Harper’s Illustrated Biochemistry, 29th Edition (2012)
SECTION IV. Structure, Function, & Replication of Informational Macromolecules
Chapter 32. Nucleotides
Victor W. Rodwell, PhD
OBJECTIVES
After studying this chapter, you should be able to:
![]() Write structural formulas to represent the amino- and oxo-tautomers of a purine and of a pyrimidine and state which tautomer predominates under physiologic conditions.
Write structural formulas to represent the amino- and oxo-tautomers of a purine and of a pyrimidine and state which tautomer predominates under physiologic conditions.
![]() Reproduce the structural formulas for the principal nucleotides present in DNA and in RNA and the less common nucleotides 5-methylcytosine, 5-hydroxymethylcytosine, and pseudouridine (Ψ).
Reproduce the structural formulas for the principal nucleotides present in DNA and in RNA and the less common nucleotides 5-methylcytosine, 5-hydroxymethylcytosine, and pseudouridine (Ψ).
![]() Represent D-ribose or 2-deoxy-D-ribose linked as either a syn or an anti conformer of a purine, name the bond between the sugar and the base, and indicate which conformer predominates under most physiologic conditions.
Represent D-ribose or 2-deoxy-D-ribose linked as either a syn or an anti conformer of a purine, name the bond between the sugar and the base, and indicate which conformer predominates under most physiologic conditions.
![]() Number the C and N atoms of a pyrimidine nucleotide and of a purine nucleoside, including using a primed numeral for C atoms of the sugars.
Number the C and N atoms of a pyrimidine nucleotide and of a purine nucleoside, including using a primed numeral for C atoms of the sugars.
![]() Compare the phosphoryl group transfer potential of each phosphoryl group of a nucleoside triphosphate.
Compare the phosphoryl group transfer potential of each phosphoryl group of a nucleoside triphosphate.
![]() Outline the physiologic roles of the cyclic phosphodiesters cAMP and cGMP.
Outline the physiologic roles of the cyclic phosphodiesters cAMP and cGMP.
![]() Appreciate that polynucleotides are directional macromolecules composed of mononucleotides linked by
Appreciate that polynucleotides are directional macromolecules composed of mononucleotides linked by ![]() -phosphodiester bonds.
-phosphodiester bonds.
![]() Understand that in the abbreviated representations of polynucleotide structures such as pTpGpT or TGCATCA, the 5′-end is always shown at the left and all phosphodiester bonds are
Understand that in the abbreviated representations of polynucleotide structures such as pTpGpT or TGCATCA, the 5′-end is always shown at the left and all phosphodiester bonds are ![]() .
.
![]() For specific synthetic analogs of purine and pyrimidine bases and their derivatives that have served as anticancer drugs, indicate in what ways these compounds inhibit metabolism.
For specific synthetic analogs of purine and pyrimidine bases and their derivatives that have served as anticancer drugs, indicate in what ways these compounds inhibit metabolism.
BIOMEDICAL IMPORTANCE
In addition to serving as precursors of nucleic acids, purine and pyrimidine nucleotides participate in metabolic functions as diverse as energy metabolism, protein synthesis, regulation of enzyme activity, and signal transduction. When linked to vitamins or vitamin derivatives, nucleotides form a portion of many coenzymes. As the principal donors and acceptors of phosphoryl groups in metabolism, nucleoside tri- and diphosphates such as ATP and ADP are the principal players in the energy transductions that accompany metabolic interconversions and oxidative phosphorylation. Linked to sugars or lipids, nucleosides constitute key biosynthetic intermediates. The sugar derivatives UDP-glucose and UDP-galactose participate in sugar interconversions and in the biosynthesis of starch and glycogen. Similarly, nucleoside-lipid derivatives such as CDP-acylglycerol are intermediates in lipid biosynthesis. Roles that nucleotides perform in metabolic regulation include ATP-dependent phosphorylation of key metabolic enzymes, allosteric regulation of enzymes by ATP, ADP, AMP, and CTP, and control by ADP of the rate of oxidative phosphorylation. The cyclic nucleotides cAMP and cGMP serve as the second messengers in hormonally regulated events, and GTP and GDP play key roles in the cascade of events that characterize signal transduction pathways. Medical applications include the use of synthetic purine and pyrimidine analogs that contain halogens, thiols, or additional nitrogen atoms in the chemotherapy of cancer and AIDS, and as suppressors of the immune response during organ transplantation.
CHEMISTRY OF PURINES, PYRIMIDINES, NUCLEOSIDES & NUCLEOTIDES
Purines & Pyrimidines Are Heterocyclic Compounds
Purines and pyrimidines are nitrogen-containing heterocycles, cyclic structures that contain, in addition to carbon, other (hetero) atoms such as nitrogen. Note that the smaller pyrimidine molecule has the longer name and the larger purine molecule the shorter name, and that their six-atom rings are numbered in opposite directions (Figure 32–1). Purines or pyrimidines with an -NH2 group are weak bases (pKa values 3-4), although the proton present at low pH is associated, not as one might expect with the exocyclic amino group, but with a ring nitrogen, typically N1 of adenine, N7 of guanine, and N3 of cytosine. The planar character of purines and pyrimidines facilitates their close association, or “stacking,” that stabilizes double-stranded DNA (see Chapter 34). The oxo and amino groups of purines and pyrimidines exhibit keto-enol and amine-imine tautomerism (Figure 32–2), although physiologic conditions strongly favor the amino and oxo forms.
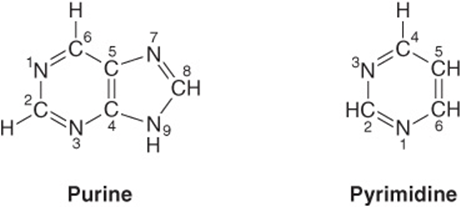
FIGURE 32–1 Purine and pyrimidine. The atoms are numbered according to the international system.

FIGURE 32–2 Tautomerism of the oxo and amino functional groups of purines and pyrimidines.
Nucleosides Are N-Glycosides
Nucleo sides are derivatives of purines and pyrimidines that have a sugar linked to a ring nitrogen of a purine or pyrimidine. Numerals with a prime (eg, 2′ or 3′) distinguish atoms of the sugar from those of the heterocycle. The sugar in ribonucleosides is D-ribose, and in deoxyribonucleosides is 2-deoxy-D-ribose. Both sugars are linked to the heterocycle by a β-N-glycosidic bond, almost always to the N-1 of a pyrimidine or to N-9 of a purine (Figure 32–3).
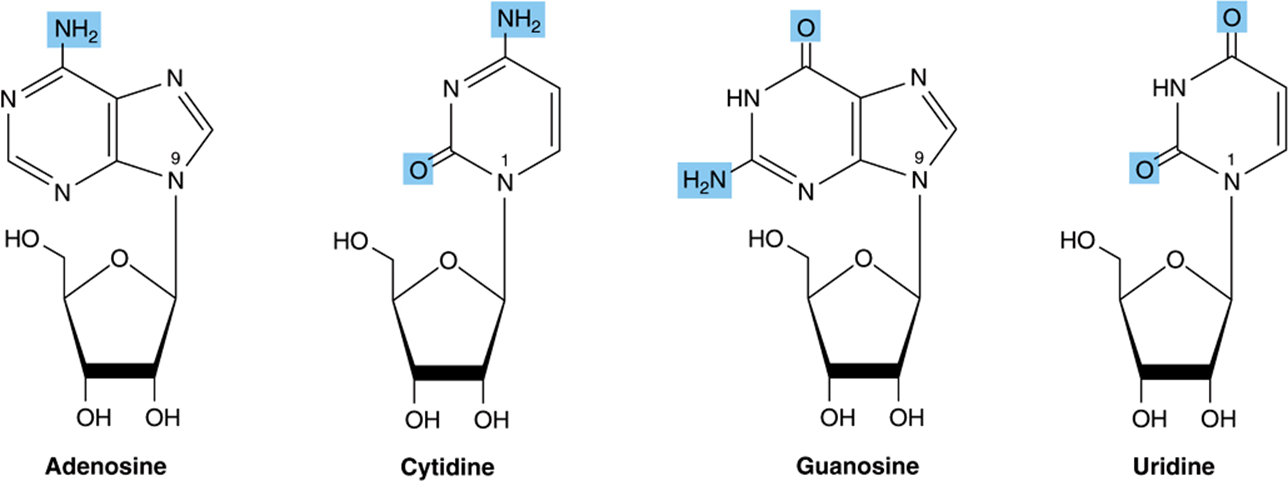
FIGURE 32–3 Ribonucleosides, drawn as the syn conformers.
Nucleotides Are Phosphorylated Nucleosides
Mononucleotides are nucleosides with a phosphoryl group esterified to a hydroxyl group of the sugar. The 3′- and 5′-nucleotides are nucleosides with a phosphoryl group on the 3′- or 5′-hydroxyl group of the sugar, respectively. Since most nucleotides are 5′-, the prefix “5’-” usually is omitted when naming them. UMP and dAMP thus represent nucleotides with a phosphoryl group on C-5 of the pentose. Additional phosphoryl groups, ligated by acid anhydride bonds to the phosphoryl group of a mononucleotide, form nucleoside diphosphates and triphosphates (Figure 32–4).
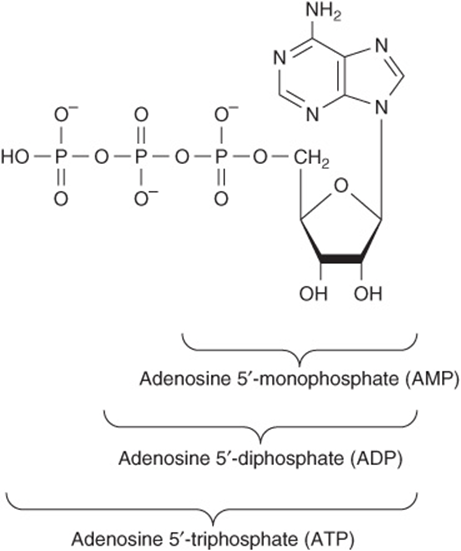
FIGURE 32–4 ATP, its diphosphate, and its monophosphate.
Heterocylic N-Glycosides Exist as Syn and Anti Conformers
Steric hindrance by the heterocycle dictates that there is no freedom of rotation about the β-N-glycosidic bond of nucleosides or nucleotides. Both therefore exist as noninterconvertible syn or anti conformers (Figure 32–5). Unlike tautomers, syn and anti conformers can only be interconverted by cleavage and reformation of the glycosidic bond. Both syn and anti conformers occur in nature, but the anti conformers predominate.
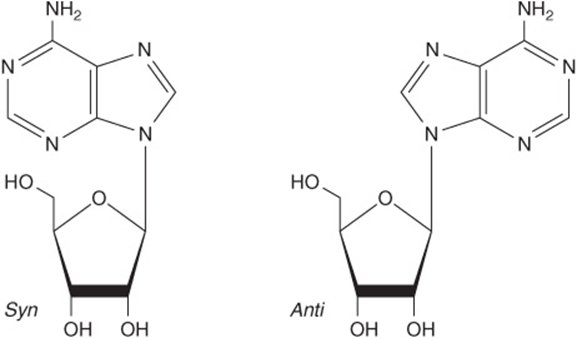
FIGURE 32–5 The syn and anti conformers of adenosine differ with respect to orientation about the N-glycosidic bond.
Table 32-1 lists the major purines and pyrimidines and their nucleoside and nucleotide derivatives. Single-letter abbreviations are used to identify adenine (A), guanine (G), cytosine (C), thymine (T), and uracil (U), whether free or present in nucleosides or nucleotides. The prefix “d” (deoxy) indicates that the sugar is 2′-deoxy-D-ribose (for example, in dATP) (Figure 32–6).
TABLE 32–1 Purine Bases, Ribonucleosides, and Ribonucleotides
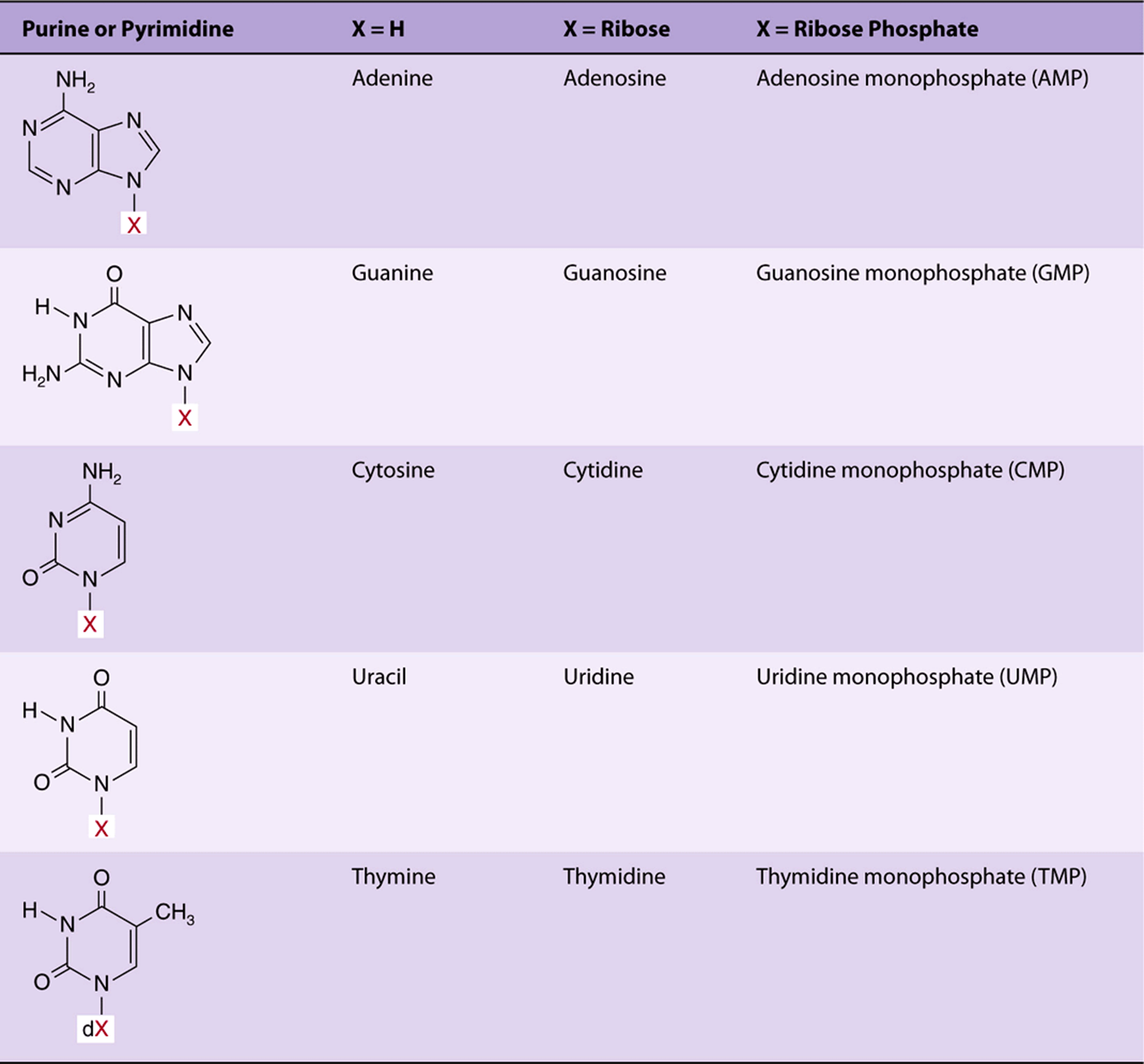
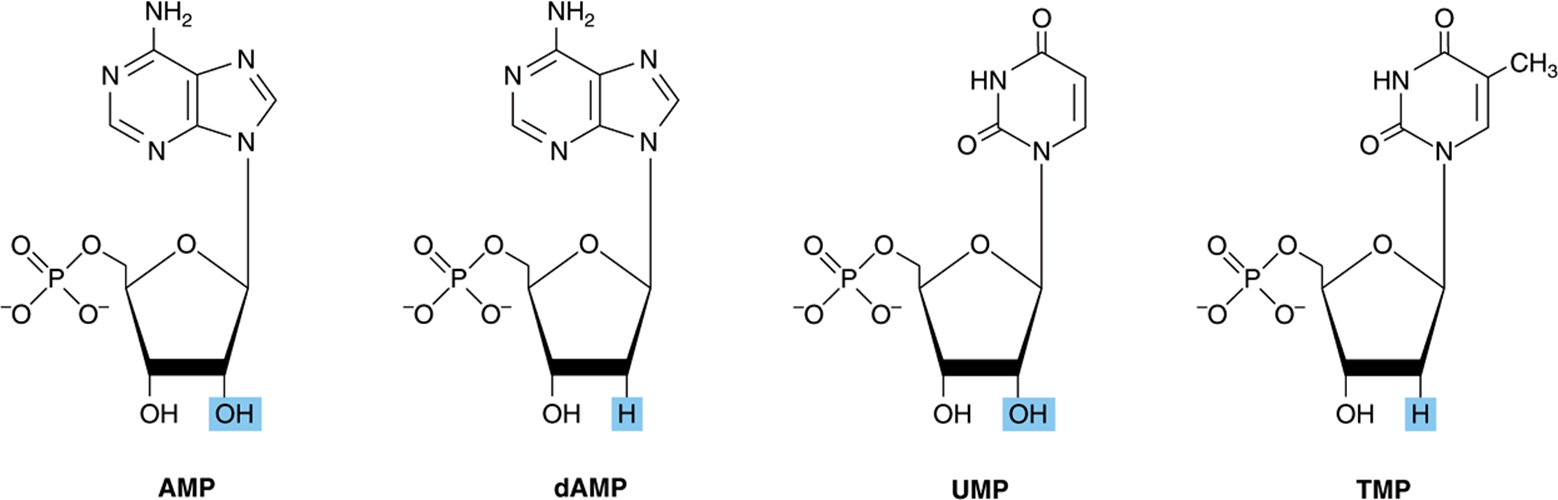
FIGURE 32–6 Structures of AMP, dAMP, UMP, and TMP.
Modification of Polynucleotides Can Generate Additional Structures
Small quantities of additional purines and pyrimidines occur in DNA and RNAs. Examples include 5-methylcytosine of bacterial and human DNA, 5-hydroxymethylcytosine of bacterial and viral nucleic acids, and mono- and the di-N-methylated adenine and guanine of mammalian messenger RNAs (Figure 32–7) that function in oligonucleotide recognition and in regulating the half-lives of RNAs. Free nucleotides include hypoxanthine, xanthine, and uric acid (Figure 32–8) that are intermediates in the catabolism of adenine and guanine (see Chapter 33). Methylated heterocycles of plants include the xanthine derivatives caffeine of coffee, theophylline of tea, and theobromine of cocoa (Figure 32–9).
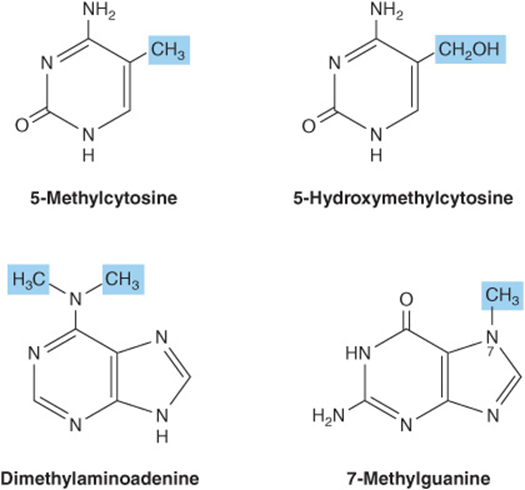
FIGURE 32–7 Four uncommon naturally occurring pyrimidines and purines.
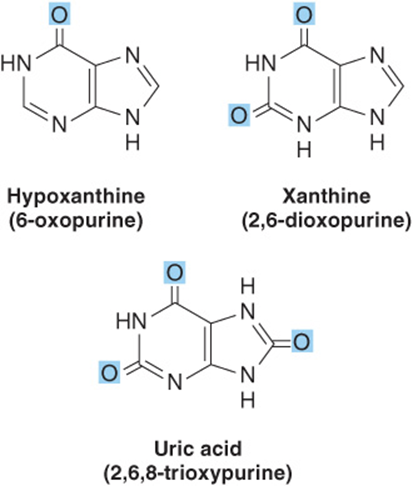
FIGURE 32–8 Structures of hypoxanthine, xanthine, and uric acid, drawn as the oxo tautomers.
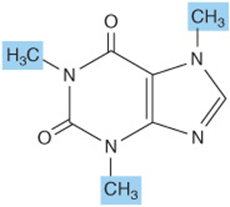
FIGURE 32–9 Caffeine, a trimethylxanthine. The dimethylxanthines theobromine and theophylline are similar but lack the methyl group at N-1 and at N-7, respectively.
Nucleotides Are Polyfunctional Acids
The primary and secondary phosphoryl groups of nucleosides have pKa values of about 1.0 and 6.2, respectively. Nucleotides therefore bear significant negative charge at physiologic pH. The pKa values of the secondary phosphoryl groups are such that they can serve both as proton donors and as proton acceptors at pH values approximately two or more units above or below neutrality.
Nucleotides Absorb Ultraviolet Light
The conjugated double bonds of purine and pyrimidine derivatives absorb ultraviolet light. While spectra are pH-dependent, at pH 7.0 all the common nucleotides absorb light at a wavelength close to 260 nm. The concentration of nucleotides and nucleic acids thus often is expressed in terms of “absorbance at 260 nm.” The mutagenic effect of ultraviolet light is due to its absorption by nucleotides in DNA that results in chemical modifications (see Chapter 35).
Nucleotides Serve Diverse Physiologic Functions
In addition to their roles as precursors of nucleic acids, ATP, GTP, UTP, CTP, and their derivatives each serve unique physiologic functions discussed in other chapters. Selected examples include the role of ATP as the principal biologic transducer of free energy, and the second messenger cAMP (Figure 32–10). The mean intracellular concentration of ATP, the most abundant free nucleotide in mammalian cells, is about 1 mmol/L. Since little cAMP is required, the intracellular cAMP concentration (about 1 nmol/L) is six orders of magnitude below that of ATP. Other examples include adenosine 3′-phosphate-5′-phosphosulfate (Figure 32–11), the sulfate donor for sulfated proteoglycans (see Chapter 48) and for sulfate conjugates of drugs; and the methyl group donor S-adenosylmethionine (Figure 32–12). GTP serves as an allosteric regulator and as an energy source for protein synthesis, and cGMP (Figure 32–10) serves as a second messenger in response to nitric oxide (NO) during relaxation of smooth muscle (see Chapter 49).
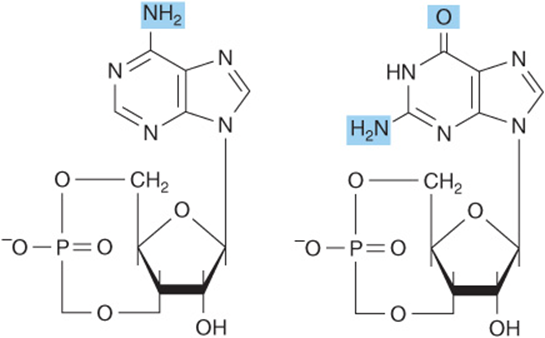
FIGURE 32–10 cAMP, 3’,5’-cyclic AMP, and cGMP, 3′, 5′-cyclic GMP.
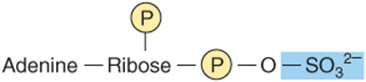
FIGURE 32–11 Adenosine 3′-phosphate-5′-phosphosulfate.
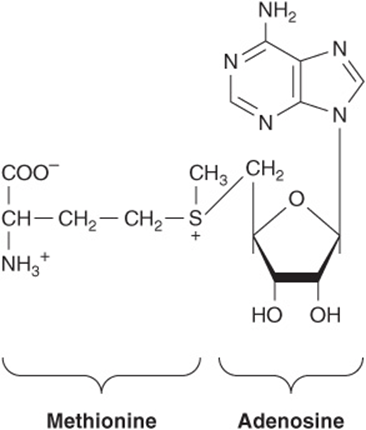
FIGURE 32–12 S-Adenosylmethionine.
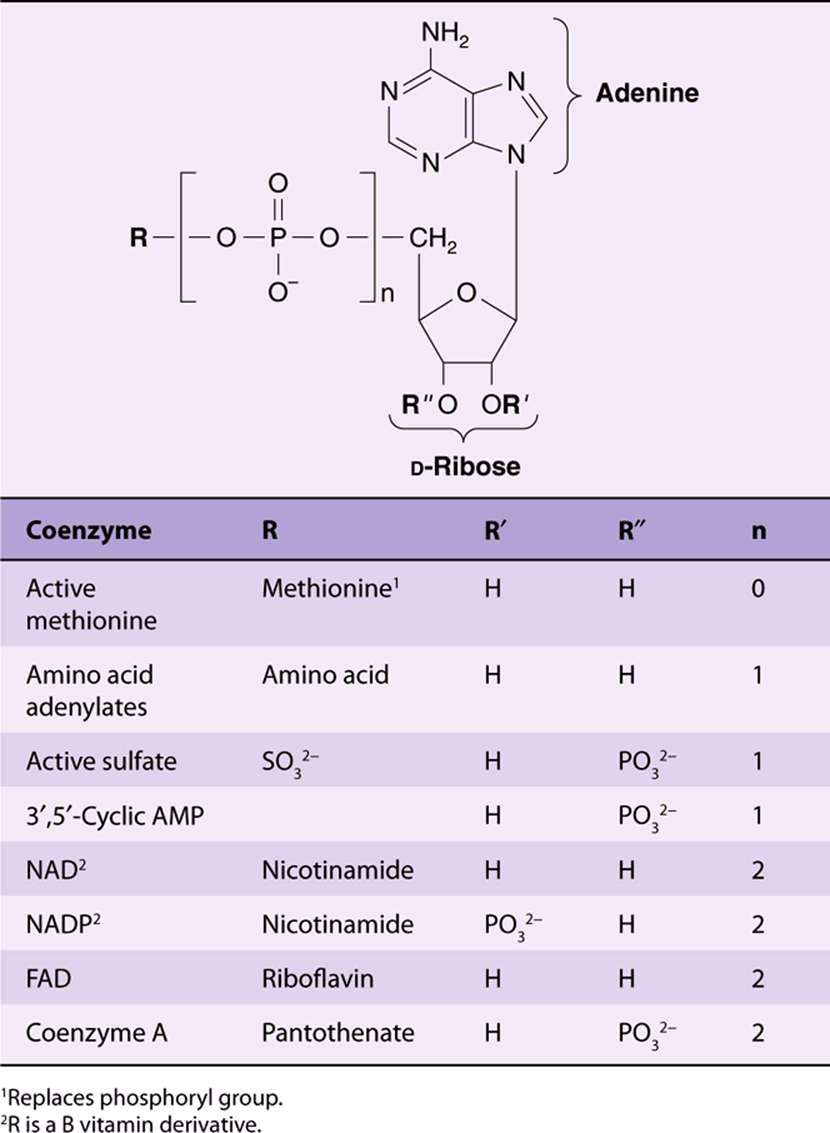
UDP-sugar derivatives participate in sugar epimerizations and in biosynthesis of glycogen (see Chapter 19), glucosyl disaccharides, and the oligosaccharides of glycoproteins and proteoglycans (see Chapters 47 and 48). UDP-glucuronic acid forms the urinary glucuronide conjugates of bilirubin (see Chapter 31) and of many drugs, including aspirin. CTP participates in biosynthesis of phosphoglycerides, sphingomyelin, and other substituted sphingosines (see Chapter 24). Finally, many coenzymes incorporate nucleotides as well as structures similar to purine and pyrimidine nucleotides (see Table 32-2).
TABLE 32-2 Many Coenzymes and Related Compounds Are Derivatives of Adenosine Monophosphate
Nucleoside Triphosphates Have High-Group Transfer Potential
Nucleotide triphosphates have two acid anhydride bonds and one ester bond. Unlike esters, acid anhydrides have a high-group transfer potential. ![]() for the hydrolysis of each of the two terminal (β and γ) phosphoryl groups of a nucleoside triphosphate is about -7 kcal/mol (-30 kJ/mol). This high group transfer potential not only permits purine and pyrimidine nucleoside triphosphates to function as group transfer reagents, most commonly of the γ-phosphoryl group, but also on occasion transfer of a nucleotide monophosphate moiety with an accompanying release of PPi. Cleavage of an acid anhydride bond typically is coupled with a highly endergonic process such as covalent bond synthesis, for example, the polymerization of nucleoside triphosphates to form a nucleic acid (see Chapter 34).
for the hydrolysis of each of the two terminal (β and γ) phosphoryl groups of a nucleoside triphosphate is about -7 kcal/mol (-30 kJ/mol). This high group transfer potential not only permits purine and pyrimidine nucleoside triphosphates to function as group transfer reagents, most commonly of the γ-phosphoryl group, but also on occasion transfer of a nucleotide monophosphate moiety with an accompanying release of PPi. Cleavage of an acid anhydride bond typically is coupled with a highly endergonic process such as covalent bond synthesis, for example, the polymerization of nucleoside triphosphates to form a nucleic acid (see Chapter 34).
SYNTHETIC NUCLEOTIDE ANALOGS ARE USED IN CHEMOTHERAPY
Synthetic analogs of purines, pyrimidines, nucleosides, and nucleotides modified in the heterocyclic ring or in the sugar moiety have numerous applications in clinical medicine. Their toxic effects reflect either inhibition of enzymes essential for nucleic acid synthesis or their incorporation into nucleic acids with resulting disruption of base-pairing. Oncologists employ 5- fluoro- or 5-iodouracil, 3-deoxyuridine, 6-thioguanine and 6- mercaptopurine, 5- or 6-azauridine, 5- or 6-azacytidine, and 8-azaguanine (Figure 32–13), which are incorporated into DNA prior to cell division. The purine analog allopurinol, used in treatment of hyperuricemia and gout, inhibits purine biosynthesis and xanthine oxidase activity. Cytarabine is used in chemotherapy of cancer, and azathioprine, which is catabolized to 6-mercaptopurine, is employed during organ transplantation to suppress immunologic rejection (Figure 32–14).
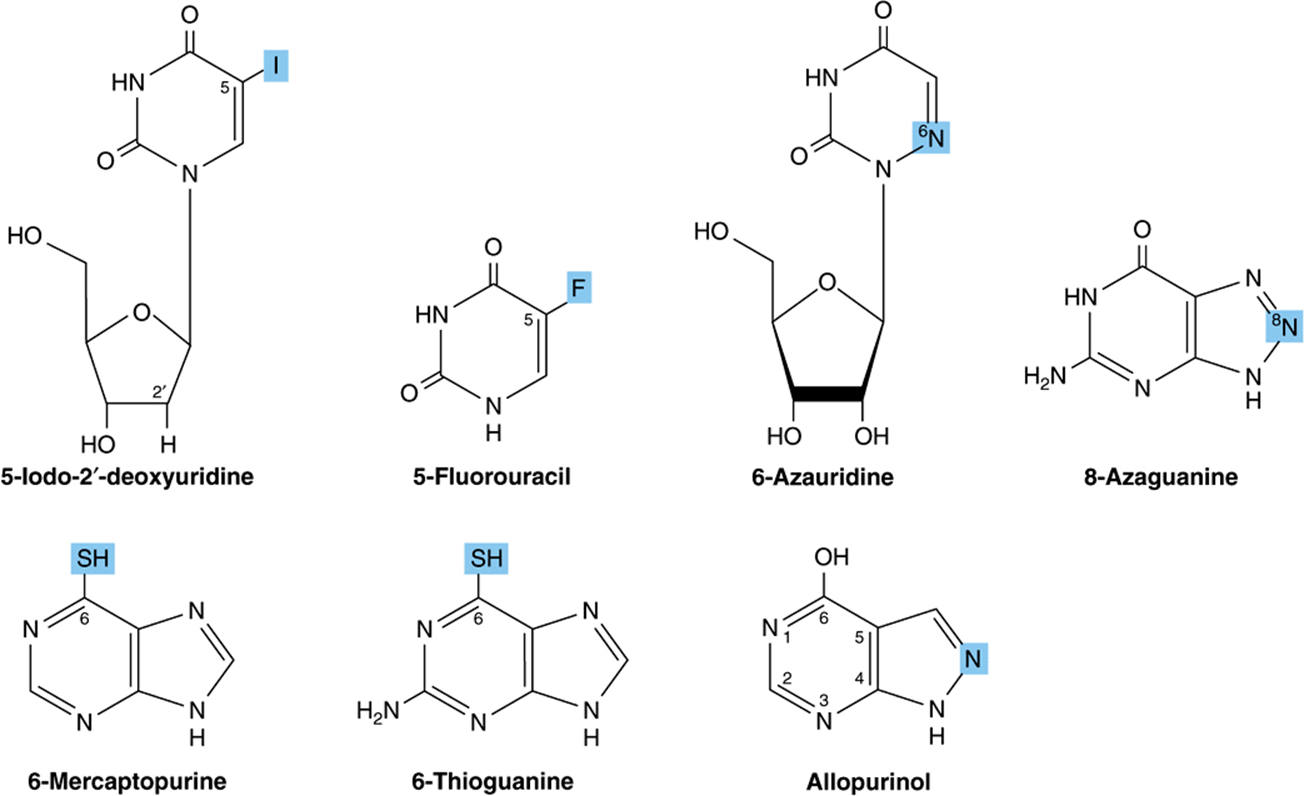
FIGURE 32–13 Selected synthetic pyrimidine and purine analogs.
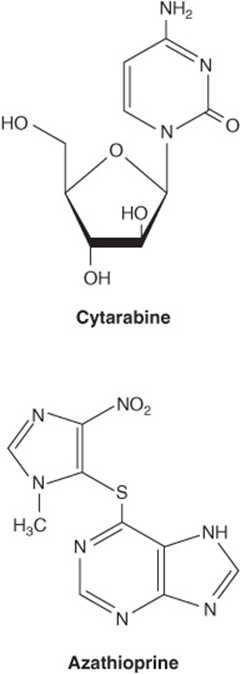
FIGURE 32–14 Arabinosylcytosine (cytarabine) and azathioprine.
Nonhydrolyzable Nucleoside Triphosphate Analogs Serve as Research Tools
Synthetic, nonhydrolyzable analogs of nucleoside triphosphates (Figure 32–15) allow investigators to distinguish the effects of nucleotides due to phosphoryl transfer from effects mediated by occupancy of allosteric nucleotide-binding sites on regulated enzymes.
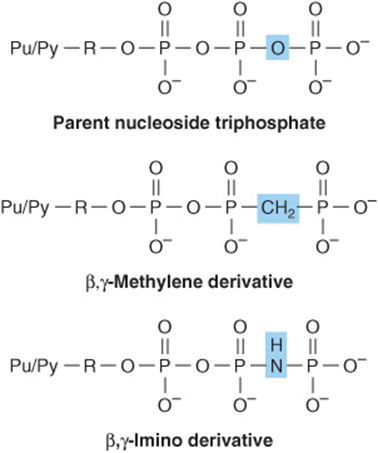
FIGURE 32–15 Synthetic derivatives of nucleoside triphosphates incapable of undergoing hydrolytic release of the terminal phosphoryl group. (Pu/Py, a purine or pyrimidine base; R, ribose or deoxyribose.) Shown are the parent (hydrolyzable) nucleoside triphosphate (top) and the unhydrolyzable β-methylene (center) and γ-imino derivatives (bottom).
DNA & RNA ARE POLYNUCLEOTIDES
The 5′-phosphoryl group of a mononucleotide can esterify a second hydroxyl group, forming a phosphodiester. Most commonly, this second hydroxyl group is the 3′-OH of the pentose of a second nucleotide. This forms a dinucleotide in which the pentose moieties are linked by a 3′,5′ -phosphodiester bond to form the “backbone” of RNA and DNA. The formation of a dinucleotide may be represented as the elimination of water between two mononucleotides. Biologic formation of dinucleotides does not, however, occur in this way because the reverse reaction, hydrolysis of the phosphodiester bond, is strongly favored on thermodynamic grounds. However, despite an extremely favorable ΔG, in the absence of catalysis by phosphodiesterases hydrolysis of the phosphodiester bonds of DNA occurs only over long periods of time. DNA therefore persists for considerable periods, and has been detected even in fossils. RNAs are far less stable than DNA since the 2′-hydroxyl group of RNA (absent from DNA) functions as a nucleophile during hydrolysis of the 3′,5′ -phosphodiester bond.
Posttranslational modification of preformed polynucleotides can generate additional structures such as pseudouridine, a nucleoside in which D-ribose is linked to C-5 of uracil by a carbon-to-carbon bond rather than by the usual β-N-glycosidic bond. The nucleotide pseudouridylic acid (ψ) arises by rearrangement of a UMP of a preformed tRNA. Similarly, methylation by S-adenosylmethionine of a UMP of preformed tRNA forms TMP (thymidine monophosphate), which contains ribose rather than deoxyribose.
Polynucleotides Are Directional Macromolecules
Phosphodiester bonds link the 3′ - and 5′ -carbons of adjacent monomers. Each end of a nucleotide polymer thus is distinct. We therefore refer to the “5 -end” or the “3 -end” of a polynucleotide, the 5′-end being the one with a free or phosphorylated 5′-hydroxyl.
The base sequence or primary structure of a polynucleotide can be represented as shown below. The phosphodiester bond is represented by P or p, bases by a single letter, and pentoses by a vertical line.
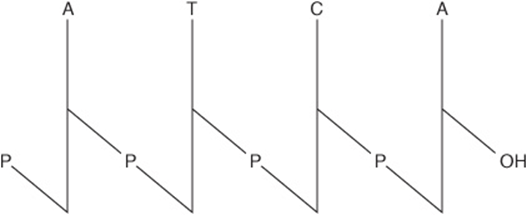
Where all the phosphodiester bonds are ![]() , a more compact notation is possible:
, a more compact notation is possible:
![]()
This representation indicates that the 5′-hydroxyl—but not the 3′-hydroxyl—is phosphorylated. The most compact representation, for example GGATC, shows only the base sequence with the 5′-end on the left and the 3′-end on the right. The phosphoryl groups are assumed to be present, but not shown.
SUMMARY
![]() Under physiologic conditions, the amino and oxo tautomers of purines, pyrimidines, and their derivatives predominate.
Under physiologic conditions, the amino and oxo tautomers of purines, pyrimidines, and their derivatives predominate.
![]() Nucleic acids contain, in addition to A, G, C, T, and U, traces of 5-methylcytosine, 5-hydroxymethylcytosine, pseudouridine (ψ), and N-methylated heterocycles.
Nucleic acids contain, in addition to A, G, C, T, and U, traces of 5-methylcytosine, 5-hydroxymethylcytosine, pseudouridine (ψ), and N-methylated heterocycles.
![]() Most nucleosides contain D-ribose or 2-deoxy-D-ribose linked to N-1 of a pyrimidine or to N-9 of a purine by a β-glycosidic bond whose syn conformers predominate.
Most nucleosides contain D-ribose or 2-deoxy-D-ribose linked to N-1 of a pyrimidine or to N-9 of a purine by a β-glycosidic bond whose syn conformers predominate.
![]() A primed numeral locates the position of the phosphate on the sugars of mononucleotides (eg, 3′-GMP, 5′-dCMP). Additional phosphoryl groups linked to the first by acid anhydride bonds form nucleoside diphosphates and triphosphates.
A primed numeral locates the position of the phosphate on the sugars of mononucleotides (eg, 3′-GMP, 5′-dCMP). Additional phosphoryl groups linked to the first by acid anhydride bonds form nucleoside diphosphates and triphosphates.
![]() Nucleoside triphosphates have high group transfer potential and participate in covalent bond syntheses. The cyclic phosphodiesters cAMP and cGMP function as intracellular second messengers.
Nucleoside triphosphates have high group transfer potential and participate in covalent bond syntheses. The cyclic phosphodiesters cAMP and cGMP function as intracellular second messengers.
![]() Mononucleotides linked by
Mononucleotides linked by ![]() -phosphodiester bonds form polynucleotides, directional macromolecules with distinct 3′- and 5′-ends. For pTpGpT or TGCATCA, the 5′-end is at the left, and all phosphodiester bonds are
-phosphodiester bonds form polynucleotides, directional macromolecules with distinct 3′- and 5′-ends. For pTpGpT or TGCATCA, the 5′-end is at the left, and all phosphodiester bonds are ![]() .
.
![]() Synthetic analogs of purine and pyrimidine bases and their derivatives serve as anticancer drugs either by inhibiting an enzyme of nucleotide biosynthesis or by being incorporated into DNA or RNA.
Synthetic analogs of purine and pyrimidine bases and their derivatives serve as anticancer drugs either by inhibiting an enzyme of nucleotide biosynthesis or by being incorporated into DNA or RNA.
REFERENCES
Adams RLP, Knowler JT, Leader DP: The Biochemistry of the Nucleic Acids, 11th ed. Chapman & Hall, 1992.
Blackburn GM, Gait MJ: Nucleic Acids in Chemistry & Biology. IRL Press, 1990.
Pacher P, Nivorozhkin A, Szabo C: Therapeutic effects of xanthine oxidase inhibitors: renaissance half a century after the discovery of allopurinol. Pharmacol Rev 2006;58:87.