Harper’s Illustrated Biochemistry, 29th Edition (2012)
SECTION V. Biochemistry of Extracellular & Intracellular Communication
Chapter 41. The Diversity of the Endocrine System
P. Anthony Weil, PhD
OBJECTIVES
After studying this chapter, you should be able to:
![]() Explain the basic principles of endocrine hormone action, including the determinants of hormone target cell response and the determinants of hormone concentration at target cells.
Explain the basic principles of endocrine hormone action, including the determinants of hormone target cell response and the determinants of hormone concentration at target cells.
![]() Understand the broad diversity and mechanisms of action of endocrine hormones.
Understand the broad diversity and mechanisms of action of endocrine hormones.
![]() Appreciate the complex steps involved in the production, transport, and storage of hormones.
Appreciate the complex steps involved in the production, transport, and storage of hormones.
ACTH
Adrenocorticotropic hormone
ANF
Atrial natriuretic factor
cAMP
Cyclic adenosine monophosphate
CBG
Corticosteroid-binding globulin
CG
Chorionic gonadotropin
cGMP
Cyclic guanosine monophosphate
CLIP
Corticotropin-like intermediate lobe peptide
DBH
Dopamine β-hydroxylase
DHEA
Dehydroepiandrosterone
DHT
Dihydrotestosterone
DIT
Diiodotyrosine
DOC
Deoxycorticosterone
EGF
Epidermal growth factor
FSH
Follicle-stimulating hormone
GH
Growth hormone
IGF-I
Insulin-like growth factor-I
LH
Luteotropic hormone
LPH
Lipotropin
MIT
Monoiodotyrosine
MSH
Melanocyte-stimulating hormone
OHSD
Hydroxysteroid dehydrogenase
PNMT
Phenylethanolamine-N-methyltransferase
POMC
Pro-opiomelanocortin
SHBG
Sex hormone-binding globulin
StAR
Steroidogenic acute regulatory (protein)
TBG
Thyroxine-binding globulin
TEBG
Testosterone-estrogen-binding globulin
TRH
Thyrotropin-releasing hormone
TSH
Thyrotropin-stimulating hormone
BIOMEDICAL IMPORTANCE
The survival of multicellular organisms depends on their ability to adapt to a constantly changing environment. Intercellular communication mechanisms are necessary requirements for this adaptation. The nervous system and the endocrine system provide this intercellular, organism-wide communication. The nervous system was originally viewed as providing a fixed communication system, whereas the endocrine system supplied hormones, which are mobile messages. In fact, there is a remarkable convergence of these regulatory systems. For example, neural regulation of the endocrine system is important in the production and secretion of some hormones; many neurotransmitters resemble hormones in their synthesis, transport, and mechanism of action; and many hormones are synthesized in the nervous system. The word “hormone” is derived from a Greek term that means to arouse to activity. As classically defined, a hormone is a substance that is synthesized in one organ and transported by the circulatory system to act on another tissue. However, this original description is too restrictive because hormones can act on adjacent cells (paracrine action) and on the cell in which they were synthesized (autocrine action) without entering the systemic circulation. A diverse array of hormones—each with distinctive mechanisms of action and properties of biosynthesis, storage, secretion, transport, and metabolism—has evolved to provide homeostatic responses. This biochemical diversity is the topic of this chapter.
THE TARGET CELL CONCEPT
There are about 200 types of differentiated cells in humans. Only a few produce hormones, but virtually all of the 75 trillion cells in a human are targets of one or more of the >50 known hormones. The concept of the target cell is a useful way of looking at hormone action. It was thought that hormones affected a single cell type—or only a few kinds of cells—and that a hormone elicited a unique biochemical or physiologic action. We now know that a given hormone can affect several different cell types; that more than one hormone can affect a given cell type; and that hormones can exert many different effects in one cell or in different cells. With the discovery of specific cell-surface and intracellular hormone receptors, the definition of a target has been expanded to include any cell in which the hormone (ligand) binds to its receptor, whether or not a biochemical or physiologic response has yet been determined.
Several factors determine the response of a target cell to a hormone. These can be thought of in two general ways: (1) as factors that affect the concentration of the hormone at the target cell (Table 41-1) and (2) as factors that affect the actual response of the target cell to the hormone (Table 41-2).
TABLE 41–1 Determinants of the Concentration of a Hormone at the Target Cell

TABLE 41-2 Determinants of the Target Cell Response

HORMONE RECEPTORS ARE OF CENTRAL IMPORTANCE
Receptors Discriminate Precisely
One of the major challenges faced in making the hormone-based communication system work is illustrated in Figure 41–1. Hormones are present at very low concentrations in the extracellular fluid, generally in the atto- to nanomolar range (10-15 to 10-9 mol/L). This concentration is much lower than that of the many structurally similar molecules (sterols, amino acids, peptides, and proteins) and other molecules that circulate at concentrations in the micro- to millimolar (10-6 to 10-3 mol/L) range. Target cells, therefore, must distinguish not only between different hormones present in small amounts but also between a given hormone and the 106- to 109-fold excess of other similar molecules. This high degree of discrimination is provided by cell-associated recognition molecules called receptors. Hormones initiate their biologic effects by binding to specific receptors, and since any effective control system also must provide a means of stopping a response, hormone-induced actions generally but not always terminate when the effector dissociates from the receptor (Figure 38–1; Type A response).
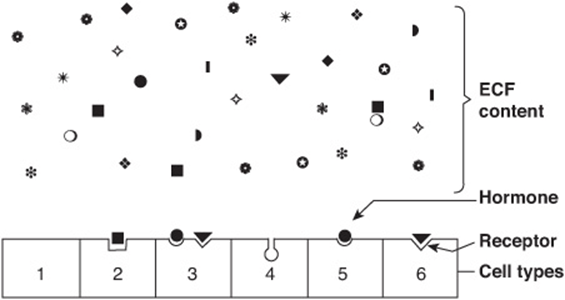
FIGURE 41–1 Specificity and selectivity of hormone receptors. Many different molecules circulate in the extracellular fluid (ECF), but only a few are recognized by hormone receptors. Receptors must select these molecules from among high concentrations of the other molecules. This simplified drawing shows that a cell may have no hormone receptors (1), have one receptor (2+5+6), have receptors for several hormones (3), or have a receptor but no hormone in the vicinity (4).
A target cell is defined by its ability to selectively bind a given hormone to its cognate receptor. Several biochemical features of this interaction are important in order for hormone-receptor interactions to be physiologically relevant: (1) binding should be specific, ie, displaceable by agonist or antagonist; (2) binding should be saturable; and (3) binding should occur within the concentration range of the expected biologic response.
Both Recognition & Coupling Domains Occur on Receptors
All receptors have at least two functional domains. A recognition domain binds the hormone ligand and a second region generates a signal that couples hormone recognition to some intracellular function. This coupling, or signal transduction, occurs in two general ways. Polypeptide and protein hormones and the catecholamines bind to receptors located in the plasma membrane and thereby generate a signal that regulates various intracellular functions, often by changing the activity of an enzyme. In contrast, steroid, retinoid, and thyroid hormones interact with intracellular receptors, and it is this ligand-receptor complex that directly provides the signal, generally to specific genes whose rate of transcription is thereby affected.
The domains responsible for hormone recognition and signal generation have been identified in the protein polypeptide and catecholamine hormone receptors. Steroid, thyroid, and retinoid hormone receptors have several functional domains: one site binds the hormone; another binds to specific DNA regions; a third is involved in the interaction with other coregulator proteins that result in the activation (or repression) of gene transcription; and a fourth may specify binding to one or more other proteins that influence the intracellular trafficking of the receptor.
The dual functions of binding and coupling ultimately define a receptor, and it is the coupling of hormone binding to signal transduction—so-called receptor-effector coupling— that provides the first step in amplification of the hormonal response. This dual purpose also distinguishes the target cell receptor from the plasma carrier proteins that bind hormone but do not generate a signal (see Table 41-6).
Receptors Are Proteins
Several classes of peptide hormone receptors have been defined. For example, the insulin receptor is a heterotetramer composed of two copies of two different protein subunits (α2β2) linked by multiple disulfide bonds in which the extracellular α subunit binds insulin and the membrane-spanning β subunit transduces the signal through the tyrosine protein kinase domain located in the cytoplasmic portion of this polypeptide. The receptors for insulin-like growth factor I (IGF-I) and epidermal growth factor (EGF) are generally similar in structure to the insulin receptor. The growth hormone and prolactin receptors also span the plasma membrane of target cells but do not contain intrinsic protein kinase activity. Ligand binding to these receptors, however, results in the association and activation of a completely different protein kinase signaling pathway, the JakStat pathway. Polypeptide hormone and catecholamine receptors, which transduce signals by altering the rate of production of cAMP through G-proteins, are characterized by the presence of seven domains that span the plasma membrane. Protein kinase activation and the generation of cyclic AMP (cAMP, 3′5’-adenylic acid; see Figure 19–5) is a downstream action of this class of receptor (see Chapter 42 for further details).
A comparison of several different steroid receptors with thyroid hormone receptors revealed a remarkable conservation of the amino acid sequence in certain regions, particularly in the DNA-binding domains. This led to the realization that receptors of the steroid or thyroid type are members of a large superfamily of nuclear receptors. Many related members of this family currently have no known ligand and thus are called orphan receptors. The nuclear receptor superfamily plays a critical role in the regulation of gene transcription by hormones, as described in Chapter 42.
HORMONES CAN BE CLASSIFIED IN SEVERAL WAYS
Hormones can be classified according to chemical composition, solubility properties, location of receptors, and the nature of the signal used to mediate hormonal action within the cell. A classification based on the last two properties is illustrated in Table 41-3; and general features of each group are illustrated in Table 41-4.
TABLE 41–3 Classification of Hormones by Mechanism of Action


TABLE 41–4 General Features of Hormone Classes
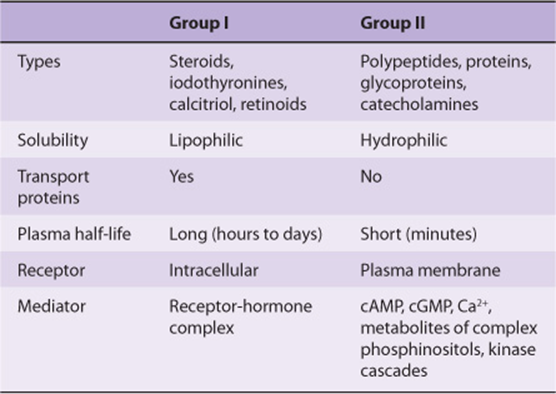
The hormones in group I are lipophilic. After secretion, these hormones associate with plasma transport or carrier proteins, a process that circumvents the problem of solubility while prolonging the plasma half-life of the hormone. The relative percentages of bound and free hormone are determined by the amount, binding affinity, and binding capacity of the transport protein. The free hormone, which is the biologically active form, readily traverses the lipophilic plasma membrane of all cells and encounters receptors in either the cytosol or nucleus of target cells. The ligand-receptor complex is assumed to be the intracellular messenger in this group.
The second major group consists of water-soluble hormones that bind to specific receptors spanning the plasma membrane of the target cell. Hormones that bind to these surface receptors of cells communicate with intracellular metabolic processes through intermediary molecules called second messengers (the hormone itself is the first messenger), which are generated as a consequence of the ligand-receptor interaction. The second messenger concept arose from an observation that epinephrine binds to the plasma membrane of certain cells and increases intracellular cAMP. This was followed by a series of experiments in which cAMP was found to mediate the effects of many hormones. Hormones that employ this mechanism are shown in group II.A of Table 41-3. Atrial natriuretic factor (ANF) uses cGMP as its second messenger (group II.B). Several hormones, many of which were previously thought to affect cAMP, appear to use ionic calcium (Ca2+) or metabolites of complex phosphoinositides (or both) as the intracellular second messenger signal. These are shown in group II.C of the table. The intracellular messenger for group II.D is a protein kinase-phosphatase cascades; several have been identified, and a given hormone may use more than one kinase cascade. A few hormones fit into more than one category, and assignments change as new information is discovered.
DIVERSITY OF THE ENDOCRINE SYSTEM
Hormones Are Synthesized in a Variety of Cellular Arrangements
Hormones are synthesized in discrete organs designed solely for this specific purpose, such as the thyroid (triiodothyronine), adrenal (glucocorticoids and mineralocorticoids), and the pituitary (TSH, FSH, LH, growth hormone, prolactin, ACTH). Some organs are designed to perform two distinct but closely related functions. For example, the ovaries produce mature oocytes and the reproductive hormones estradiol and progesterone. The testes produce mature spermatozoa and testosterone. Hormones are also produced in specialized cells within other organs such as the small intestine (glucagon-like peptide), thyroid (calcitonin), and kidney (angiotensin II). Finally, the synthesis of some hormones requires the parenchymal cells of more than one organ—eg, the skin, liver, and kidney are required for the production of 1,25(OH)2-D3 (calcitriol). Examples of this diversity in the approach to hormone synthesis, each of which has evolved to fulfill a specific purpose, are discussed below.
Hormones Are Chemically Diverse
Hormones are synthesized from a wide variety of chemical building blocks. A large series is derived from cholesterol. These include the glucocorticoids, mineralocorticoids, estrogens, progestins, and 1,25(OH)2-D3 (Figure 41–2).In some cases, a steroid hormone is the precursor molecule for another hormone. For example, progesterone is a hormone in its own right but is also a precursor in the formation of glucocorticoids, mineralocorticoids, testosterone, and estrogens. Testosterone is an obligatory intermediate in the biosynthesis of estradiol and in the formation of dihydrotestosterone (DHT). In these examples, described in detail below, the final product is determined by the cell type and the associated set of enzymes in which the precursor exists.
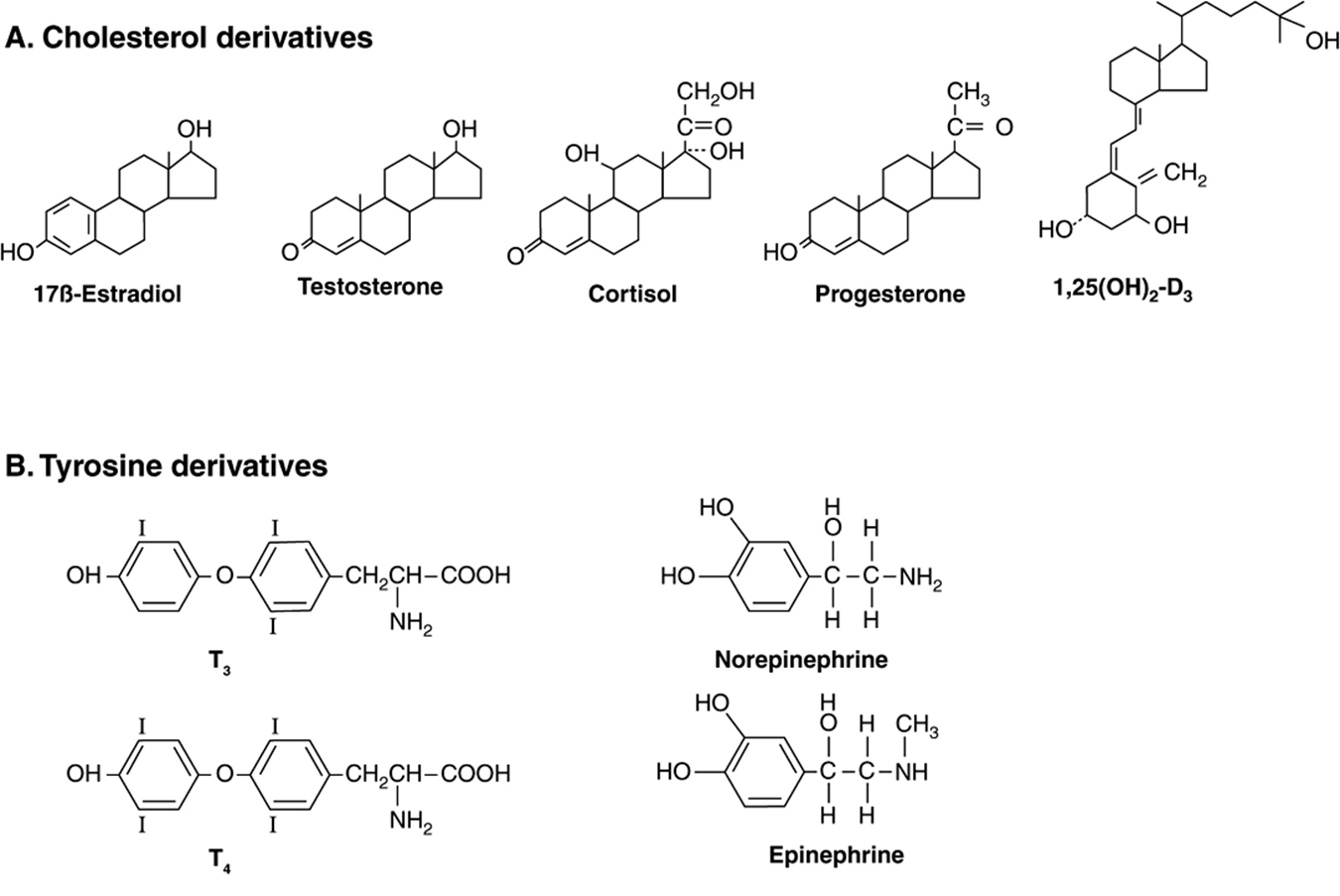
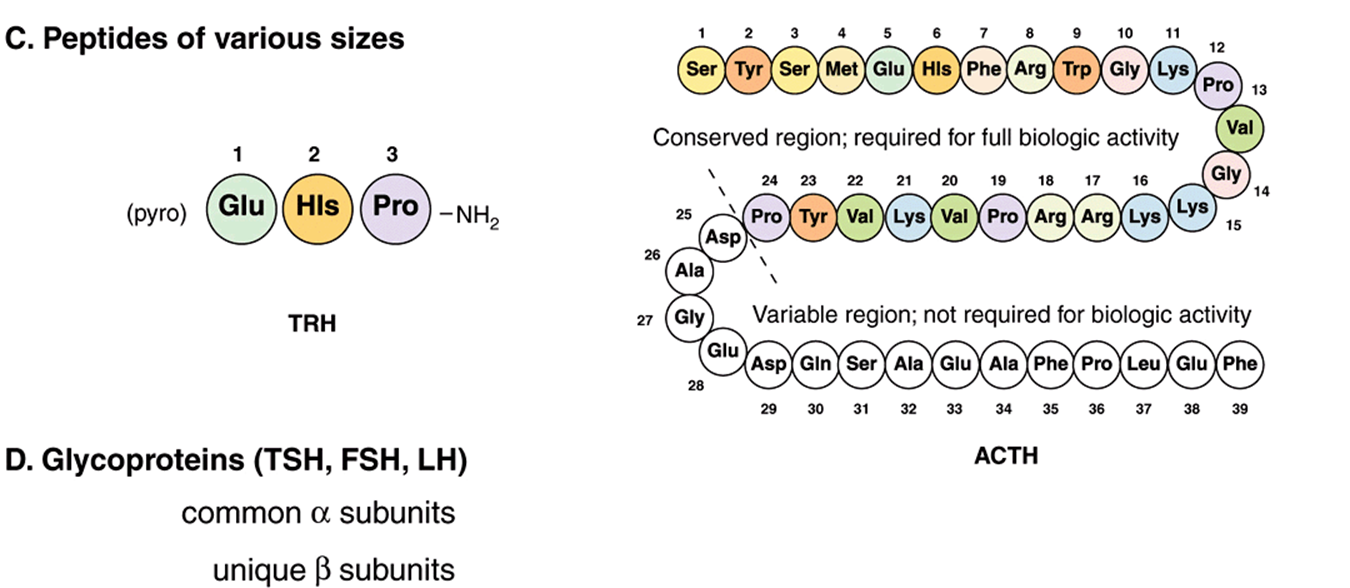
FIGURE 41–2 Chemical diversity of hormones: (A) cholesterol derivatives; (B) tyrosine derivatives; (C) peptides of various sizes; (D) glycoproteins (TSH, FSH, and LH) with common α subunits and unique β subunits.
The amino acid tyrosine is the starting point in the synthesis of both the catecholamines and thyroid hormones tetraiodothyronine (thyroxine; T4) and triiodothyronine (T3) (Figure 41–2). T3 and T4 are unique in that they require the addition of iodine (as I–) for bioactivity. Since dietary iodine is very scarce in many parts of the world, an intricate mechanism for accumulating and retaining I– has evolved.
Many hormones are polypeptides or glycoproteins. These range in size from the small thyrotropin-releasing hormone (TRH), a tripeptide, to single-chain polypeptides like adrenocorticotropic hormone (ACTH; 39 amino acids), parathyroid hormone (PTH; 84 amino acids), and growth hormone (GH; 191 amino acids) (Figure 41–2). Insulin is an AB chain heterodimer of 21 and 30 amino acids, respectively. Follicle-stimulating hormone (FSH), luteinizing hormone (LH), thyroid-stimulating hormone (TSH), and chorionic gonadotropin (CG) are glycoprotein hormones of αβ heterodimeric structure. The α chain is identical in all of these hormones, and distinct β chains impart hormone uniqueness. These hormones have a molecular mass in the range of 25-30 kDa depending on the degree of glycosylation and the length of the β chain.
Hormones Are Synthesized & Modified for Full Activity in a Variety of Ways
Some hormones are synthesized in final form and secreted immediately. Included in this class are hormones derived from cholesterol. Some, such as the catecholamines are synthesized in final form and stored in the producing cells, while others, like insulin, are synthesized from precursor molecules in the producing cell, and then are processed and secreted upon a physiologic cue (plasma glucose concentrations). Finally, still others are converted to active forms from precursor molecules in the periphery (T3 and DHT). All of these examples are discussed in more detail below.
MANY HORMONES ARE MADE FROM CHOLESTEROL
Adrenal Steroidogenesis
The adrenal steroid hormones are synthesized from cholesterol, which is mostly derived from the plasma, but a small portion is synthesized in situ from acetyl-CoA via mevalonate and squalene. Much of the cholesterol in the adrenal is esterified and stored in cytoplasmic lipid droplets. Upon stimulation of the adrenal by ACTH, an esterase is activated, and the free cholesterol formed is transported into the mitochondrion, where a cytochrome P450 side chain cleavage enzyme (P450scc) converts cholesterol to pregnenolone. Cleavage of the side chain involves sequential hydroxylations, first at C22 and then at C20, followed by side chain cleavage (removal of the six-carbon fragment isocaproaldehyde) to give the 21-carbon steroid (Figure 41–3, top). An ACTH-dependent steroidogenic acute regulatory (StAR) protein is essential for the transport of cholesterol to P450scc in the inner mitochondrial membrane.
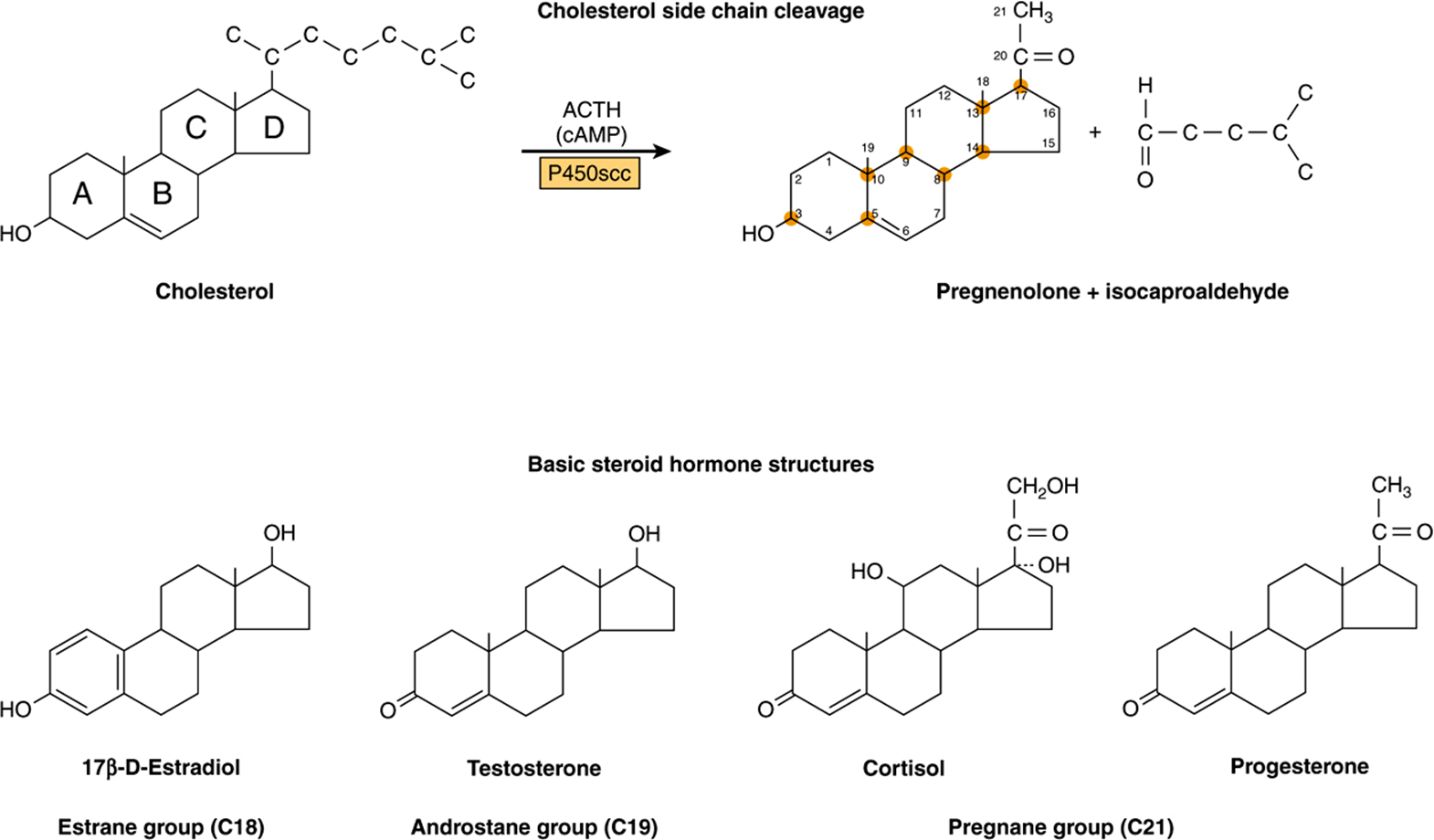
FIGURE 41–3 Cholesterol side-chain cleavage and basic steroid hormone structures. The basic sterol rings are identified by the letters A-D. The carbon atoms are numbered 1-21, starting with the A ring.
All mammalian steroid hormones are formed from cholesterol via pregnenolone through a series of reactions that occur in either the mitochondria or endoplasmic reticulum of the producing cell. Hydroxylases that require molecular oxygen and NADPH are essential, and dehydrogenases, an isomerase, and a lyase reaction are also necessary for certain steps. There is cellular specificity in adrenal steroidogenesis. For instance, 18-hydroxylase and 19-hydroxysteroid dehydrogenises, which are required for aldosterone synthesis, are found only in the zona glomerulosa cells (the outer region of the adrenal cortex), so that the biosynthesis of this mineralocorticoid is confined to this region. A schematic representation of the pathways involved in the synthesis of the three major classes of adrenal steroids is presented in Figure 41–4. The enzymes are shown in the rectangular boxes, and the modifications at each step are shaded.
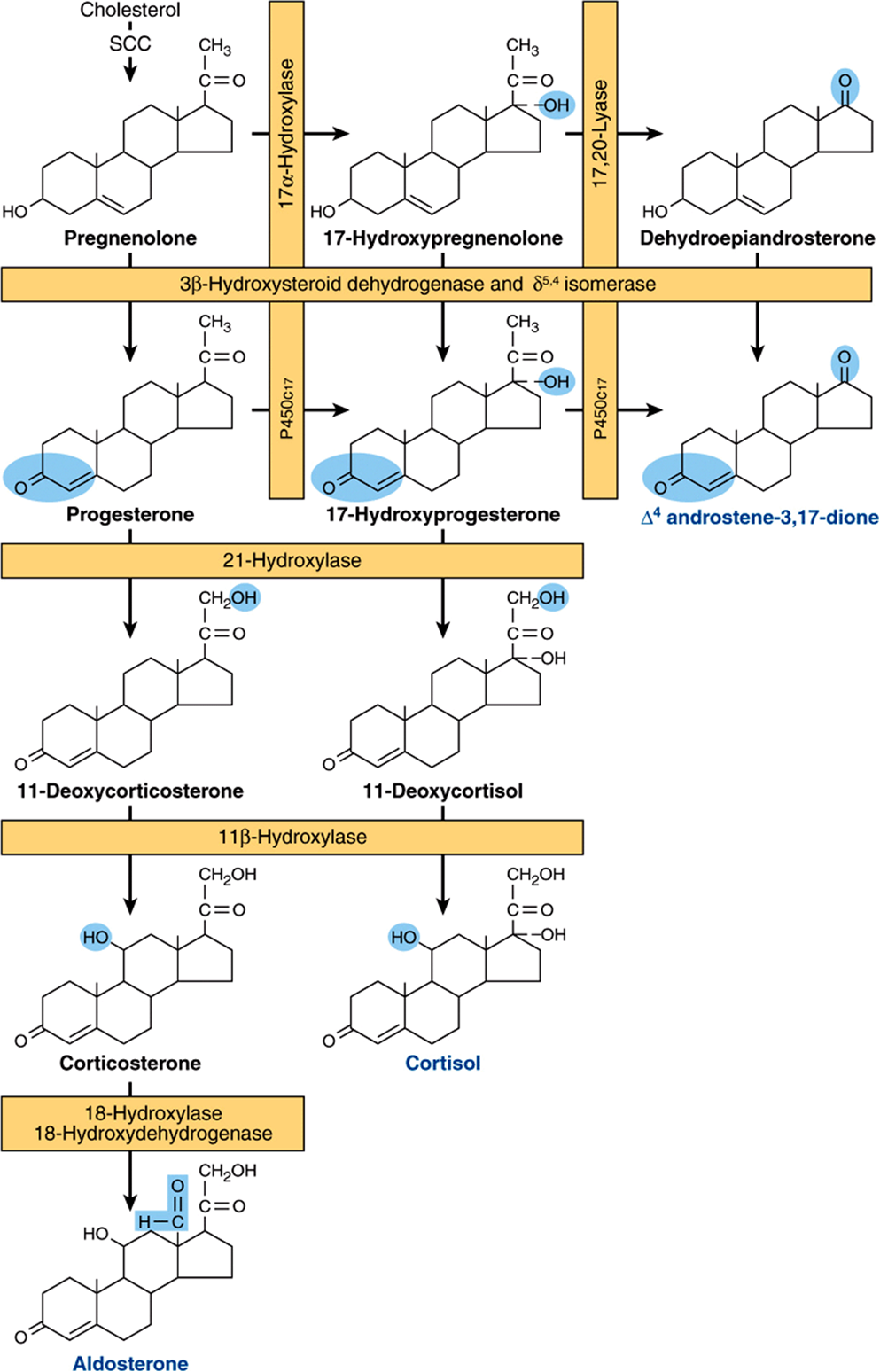
FIGURE 41–4 Pathways involved in the synthesis of the three major classes of adrenal steroids (mineralocorticoids, glucocorticoids, and androgens). Enzymes are shown in the rectangular boxes, and the modifications at each step are shaded. Note that the 17α-hydroxylase and 17,20-lyase activities are both part of one enzyme, designated P450c17. (Slightly modified and reproduced, with permission, from Harding BW: In: Endocrinology, vol 2. DeGroot LJ (editors). Grune & Stratton, 1979. Copyright © 1979 Elsevier Inc. Reprinted with permission from Elsevier.)
Mineralocorticoid Synthesis
Synthesis of aldosterone follows the mineralocorticoid pathway and occurs in the zona glomerulosa. Pregnenolone is converted to progesterone by the action of two smooth endoplasmic reticulum enzymes, 3β-hydroxysteroid dehydrogenase (3β-OHSD) and Δ5,4-isomerase. Progesterone is hydroxylated at the C21 position to form 11-deoxycorticosterone (DOC), which is an active (Na+-retaining) mineralocorticoid. The next hydroxylation, at C11, produces corticosterone, which has glucocorticoid activity and is a weak mineralocorticoid (it has <5% of the potency of aldosterone). In some species (eg, rodents), it is the most potent glucocorticoid. C21 hydroxylation is necessary for both mineralocorticoid and glucocorticoid activity, but most steroids with a C17 hydroxyl group have more glucocorticoid and less mineralocorticoid action. In the zona glomerulosa, which does not have the smooth endoplasmic reticulum enzyme 17α-hydroxylase, a mitochondrial 18-hydroxylase is present. The 18-hydroxylase (aldosterone synthase) acts on corticosterone to form 18-hydroxycorticosterone, which is changed to aldosterone by conversion of the 18-alcohol to an aldehyde. This unique distribution of enzymes and the special regulation of the zona glomerulosa by K+ and angiotensin II have led some investigators to suggest that, in addition to the adrenal being two glands, the adrenal cortex is actually two separate organs.
Glucocorticoid Synthesis
Cortisol synthesis requires three hydroxylases located in the fasciculata and reticularis zones of the adrenal cortex that act sequentially on the C17, C21; and C11 positions. The first two reactions are rapid, while C11 hydroxylation is relatively slow. If the C11 position is hydroxylated first, the action of 17α-hydroxylase is impeded and the mineralocorticoid pathway is followed (forming corticosterone or aldosterone, depending on the cell type). 17α-Hydroxylase is a smooth endoplasmic reticulum enzyme that acts upon either progesterone or, more commonly, pregnenolone. 17α-Hydroxyprogesterone is hydroxylated at C21 to form 11-deoxycortisol, which is then hydroxylated at C11 to form cortisol, the most potent natural glucocorticoid hormone in humans. 21-Hydroxylase is a smooth endoplasmic reticulum enzyme, whereas 11β-hydroxylase is a mitochondrial enzyme. Steroidogenesis thus involves the repeated shuttling of substrates into and out of the mitochondria.
Androgen Synthesis
The major androgen or androgen precursor produced by the adrenal cortex is dehydroepiandrosterone (DHEA). Most 17-hydroxypregnenolone follows the glucocorticoid pathway, but a small fraction is subjected to oxidative fission and removal of the two-carbon side chain through the action of 17,20-lyase. The lyase activity is actually part of the same enzyme (P450c17) that catalyzes 17α-hydroxylation. This is therefore a dual-function protein. The lyase activity is important in both the adrenals and the gonads and acts exclusively on 17α-hydroxy-containing molecules. Adrenal androgen production increases markedly if glucocorticoid biosynthesis is impeded by the lack of one of the hydroxylases (adrenogenital syndrome). DHEA is really a prohormone since the actions of 3β-OHSD and Δ5,4-isomerase convert the weak androgen DHEA into the more potent androstenedione. Small amounts of androstenedione are also formed in the adrenal by the action of the lyase on 17α-hydroxyprogesterone. Reduction of androstenedione at the C1 7 position results in the formation of testosterone, the most potent adrenal androgen. Small amounts of testosterone are produced in the adrenal by this mechanism, but most of this conversion occurs in the testes.
Testicular Steroidogenesis
Testicular androgens are synthesized in the interstitial tissue by the Leydig cells. The immediate precursor of the gonadal steroids, as for the adrenal steroids, is cholesterol. The ratelimiting step, as in the adrenal, is delivery of cholesterol to the inner membrane of the mitochondria by the transport protein StAR. Once in the proper location, cholesterol is acted upon by the side chain cleavage enzyme P450scc. The conversion of cholesterol to pregnenolone is identical in adrenal, ovary, and testis. In the latter two tissues, however, the reaction is promoted by LH rather than ACTH.
The conversion of pregnenolone to testosterone requires the action of five enzyme activities contained in three proteins: (1) 3β-hydroxysteroid dehydrogenase (β-OHSD) and Δ5,4-isomerase; (2) 17α-hydroxylase and 17,20-lyase; and (3) 17β-hydroxysteroid dehydrogenase (17β-OHSD). This sequence, referred to as the progesterone (or Δ4) pathway, is shown on the right side of Figure 41–5. Pregnenolone can also be converted to testosterone by the dehydroepiandrosterone (or Δ5) pathway, which is illustrated on the left side of Figure 41–5. The Δ5 route appears to be most used in human testes.
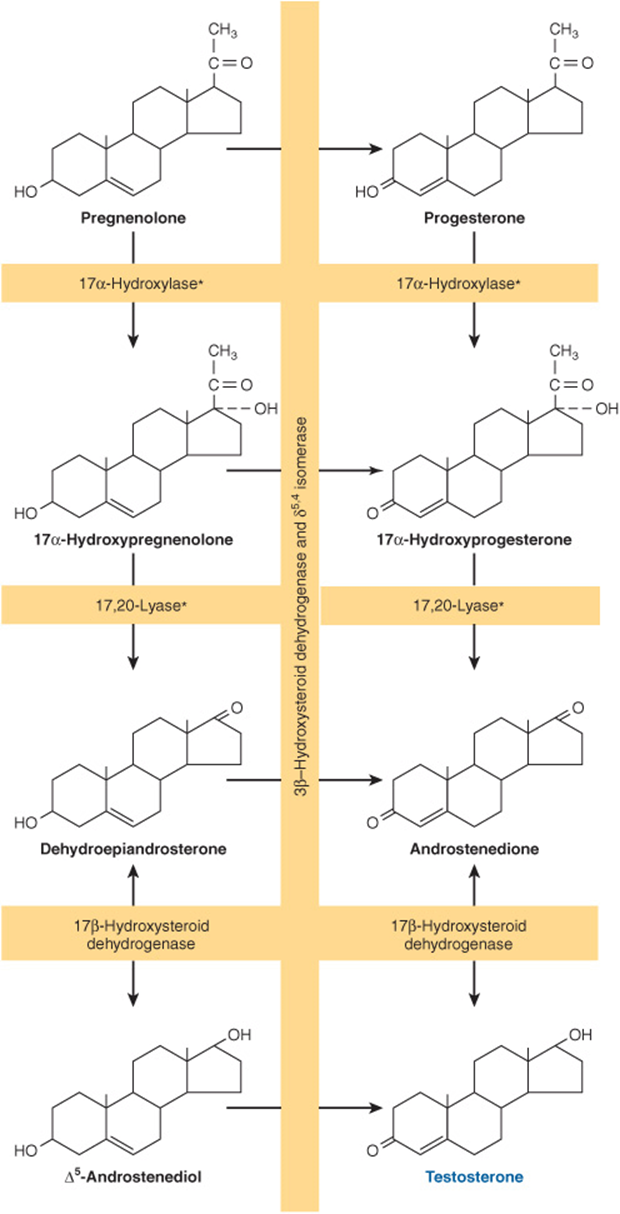
FIGURE 41–5 Pathways of testosterone biosynthesis. The pathway on the left side of the figure is called the Δ5 or dehydroepiandrosterone pathway; the pathway on the right side is called the Δ4 or progesterone pathway. The asterisk indicates that the 17α-hydroxylase and 17,20-lyase activities reside in a single protein, P450c17.
The five enzyme activities are localized in the microsomal fraction in rat testes, and there is a close functional association between the activities of 3β-OHSD and Δ5,4-isomerase and between those of a 17α-hydroxylase and 17,20-lyase. These enzyme pairs, both contained in a single protein, are shown in the general reaction sequence in Figure 41–5.
DHT Is Formed from Testosterone in Peripheral Tissues
Testosterone is metabolized by two pathways. One involves oxidation at the 17 position, and the other involves reduction of the A ring double bond and the 3-ketone. Metabolism by the first pathway occurs in many tissues, including liver, and produces 17-ketosteroids that are generally inactive or less active than the parent compound. Metabolism by the second pathway, which is less efficient, occurs primarily in target tissues and produces the potent metabolite DHT.
The most significant metabolic product of testosterone is DHT, since in many tissues, including prostate, external genitalia, and some areas of the skin, this is the active form of the hormone. The plasma content of DHT in the adult male is about one-tenth that of testosterone, and ~400 μg of DHT is produced daily as compared with about 5 mg of testosterone. About 50-100 μg of DHT are secreted by the testes. The rest is produced peripherally from testosterone in a reaction catalyzed by the NADPH-dependent 5α-reductase (Figure 41–6). Testosterone can thus be considered a prohormone since it is converted into a much more potent compound (DHT) and since most of this conversion occurs outside the testes. Some estradiol is formed from the peripheral aromatization of testosterone, particularly in males.
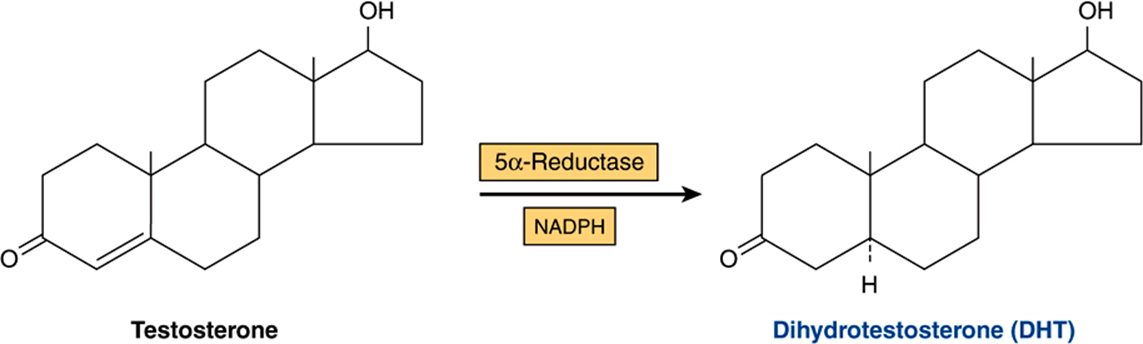
FIGURE 41–6 Dihydrotestosterone is formed from testosterone through action of the enzyme 5α-reductase.
Ovarian Steroidogenesis
The estrogens are a family of hormones synthesized in a variety of tissues. 17β-Estradiol is the primary estrogen of ovarian origin. In some species, estrone, synthesized in numerous tissues, is more abundant. In pregnancy, relatively more estriol is produced, and this comes from the placenta. The general pathway and the subcellular localization of the enzymes involved in the early steps of estradiol synthesis are the same as those involved in androgen biosynthesis. Features unique to the ovary are illustrated in Figure 41–7.
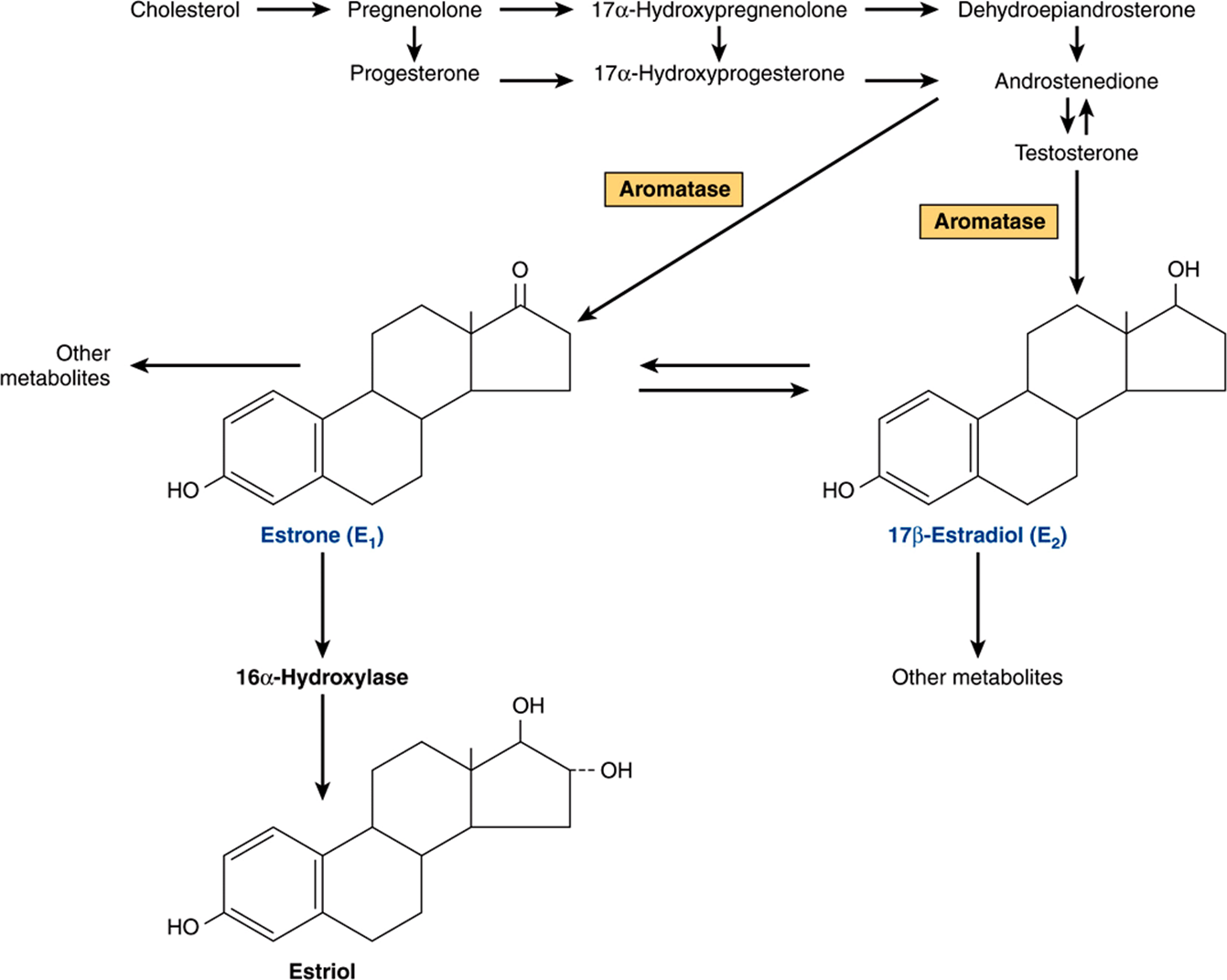
FIGURE 41–7 Biosynthesis of estrogens. (Slightly modified and reproduced, with permission, from Ganong WF: Review of Medical Physiology, 21st ed. McGraw-Hill, 2005.)
Estrogens are formed by the aromatization of androgens in a complex process that involves three hydroxylation steps, each of which requires O2 and NADPH. The aromatase enzyme complex is thought to include a P450 monooxygenase. Estradiol is formed if the substrate of this enzyme complex is testosterone, whereas estrone results from the aromatization of androstenedione.
The cellular source of the various ovarian steroids has been difficult to unravel, but a transfer of substrates between two cell types is involved. Theca cells are the source of androstenedione and testosterone. These are converted by the aro-matase enzyme in granulosa cells to estrone and estradiol, respectively. Progesterone, a precursor for all steroid hormones, is produced and secreted by the corpus luteum as an end-product hormone because these cells do not contain the enzymes necessary to convert progesterone to other steroid hormones (Figure 41–8).
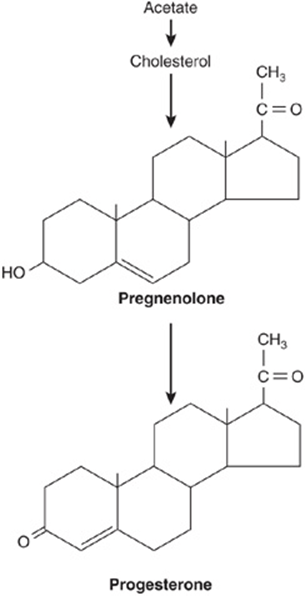
FIGURE 41–8 Biosynthesis of progesterone in the corpus luteum.
Significant amounts of estrogens are produced by the peripheral aromatization of androgens. In human males, the peripheral aromatization of testosterone to estradiol (E2) accounts for 80% of the production of the latter. In females, adrenal androgens are important substrates since as much as 50% of the E2 produced during pregnancy comes from the aromatization of androgens. Finally, conversion of androstenedione to estrone is the major source of estrogens in postmenopausal women. Aromatase activity is present in adipose cells and also in liver, skin, and other tissues. Increased activity of this enzyme may contribute to the “estrogenization” that characterizes such diseases as cirrhosis of the liver, hyperthyroidism, aging, and obesity. Aromatase inhibitors show promise as therapeutic agents in breast cancer and possibly in other female reproductive tract malignancies.
1,25(OH)2-D3 (Calcitriol) Is Synthesized from a Cholesterol Derivative
1,25(OH)2-D3 is produced by a complex series of enzymatic reactions that involve the plasma transport of precursor molecules to a number of different tissues (Figure 41–9). One of these precursors is vitamin D—really not a vitamin, but this common name persists. The active molecule, 1,25(OH)2-D3, is transported to other organs where it activates biologic processes in a manner similar to that employed by the steroid hormones.
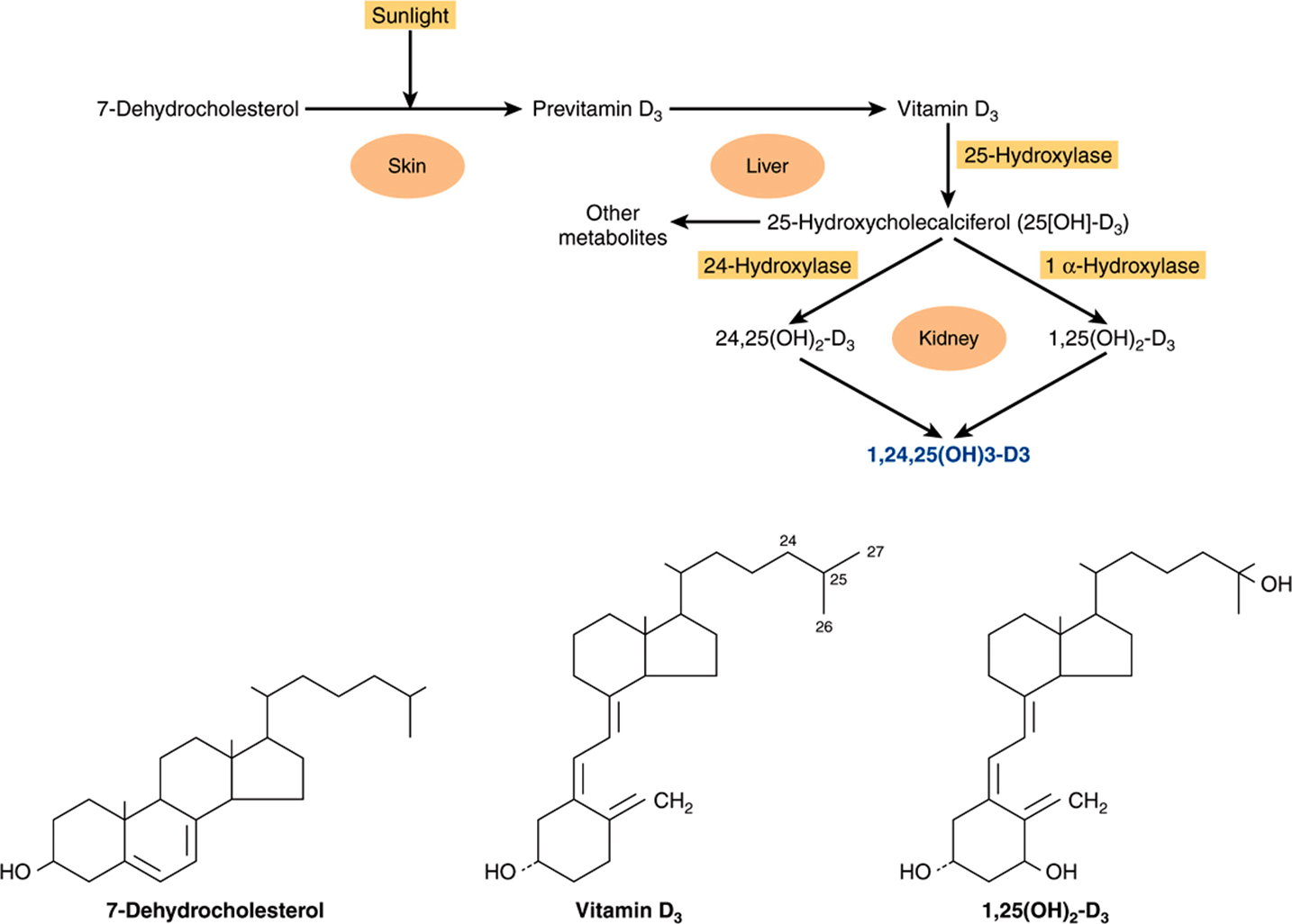
FIGURE 41–9 Formation and hydroxylation of vitamin D3. 25-Hydroxylation takes place in the liver, and the other hydroxylations occur in the kidneys. 25,26(OH)2-D3 and 1,25,26(OH)3-D3 are probably formed as well. The structures of 7-dehydrocholesterol, vitamin D3, and 1,25(OH)2-D3 are also shown. (Modified and reproduced, with permission, from Ganong WF: Review of Medical Physiology, 21st ed. McGraw-Hill, 2005.)
Skin
Small amounts of the precursor for 1,25(OH)2-D3 synthesis are present in food (fish liver oil, and egg yolk), but most of the precursor for 1,25(OH)2-D3 synthesis is produced in the malpighian layer of the epidermis from 7-dehydrocholesterol in an ultraviolet light-mediated, nonenzymatic photolysis reaction. The extent of this conversion is related directly to the intensity of the exposure and inversely to the extent of pigmentation in the skin. There is an age-related loss of 7-dehydrocholesterol in the epidermis that may be related to the negative calcium balance associated with old age.
Liver
A specific transport protein called the vitamin D-binding protein binds vitamin D3 and its metabolites and moves vitamin D3 from the skin or intestine to the liver, where it undergoes 25-hydroxylation, the first obligatory reaction in the production of 1,25(OH)2-D3. 25-Hydroxylation occurs in the endoplasmic reticulum in a reaction that requires magnesium, NADPH, molecular oxygen, and an uncharacterized cytoplasmic factor. Two enzymes are involved: an NADPH-dependent cytochrome P450 reductase and a cytochrome P450. This reaction is not regulated, and it also occurs with low efficiency in kidney and intestine. The 25(OH)2-D3 enters the circulation, where it is the major form of vitamin D found in plasma, and is transported to the kidney by the vitamin D-binding protein.
Kidney
25(OH)2-D3 is a weak agonist and must be modified by hydroxylation at position C1 for full biologic activity. This is accomplished in mitochondria of the renal proximal convoluted tubule by a three-component monooxygenase reaction that requires NADPH, Mg2+, molecular oxygen, and at least three enzymes: (1) a flavoprotein, renal ferredoxin reductase; (2) an iron sulfur protein, renal ferredoxin; and (3) cytochrome P450. This system produces 1,25(OH)2-D3; which is the most potent naturally occurring metabolite of vitamin D.
CATECHOLAMINES & THYROID HORMONES ARE MADE FROM TYROSINE
Catecholamines Are Synthesized in Final Form & Stored in Secretion Granules
Three amines—dopamine, norepinephrine, and epinephrine—are synthesized from tyrosine in the chromaffin cells of the adrenal medulla. The major product of the adrenal medulla is epinephrine. This compound constitutes about 80% of the catecholamines in the medulla, and it is not made in extramedullary tissue. In contrast, most of the norepinephrine present in organs innervated by sympathetic nerves is made in situ (about 80% of the total), and most of the rest is made in other nerve endings and reaches the target sites via the circulation. Epinephrine and norepinephrine may be produced and stored in different cells in the adrenal medulla and other chromaffin tissues.
The conversion of tyrosine to epinephrine requires four sequential steps: (1) ring hydroxylation; (2) decarboxylation; (3) side-chain hydroxylation to form norepinephrine; and (4) N-methylation to form epinephrine. The biosynthetic pathway and the enzymes involved are illustrated in Figure 41–10.
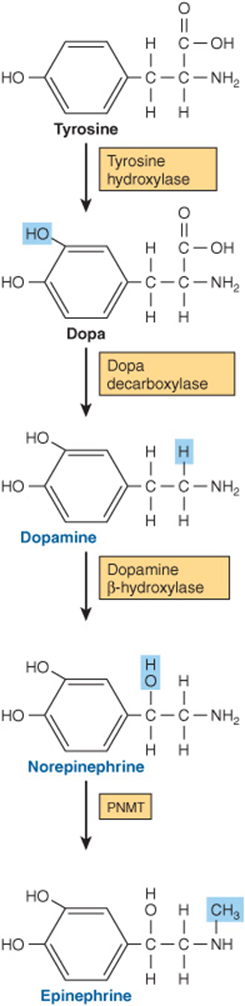
FIGURE 41–10 Biosynthesis of catecholamines. (PNMT, phenylethanolamine-N-methyltransferase.)
Tyrosine Hydroxylase Is Rate-Limiting for Catecholamine Biosynthesis
Tyrosine is the immediate precursor of catecholamines, and tyrosine hydroxylase is the rate-limiting enzyme in catecholamine biosynthesis. Tyrosine hydroxylase is found in both soluble and particle-bound forms only in tissues that synthesize catecholamines; it functions as an oxidoreductase, with tetrahydropteridine as a cofactor, to convert L-tyrosine to L-dihydroxyphenylalanine (L-dopa). As the rate-limiting enzyme, tyrosine hydroxylase is regulated in a variety of ways. The most important mechanism involves feedback inhibition by the catecholamines, which compete with the enzyme for the pteridine cofactor. Catecholamines cannot cross the blood-brain barrier; hence, in the brain they must be synthesized locally. In certain central nervous system diseases (eg, Parkinson’s disease), there is a local deficiency of dopamine synthesis. L-Dopa, the precursor of dopamine, readily crosses the blood-brain barrier and so is an important agent in the treatment of Parkinson disease.
Dopa Decarboxylase Is Present in All Tissues
This soluble enzyme requires pyridoxal phosphate for the conversion of L-dopa to 3,4-dihydroxyphenylethylamine (dopamine). Compounds that resemble L-dopa, such as α-methyldopa, are competitive inhibitors of this reaction. α-Methyldopa is effective in treating some kinds of hypertension.
Dopamine β-Hydroxylase (DBH) Catalyzes the Conversion of Dopamine to Norepinephrine
DBH is a monooxygenase and uses ascorbate as an electron donor, copper at the active site, and fumarate as modulator. DBH is in the particulate fraction of the medullary cells, probably in the secretion granule; thus, the conversion of dopamine to norepinephrine occurs in this organelle.
Phenylethanolamine-N-Methyltransferase (PNMT) Catalyzes the Production of Epinephrine
PNMT catalyzes the N-methylation of norepinephrine to form epinephrine in the epinephrine-forming cells of the adrenal medulla. Since PNMT is soluble, it is assumed that norepinephrine-to-epinephrine conversion occurs in the cytoplasm. The synthesis of PNMT is induced by glucocorticoid hormones that reach the medulla via the intra-adrenal portal system. This special system provides for a 100-fold steroid concentration gradient over systemic arterial blood, and this high intra-adrenal concentration appears to be necessary for the induction of PNMT.
T3 & T4 Illustrate the Diversity in Hormone Synthesis
The formation of triiodothyronine (T3) and tetraiodothyronine (thyroxine; T4) (see Figure 41–2) illustrates many of the principles of diversity discussed in this chapter. These hormones require a rare element (iodine) for bioactivity; they are synthesized as part of a very large precursor molecule (thyroglobulin); they are stored in an intracellular reservoir (colloid); and there is peripheral conversion of T4 to T3, which is a much more active hormone.
The thyroid hormones T3 and T4 are unique in that iodine (as iodide) is an essential component of both. In most parts of the world, iodine is a scarce component of soil, and for that reason there is little in food. A complex mechanism has evolved to acquire and retain this crucial element and to convert it into a form suitable for incorporation into organic compounds. At the same time, the thyroid must synthesize thyronine from tyrosine, and this synthesis takes place in thyroglobulin (Figure 41–11).
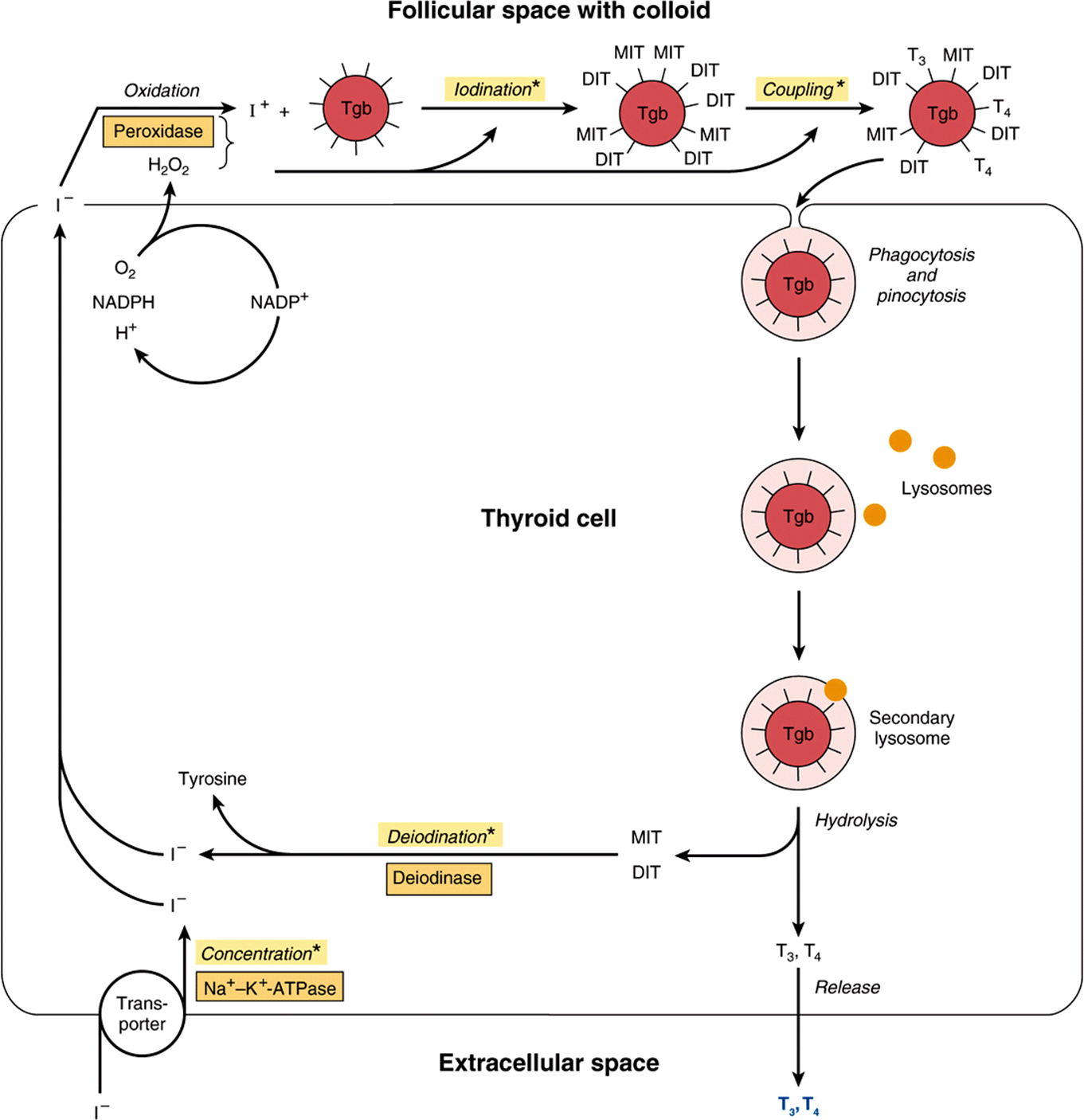
FIGURE 41–11 Model of iodide metabolism in the thyroid follicle. A follicular cell is shown facing the follicular lumen (top) and the extracellular space (bottom). Iodide enters the thyroid primarily through a transporter (bottom left). Thyroid hormone synthesis occurs in the follicular space through a series of reactions, many of which are peroxidase-mediated. Thyroid hormones, stored in the colloid in the follicular space, are released from thyroglobulin by hydrolysis inside the thyroid cell. (DIT, diiodotyrosine; MIT, monoiodotyrosine; Tgb, thyroglobulin; T3, triiodothyronine; T4, tetraiodothyronine.) Asterisks indicate steps or processes where inherited enzyme deficiencies cause congenital goiter and often result in hypothyroidism.
Thyroglobulin is the precursor of T4 and T3. It is a large iodinated, glycosylated protein with a molecular mass of 660 kDa. Carbohydrate accounts for 8-10% of the weight of thyroglobulin and iodide for about 0.2-1%, depending upon the iodine content in the diet. Thyroglobulin is composed of two large subunits. It contains 115 tyrosine residues, each of which is a potential site of iodination. About 70% of the iodide in thyroglobulin exists in the inactive precursors, monoiodotyrosine (MIT) and diiodotyrosine (DIT), while 30% is in the iodothyronyl residues, T4and T3; When iodine supplies are sufficient, the T4:T3 ratio is about 7:1. In iodine deficiency, this ratio decreases, as does the DIT:MIT ratio. Thyroglobulin, a large molecule of about 5000 amino acids, provides the conformation required for tyrosyl coupling and iodide organification necessary in the formation of the diaminoacid thyroid hormones. It is synthesized in the basal portion of the cell and moves to the lumen, where it is a storage form of T3 and T4 in the colloid; several weeks’ supply of these hormones exist in the normal thyroid. Within minutes after stimulation of the thyroid by TSH, colloid reenters the cell and there is a marked increase of phagolysosome activity. Various acid proteases and peptidases hydrolyze the thyroglobulin into its constituent amino acids, including T4 and T3, which are discharged into the extracellular space (see Figure 41–11). Thyroglobulin is thus a very large prohormone.
Iodide Metabolism Involves Several Discrete Steps
The thyroid is able to concentrate I– against a strong electrochemical gradient. This is an energy-dependent process and is linked to the Na+-K+-ATPase-dependent thyroidal I– transporter. The ratio of iodide in thyroid to iodide in serum (T:S ratio) is a reflection of the activity of this transporter. This activity is primarily controlled by TSH and ranges from 500:1 in animals chronically stimulated with TSH to 5:1 or less in hypophysectomized animals (no TSH). The T:S ratio in humans on a normal iodine diet is about 25:1.
The thyroid is the only tissue that can oxidize I– to a higher valence state, an obligatory step in I– organification and thyroid hormone biosynthesis. This step involves a heme-containing peroxidase and occurs at the luminal surface of the follicular cell. Thyroperoxidase, a tetrameric protein with a molecular mass of 60 kDa, requires hydrogen peroxide as an oxidizing agent. The H2O2 is produced by an NADPH-dependent enzyme resembling cytochrome; reductase. A number of compounds inhibit I– oxidation and therefore its subsequent incorporation into MIT and DIT. The most important of these are the thiourea drugs. They are used as antithyroid drugs because of their ability to inhibit thyroid hormone biosynthesis at this step. Once iodination occurs, the iodine does not readily leave the thyroid. Free tyrosine can be iodinated, but it is not incorporated into proteins since no tRNA recognizes iodinated tyrosine.
The coupling of two DIT molecules to form T4—or of an MIT and DIT to form T3—occurs within the thyroglobulin molecule. A separate coupling enzyme has not been found, and since this is an oxidative process it is assumed that the same thyroperoxidase catalyzes this reaction by stimulating free radical formation of iodotyrosine. This hypothesis is supported by the observation that the same drugs which inhibit I–oxidation also inhibit coupling. The formed thyroid hormones remain as integral parts of thyroglobulin until the latter is degraded, as described above.
A deiodinase removes I– from the inactive mono and diiodothyronine molecules in the thyroid. This mechanism provides a substantial amount of the I– used in T3 and T4 biosynthesis. A peripheral deiodinase in target tissues such as pituitary, kidney, and liver selectively removes I– from the 5′ position of T4 to make T3 (see Figure 41–2), which is a much more active molecule. In this sense, T4 can be thought of as a prohormone, though it does have some intrinsic activity.
Several Hormones Are Made from Larger Peptide Precursors
Formation of the critical disulfide bridges in insulin requires that this hormone be first synthesized as part of a larger precursor molecule, proinsulin. This is conceptually similar to the example of the thyroid hormones, which can only be formed in the context of a much larger molecule. Several other hormones are synthesized as parts of large precursor molecules, not because of some special structural requirement but rather as a mechanism for controlling the available amount of the active hormone. PTH and angiotensin II are examples of this type of regulation. Another interesting example is the POMC protein, which can be processed into many different hormones in a tissue-specific manner. These examples are discussed in detail below.
Insulin Is Synthesized as a Preprohormone & Modified Within the β Cell
Insulin has an AB heterodimeric structure with one intrachain (A6-A11) and two interchain disulfide bridges (A7-B7 and A20-B19) (Figure 41–12). The A and B chains could be synthesized in the laboratory, but attempts at a biochemical synthesis of the mature insulin molecule yielded very poor results. The reason for this became apparent when it was discovered that insulin is synthesized as a preprohormone (molecular weight ~11,500), which is the prototype for peptides that are processed from larger precursor molecules. The hydrophobic 23-amino-acid pre-, or leader, sequence directs the molecule into the cisternae of the endoplasmic reticulum and then is removed. This results in the 9000-MW proinsulin molecule, which provides the conformation necessary for the proper and efficient formation of the disulfide bridges. As shown in Figure 41–12, the sequence of proinsulin, starting from the amino terminal, is B chain—connecting (C) peptide—A chain. The proinsulin molecule undergoes a series of site-specific peptide cleavages that result in the formation of equimolar amounts of mature insulin and C-peptide. These enzymatic cleavages are summarized in Figure 41–12.
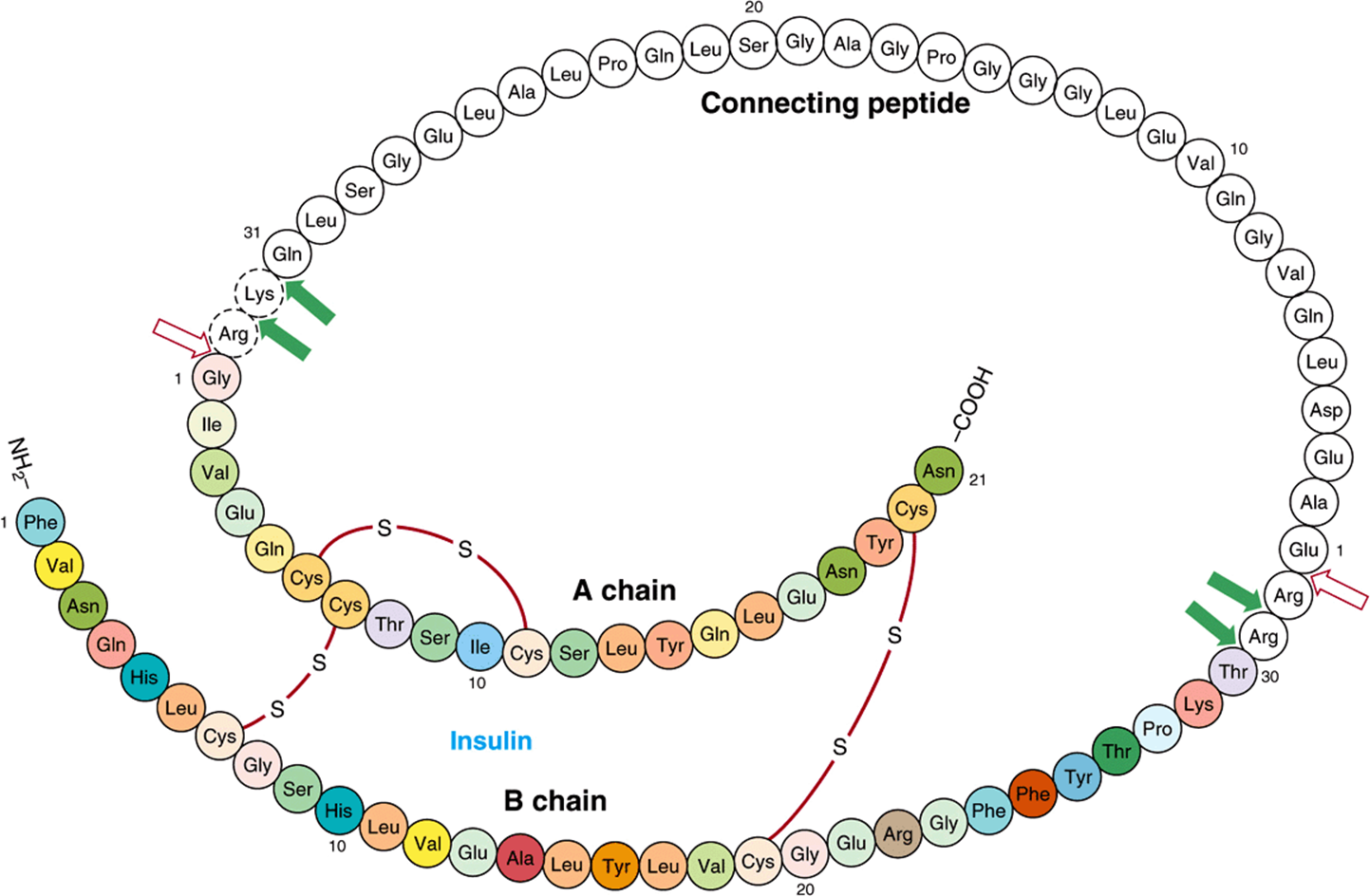
FIGURE 41–12 Structure of human proinsulin. Insulin and C-peptide molecules are connected at two sites by dipeptide links. An initial cleavage by a trypsin-like enzyme (open arrows) followed by several cleavages by a carboxypeptidase-like enzyme (solid arrows) results in the production of the heterodimeric (AB) insulin molecule (colored) and the C-peptide (white).
PTH Is Secreted as an 84-Amino-Acid Peptide
The immediate precursor of PTH is proPTH, which differs from the native 84-amino-acid hormone by having a highly basic hexapeptide amino terminal extension. The primary gene product and the immediate precursor for proPTH is the 115-amino-acid preproPTH. This differs from proPTH by having an additional 25-amino-acid amino terminal extension that, in common with the other leader or signal sequences characteristic of secreted proteins, is hydrophobic. The complete structure of preproPTH and the sequences of proPTH and PTH are illustrated in Figure 41–13. PTH1-34 has full biologic activity, and the region 25–34 is primarily responsible for receptor binding.
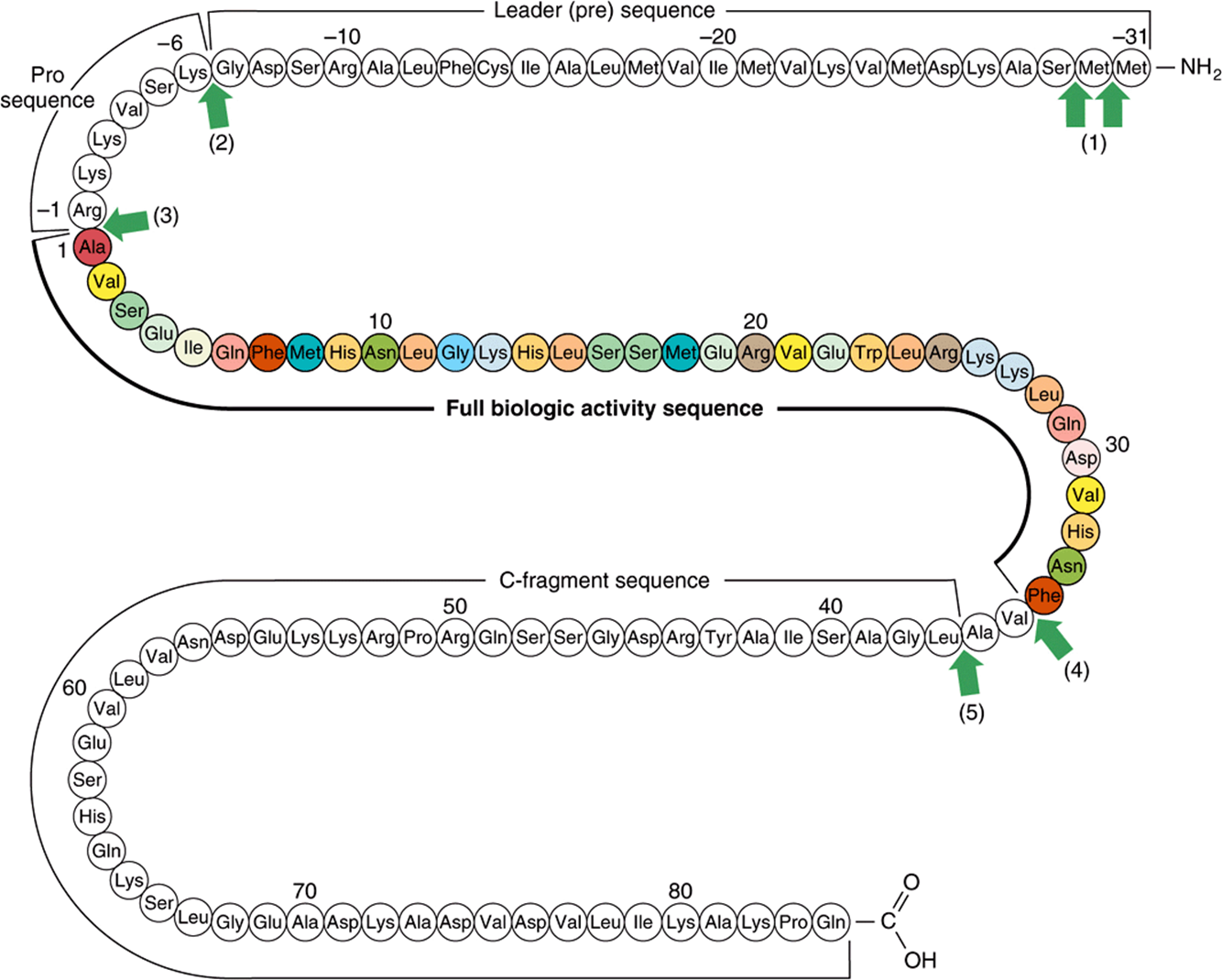
FIGURE 41–13 Structure of bovine preproparathyroid hormone. Arrows indicate sites cleaved by processing enzymes in the parathyroid gland and in the liver after secretion of the hormone (1-5). The biologically active region of the molecule (colored) is flanked by sequence not required for activity on target receptors. (Slightly modified and reproduced, with permission, from Habener JF: Recent advances in parathyroid hormone research. Clin Biochem 1981;14:223. Copyright © 1981. Reprinted with permission from Elsevier.)
The biosynthesis of PTH and its subsequent secretion are regulated by the plasma ionized calcium (Ca2+) concentration through a complex process. An acute decrease of Ca2+ results in a marked increase of PTH mRNA, and this is followed by an increased rate of PTH synthesis and secretion. However, about 80-90% of the proPTH synthesized cannot be accounted for as intact PTH in cells or in the incubation medium of experimental systems. This finding led to the conclusion that most of the proPTH synthesized is quickly degraded. It was later discovered that this rate of degradation decreases when Ca2+ concentrations are low, and it increases when Ca2+ concentrations are high. A Ca2+ receptor on the surface of the parathyroid cell mediates these effects. Very specific fragments of PTH are generated during its proteolytic digestion (Figure 41–13). A number of proteolytic enzymes, including cathepsins B and D, have been identified in parathyroid tissue. Cathepsin B cleaves PTH into two fragments: PTH1-36 and PTH37-84. PTH37-84 is not further degraded; however, PTH1-36 is rapidly and progressively cleaved into di- and tripeptides. Most of the proteolysis of PTH occurs within the gland, but a number of studies confirm that PTH, once secreted, is proteolytically degraded in other tissues, especially the liver, by similar mechanisms.
Angiotensin II Is Also Synthesized from a Large Precursor
The renin-angiotensin system is involved in the regulation of blood pressure and electrolyte metabolism (through production of aldosterone). The primary hormone involved in these processes is angiotensin II, an octapeptide made from angiotensinogen (Figure 41–14). Angiotensinogen, a large α2-globulin made in liver, is the substrate for renin, an enzyme produced in the juxtaglomerular cells of the renal afferent arteriole. The position of these cells makes them particularly sensitive to blood pressure changes, and many of the physiologic regulators of renin release act through renal baroreceptors. The juxtaglomerular cells are also sensitive to changes of Na+ and Cl– concentration in the renal tubular fluid; therefore, any combination of factors that decreases fluid volume (dehydration, decreased blood pressure, fluid, or blood loss) or decreases NaCl concentration stimulates renin release. Renal sympathetic nerves that terminate in the juxtaglomerular cells mediate the central nervous system and postural effects on renin release independently of the baroreceptor and salt effects, a mechanism that involves the β-adrenergic receptor. Renin acts upon the substrate angiotensinogen to produce the decapeptide angiotensin I.
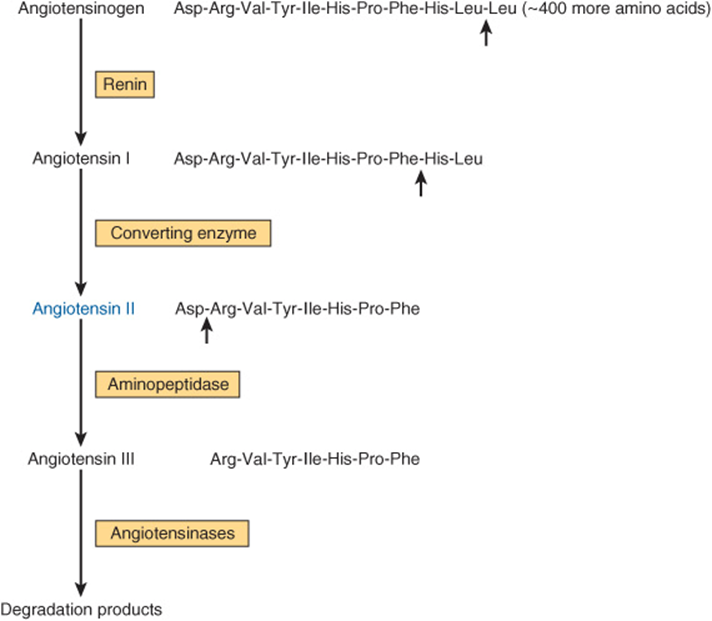
FIGURE 41–14 Formation and metabolism of angiotensins. Small arrows indicate cleavage sites.
Angiotensin-converting enzyme, a glycoprotein found in lung, endothelial cells, and plasma, removes two carboxyl terminal amino acids from the decapeptide angiotensin I to form angiotensin II in a step that is not thought to be rate limiting. Various nonapeptide analogs of angiotensin I and other compounds act as competitive inhibitors of converting enzyme and are used to treat renin-dependent hypertension. These are referred to as angiotensin-converting enzyme (ACE) inhibitors. Angiotensin II increases blood pressure by causing vasoconstriction of the arteriole and is a very potent vasoactive substance. It inhibits renin release from the juxtaglomerular cells and is a potent stimulator of aldosterone production. This results in Na+ retention, volume expansion, and increased blood pressure.
In some species, angiotensin II is converted to the heptapeptide angiotensin III (Figure 41–14), an equally potent stimulator of aldosterone production. In humans, the plasma level of angiotensin II is four times greater than that of angiotensin III, so most effects are exerted by the octapeptide. Angiotensins II and III are rapidly inactivated by angiotensinases.
Angiotensin II binds to specific adrenal cortex glomerulosa cell receptors. The hormone-receptor interaction does not activate adenylyl cyclase, and cAMP does not appear to mediate the action of this hormone. The actions of angiotensin II, which are to stimulate the conversion of cholesterol to pregnenolone and of corticosterone to 18-hydroxycorticosterone and aldosterone, may involve changes in the concentration of intracellular calcium and of phospholipid metabolites by mechanisms similar to those described in Chapter 42.
Complex Processing Generates the Pro-Opiomelanocortin (POMC) Peptide Family
The POMC family consists of peptides that act as hormones (ACTH, LPH, MSH) and others that may serve as neurotransmitters or neuromodulators (endorphins) (Figure 41–15). POMC is synthesized as a precursor molecule of 285 amino acids and is processed differently in various regions of the pituitary.
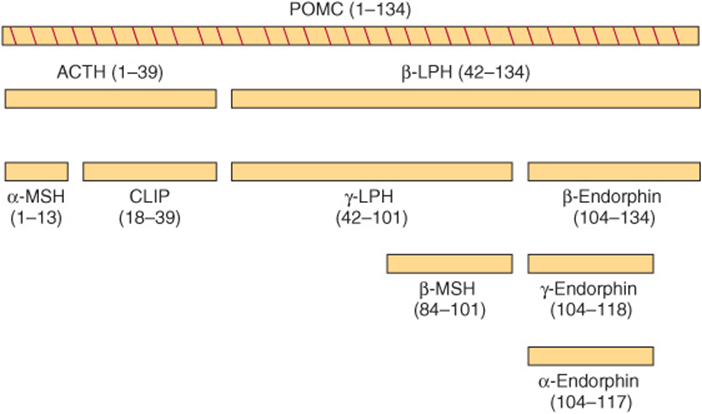
FIGURE 41–15 Products of pro-opiomelanocortin (POMC) cleavage. (CLIP, corticotropin-like intermediate lobe peptide; LPH, lipotropin; MSH, melanocyte-stimulating hormone.)
The POMC gene is expressed in the anterior and intermediate lobes of the pituitary. The most conserved sequences between species are within the amino terminal fragment, the ACTH region, and the β-endorphin region. POMC or related products are found in several other vertebrate tissues, including the brain, placenta, gastrointestinal tract, reproductive tract, lung, and lymphocytes.
The POMC protein is processed differently in the anterior lobe than in the intermediate lobe. The intermediate lobe of the pituitary is rudimentary in adult humans, but it is active in human fetuses and in pregnant women during late gestation and is also active in many animal species. Processing of the POMC protein in the peripheral tissues (gut, placenta, and male reproductive tract) resembles that in the intermediate lobe. There are three basic peptide groups: (1) ACTH, which can give rise to α-MSH and corticotropin-like intermediate lobe peptide (CLIP); (2) β-lipotropin (β-LPH), which can yield γ-LPH, β-MSH, and β-endorphin (and thus α- and γ-endorphins); and (3) a large amino terminal peptide, which generates γ-MSH (not shown). The diversity of these products is due to the many dibasic amino acid clusters that are potential cleavage sites for trypsin-like enzymes. Each of the peptides mentioned is preceded by Lys-Arg, Arg-Lys, Arg-Arg, or Lys-Lys residues. After the prehormone segment is cleaved, the next cleavage, in both anterior and intermediate lobes, is between ACTH and β-LPH, resulting in an amino terminal peptide with ACTH and a β-LPH segment (Figure 41–15). ACTH1-39 is subsequently cleaved from the amino terminal peptide, and in the anterior lobe essentially no further cleavages occur. In the intermediate lobe, ACTH1-39 is cleaved into α-MSH (residues 1-13) and CLIP (18-39); β-LPH (42-134) is converted to γ-LPH (42-101) and β-endorphin (104-134). β-MSH (84-101) is derived from γ-LPH, while γ-MSH (50-74) is derived from a POMC N-terminal fragment (1-74).
There are extensive additional tissue-specific modifications of these peptides that affect activity. These modifications include phosphorylation, acetylation, glycosylation, and amidation.
Mutations of the α-MSH receptor are linked to a common, early-onset form of obesity. This observation has redirected attention to the POMC peptide hormones.
THERE IS VARIATION IN THE STORAGE & SECRETION OF HORMONES
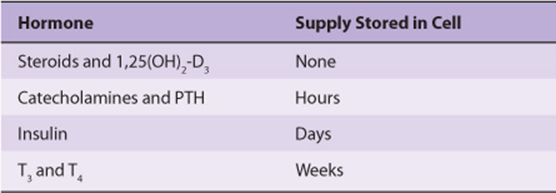
As mentioned above, the steroid hormones and 1,25(OH)2 -D3 are synthesized in their final active form. They are also secreted as they are made, and thus there is no intracellular reservoir of these hormones. The catecholamines, also synthesized in active form, are stored in granules in the chromaffin cells in the adrenal medulla. In response to appropriate neural stimulation, these granules are released from the cell through exocytosis, and the catecholamines are released into the circulation. A several-hour reserve supply of catecholamines exists in the chromaffin cells.
PTH also exists in storage vesicles. As much as 80-90% of the pro PTH synthesized is degraded before it enters this final storage compartment, especially when Ca2+ levels are high in the parathyroid cell (see above). PTH is secreted when Ca2+ is low in the parathyroid cells, which contain a several-hour supply of the hormone.
The human pancreas secretes about 40-50 units of insulin daily; this represents about 15-20% of the hormone stored in the β cells. Insulin and the C-peptide (see Figure 41–12) are normally secreted in equimolar amounts. Stimuli such as glucose, which provokes insulin secretion, therefore trigger the processing of proinsulin to insulin as an essential part of the secretory response.
A several-week supply of T3 and T4 exists in the thyroglobulin that is stored in colloid in the lumen of the thyroid follicles. These hormones can be released upon stimulation by TSH. This is the most exaggerated example of a prohormone, as a molecule containing ~5000 amino acids must be first synthesized, then degraded, to supply a few molecules of the active hormones T3 and T4.
The diversity in storage and secretion of hormones is illustrated in Table 41-5.
TABLE 41–5 Diversity in the Storage of Hormones
SOME HORMONES HAVE PLASMA TRANSPORT PROTEINS
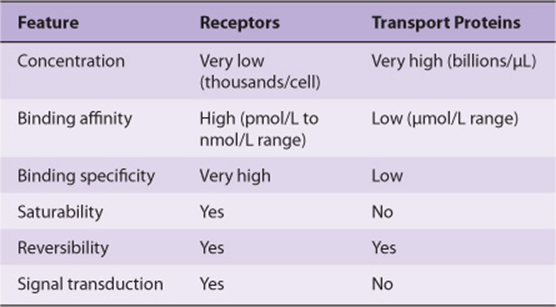
The class I hormones are hydrophobic in chemical nature and thus are not very soluble in plasma. These hormones, principally the steroids and thyroid hormones, have specialized plasma transport proteins that serve several purposes. First, these proteins circumvent the solubility problem and thereby deliver the hormone to the target cell. They also provide a circulating reservoir of the hormone that can be substantial, as in the case of the thyroid hormones. Hormones, when bound to the transport proteins, cannot be metabolized, thereby prolonging their plasma half-life (t½). The binding affinity of a given hormone to its transporter determines the bound versus free ratio of the hormone. This is important because only the free form of a hormone is biologically active. In general, the concentration of free hormone in plasma is very low, in the range of 10-15 to 10-9 mol/L. It is important to distinguish between plasma transport proteins and hormone receptors. Both bind hormones but with very different characteristics (Table 41-6).
TABLE 41–6 Comparison of Receptors with Transport Proteins
The hydrophilic hormones—generally class II and of peptide structure—are freely soluble in plasma and do not require transport proteins. Hormones such as insulin, growth hormone, ACTH, and TSH circulate in the free, active form and have very short plasma half-lives. A notable exception is IGF-I, which is transported bound to members of a family of binding proteins.
Thyroid Hormones Are Transported by Thyroid-Binding Globulin
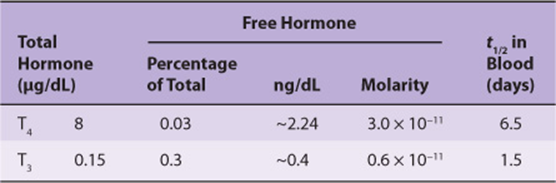
Many of the principles discussed above are illustrated in a discussion of thyroid-binding proteins. One-half to two-thirds of T4 and T3 in the body is in an extrathyroidal reservoir. Most of this circulates in bound form, ie, bound to a specific binding protein, thyroxine-binding globulin (TBG). TBG, a glycoprotein with a molecular mass of 50 kDa, binds T4 and T3and has the capacity to bind 20 μg/dL of plasma. Under normal circumstances, TBG binds—noncovalently—nearly all of the T4 and T3 in plasma, and it binds T4 with greater affinity than T3 (Table 41-7). The plasma half-life of T4 is correspondingly four to five times that of T3. The small, unbound (free) fraction is responsible for the biologic activity. Thus, in spite of the great difference in total amount, the free fraction of T3approximates that of T4, and given that T3 is intrinsically more active than T4, most biologic activity is attributed to T3. TBG does not bind any other hormones.
TABLE 41–7 Comparison of T4 and T3 in Plasma
Glucocorticoids Are Transported by Corticosteroid-Binding Globulin
Hydrocortisone (cortisol) also circulates in plasma in protein-bound and free forms. The main plasma binding protein is an α-globulin called transcortin, or corticosteroid-binding globulin (CBG). CBG is produced in the liver, and its synthesis, like that of TBG, is increased by estrogens. CBG binds most of the hormone when plasma cortisol levels are within the normal range; much smaller amounts of cortisol are bound to albumin. The avidity of binding helps determine the biologic half-lives of various glucocorticoids. Cortisol binds tightly to CBG and has a t½ of 1.5-2 h, while corticosterone, which binds less tightly, has a t½ of <1 h (Table 41-8). The unbound (free) cortisol constitutes ~8% of the total and represents the biologically active fraction. Binding to CBG is not restricted to glucocorticoids. Deoxycorticosterone and progesterone interact with CBG with sufficient affinity to compete for cortisol binding. Aldosterone, the most potent natural mineralo-corticoid, does not have a specific plasma transport protein. Gonadal steroids bind very weakly to CBG (Table 41-8).
TABLE 41–8 Approximate Affinities of Steroids for Serum-Binding Proteins
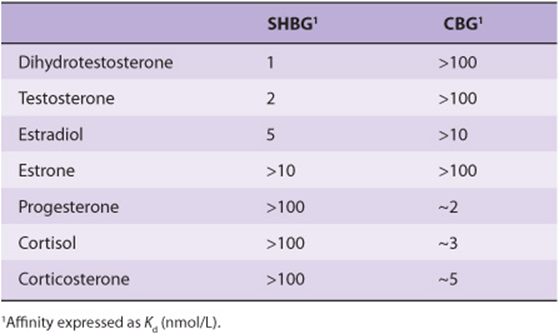
Gonadal Steroids Are Transported by Sex-Hormone-Binding Globulin
Most mammals, humans included, have a plasma β-globulin that binds testosterone with specificity, relatively high affinity, and limited capacity (Table 41-8). This protein, usually called sex-hormone-binding globulin (SHBG) or testosteroneestrogen-binding globulin (TEBG), is produced in the liver. Its production is increased by estrogens (women have twice the serum concentration of SHBG as men), certain types of liver disease, and hyperthyroidism; it is decreased by androgens, advancing age, and hypothyroidism. Many of these conditions also affect the production of CBG and TBG. Since SHBG and albumin bind 97-99% of circulating testosterone, only a small fraction of the hormone in circulation is in the free (biologically active) form. The primary function of SHBG may be to restrict the free concentration of testosterone in the serum. Testosterone binds to SHBG with higher affinity than does estradiol (Table 41-8). Therefore, a change in the level of SHBG causes a greater change in the free testosterone level than in the free estradiol level.
Estrogens are bound to SHBG and progestins to CBG. SHBG binds estradiol about five times less avidly than it binds testosterone or DHT, while progesterone and cortisol have little affinity for this protein (Table 41-8). In contrast, progesterone and cortisol bind with nearly equal affinity to CBG, which in turn has little avidity for estradiol and even less for testosterone, DHT, or estrone.
These binding proteins also provide a circulating reservoir of hormone, and because of the relatively large binding capacity they probably buffer against sudden changes in the plasma level. Because the metabolic clearance rates of these steroids are inversely related to the affinity of their binding to SHBG, estrone is cleared more rapidly than estradiol, which in turn is cleared more rapidly than testosterone or DHT.
SUMMARY
![]() The presence of a specific receptor defines the target cells for a given hormone.
The presence of a specific receptor defines the target cells for a given hormone.
![]() Receptors are proteins that bind specific hormones and generate an intracellular signal (receptor-effector coupling).
Receptors are proteins that bind specific hormones and generate an intracellular signal (receptor-effector coupling).
![]() Some hormones have intracellular receptors; others bind to receptors on the plasma membrane.
Some hormones have intracellular receptors; others bind to receptors on the plasma membrane.
![]() Hormones are synthesized from a number of precursor molecules, including cholesterol, tyrosine per se, and all the constituent amino acids of peptides and proteins.
Hormones are synthesized from a number of precursor molecules, including cholesterol, tyrosine per se, and all the constituent amino acids of peptides and proteins.
![]() A number of modification processes alter the activity of hormones. For example, many hormones are synthesized from larger precursor molecules.
A number of modification processes alter the activity of hormones. For example, many hormones are synthesized from larger precursor molecules.
![]() The complement of enzymes in a particular cell type allows for the production of a specific class of steroid hormone.
The complement of enzymes in a particular cell type allows for the production of a specific class of steroid hormone.
![]() Most of the lipid-soluble hormones are bound to rather specific plasma transport proteins.
Most of the lipid-soluble hormones are bound to rather specific plasma transport proteins.
REFERENCES
Bain DL, Heneghan AF, Connaghan-Jones KD, et al: Nuclear receptor structure: implications for function. Ann Rev Physiol 2007;69:201.
Bartalina L: Thyroid hormone-binding proteins: update 1994. Endocr Rev 1994;13:140.
Beato M, Herrlich P, Schütz G: Steroid hormone receptors: many actors in search of a plot. Cell 1995;83:851.
Cheung E, Kraus WL: Genomic Analyses of Hormone Signaling and Gene Regulation. Annu Rev Physiol 2010;72:191-218.
Cristina Casals-Casas C, Desvergne B: Endocrine Disruptors: From Endocrine to Metabolic Disruption. Annu Rev Physiol 2011;73:23.1-23.28.
Dai G, Carrasco L, Carrasco N: Cloning and characterization of the thyroid iodide transporter. Nature 1996;379:458.
DeLuca HR: The vitamin D story: a collaborative effort of basic science and clinical medicine. FASEB J 1988;2:224.
Douglass J, Civelli O, Herbert E: Polyprotein gene expression: Generation of diversity of neuroendocrine peptides. Annu Rev Biochem 1984;53:665.
Farooqi IS, O’Rahilly S: Monogenic obesity in humans. Ann Rev Med 2005;56:443.
Miller WL: Molecular biology of steroid hormone biosynthesis. Endocr Rev 1988;9:295.
Nagatsu T: Genes for human catecholamine-synthesizing enzymes. Neurosci Res 1991;12:315.
Russell DW, Wilson JD: Steroid 5 alpha-reductase: two genes/two enzymes. Annu Rev Biochem 1994;63:25.
Russell J, Bar A, Sherwood LM, et al: Interaction between calcium and 1,25-dihydroxyvitamin D3 in the regulation of preproparathyroid hormone and vitamin D receptor mRNA in avian parathyroids. Endocrinology 1993;132:2639.
Steiner DF, Smeekens SP, Ohagi S, et al: The new enzymology of precursor processing endoproteases. J Biol Chem 1992;267:23435.
Taguchi A, White M: Insulin-like signaling, nutrient homeostasis, and life span. Ann Rev Physiol 2008;70:191.