Harper’s Illustrated Biochemistry, 29th Edition (2012)
Chapter 2. Water & pH
Peter J. Kennelly, PhD & Victor W. Rodwell, PhD
OBJECTIVES
After studying this chapter, you should be able to:
![]() Describe the properties of water that account for its surface tension, viscosity, liquid state at ambient temperature, and solvent power.
Describe the properties of water that account for its surface tension, viscosity, liquid state at ambient temperature, and solvent power.
![]() Use structural formulas to represent several organic compounds that can serve as hydrogen bond donors or acceptors.
Use structural formulas to represent several organic compounds that can serve as hydrogen bond donors or acceptors.
![]() Explain the role played by entropy in the orientation, in an aqueous environment, of the polar and nonpolar regions of macromolecules.
Explain the role played by entropy in the orientation, in an aqueous environment, of the polar and nonpolar regions of macromolecules.
![]() Indicate the quantitative contributions of salt bridges, hydrophobic interactions, and van der Waals forces to the stability of macromolecules.
Indicate the quantitative contributions of salt bridges, hydrophobic interactions, and van der Waals forces to the stability of macromolecules.
![]() Explain the relationship of pH to acidity, alkalinity, and the quantitative determinants that characterize weak and strong acids.
Explain the relationship of pH to acidity, alkalinity, and the quantitative determinants that characterize weak and strong acids.
![]() Calculate the shift in pH that accompanies the addition of a given quantity of acid or base to the pH of a buffered solution.
Calculate the shift in pH that accompanies the addition of a given quantity of acid or base to the pH of a buffered solution.
![]() Describe what buffers do, how they do it, and the conditions under which a buffer is most effective under physiologic or other conditions.
Describe what buffers do, how they do it, and the conditions under which a buffer is most effective under physiologic or other conditions.
![]() Illustrate how the Henderson–Hasselbalch equation can be used to calculate the net charge on a polyelectrolyte at a given pH.
Illustrate how the Henderson–Hasselbalch equation can be used to calculate the net charge on a polyelectrolyte at a given pH.
BIOMEDICAL IMPORTANCE
Water is the predominant chemical component of living organisms. Its unique physical properties, which include the ability to solvate a wide range of organic and inorganic molecules, derive from water’s dipolar structure and exceptional capacity for forming hydrogen bonds. The manner in which water interacts with a solvated biomolecule influences the structure both of the biomolecule and of water itself. An excellent nucleophile, water is a reactant or product in many metabolic reactions. Regulation of water balance depends upon hypothalamic mechanisms that control thirst, on antidiuretic hormone (ADH), on retention or excretion of water by the kidneys, and on evaporative loss. Nephrogenic diabetes insipidus, which involves the inability to concentrate urine or adjust to subtle changes in extracellular fluid osmolarity, results from the unresponsiveness of renal tubular osmoreceptors to ADH.
Water has a slight propensity to dissociate into hydroxide ions and protons. The concentration of protons, or acidity, of aqueous solutions is generally reported using the logarithmic pH scale. Bicarbonate and other buffers normally maintain the pH of extracellular fluid between 7.35 and 7.45. Suspected disturbances of acid–base balance are verified by measuring the pH of arterial blood and the CO2 content of venous blood. Causes of acidosis (blood ![]() ) include diabetic ketosis and lactic acidosis. Alkalosis
) include diabetic ketosis and lactic acidosis. Alkalosis ![]() may follow vomiting of acidic gastric contents.
may follow vomiting of acidic gastric contents.
WATER IS AN IDEAL BIOLOGIC SOLVENT
Water Molecules Form Dipoles
A water molecule is an irregular, slightly skewed tetrahedron with oxygen at its center (Figure 2–1). The two hydrogens and the unshared electrons of the remaining two sp3-hybridized orbitals occupy the corners of the tetrahedron. The 105° angle between the hydrogen differs slightly from the ideal tetrahedral angle, 109.5°. Ammonia is also tetrahedral, with a 107° angle between its hydrogens. The strongly electronegative oxygen atoms in water attract electrons away from the hydrogen nuclei, leaving them with a partial positive charge, while its two unshared electron pairs constitute a region of local negative charge.
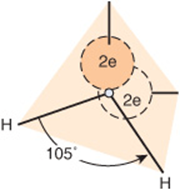
FIGURE 2–1 The water molecule has tetrahedral geometry.
A molecule with electrical charge distributed asymmetrically about its structure is referred to as a dipole. Water’s strong dipole is responsible for its high dielectric constant. As described quantitatively by Coulomb’s law, the strength of interaction F between oppositely charged particles is inversely proportionate to the dielectric constant ε of the surrounding medium. The dielectric constant for a vacuum is unity; for hexane it is 1.9; for ethanol, 24.3; and for water, 78.5. Water therefore greatly decreases the force of attraction between charged and polar species relative to water-free environments with lower dielectric constants. Its strong dipole and high dielectric constant enable water to dissolve large quantities of charged compounds such as salts.
Water Molecules Form Hydrogen Bonds
A partially unshielded hydrogen nucleus covalently bound to an electron-withdrawing oxygen or nitrogen atom can interact with an unshared electron pair on another oxygen or nitrogen atom to form a hydrogen bond. Since water molecules contain both of these features, hydrogen bonding favors the self-association of water molecules into ordered arrays (Figure 2–2). Hydrogen bonding profoundly influences the physical properties of water and accounts for its exceptionally high viscosity, surface tension, and boiling point. On average, each molecule in liquid water associates through hydrogen bonds with 3.5 others. These bonds are both relatively weak and transient, with a half-life of a few nanoseconds or less. Rupture of a hydrogen bond in liquid water requires only about 4.5 kcal/mol, less than 5% of the energy required to rupture a covalent O—H bond.
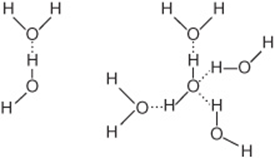
FIGURE 2–2 Left: Association of two dipolar water molecules by a hydrogen bond (dotted line). Right: Hydrogen-bonded cluster of four water molecules. Note that water can serve simultaneously both as a hydrogen donor and as a hydrogen acceptor.
Hydrogen bonding enables water to dissolve many organic biomolecules that contain functional groups which can participate in hydrogen bonding. The oxygen atoms of aldehydes, ketones, and amides, for example, provide lone pairs of electrons that can serve as hydrogen acceptors. Alcohols, carboxylic acids, and amines can serve both as hydrogen acceptors and as donors of unshielded hydrogen atoms for formation of hydrogen bonds (Figure 2–3).
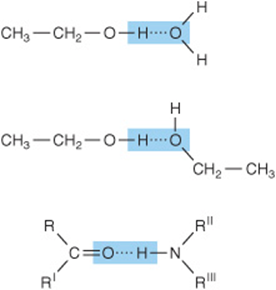
FIGURE 2–3 Additional polar groups participate in hydrogen bonding. Shown are hydrogen bonds formed between alcohol and water, between two molecules of ethanol, and between the peptide carbonyl oxygen and the peptide nitrogen hydrogen of an adjacent amino acid.
INTERACTION WITH WATER INFLUENCES THE STRUCTURE OF BIOMOLECULES
Covalent and Noncovalent Bonds Stabilize Biologic Molecules
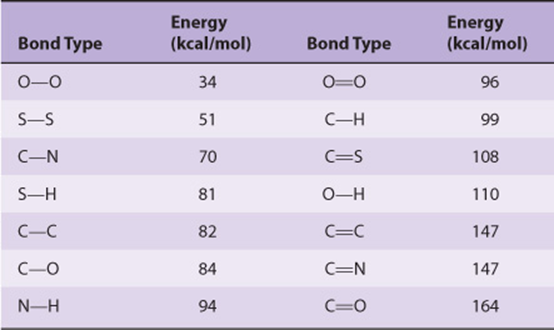
The covalent bond is the strongest force that holds molecules together (Table 2–1). Noncovalent forces, while of lesser magnitude, make significant contributions to the structure, stability, and functional competence of macromolecules in living cells. These forces, which can be either attractive or repulsive, involve interactions both within the biomolecule and between it and the water that forms the principal component of the surrounding environment.
TABLE 2–1 Bond Energies for Atoms of Biologic Significance
Biomolecules Fold to Position Polar & Charged Groups on Their Surfaces
Most biomolecules are amphipathic; that is, they possess regions rich in charged or polar functional groups as well as regions with hydrophobic character. Proteins tend to fold with the R-groups of amino acids with hydrophobic side chains in the interior. Amino acids with charged or polar amino acid side chains (eg, arginine, glutamate, serine) generally are present on the surface in contact with water. A similar pattern prevails in a phospholipid bilayer, where the charged “head groups” of phosphatidyl serine or phosphatidyl ethanolamine contact water while their hydrophobic fatty acyl side chains cluster together, excluding water. This pattern maximizes the opportunities for the formation of energetically favorable charge–dipole, dipole–dipole, and hydrogen bonding interactions between polar groups on the biomolecule and water. It also minimizes energetically unfavorable contacts between water and hydrophobic groups.
Hydrophobic Interactions
Hydrophobic interaction refers to the tendency of nonpolar compounds to self-associate in an aqueous environment. This self-association is driven neither by mutual attraction nor by what are sometimes incorrectly referred to as “hydrophobic bonds.” Self-association minimizes the disruption of energetically favorable interactions between the surrounding water molecules.
While the hydrogens of nonpolar groups such as the methylene groups of hydrocarbons do not form hydrogen bonds, they do affect the structure of the water that surrounds them. Water molecules adjacent to a hydrophobic group are restricted in the number of orientations (degrees of freedom) that permit them to participate in the maximum number of energetically favorable hydrogen bonds. Maximal formation of multiple hydrogen bonds, which maximizes enthalpy, can be maintained only by increasing the order of the adjacent water molecules, with an accompanying decrease in entropy.
It follows from the second law of thermodynamics that the optimal free energy of a hydrocarbon–water mixture is a function of both maximal enthalpy (from hydrogen bonding) and minimum entropy (maximum degrees of freedom). Thus, nonpolar molecules tend to form droplets that minimize exposed surface area and reduce the number of water molecules whose motional freedom becomes restricted. Similarly, in the aqueous environment of the living cell the hydrophobic portions of biopolymers tend to be buried inside the structure of the molecule, or within a lipid bilayer, minimizing contact with water.
Electrostatic Interactions
Interactions between charged groups help shape biomolecular structure. Electrostatic interactions between oppositely charged groups within or between biomolecules are termed salt bridges. Salt bridges are comparable in strength to hydrogen bonds but act over larger distances. They therefore often facilitate the binding of charged molecules and ions to proteins and nucleic acids.
van der Waals Forces
van der Waals forces arise from attractions between transient dipoles generated by the rapid movement of electrons of all neutral atoms. Significantly weaker than hydrogen bonds but potentially extremely numerous, van der Waals forces decrease as the sixth power of the distance separating atoms (Figure 2–4). Thus, they act over very short distances, typically 2–4 Å.
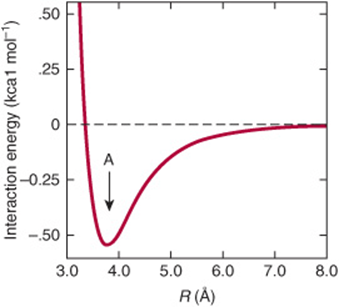
FIGURE 2–4 The strength of van der Waals interactions varies with the distance, R, between interacting species. The force of interaction between interacting species increases with decreasing distance until they are separated by the van der Waals contact distance (see arrow marked A). Repulsion due to interaction between the electrons of each atom or molecule then supervenes. While individual van der Waals interactions are extremely weak, the cumulative effect is nevertheless substantial for macromolecules such as DNA and proteins with many atoms in close contact.
Multiple Forces Stabilize Biomolecules
The DNA double helix illustrates the contribution of multiple forces to the structure of biomolecules. While each individual DNA strand is held together by covalent bonds, the two strands of the helix are held together exclusively by noncovalent interactions such as hydrogen bonds between nucleotide bases (Watson–Crick base pairing) and van der Waals interactions between the stacked purine and pyrimidine bases. The double helix presents the charged phosphate groups and polar hydroxyl groups from the ribose sugars of the DNA backbone to water while burying the relatively hydrophobic nucleotide bases inside. The extended backbone maximizes the distance between negatively charged phosphates, minimizing unfavorable electrostatic interactions.
WATER IS AN EXCELLENT NUCLEOPHILE
Metabolic reactions often involve the attack by lone pairs of electrons residing on electron-rich molecules termed nucleophiles upon electron-poor atoms called electrophiles. Nucleophiles and electrophiles do not necessarily possess a formal negative or positive charge. Water, whose two lone pairs of sp3 electrons bear a partial negative charge (Figure 2–1), is an excellent nucleophile. Other nucleophiles of biologic importance include the oxygen atoms of phosphates, alcohols, and carboxylic acids; the sulfur of thiols; and the nitrogen of amines and the imidazole ring of histidine. Common electrophiles include the carbonyl carbons in amides, esters, aldehydes, and ketones and the phosphorus atoms of phosphoesters.
Nucleophilic attack by water typically results in the cleavage of the amide, glycoside, or ester bonds that hold biopolymers together. This process is termed hydrolysis. Conversely, when monomer units are joined together to form biopolymers such as proteins or glycogen, water is a product, for example, during the formation of a peptide bond between two amino acids:
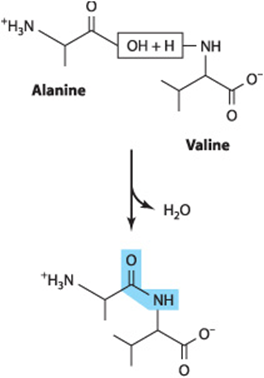
While hydrolysis is a thermodynamically favored reaction, the amide and phosphoester bonds of polypeptides and oligonucleotides are stable in the aqueous environment of the cell. This seemingly paradoxic behavior reflects the fact that the thermodynamics governing the equilibrium of a reaction do not determine the rate at which it will proceed. In the cell, protein catalysts called enzymes accelerate the rate of hydrolytic reactions when needed. Proteasescatalyze the hydrolysis of proteins into their component amino acids, while nucleases catalyze the hydrolysis of the phosphoester bonds in DNA and RNA. Careful control of the activities of these enzymes is required to ensure that they act only on appropriate target molecules at appropriate times.
Many Metabolic Reactions Involve Group Transfer
Many of the enzymic reactions responsible for synthesis and breakdown of biomolecules involve the transfer of a chemical group G from a donor D to an acceptor A to form an acceptor group complex, A—G:
![]()
The hydrolysis and phosphorolysis of glycogen, for example, involve the transfer of glucosyl groups to water or to orthophosphate. The equilibrium constant for the hydrolysis of covalent bonds strongly favors the formation of split products. Conversely, in many cases the group transfer reactions responsible for the biosynthesis of macromolecules involve the thermodynamically unfavored formation of covalent bonds. Enzyme catalysts play a critical role in surmounting these barriers by virtue of their capacity to directly link two normally separate reactions together. By marrying an energetically unfavorable group transfer reaction with a thermodynamically favorable reaction, such as the hydrolysis of ATP, a new coupled reaction can be generated whose net overall change in free energy favors biopolymer synthesis.
Given the nucleophilic character of water and its high concentration in cells, why are biopolymers such as proteins and DNA relatively stable? And how can synthesis of biopolymers occur in an aqueous, seemingly prohydrolytic, environment? Central to both questions are the properties of enzymes. In the absence of enzymic catalysis, even reactions that are highly favored thermodynamically do not necessarily take place rapidly. Precise and differential control of enzyme activity and the sequestration of enzymes in specific organelles determine under what physiologic conditions a given biopolymer will be synthesized or degraded. Newly synthesized biopolymers are not immediately hydrolyzed because the active sites of biosynthetic enzymes sequester substrates in an environment from which water can be excluded.
Water Molecules Exhibit a Slight but Important Tendency to Dissociate
The ability of water to ionize, while slight, is of central importance for life. Since water can act both as an acid and as a base, its ionization may be represented as an intermolecular proton transfer that forms a hydronium ion (H3O+) and a hydroxide ion (OH–):
![]()
The transferred proton is actually associated with a cluster of water molecules. Protons exist in solution not only as H3O+, but also as multimers such as ![]() and
and ![]() The proton is nevertheless routinely represented as H+, even though it is in fact highly hydrated.
The proton is nevertheless routinely represented as H+, even though it is in fact highly hydrated.
Since hydronium and hydroxide ions continuously recombine to form water molecules, an individual hydrogen or oxygen cannot be stated to be present as an ion or as part of a water molecule. At one instant it is an ion; an instant later it is part of a water molecule. Individual ions or molecules are therefore not considered. We refer instead to the probability that at any instant in time a given hydrogen will be present as an ion or as part of a water molecule. Since 1 g of water contains 3.46 × 1022 molecules, the ionization of water can be described statistically. To state that the probability that a hydrogen exists as an ion is 0.01 means that at any given moment in time, a hydrogen atom has 1 chance in 100 of being an ion and 99 chances out of 100 of being part of a water molecule. The actual probability of a hydrogen atom in pure water existing as a hydrogen ion is approximately 1.8 × 10–9. The probability of its being part of a water molecule thus is almost unity. Stated another way, for every hydrogen ion or hydroxide ion in pure water, there are 1.8 billion or 1.8 × 109 water molecules. Hydrogen ions and hydroxide ions nevertheless contribute significantly to the properties of water.
For dissociation of water,
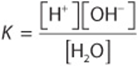
where the brackets represent molar concentrations (strictly speaking, molar activities) and K is the dissociation constant. Since 1 mole (mol) of water weighs 18 g, 1 liter (L) (1000 g) of water contains ![]() mol. Pure water thus is 55.56 molar. Since the probability that a hydrogen in pure water will exist as a hydrogen ion is 1.8 × 10–9, the molar concentration of H+ ions (or of OH– ions) in pure water is the product of the probability, 1.8 × 10–9, times the molar concentration of water, 55.56 mol/L. The result is 1.0 × 10–7 mol/L.
mol. Pure water thus is 55.56 molar. Since the probability that a hydrogen in pure water will exist as a hydrogen ion is 1.8 × 10–9, the molar concentration of H+ ions (or of OH– ions) in pure water is the product of the probability, 1.8 × 10–9, times the molar concentration of water, 55.56 mol/L. The result is 1.0 × 10–7 mol/L.
We can now calculate K for pure water:
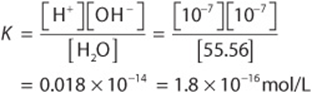
The molar concentration of water, 55.56 mol/L, is too great to be significantly affected by dissociation. It is therefore considered to be essentially constant. This constant may therefore be incorporated into the dissociation constant K to provide a useful new constant Kw termed the ion product for water. The relationship between Kw and K is shown below:
Note that the dimensions of K are moles per liter and those of Kw are moles2 per liter2. As its name suggests, the ion product Kw is numerically equal to the product of the molar concentrations of H+ and OH–:
![]()
At 25°C, ![]() , or 10–14 (mol/L)2. At temperatures below 25°C, Kw is somewhat less than 10–14, and at temperatures above 25°C it is somewhat greater than 10–14. Within the stated limitations of the effect of temperature, Kw equals 10–14 (mol/L)2 for all aqueous solutions, even solutions of acids or bases. We use Kw to calculate the pH of acidic and basic solutions.
, or 10–14 (mol/L)2. At temperatures below 25°C, Kw is somewhat less than 10–14, and at temperatures above 25°C it is somewhat greater than 10–14. Within the stated limitations of the effect of temperature, Kw equals 10–14 (mol/L)2 for all aqueous solutions, even solutions of acids or bases. We use Kw to calculate the pH of acidic and basic solutions.
pH IS THE NEGATIVE LOG OF THE HYDROGEN ION CONCENTRATION
The term pH was introduced in 1909 by Sörensen, who defined pH as the negative log of the hydrogen ion concentration:
![]()
This definition, while not rigorous, suffices for many biochemical purposes. To calculate the pH of a solution:
1. Calculate the hydrogen ion concentration [H+].
2. Calculate the base 10 logarithm of [H+].
3. pH is the negative of the value found in step 2.
For example, for pure water at 25°C,
![]()
This value is also known as the power (English), puissant (French), or potennz (German) of the exponent, hence the use of the term “p.”
Low pH values correspond to high concentrations of H+ and high pH values correspond to low concentrations of H+.
Acids are proton donors and bases are proton acceptors. Strong acids (eg, HCl, H2SO4) completely dissociate into anions and protons even in strongly acidic solutions (low pH). Weak acids dissociate only partially in acidic solutions. Similarly, strong bases (eg, KOH, NaOH)—but not weak bases, eg: Ca(OH)2, are completely dissociated even at high pH. Many biochemicals are weak acids. Exceptions include phosphorylated intermediates, whose phosphoryl group contains two dissociable protons, the first of which is strongly acidic.
The following examples illustrate how to calculate the pH of acidic and basic solutions.
Example 1: What is the pH of a solution whose hydrogen ion concentration is 3.2 × 10–4 mol/L?
Example 2: What is the pH of a solution whose hydroxide ion concentration is 4.0 × 10–4 mol/L? We first define a quantity pOH that is equal to –log [OH–] and that may be derived from the definition of Kw :
![]()
Therefore,
![]()
or
![]()
To solve the problem by this approach:
Now
![]()
The examples above illustrate how the logarithmic pH scale facilitates recording and comparing hydrogen ion concentrations that differ by orders of magnitude from one another, ie, 0.00032 M (pH 3.5) and 0.000000000025 M (pH 10.6).
Example 3: What are the pH values of (a) 2.0 × 10–2 mol/L KOH and of (b) 2.0 × 10–6 mol/L KOH? The OH– arises from two sources, KOH and water. Since pH is determined by the total [H+] (and pOH by the total [OH–]), both sources must be considered. In the first case (a), the contribution of water to the total [OH–] is negligible. The same cannot be said for the second case (b):
Once a decision has been reached about the significance of the contribution by water, pH may be calculated as above.
The above examples assume that the strong base KOH is completely dissociated in solution and that the concentration of OH– ions was thus equal to that due to the KOH plus that present initially in the water. This assumption is valid for dilute solutions of strong bases or acids, but not for weak bases or acids. Since weak electrolytes dissociate only slightly in solution, we must use the dissociation constant to calculate the concentration of [H+] (or [OH–]) produced by a given molarity of a weak acid (or base) before calculating total [H+] (or total [OH–]) and subsequently pH.
Functional Groups That Are Weak Acids Have Great Physiologic Significance
Many biochemicals possess functional groups that are weak acids or bases. Carboxyl groups, amino groups, and phosphate esters, whose second dissociation falls within the physiologic range, are present in proteins and nucleic acids, most coenzymes, and most intermediary metabolites. Knowledge of the dissociation of weak acids and bases thus is basic to understanding the influence of intracellular pH on structure and biologic activity. Charge-based separations such as electrophoresis and ion exchange chromatography are also best understood in terms of the dissociation behavior of functional groups.
We term the protonated species (eg, HA or ![]() ) the acid and the unprotonated species (eg, A– or R—NH2) its conjugate base. Similarly, we may refer to a base (eg, A– or
) the acid and the unprotonated species (eg, A– or R—NH2) its conjugate base. Similarly, we may refer to a base (eg, A– or ![]() ) and its conjugate acid (eg, HA or
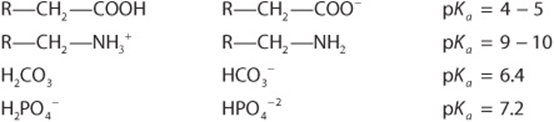
) and its conjugate acid (eg, HA or ![]() ). Representative weak acids (left), their conjugate bases (center), and pKa values (right) include the following:
). Representative weak acids (left), their conjugate bases (center), and pKa values (right) include the following:
We express the relative strengths of weak acids and bases in terms of their dissociation constants. Shown below are the expressions for the dissociation constant (Ka) for two representative weak acids, R—COOH and ![]()
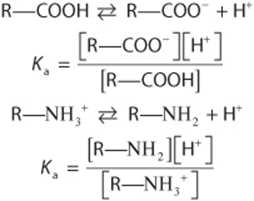
Since the numeric values of Ka for weak acids are negative exponential numbers, we express Ka as pKa, where
![]()
Note that pKa is related to Ka as pH is to [H+]. The stronger the acid, the lower is its pKa value.
pKa is used to express the relative strengths of both acids and bases. For any weak acid, its conjugate is a strong base. Similarly, the conjugate of a strong base is a weak acid. The relative strengths of bases are expressed in terms of the pKa of their conjugate acids. For polyprotic compounds containing more than one dissociable proton, a numerical subscript is assigned to each dissociation, numbered starting from unity in decreasing order of relative acidity. For a dissociation of the type
![]()
the pKa is the pH at which the concentration of the acid ![]() equals that of the base R—NH2.
equals that of the base R—NH2.
From the above equations that relate Ka to [H+] and to the concentrations of undissociated acid and its conjugate base, when
![]()
or when
![]()
then
![]()
Thus, when the associated (protonated) and dissociated (conjugate base) species are present at equal concentrations, the prevailing hydrogen ion concentration [H+] is numerically equal to the dissociation constant, Ka. If the logarithms of both sides of the above equation are taken and both sides are multiplied by –1, the expressions would be as follows:
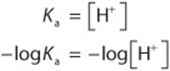
Since –log Ka is defined as pKa, and –log [H+] defines pH, the equation may be rewritten as
![]()
ie, the pKa of an acid group is the pH at which the protonated and unprotonated species are present at equal concentrations. The pKa for an acid may be determined by adding 0.5 equivalent of alkali per equivalent of acid. The resulting pH will equal the pKa of the acid.
The Henderson-Hasselbalch Equation Describes the Behavior of Weak Acids & Buffers
The Henderson–Hasselbalch equation is derived below.
A weak acid, HA, ionizes as follows:
![]()
The equilibrium constant for this dissociation is
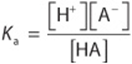
Cross-multiplication gives
![]()
Divide both sides by [A–]:
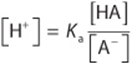
Take the log of both sides:
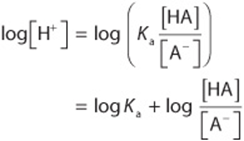
Multiply through by –1:
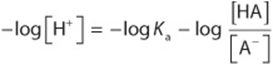
Substitute pH and pKa for –log [H+] and –log Ka, respectively; then
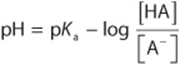
Inversion of the last term removes the minus sign and gives the Henderson–Hasselbalch equation
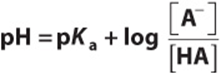
The Henderson–Hasselbalch equation has great predictive value in protonic equilibria. For example,
1. When an acid is exactly half-neutralized, ![]() . Under these conditions,
. Under these conditions,
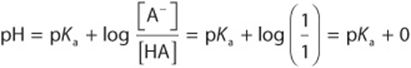
Therefore, at half-neutralization, ![]() .
.
2. When the ratio ![]() ,
,
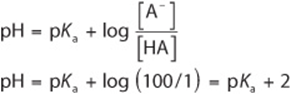
3. When the ratio ![]() ,
,
![]()
If the equation is evaluated at ratios of [A–]/[HA] ranging from 103 to 10–3 and the calculated pH values are plotted, the resulting graph describes the titration curve for a weak acid (Figure 2–5).
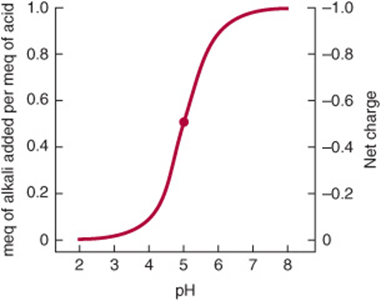
FIGURE 2–5 Titration curve for an acid of the type HA. The heavy dot in the center of the curve indicates the pKa 5.0.
Solutions of Weak Acids & Their Salts Buffer Changes in pH
Solutions of weak acids or bases and their conjugates exhibit buffering, the ability to resist a change in pH following addition of strong acid or base. Since many metabolic reactions are accompanied by the release or uptake of protons, most intracellular reactions are buffered. Oxidative metabolism produces CO2, the anhydride of carbonic acid, which if not buffered would produce severe acidosis. Maintenance of a constant pH involves buffering by phosphate, bicarbonate, and proteins, which accept or release protons to resist a change in pH. For experiments using tissue extracts or enzymes, constant pH is maintained by the addition of buffers such as MES ([2-N-morpholino]-ethanesulfonic acid, pKa 6.1), inorganic orthophosphate (pKa2 7.2), HEPES (N-hydroxyethylpiperazine-N’-2-ethanesulfonic acid, pKa 6.8), or Tris (tris[hydroxymethyl]aminomethane, pKa 8.3). The value of pKa relative to the desired pH is the major determinant of which buffer is selected.
Buffering can be observed by using a pH meter while titrating a weak acid or base (Figure 2–5). We can also calculate the pH shift that accompanies addition of acid or base to a buffered solution. In the example, the buffered solution (a weak acid, ![]() , and its conjugate base) is initially at one of four pH values. We will calculate the pH shift that results when 0.1 meq of KOH is added to 1 meq of each solution:
, and its conjugate base) is initially at one of four pH values. We will calculate the pH shift that results when 0.1 meq of KOH is added to 1 meq of each solution:
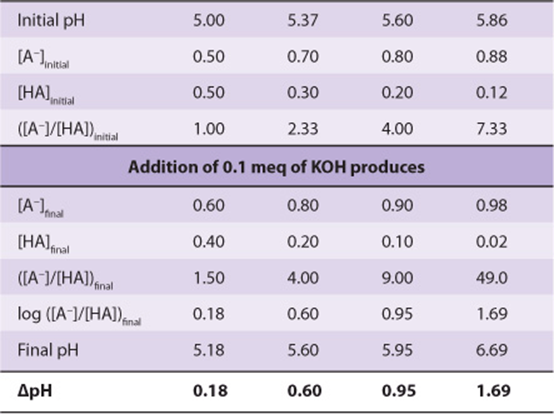
Notice that the change in pH per milliequivalent of OH– added depends on the initial pH. The solution resists changes in pH most effectively at pH values close to the pKa. A solution of a weak acid and its conjugate base buffers most effectively in the pH range ![]() pH unit.
pH unit.
Figure 2–5 also illustrates the net charge on one molecule of the acid as a function of pH. A fractional charge of –0.5 does not mean that an individual molecule bears a fractional charge but that the probability is 0.5 that a given molecule has a unit negative charge at any given moment in time. Consideration of the net charge on macromolecules as a function of pH provides the basis for separatory techniques such as ion exchange chromatography and electrophoresis.
Acid Strength Depends on Molecular Structure
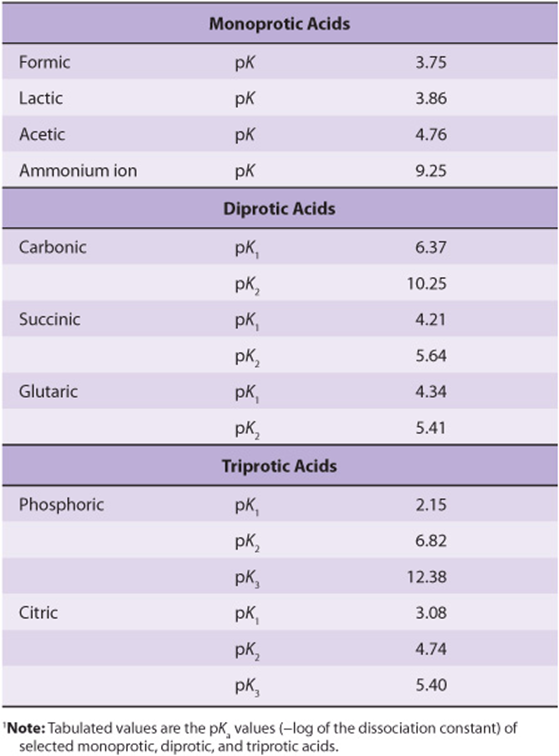
Many acids of biologic interest possess more than one dissociating group. The presence of adjacent negative charge hinders the release of a proton from a nearby group, raising its pKa. This is apparent from the pKa values for the three dissociating groups of phosphoric acid and citric acid (Table 2–2). The effect of adjacent charge decreases with distance. The second pKa for succinic acid, which has two methylene groups between its carboxyl groups, is 5.6, whereas the second pKa for glutaric acid, which has one additional methylene group, is 5.4.
TABLE 2–2 Relative Strengths of Selected Acids of Biologic Significance1
pKa Values Depend on the Properties of the Medium
The pKa of a functional group is also profoundly influenced by the surrounding medium. The medium may either raise or lower the pKa depending on whether the undissociated acid or its conjugate base is the charged species. The effect of dielectric constant on pKa may be observed by adding ethanol to water. The pKa of a carboxylic acid increases, whereas that of an amine decreases because ethanol decreases the ability of water to solvate a charged species. The pKa values of dissociating groups in the interiors of proteins thus are profoundly affected by their local environment, including the presence or absence of water.
SUMMARY
![]() Water forms hydrogen-bonded clusters with itself and with other proton donors or acceptors. Hydrogen bonds account for the surface tension, viscosity, liquid state at room temperature, and solvent power of water.
Water forms hydrogen-bonded clusters with itself and with other proton donors or acceptors. Hydrogen bonds account for the surface tension, viscosity, liquid state at room temperature, and solvent power of water.
![]() Compounds that contain O or N can serve as hydrogen bond donors and/or acceptors.
Compounds that contain O or N can serve as hydrogen bond donors and/or acceptors.
![]() Macromolecules exchange internal surface hydrogen bonds for hydrogen bonds to water. Entropic forces dictate that macromolecules expose polar regions to an aqueous interface and bury nonpolar regions.
Macromolecules exchange internal surface hydrogen bonds for hydrogen bonds to water. Entropic forces dictate that macromolecules expose polar regions to an aqueous interface and bury nonpolar regions.
![]() Salt bridges, hydrophobic interactions, and van der Waals forces participate in maintaining molecular structure.
Salt bridges, hydrophobic interactions, and van der Waals forces participate in maintaining molecular structure.
![]() pH is the negative log of [H+]. A low pH characterizes an acidic solution, and a high pH denotes a basic solution.
pH is the negative log of [H+]. A low pH characterizes an acidic solution, and a high pH denotes a basic solution.
![]() The strength of weak acids is expressed by pKa, the negative log of the acid dissociation constant. Strong acids have low pKa values and weak acids have high pKa values.
The strength of weak acids is expressed by pKa, the negative log of the acid dissociation constant. Strong acids have low pKa values and weak acids have high pKa values.
![]() Buffers resist a change in pH when protons are produced or consumed. Maximum buffering capacity occurs ± 1 pH unit on either side of pKa. Physiologic buffers include bicarbonate, orthophosphate, and proteins.
Buffers resist a change in pH when protons are produced or consumed. Maximum buffering capacity occurs ± 1 pH unit on either side of pKa. Physiologic buffers include bicarbonate, orthophosphate, and proteins.
REFERENCES
Reese KM: Whence came the symbol pH. Chem & Eng News 2004;82:64.
Segel IM: Biochemical Calculations. Wiley, 1968.
Skinner JL: Following the motions of water molecules in aqueous solutions. Science 2010;328:985.
Stillinger FH: Water revisited. Science 1980;209:451. Suresh SJ, Naik VM: Hydrogen bond thermodynamic properties of water from dielectric constant data. J Chem Phys 2000;113:9727.
Wiggins PM: Role of water in some biological processes. Microbiol Rev 1990;54:432.