Harper’s Illustrated Biochemistry, 29th Edition (2012)
SECTION IV. Structure, Function, & Replication of Informational Macromolecules
Chapter 33. Metabolism of Purine & Pyrimidine Nucleotides
Victor W. Rodwell, PhD
OBJECTIVES
After studying this chapter, you should be able to:
![]() Compare and contrast the roles of dietary nucleic acids and of de novo biosynthesis in the production of purines and pyrimidines destined for polynucleotide biosynthesis.
Compare and contrast the roles of dietary nucleic acids and of de novo biosynthesis in the production of purines and pyrimidines destined for polynucleotide biosynthesis.
![]() Explain why antifolate drugs and analogs of the amino acid glutamine inhibit purine biosynthesis.
Explain why antifolate drugs and analogs of the amino acid glutamine inhibit purine biosynthesis.
![]() Outline the sequence of reactions that convert IMP, first to AMP and GMP, and subsequently to their corresponding nucleoside triphosphates.
Outline the sequence of reactions that convert IMP, first to AMP and GMP, and subsequently to their corresponding nucleoside triphosphates.
![]() Describe the formation from ribonucleotides of deoxyribonucleotides (dNTPs).
Describe the formation from ribonucleotides of deoxyribonucleotides (dNTPs).
![]() Indicate the regulatory role of PRPP in hepatic purine biosynthesis and the specific reaction of hepatic purine biosynthesis that is feedback inhibited by AMP and by GMP.
Indicate the regulatory role of PRPP in hepatic purine biosynthesis and the specific reaction of hepatic purine biosynthesis that is feedback inhibited by AMP and by GMP.
![]() State the relevance of coordinated control of purine and pyrimidine nucleotide biosynthesis.
State the relevance of coordinated control of purine and pyrimidine nucleotide biosynthesis.
![]() Identify reactions that are inhibited by anticancer drugs.
Identify reactions that are inhibited by anticancer drugs.
![]() Write the structure of the end product of purine catabolism. Comment on its solubility and indicate its role in gout, Lesch-Nyhan syndrome, and von Gierke disease.
Write the structure of the end product of purine catabolism. Comment on its solubility and indicate its role in gout, Lesch-Nyhan syndrome, and von Gierke disease.
![]() Identify reactions whose impairment leads to modified pathologic signs and symptoms.
Identify reactions whose impairment leads to modified pathologic signs and symptoms.
![]() Indicate why there are few clinically significant disorders of pyrimidine catabolism.
Indicate why there are few clinically significant disorders of pyrimidine catabolism.
BIOMEDICAL IMPORTANCE
Even when humans consume a diet rich in nucleoproteins, dietary purines and pyrimidines are not incorporated directly into tissue nucleic acids. Humans synthesize the nucleic acids, ATP, NAD+, coenzyme A, etc, from amphibolic intermediates. However, injected purine or pyrimidine analogs, including potential anticancer drugs, may be incorporated into DNA. The biosyntheses of purine and pyrimidine ribonucleotide triphosphates (NTPs) and dNTPs are precisely regulated events. Coordinated feedback mechanisms ensure their production in appropriate quantities and at times that match varying physiologic demand (eg, cell division). Human diseases that involve abnormalities in purine metabolism include gout, Lesch-Nyhan syndrome, adenosine deaminase deficiency, and purine nucleoside phosphorylase deficiency. Diseases of pyrimidine biosynthesis are rarer, but include orotic acidurias. Unlike the low solubility of uric acid formed by catabolism of purines, the end-products of pyrimidine catabolism (carbon dioxide, ammonia, β-alanine, and γ-aminoisobutyrate) are highly water soluble. One genetic disorder of pyrimidine catabolism is β-hydroxybutyric aciduria, due to total or partial deficiency of the enzyme dihydropyrimidine dehydrogenase. This disorder of pyrimidine catabolism, also known as combined uraciluria-thyminuria, is also a disorder of β-amino acid biosynthesis, since the formation of β-alanine and of β-aminoisobutyrate is impaired. A nongenetic form can be triggered by administration of 5-fluorouracil to patients with low levels of dihydropyrimidine dehydrogenase.
PURINES & PYRIMIDINES ARE DIETARILY NONESSENTIAL
Normal human tissues can synthesize purines and pyrimidines from amphibolic intermediates in quantities and at times appropriate to meet variable physiologic demand. Ingested nucleic acids and nucleotides therefore are dietarily nones-sential. Following their degradation in the intestinal tract, the resulting mononucleotides may be absorbed or converted to purine and pyrimidine bases. The purine bases are then oxidized to uric acid, which may be absorbed and excreted in the urine. While little or no dietary purine or pyrimidine is incorporated into tissue nucleic acids, injected compounds are incorporated. The incorporation of injected [3H]thymidine into newly synthesized DNA thus can be used to measure the rate of DNA synthesis.
BIOSYNTHESIS OF PURINE NUCLEOTIDES
With the exception of parasitic protozoa, all forms of life synthesize purine and pyrimidine nucleotides. Synthesis from amphibolic intermediates proceeds at controlled rates appropriate for all cellular functions. To achieve homeostasis, intracellular mechanisms sense and regulate the pool sizes of NTPs, which rise during growth or tissue regeneration when cells are rapidly dividing.
Purine and pyrimidine nucleotides are synthesized in vivo at rates consistent with physiologic need. Early investigations of nucleotide biosynthesis first employed birds, and later Escherichia coli. Isotopic precursors of uric acid fed to pigeons established the source of each atom of a purine (Figure 33–1) and initiated study of the intermediates of purine biosynthesis. Avian tissues also served as a source of cloned genes that encode enzymes of purine biosynthesis and the regulatory proteins that control the rate of purine biosynthesis.
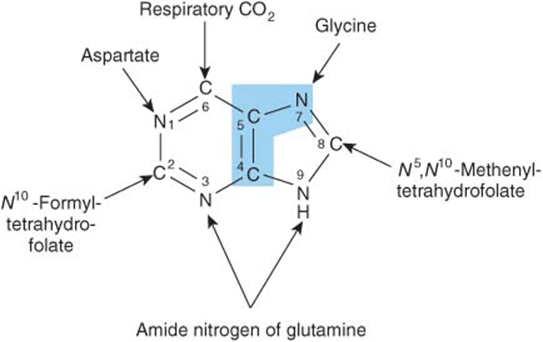
FIGURE 33–1 Sources of the nitrogen and carbon atoms of the purine ring. Atoms 4, 5, and 7 (blue highlight) derive from glycine.
The three processes that contribute to purine nucleotide biosynthesis are, in order of decreasing importance.
1. Synthesis from amphibolic intermediates (synthesis de novo).
2. Phosphoribosylation of purines.
3. Phosphorylation of purine nucleosides.
INOSINE MONOPHOSPHATE (IMP) IS SYNTHESIZED FROM AMPHIBOLIC INTERMEDIATES
Figure 33–2 illustrates the intermediates and the 11 enzyme-catalyzed reactions that convert α-D-ribose 5-phosphate to inosine monophosphate (IMP). The first intermediate formed in the de novo pathway for purine biosynthesis is 5-phosphoribosyl 5-pyrophosphate (PRPP; structure II, Figure 33–2). PRPP is also an intermediate in the biosynthesis of pyrimidine nucleotides, NAD+, and NADP+. Separate branches then lead from IMP to AMP and GMP (Figure 33–3). Subsequent phosphoryl transfer from ATP converts AMP and GMP to ADP and GDP. Conversion of GDP to GTP involves a second phosphoryl transfer from ATP, whereas conversion of ADP to ATP is achieved primarily by oxidative phosphorylation (see Chapter 13).
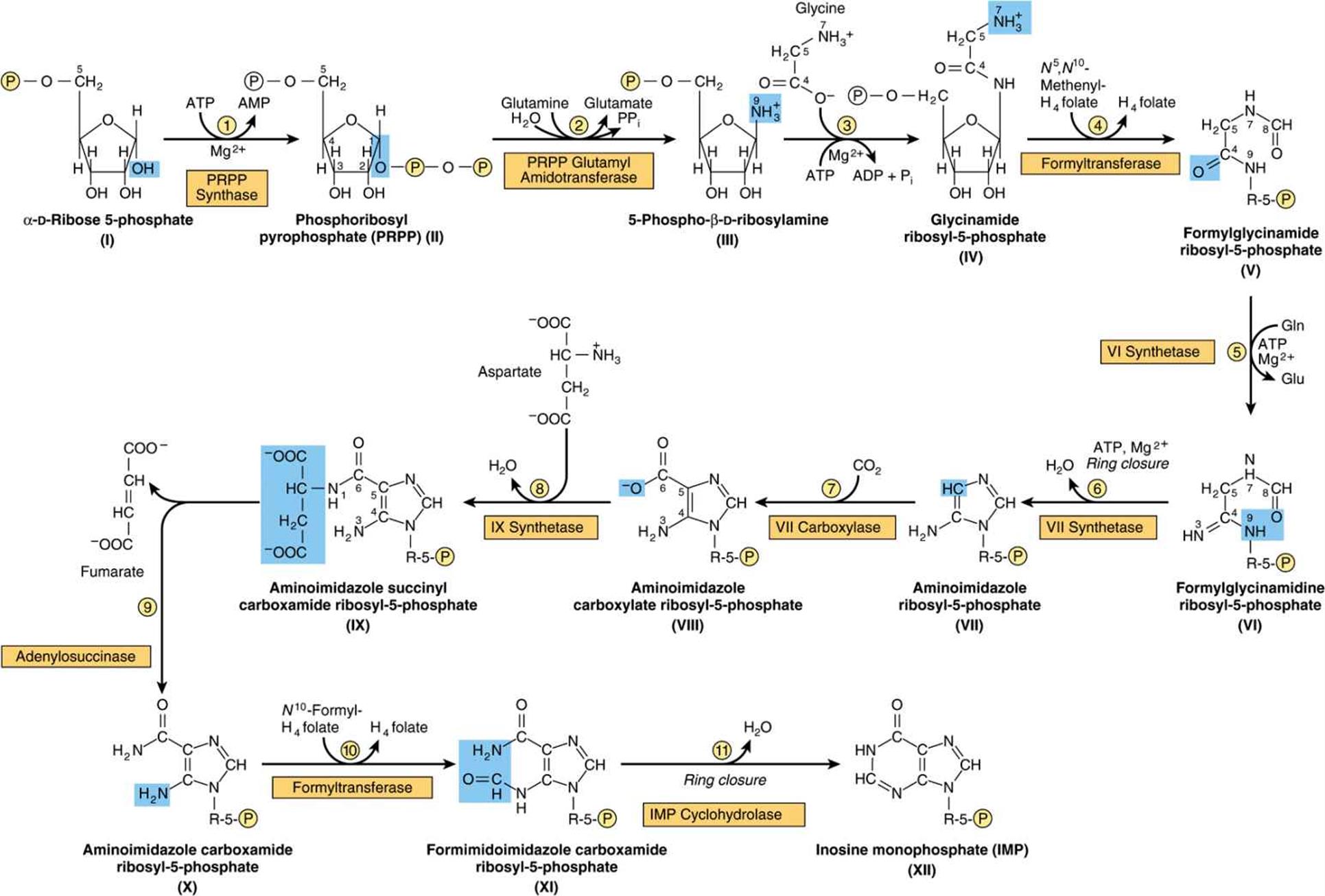
FIGURE 33–2 Purine biosynthesis from ribose 5-phosphate and ATP. See the text for explanations. ![]()
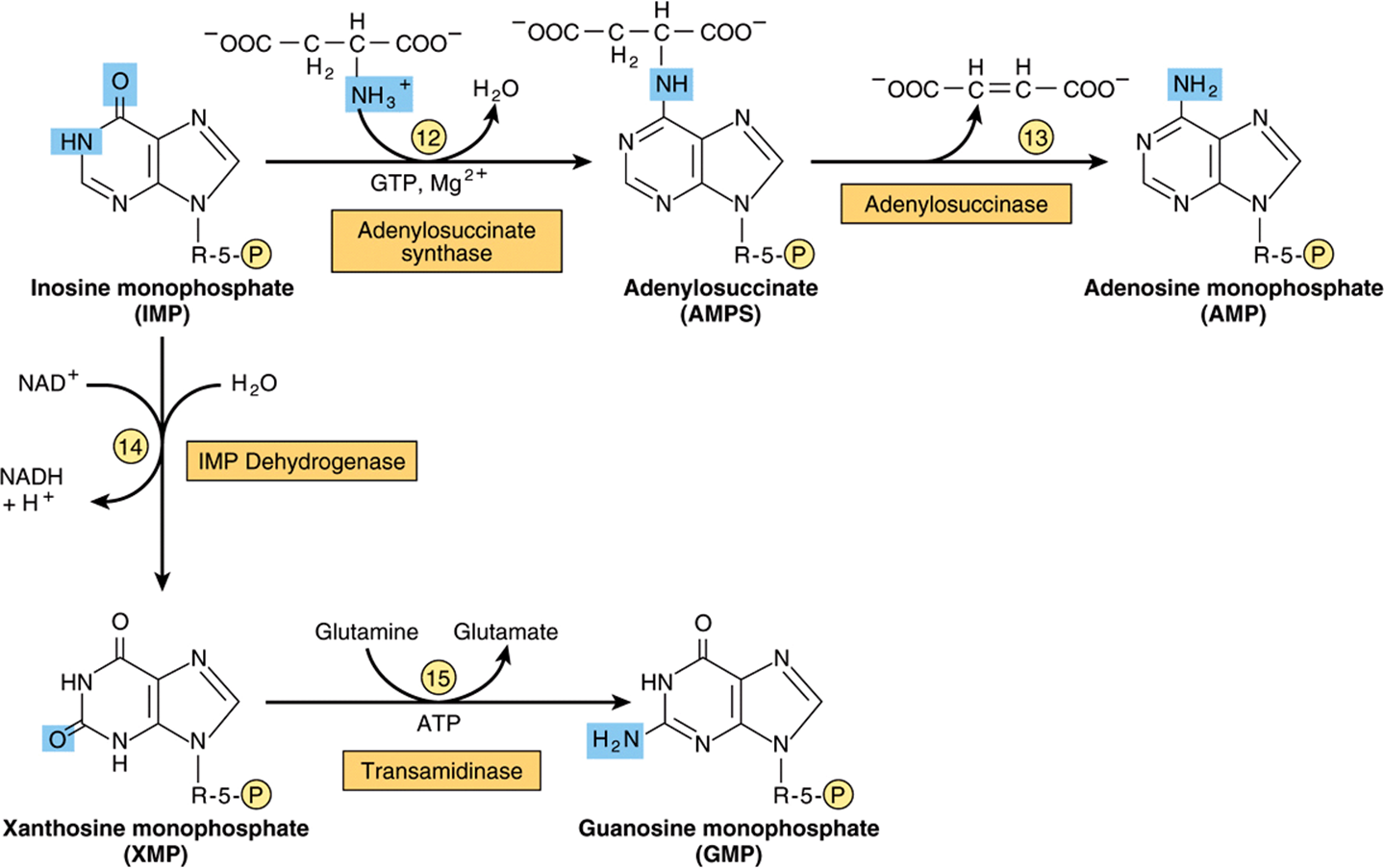
FIGURE 33–3 Conversion of IMP to AMP and GMP.
Multifunctional Catalysts Participate in Purine Nucleotide Biosynthesis
In prokaryotes, each reaction of Figure 33–2 is catalyzed by a different polypeptide. By contrast, the enzymes of eukaryotes are polypeptides that possess multiple catalytic activities whose adjacent catalytic sites facilitate channeling of intermediates between sites. Three distinct multifunctional enzymes catalyze reactions ![]() ,
, ![]() , and
, and ![]() ; reactions
; reactions ![]() and
and ![]() ; and reactions
; and reactions ![]() and
and ![]() of Figure 33–2.
of Figure 33–2.
Antifolate Drugs and Glutamine Analogs Block Purine Nucleotide Biosynthesis
The carbons added in reactions ![]() and
and ![]() of Figure 33–2 are contributed by derivatives of tetrahydrofolate. Purine deficiency states, while rare in humans, generally reflect a deficiency of folic acid. Compounds that inhibit formation of tetrahydrofolates and therefore block purine synthesis have been used in cancer chemotherapy. Inhibitory compounds and the reactions they inhibit include azaserine (reaction
of Figure 33–2 are contributed by derivatives of tetrahydrofolate. Purine deficiency states, while rare in humans, generally reflect a deficiency of folic acid. Compounds that inhibit formation of tetrahydrofolates and therefore block purine synthesis have been used in cancer chemotherapy. Inhibitory compounds and the reactions they inhibit include azaserine (reaction ![]() , Figure 33–2), diazanorleucine(reaction
, Figure 33–2), diazanorleucine(reaction ![]() , Figure 33–2), 6-mercaptopurine (reactions
, Figure 33–2), 6-mercaptopurine (reactions ![]() and
and ![]() , Figure 33–3), and mycophenolic acid (reaction
, Figure 33–3), and mycophenolic acid (reaction ![]() , Figure 33–3).
, Figure 33–3).
“SALVAGE REACTIONS” CONVERT PURINES & THEIR NUCLEOSIDES TO MONONUCLEOTIDES
Conversion of purines, their ribonucleosides, and their deoxyribonucleosides to mononucleotides involves “salvage reactions” that require far less energy than de novo synthesis. The more important mechanism involves phosphoribosylation by PRPP (structure II, Figure 33–2) of a free purine (Pu) to form a purine 5′-mononucleotide (Pu-RP).
![]()
Phosphoryl transfer from ATP, catalyzed by adenosine-and hypoxanthine-phosphoribosyl transferases, converts adenine, hypoxanthine, and guanine to their mononucleotides (Figure 33–4).
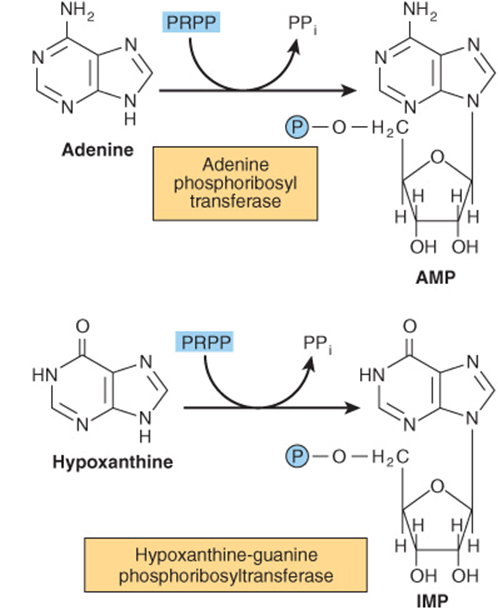
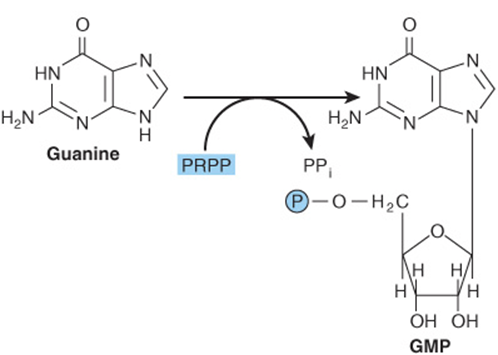
FIGURE 33–4 Phosphoribosylation of adenine, hypoxanthine, and guanine to form AMP, IMP, and GMP, respectively.
A second salvage mechanism involves phosphoryl transfer from ATP to a purine ribonucleo side (Pu-R):
![]()
Phosphorylation of the purine nucleotides, catalyzed by adenosine kinase, converts adenosine and deoxyadenosine to AMP and dAMP. Similarly, deoxycytidine kinase phosphorylates deoxycytidine and 2′-deoxyguanosine, forming dCMP and dGMP.
Liver, the major site of purine nucleotide biosynthesis, provides purines and purine nucleosides for salvage and for utilization by tissues incapable of their biosynthesis. Human brain tissue has a low level of PRPP glutamyl amidotransferase (reaction ![]() , Figure 33–2) and hence depends in part on exogenous purines. Erythrocytes and polymorphonuclear leukocytes cannot synthesize 5-phosphoribosylamine (structure III, Figure 33–2) and therefore utilize exogenous purines to form nucleotides.
, Figure 33–2) and hence depends in part on exogenous purines. Erythrocytes and polymorphonuclear leukocytes cannot synthesize 5-phosphoribosylamine (structure III, Figure 33–2) and therefore utilize exogenous purines to form nucleotides.
HEPATIC PURINE BIOSYNTHESIS IS STRINGENTLY REGULATED
AMP & GMP Feedback Regulate PRPP Glutamyl Amidotransferase
Biosynthesis of IMP is energetically expensive. In addition to ATP, glycine, glutamine, aspartate, and reduced tetrahydrofolate derivatives all are consumed. It thus is of survival advantage to closely regulate purine biosynthesis in response to varying physiological need. The overall determinant of the rate of de novo purine nucleotide biosynthesis is the concentration of PRPP. This depends on the rate of PRPP synthesis, utilization, degradation, and regulation. The rate of PRPP synthesis depends on the availability of ribose 5-phosphate and on the activity of PRPP synthase, (reaction ![]() Figure 33–5), an enzyme whose activity is feedback inhibited by AMP, ADP, GMP, and GDP. Elevated levels of these nucleoside phosphates thus signal a physiologically appropriate overall decrease in their biosynthesis.
Figure 33–5), an enzyme whose activity is feedback inhibited by AMP, ADP, GMP, and GDP. Elevated levels of these nucleoside phosphates thus signal a physiologically appropriate overall decrease in their biosynthesis.
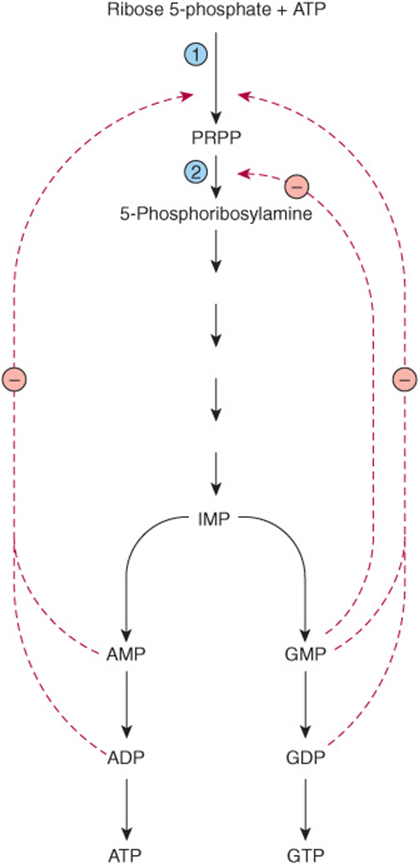
FIGURE 33–5 Control of the rate of de novo purine nucleotide biosynthesis. Reactions ![]() and
and ![]() are catalyzed by PRPP synthase and by PRPP glutamyl amidotransferase, respectively. Solid lines represent chemical flow. Broken red lines represent feedback inhibition by intermediates of the pathway.
are catalyzed by PRPP synthase and by PRPP glutamyl amidotransferase, respectively. Solid lines represent chemical flow. Broken red lines represent feedback inhibition by intermediates of the pathway.
AMP & GMP Feedback Regulate Their Formation from IMP
In addition to regulation at the level of PRPP biosynthesis, additional mechanisms regulate conversion of IMP to ATP and GTP. These are summarized in Figure 33–6. AMP feedback inhibits adenylosuccinate synthase (reaction ![]() , Figure 33–3), and GMP inhibits IMP dehydrogenase (reaction
, Figure 33–3), and GMP inhibits IMP dehydrogenase (reaction ![]() , Figure 33–3). Furthermore, conversion of IMP to adenylosuccinate en route to AMP (reaction
, Figure 33–3). Furthermore, conversion of IMP to adenylosuccinate en route to AMP (reaction ![]() , Figure 33–3) requires GTP, and conversion of xanthinylate (XMP) to GMP requires ATP. This cross-regulation between the pathways of IMP metabolism thus serves to balance the biosynthesis of purine nucleoside triphosphates by decreasing the synthesis of one purine nucleotide when there is a deficiency of the other nucleotide. AMP and GMP also inhibit hypoxanthine-guanine phosphoribosyltransferase, which converts hypoxanthine and guanine to IMP and GMP (Figure 33–4), and GMP feedback inhibits PRPP glutamyl amidotransferase (reaction
, Figure 33–3) requires GTP, and conversion of xanthinylate (XMP) to GMP requires ATP. This cross-regulation between the pathways of IMP metabolism thus serves to balance the biosynthesis of purine nucleoside triphosphates by decreasing the synthesis of one purine nucleotide when there is a deficiency of the other nucleotide. AMP and GMP also inhibit hypoxanthine-guanine phosphoribosyltransferase, which converts hypoxanthine and guanine to IMP and GMP (Figure 33–4), and GMP feedback inhibits PRPP glutamyl amidotransferase (reaction ![]() , Figure 33–2).
, Figure 33–2).
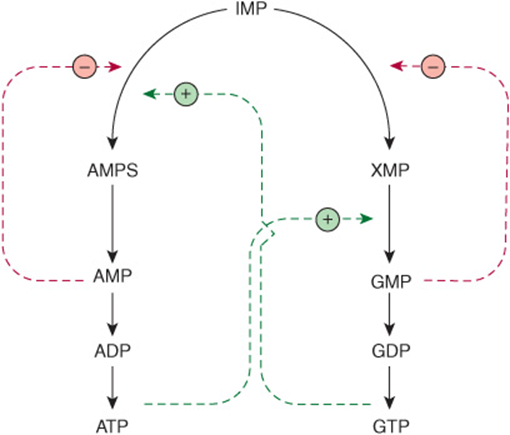
FIGURE 33–6 Regulation of the conversion of IMP to adenosine nucleotides and guanosine nucleotides. Solid lines represent chemical flow. Broken green lines represent positive feedback loops ![]() , and broken red lines represent negative feedback loops
, and broken red lines represent negative feedback loops ![]() . Abbreviations include AMPS (adenylosuccinate) and XMP (xanthosine monophosphate), whose structures are given in Figure 33–3.
. Abbreviations include AMPS (adenylosuccinate) and XMP (xanthosine monophosphate), whose structures are given in Figure 33–3.
REDUCTION OF RIBONUCLEOSIDE DIPHOSPHATES FORMS DEOXYRIBONUCLEOSIDE DIPHOSPHATES
Reduction of the 2′-hydroxyl of purine and pyrimidine ribonucleotides, catalyzed by the ribonucleotide reductase complex (Figure 33–7), provides the deoxyribonucleoside diphosphates (dNDPs) needed for both the synthesis and repair of DNA (see Chapter 35). The enzyme complex is functional only when cells are actively synthesizing DNA. Reduction requires thioredoxin, thioredoxin reductase, and NADPH. The immediate reductant, reduced thioredoxin, is produced by NADPH:thioredoxin reductase (Figure 33–7). The reduction of ribonucleoside diphosphates (NDPs) to dNDPs is subject to complex regulatory controls that achieve balanced production of dNTPs for synthesis of DNA (Figure 33–8).
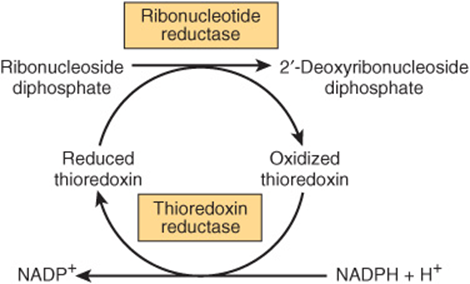
FIGURE 33–7 Reduction of ribonucleoside diphosphates to 2′-deoxyribonucleoside diphosphates.
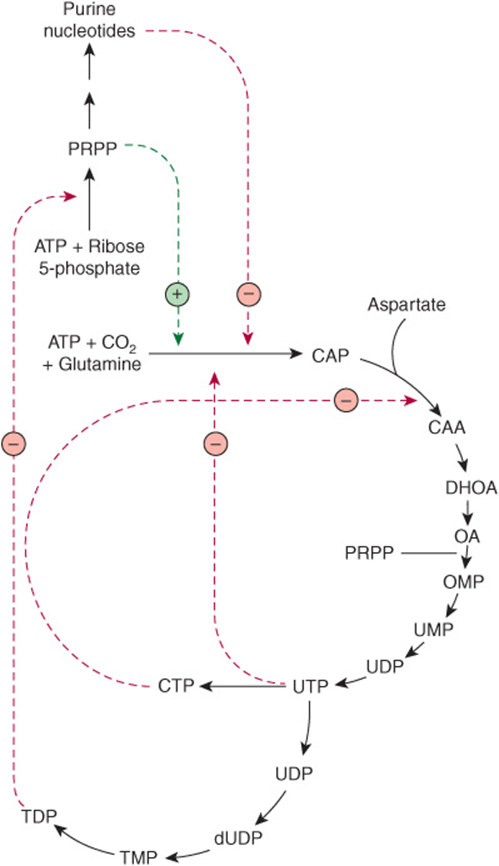
FIGURE 33–8 Regulatory aspects of the biosynthesis of purine and pyrimidine ribonucleotides and reduction to their respective 2′-deoxyribonucleotides. The broken green line represents a positive feedback loop. Broken red lines represent negative feedback loops. Abbreviations for the intermediates in the biosynthesis of pyrimidine nucleotides whose structures are given in Figure 33–9 are: (CAA, carbamoyl aspartate; DHOA, dihydroorotate; OA, orotic acid; OMP, orotidine monophosphate; and PRPP phosphoribosyl pyrophosphate).
BIOSYNTHESIS OF PYRIMIDINE NUCLEOTIDES
Figure 33–9 illustrates the intermediates and enzymes of pyrimidine nucleotide biosynthesis. The catalyst for the initial reaction is cytosolic carbamoyl phosphate synthase II, a different enzyme from the mitochondrial carbamoyl phosphate synthase I of urea synthesis (s ee Figure 28–13). Compartmentation thus provides an independent pool of carbamoyl phosphate for each process. PRPP, an early participant in purine nucleotide synthesis (Figure 33–2), is a much later participant in pyrimidine biosynthesis. Inspection of the reaction components in Figure 33–9 will reveal that, like the biosynthesis of pyrimidines, the biosynthesis of the purine nucleosides is energetically costly.
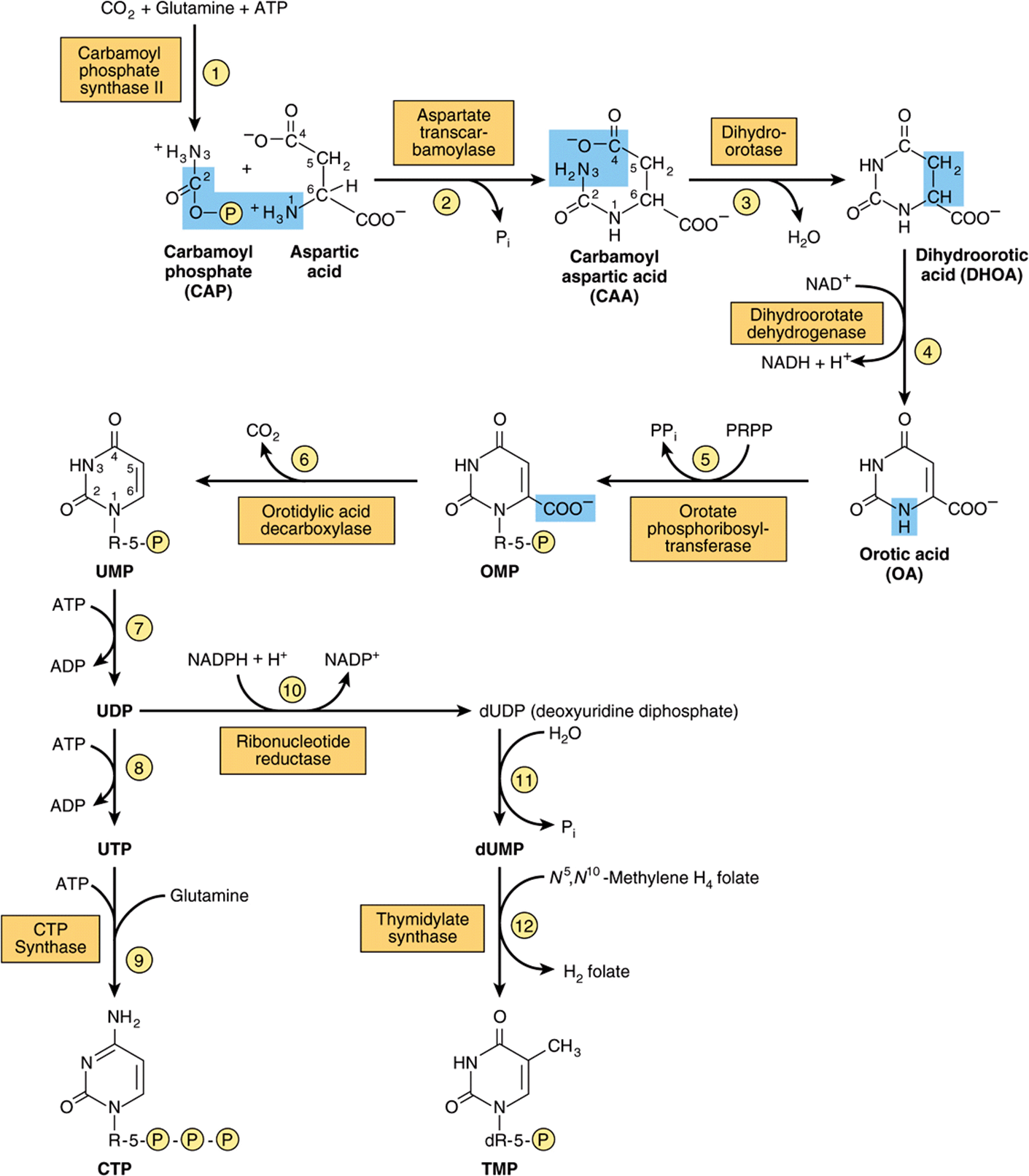
FIGURE 33–9 The biosynthetic pathway for pyrimidine nucleotides.
Multifunctional Proteins Catalyze the Early Reactions of Pyrimidine Biosynthesis
Five of the first six enzyme activities of pyrimidine biosynthesis reside on multifunctional polypeptides. One polypeptide catalyzes the first three reactions of Figure 33–9. A second bifunctional enzyme catalyzes reactions ![]() and
and ![]() of Figure 33–9. The close proximity of multiple active sites on a multifunctional polypeptide facilitates efficient channeling of the intermediates of pyrimidine biosynthesis.
of Figure 33–9. The close proximity of multiple active sites on a multifunctional polypeptide facilitates efficient channeling of the intermediates of pyrimidine biosynthesis.
THE DEOXYRIBONUCLEOSIDES OF URACIL & CYTOSINE ARE SALVAGED
Adenine, guanine and hypoxanthine released during the turnover of nucleic acids, especially messenger RNAs, are reconverted to nucleoside triphosphates via so-called salvage pathways. While mammalian cells reutilize few freepyrimidines, “salvage reactions” convert the pyrimidine ribonucleosides uridine and cytidine and the pyrimidine deoxyribonucleosides thymidine and deoxycytidine to their respective nucleotides.
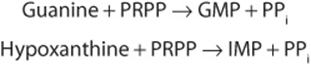
Phosphoryltransferases (kinases) catalyze transfer of the γ-phosphoryl group of ATP to the diphosphates of 2′-deoxycytidine, 2′-deoxyguanosine, and 2′-deoxyadenosine, converting them to the corresponding nucleoside triphosphates.
![]()
Methotrexate Blocks Reduction of Dihydrofolate
The reaction catalyzed by thymidylate synthase (reaction ![]() of Figure 33–9) is the only reaction of pyrimidine nucleotide biosynthesis that requires a tetrahydrofolate derivative. During this reaction the methylene group of N5, N10-methylene-tetra-hydrofolate is reduced to the methyl group that is transferred to the 5-position of the pyrimidine ring, and tetrahydrofolate is oxidized to dihydrofolate. For further pyrimidine synthesis to occur, dihydrofolate must be reduced back to tetrahydrofolate. This reduction, catalyzed by dihydrofolate reductase, is inhibited by methotrexate. Dividing cells, which must generate TMP and dihydrofolate, thus are especially sensitive to inhibitors of dihydrofolate reductase such as the anticancer drug methotrexate.
of Figure 33–9) is the only reaction of pyrimidine nucleotide biosynthesis that requires a tetrahydrofolate derivative. During this reaction the methylene group of N5, N10-methylene-tetra-hydrofolate is reduced to the methyl group that is transferred to the 5-position of the pyrimidine ring, and tetrahydrofolate is oxidized to dihydrofolate. For further pyrimidine synthesis to occur, dihydrofolate must be reduced back to tetrahydrofolate. This reduction, catalyzed by dihydrofolate reductase, is inhibited by methotrexate. Dividing cells, which must generate TMP and dihydrofolate, thus are especially sensitive to inhibitors of dihydrofolate reductase such as the anticancer drug methotrexate.
Certain Pyrimidine Analogs Are Substrates for Enzymes of Pyrimidine Nucleotide Biosynthesis
Allopurinol and the anticancer drug 5-fluorouracil (see Figure 32–13) are alternate substrates for orotate phosphoribosyltransferase (reaction ![]() , Figure 33–9). Both drugs are phosphoribosylated, and allopurinol is converted to a nucleotide in which the ribosyl phosphate is attached to N-1 of the pyrimidine ring.
, Figure 33–9). Both drugs are phosphoribosylated, and allopurinol is converted to a nucleotide in which the ribosyl phosphate is attached to N-1 of the pyrimidine ring.
REGULATION OF PYRIMIDINE NUCLEOTIDE BIOSYNTHESIS
Gene Expression & Enzyme Activity Both Are Regulated
The activities of the first and second enzymes of pyrimidine nucleotide biosynthesis are controlled by allosteric regulation. Carbamoyl phosphate synthase II (reaction ![]() , Figure 33–9) is inhibited by UTP and purine nucleotides but activated by PRPP. Aspartate transcarbamoylase (reaction
, Figure 33–9) is inhibited by UTP and purine nucleotides but activated by PRPP. Aspartate transcarbamoylase (reaction ![]() , Figure 33–9) is inhibited by CTP but activated by ATP (Figure 33–10). In addition, the first three and the last two enzymes of the pathway are regulated by coordinate repression and derepression.
, Figure 33–9) is inhibited by CTP but activated by ATP (Figure 33–10). In addition, the first three and the last two enzymes of the pathway are regulated by coordinate repression and derepression.
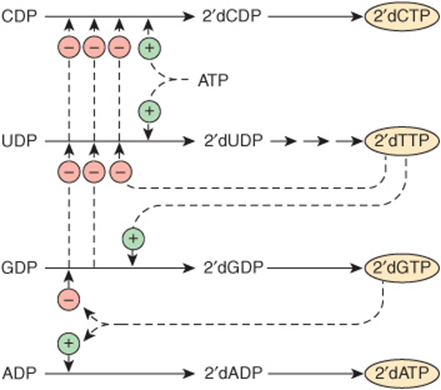
FIGURE 33–10 Control of pyrimidine nucleotide biosynthesis. Solid lines represent chemical flow. Broken green lines represent positive ![]() , and broken red lines negative
, and broken red lines negative ![]() feedback regulation.
feedback regulation.
Purine & Pyrimidine Nucleotide Biosynthesis Are Coordinately Regulated
Purine and pyrimidine biosynthesis parallel one another quantitatively, that is, mole for mole, suggesting coordinated control of their biosynthesis. Several sites of cross-regulation characterize the pathways that lead to the biosynthesis of purine and pyrimidine nucleotides. PRPP synthase (reaction ![]() , Figure 33–2), which forms a precursor essential for both processes, is feedback inhibited by both purine and pyrimidine nucleotides.
, Figure 33–2), which forms a precursor essential for both processes, is feedback inhibited by both purine and pyrimidine nucleotides.
HUMANS CATABOLIZE PURINES TO URIC ACID
Humans convert adenosine and guanosine to uric acid (Figure 33–11). Adenosine is first converted to inosine by adenosine deaminase. In mammals other than higher primates, uricase converts uric acid to the water-soluble product allantoin. However, since humans lack uricase, the end product of purine catabolism in humans is uric acid.
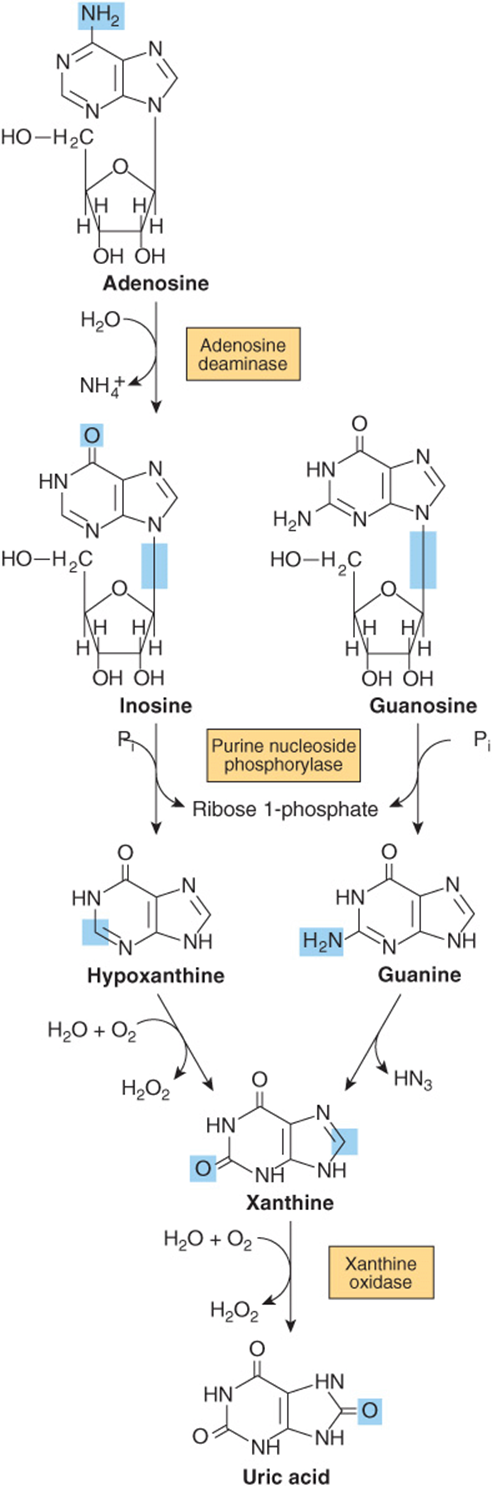
FIGURE 33–11 Formation of uric acid from purine nucleosides by way of the purine bases hypoxanthine, xanthine, and guanine. Purine deoxyribonucleosides are degraded by the same catabolic pathway and enzymes, all of which exist in the mucosa of the mammalian gastrointestinal tract.
GOUT IS A METABOLIC DISORDER OF PURINE CATABOLISM
Various genetic defects in PRPP synthase (reaction ![]() , Figure 33–2) present clinically as gout. Each defect—for example, an elevated Vmax, increased affinity for ribose 5-phosphate, or resistance to feedback inhibition—results in overproduction and overexcretion of purine catabolites. When serum urate levels exceed the solubility limit, sodium urate crystalizes in soft tissues and joints and causes an inflammatory reaction, gouty arthritis. However, most cases of gout reflect abnormalities in renal handling of uric acid.
, Figure 33–2) present clinically as gout. Each defect—for example, an elevated Vmax, increased affinity for ribose 5-phosphate, or resistance to feedback inhibition—results in overproduction and overexcretion of purine catabolites. When serum urate levels exceed the solubility limit, sodium urate crystalizes in soft tissues and joints and causes an inflammatory reaction, gouty arthritis. However, most cases of gout reflect abnormalities in renal handling of uric acid.
OTHER DISORDERS OF PURINE CATABOLISM
While purine deficiency states are rare in human subjects, there are numerous genetic disorders of purine catabolism. Hyperuricemias may be differentiated based on whether patients excrete normal or excessive quantities of total urates. Some hyperuricemias reflect specific enzyme defects. Others are secondary to diseases such as cancer or psoriasis that enhance tissue turnover.
Lesch-Nyhan Syndrome
The Lesch-Nyhan syndrome, an overproduction hyperuricemia characterized by frequent episodes of uric acid lithiasis and a bizarre syndrome of self-mutilation, reflects a defect in hypoxanthine-guanine phosphoribosyl transferase, an enzyme of purine salvage (Figure 33–4). The accompanying rise in intracellular PRPP results in purine overproduction. Mutations that decrease or abolish hypoxanthine-guanine phosphoribosyltransferase activity include deletions, frame-shift mutations, base substitutions, and aberrant mRNA splicing.
von Gierke Disease
Purine overproduction and hyperuricemia in von Gierke disease (glucose-6-phosphatase deficiency) occurs secondary to enhanced generation of the PRPP precursor ribose 5-phosphate. An associated lactic acidosis elevates the renal threshold for urate, elevating total body urates.
Hypouricemia
Hypouricemia and increased excretion of hypoxanthine and xanthine are associated with xanthine oxidase deficiency (Figure 33–11) due to a genetic defect or to severe liver damage. Patients with a severe enzyme deficiency may exhibit xanthinuria and xanthine lithiasis.
Adenosine Deaminase & Purine Nucleoside Phosphorylase Deficiency
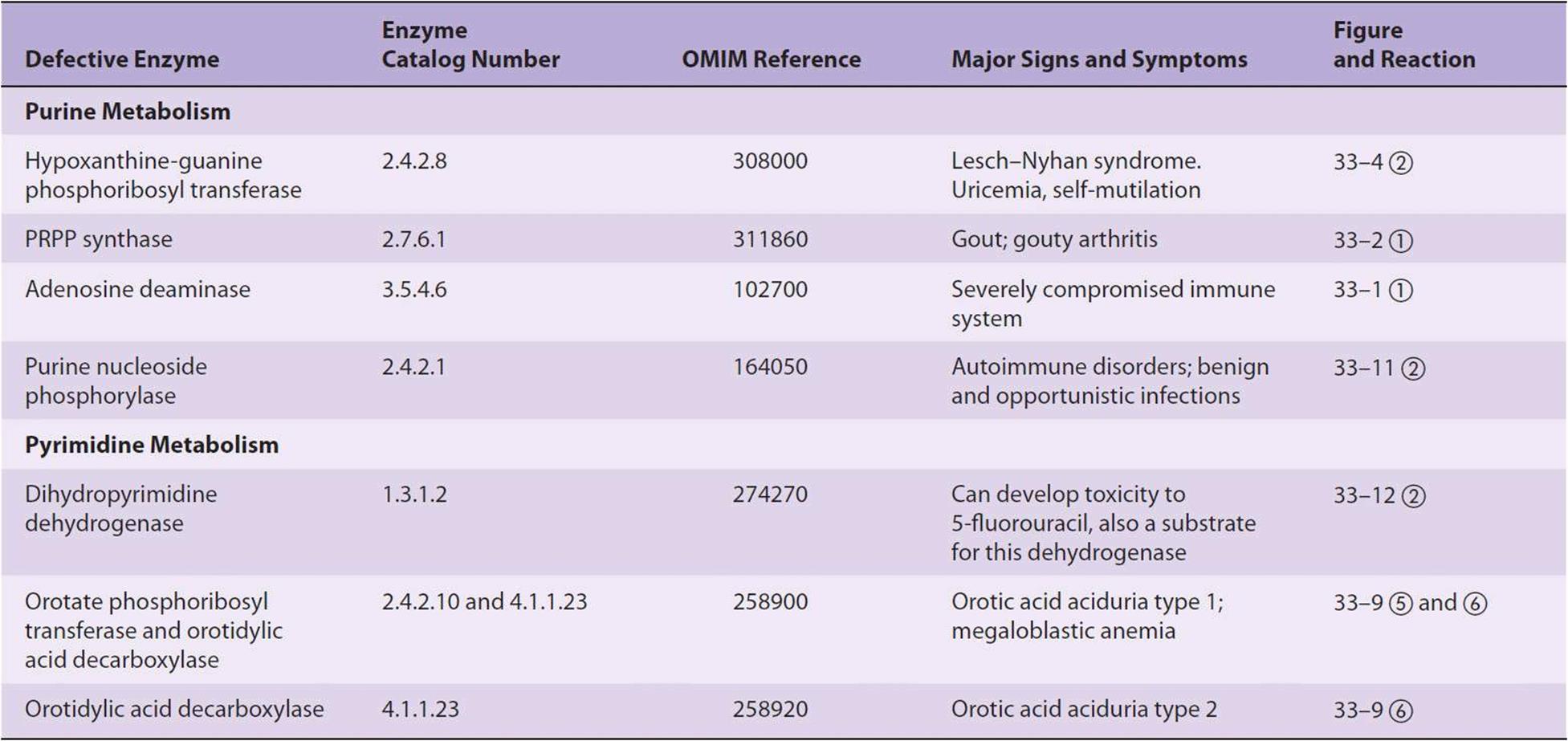
Adenosine deaminase deficiency (Figure 33–11) is associated with an immunodeficiency disease in which both thymus-derived lymphocytes (T cells) and bone marrow-derived lymphocytes (B cells) are sparse and dysfunctional. Patients suffer from severe immunodeficiency. In the absence of enzyme replacement or bone marrow transplantation, infants often succumb to fatal infections. Purine nucleoside phosphorylase deficiency is associated with a severe deficiency of T cells but apparently normal B cell function. Immune dysfunctions appear to result from accumulation of dGTP and dATP, which inhibit ribonucleotide reductase and thereby deplete cells of DNA precursors. Table 33-1 summarizes known disorders of purine metabolism.
TABLE 33–1 Metabolic Disorders of Purine and Pyrimidine Metabolism
CATABOLISM OF PYRIMIDINES PRODUCES WATER-SOLUBLE METABOLITES
Unlike the low solubility products of purine catabolism, catabolism of the pyrimidines forms highly water-soluble products—CO2, NH3, β-alanine, and β-aminoisobutyrate (Figure 33–12). Humans transaminate β-aminoisobutyrate to methylmalonate semialdehyde, which then forms succinyl-CoA (see Figure 20–2). Excretion of β-aminoisobutyrate increases in leukemia and severe x-ray radiation exposure due to increased destruction of DNA. However, many persons of Chinese or Japanese ancestry routinely excrete β-aminoisobutyrate. Disorders of β-alanine and β-aminoisobutryrate metabolism arise from defects in enzymes of pyrimidine catabolism. These include β-hydroxybutyric aciduria, a disorder due to total or partial deficiency of the enzyme dihydropyrimidine dehydrogenase (Figure 33–12). The genetic disease reflects an absence of the enzyme. A disorder of pyrimidine catabolism, known also as combined uraciluria-thyminuria, it is also a disorder of β-amino acid metabolism, since the formation of β-alanine and of β-aminoisobutyrate is impaired. When due to an inborn error, there are serious neurological complications. A nongenetic form is triggered by the administration of the anticancer drug 5-fluorouracil (see Figure 32–13) to patients with low levels of dihydropyrimidine dehydrogenase.
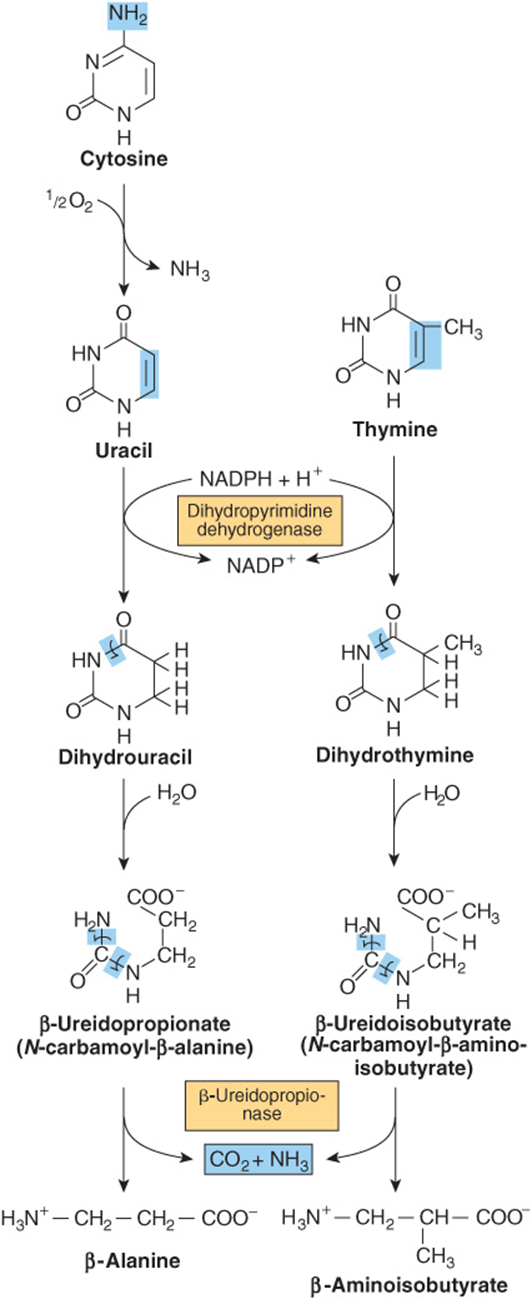
FIGURE 33–12 Catabolism of pyrimidines. Hepatic β-ureidopropionase catalyzes the formation of both β-alanine and β-aminoisobutryrate from their pyrimidine precursors.
Pseudouridine Is Excreted Unchanged
No human enzyme catalyzes hydrolysis or phosphorolysis of the pseudouridine (ψ) derived from the degradation of RNA molecules. This unusual nucleotide therefore is excreted unchanged in the urine of normal subjects. Pseudouridine was indeed first isolated from human urine (Figure 33–13).
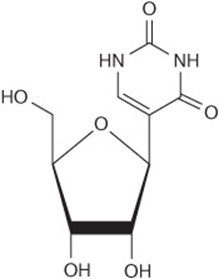
FIGURE 33–13 Pseudouridine, in which ribose is linked to C5 of uridine.
OVERPRODUCTION OF PYRIMIDINE CATABOLITES IS ONLY RARELY ASSOCIATED WITH CLINICALLY SIGNIFICANT ABNORMALITIES
Since the end products of pyrimidine catabolism are highly water soluble, pyrimidine overproduction results in few clinical signs or symptoms. Table 33-1 lists exceptions. In hyperuricemia associated with severe overproduction of PRPP, there is overproduction of pyrimidine nucleotides and increased excretion of β-alanine. Since N5,N10-methylenetetrahydrofolate is required for thymidylate synthesis, disorders of folate and vitamin B12 metabolism result in deficiencies of TMP.
Orotic Aciduria
The orotic aciduria that accompanies the Reye syndrome probably is a consequence of the inability of severely damaged mitochondria to utilize carbamoyl phosphate, which then becomes available for cytosolic overproduction of orotic acid. Type-I orotic aciduria reflects a deficiency of both orotate phosphoribosyltransferase and orotidylate decarboxylase (reactions ![]() and
and ![]() , Figure 33–9); the rarer Type-II orotic aciduria is due to a deficiency only of orotidylate decarboxylase (reaction
, Figure 33–9); the rarer Type-II orotic aciduria is due to a deficiency only of orotidylate decarboxylase (reaction ![]() , Figure 33–9).
, Figure 33–9).
Deficiency of a Urea Cycle Enzyme Results in Excretion of Pyrimidine Precursors
Increased excretion of orotic acid, uracil, and uridine accompanies a deficiency in liver mitochondrial ornithine transcarbamoylase (see reaction ![]() , Figure 28–13). Excess carbamoyl phosphate exits to the cytosol, where it stimulates pyrimidine nucleotide biosynthesis. The resulting mild orotic aciduria is increased by high-nitrogen foods.
, Figure 28–13). Excess carbamoyl phosphate exits to the cytosol, where it stimulates pyrimidine nucleotide biosynthesis. The resulting mild orotic aciduria is increased by high-nitrogen foods.
Drugs May Precipitate Orotic Aciduria
Allopurinol (see Figure 32–13), an alternative substrate for orotate phosphoribosyltransferase (reaction ![]() , Figure 33–9), competes with orotic acid. The resulting nucleotide product also inhibits orotidylate decarboxylase (reaction
, Figure 33–9), competes with orotic acid. The resulting nucleotide product also inhibits orotidylate decarboxylase (reaction ![]() , Figure 33–9), resulting in orotic aciduria and orotidinuria. 6-Azauridine, following conversion to 6-azauridylate, also competitively inhibits orotidylate decarboxylase (reaction ©, Figure 33–9), enhancing excretion of orotic acid and orotidine. Four genes that encode urate transporters have been identified. Two of the encoded proteins are localized to the apical membrane of proximal tubular cells.
, Figure 33–9), resulting in orotic aciduria and orotidinuria. 6-Azauridine, following conversion to 6-azauridylate, also competitively inhibits orotidylate decarboxylase (reaction ©, Figure 33–9), enhancing excretion of orotic acid and orotidine. Four genes that encode urate transporters have been identified. Two of the encoded proteins are localized to the apical membrane of proximal tubular cells.
SUMMARY
![]() Ingested nucleic acids are degraded to purines and pyrimidines. New purines and pyrimidines are formed from amphibolic intermediates and thus are dietarily nonessential.
Ingested nucleic acids are degraded to purines and pyrimidines. New purines and pyrimidines are formed from amphibolic intermediates and thus are dietarily nonessential.
![]() Several reactions of IMP biosynthesis require folate derivatives and glutamine. Consequently, antifolate drugs and glutamine analogs inhibit purine biosynthesis.
Several reactions of IMP biosynthesis require folate derivatives and glutamine. Consequently, antifolate drugs and glutamine analogs inhibit purine biosynthesis.
![]() Oxidation and amination of IMP forms AMP and GMP, and subsequent phosphoryl transfer from ATP forms ADP and GDP. Further phosphoryl transfer from ATP to GDP forms GTP. ADP is converted to ATP by oxidative phosphorylation. Reduction of NDPs forms dNDPs.
Oxidation and amination of IMP forms AMP and GMP, and subsequent phosphoryl transfer from ATP forms ADP and GDP. Further phosphoryl transfer from ATP to GDP forms GTP. ADP is converted to ATP by oxidative phosphorylation. Reduction of NDPs forms dNDPs.
![]() Hepatic purine nucleotide biosynthesis is stringently regulated by the pool size of PRPP and by feedback inhibition of PRPP glutamyl amidotransferase by AMP and GMP.
Hepatic purine nucleotide biosynthesis is stringently regulated by the pool size of PRPP and by feedback inhibition of PRPP glutamyl amidotransferase by AMP and GMP.
![]() Coordinated regulation of purine and pyrimidine nucleotide biosynthesis ensures their presence in proportions appropriate for nucleic acid biosynthesis and other metabolic needs.
Coordinated regulation of purine and pyrimidine nucleotide biosynthesis ensures their presence in proportions appropriate for nucleic acid biosynthesis and other metabolic needs.
![]() Humans catabolize purines to uric acid (pKa 5.8), present as the relatively insoluble acid at acidic pH or as its more soluble sodium urate salt at a pH near neutrality. Urate crystals are diagnostic of gout. Other disorders of purine catabolism include Lesch-Nyhan syndrome, von Gierke disease, and hypouricemias.
Humans catabolize purines to uric acid (pKa 5.8), present as the relatively insoluble acid at acidic pH or as its more soluble sodium urate salt at a pH near neutrality. Urate crystals are diagnostic of gout. Other disorders of purine catabolism include Lesch-Nyhan syndrome, von Gierke disease, and hypouricemias.
![]() Since pyrimidine catabolites are water-soluble, their overproduction does not result in clinical abnormalities. Excretion of pyrimidine precursors can, however, result from a deficiency of ornithine transcarbamoylase because excess carbamoyl phosphate is available for pyrimidine biosynthesis.
Since pyrimidine catabolites are water-soluble, their overproduction does not result in clinical abnormalities. Excretion of pyrimidine precursors can, however, result from a deficiency of ornithine transcarbamoylase because excess carbamoyl phosphate is available for pyrimidine biosynthesis.
REFERENCES
Chow EL, Cherry JD, Harrison R, et al: Reassessing Reye syndrome. Arch Pediatr Adolesc Med 2003;157:1241.
Christopherson RI, Lyons SD, Wilson PK: Inhibitors of de novo nucleotide biosynthesis as drugs. Acc Chem Res 2002;35:961.
Kamal MA, Christopherson RI: Accumulation of 5-phosphoribosyl-1-pyrophosphate in human CCRF-CEM leukemia cells treated with antifolates. Int J Biochem Cell Biol 2004;36:957.
Lipkowitz MS, Leal-Pinto E, Rappoport JZ, et al: Functional reconstitution, membrane targeting, genomic structure, and chromosomal localization of a human urate transporter. J Clin Invest 2001;107:1103.
Martinez J, Dugaiczyk LJ, Zielinski R, et al: Human genetic disorders, a phylogenetic perspective. J Mol Biol 2001;308:587.
Moyer RA, John DS: Acute gout precipitated by total parenteral nutrition. J Rheumatol 2003;30:849.
Nofech-Mozes Y, Blaser SI, Kobayashi J, et al: Neurologic abnormalities in patients with adenosine deaminase deficiency. Pediatr Neurol 2007;37:218.
Rafey MA, Lipkowitz MS, Leal-Pinto E, et al: Uric acid transport. Curr Opin Nephrol Hypertens 2003;12:511.
Scriver CR, Sly WS, Childs B, et al (editors): The Metabolic and Molecular Bases of Inherited Disease, 8th ed. McGraw-Hill, 2001.
Torres RJ, Puig JG: Hypoxanthine-guanine phosophoribosyltransferase (HPRT) deficiency: Lesch-Nyhan syndrome. Orphanet J Rare Dis 2007;2:48.
Wu VC, Huang JW, Hsueh PR, et al: Renal hypouricemia is an ominous sign in patients with severe acute respiratory syndrome. Am J Kidney Dis 2005;45:88.