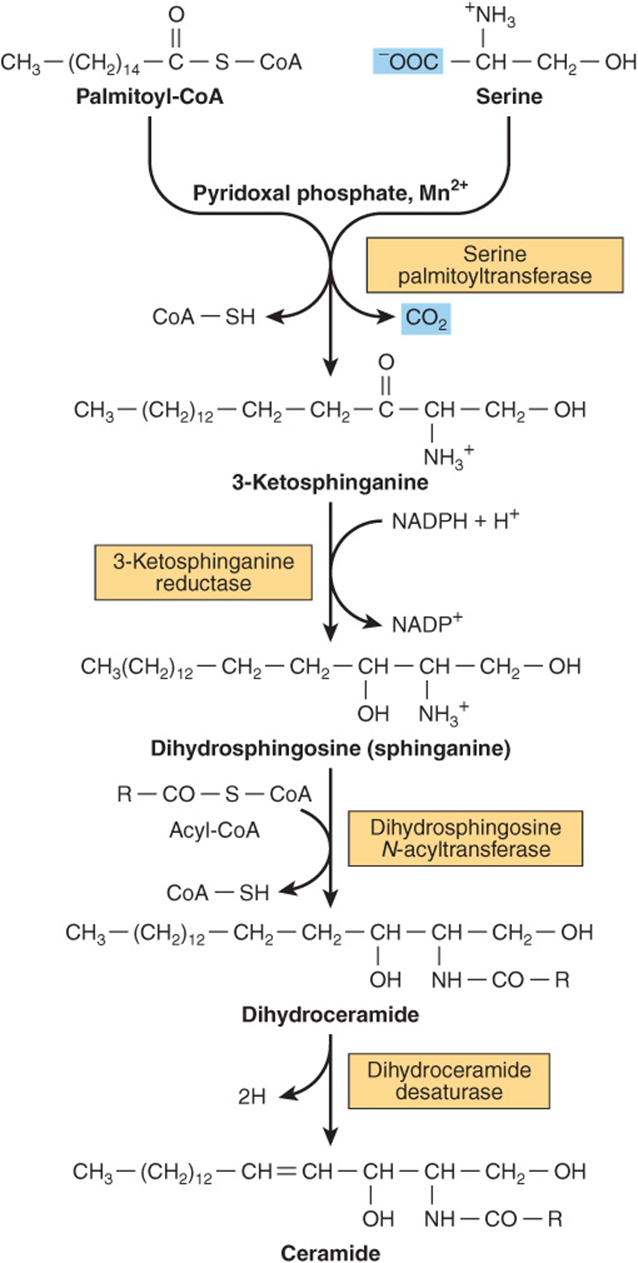
FIGURE 24–7 Biosynthesis of ceramide.
Sphingomyelins (Figure 15–131) are phospholipids and are formed when ceramide reacts with phosphatidylcholine to form sphingomyelin plus diacylglycerol (Figure 24–8A). This occurs mainly in the Golgi apparatus and to a lesser extent in the plasma membrane.
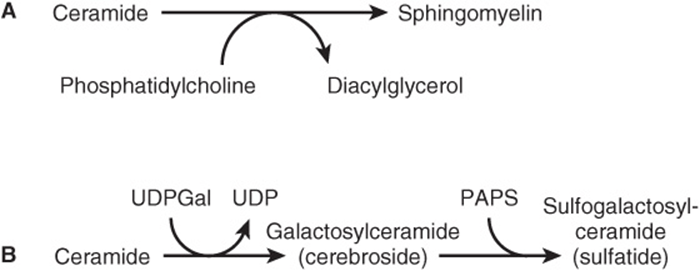
FIGURE 24–8 Biosynthesis of (A) sphingomyelin, (B) galactosylceramide and its sulfo derivative. (PAPS, “active sulfate,” adenosine 3′-phosphate-5′-phosphosulfate.)
Glycosphingolipids Are a Combination of Ceramide with One or More Sugar Residues
The simplest glycosphingolipids (cerebrosides) are galactosylceramide (GalCer) and glucosylceramide (GlcCer). GalCer is a major lipid of myelin, whereas GlcCer is the major glycosphingolipid of extraneural tissues and a precursor of most of the more complex glycosphingolipids. GalCer (Figure 24–8B) is formed in a reaction between ceramide and UDPGal (formed by epimerization from UDPGlc—Figure 21–6).
Sulfogalactosylceramide and other sulfolipids such as the sulfo(galacto)-glycerolipids and the steroid sulfates are formed after further reactions involving 3′-phosphoadenosine-5’-phosphosulfate (PAPS; “active sulfate”). Gangliosides are synthesized from ceramide by the stepwise addition of activated sugars (eg, UDPGlc and UDPGal) and a sialic acid, usually N-acetylneuraminic acid (Figure 24–9). A large number of gangliosides of increasing molecular weight may be formed. Most of the enzymes transferring sugars from nucleotide sugars (glycosyl transferases) are found in the Golgi apparatus.
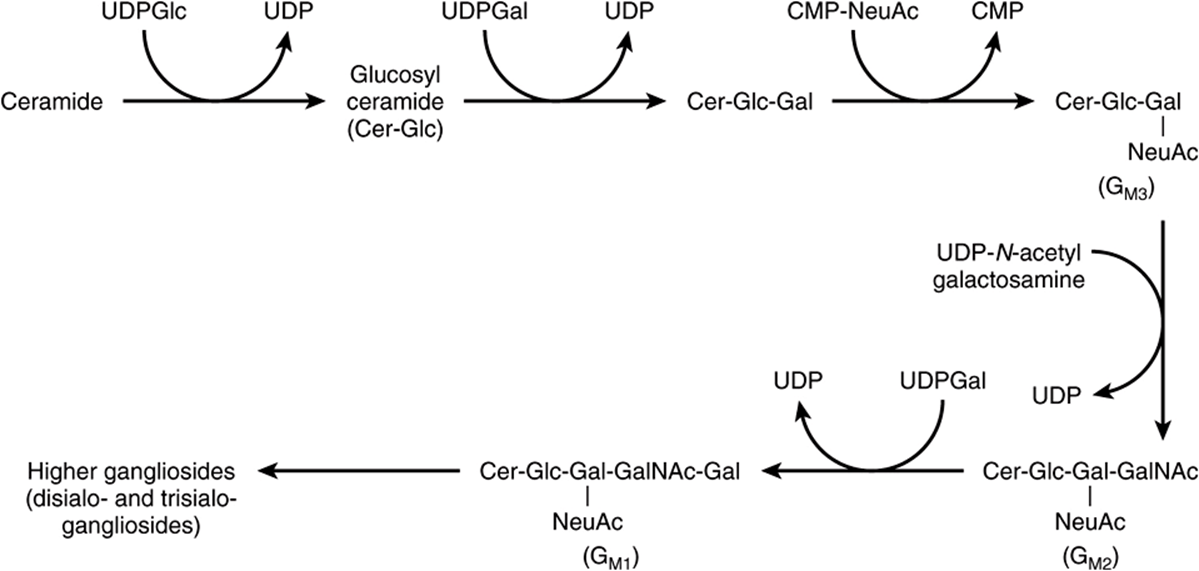
FIGURE 24–9 Biosynthesis of gangliosides. (NeuAc, N-acetylneuraminic acid.)
Glycosphingolipids are constituents of the outer leaflet of plasma membranes and are important in cell adhesion and cell recognition. Some are antigens, eg, ABO blood group substances. Certain gangliosides function as receptors for bacterial toxins (eg, for cholera toxin, which subsequently activates adenylyl cyclase).
CLINICAL ASPECTS
Deficiency of Lung Surfactant Causes Respiratory Distress Syndrome
Lung surfactant is composed mainly of lipid with some proteins and carbohydrate and prevents the alveoli from collapsing. The phospholipid dipalmitoyl-phosphatidylcholine decreases surface tension at the air-liquid interface and thus greatly reduces the work of breathing, but other surfactant lipid and protein components are also important in surfactant function. Deficiency of lung surfactant in the lungs of many preterm newborns gives rise to infant respiratory distress syndrome (IRDS). Administration of either natural or artificial surfactant is of therapeutic benefit.
Phospholipids & Sphingolipids Are Involved in Multiple Sclerosis and Lipidoses
Certain diseases are characterized by abnormal quantities of these lipids in the tissues, often in the nervous system. They may be classified into two groups: (1) true demyelinating diseases and (2) sphingolipidoses.
In multiple sclerosis, which is a demyelinating disease, there is loss of both phospholipids (particularly ethanolamine plasmalogen) and of sphingolipids from white matter. Thus, the lipid composition of white matter resembles that of gray matter. The cerebrospinal fluid shows raised phospholipid levels.
The sphingolipidoses (lipid storage diseases) are a group of inherited diseases that are caused by a genetic defect in the catabolism of lipids containing sphingosine. They are part of a larger group of lysosomal disorders and exhibit several constant features: (1) complex lipids containing ceramide accumulate in cells, particularly neurons, causing neurodegeneration and shortening the lifespan. (2) The rate of synthesis of the stored lipid is normal. (3) The enzymatic defect is in the lysosomal degradation pathway of sphingolipids. (4) The extent to which the activity of the affected enzyme is decreased is similar in all tissues. There is no effective treatment for many of the diseases, although some success has been achieved with enzyme replacement therapy and bone marrow transplantation in the treatment of Gaucher’s and Fabry’s diseases. Other promising approaches are substrate deprivation therapy to inhibit the synthesis of sphingolipids and chemical chaperone therapy. Gene therapy for lysosomal disorders is also currently under investigation. Some examples of the more important lipid storage diseases are shown in Table 24-1.
TABLE 24–1 Examples of Sphingolipidoses
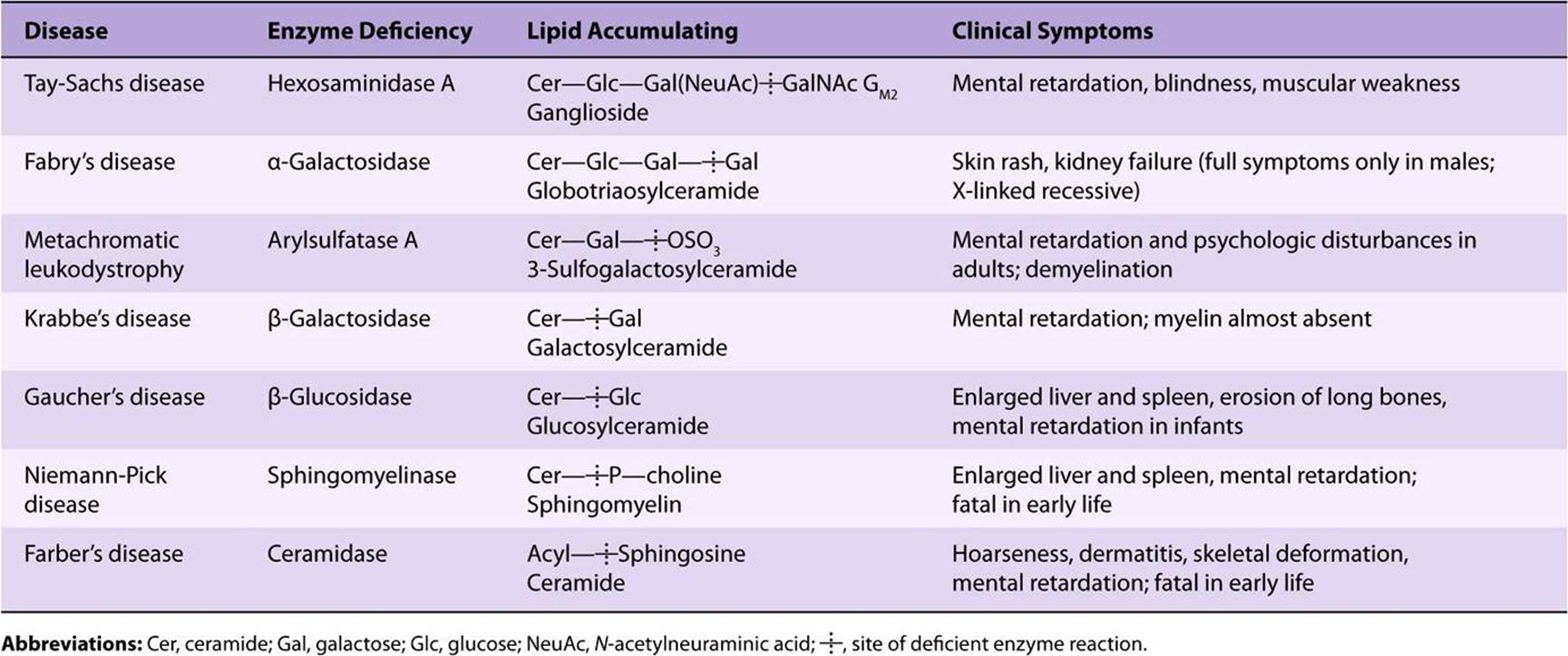
Multiple sulfatase deficiency results in accumulation of sulfogalactosylceramide, steroid sulfates, and proteoglycans owing to a combined deficiency of arylsulfatases A, B, and C and steroid sulfatase.
SUMMARY
![]() Triacylglycerols are the major energy-storing lipids, whereas phosphoglycerols, sphingomyelin, and glycosphingolipids are amphipathic and have structural functions in cell membranes as well as other specialized roles.
Triacylglycerols are the major energy-storing lipids, whereas phosphoglycerols, sphingomyelin, and glycosphingolipids are amphipathic and have structural functions in cell membranes as well as other specialized roles.
![]() Triacylglycerols and some phosphoglycerols are synthesized by progressive acylation of glycerol 3-phosphate. The pathway bifurcates at phosphatidate, forming inositol phospholipids and cardiolipin on the one hand and triacylglycerol and choline and ethanolamine phospholipids on the other.
Triacylglycerols and some phosphoglycerols are synthesized by progressive acylation of glycerol 3-phosphate. The pathway bifurcates at phosphatidate, forming inositol phospholipids and cardiolipin on the one hand and triacylglycerol and choline and ethanolamine phospholipids on the other.
![]() Plasmalogens and platelet-activating factor (PAF) are ether phospholipids formed from dihydroxyacetone phosphate.
Plasmalogens and platelet-activating factor (PAF) are ether phospholipids formed from dihydroxyacetone phosphate.
![]() Sphingolipids are formed from ceramide (N-acylsphingosine). Sphingomyelin is present in membranes of organelles involved in secretory processes (eg, Golgi apparatus). The simplest glycosphingolipids are a combination of ceramide plus a sugar residue (eg, GalCer in myelin). Gangliosides are more complex glycosphingolipids containing more sugar residues plus sialic acid. They are present in the outer layer of the plasma membrane, where they contribute to the glycocalyx and are important as antigens and cell receptors.
Sphingolipids are formed from ceramide (N-acylsphingosine). Sphingomyelin is present in membranes of organelles involved in secretory processes (eg, Golgi apparatus). The simplest glycosphingolipids are a combination of ceramide plus a sugar residue (eg, GalCer in myelin). Gangliosides are more complex glycosphingolipids containing more sugar residues plus sialic acid. They are present in the outer layer of the plasma membrane, where they contribute to the glycocalyx and are important as antigens and cell receptors.
![]() Phospholipids and sphingolipids are involved in several disease processes, including infant respiratory distress syndrome (lack of lung surfactant), multiple sclerosis (demyelination), and sphingolipidoses (inability to break down sphingolipids in lysosomes due to inherited defects in hydrolase enzymes).
Phospholipids and sphingolipids are involved in several disease processes, including infant respiratory distress syndrome (lack of lung surfactant), multiple sclerosis (demyelination), and sphingolipidoses (inability to break down sphingolipids in lysosomes due to inherited defects in hydrolase enzymes).
REFERENCES
McPhail LC: Glycerolipid in signal transduction. Biochemistry of Lipids, Lipoproteins and Membranes, 4th ed. Vance DE, Vance JE (editors). Elsevier, 2002:315-340.
Merrill AH: Sphingolipids. Biochemistry of Lipids, Lipoproteins and Membranes, 5th ed. Vance DE, Vance JE (editors). Elsevier, 2008:363-398.
Meyer KC, Zimmerman JJ: Inflammation and surfactant. Paediatr Respir Rev 2002;3:308.
Prescott SM, Zimmerman GA, Stafforini DM, et al: Platelet-activating factor and related lipid mediators. Annu Rev Biochem 2000;69:419.
Ruvolo PP: Intracellular signal transduction pathways activated by ceramide and its metabolites. Pharmacol Res 2003;47:383.
Scriver CR, Beaudet AL, Sly WS, et al (editors): The Metabolic and Molecular Bases of Inherited Disease, 8th ed. McGraw-Hill, 2001.
Vance DE, Vance JE (editors): Phospholipid biosynthesis in eukaryotes. In: Biochemistry of Lipids, Lipoproteins and Membranes, 5th ed. Elsevier, 2008: 213-244.
van Echten G, Sandhoff K: Ganglioside metabolism. Enzymology, topology, and regulation. J Biol Chem 1993;268:5341.