Harper’s Illustrated Biochemistry, 29th Edition (2012)
SECTION VI. Special Topics
Chapter 47. Glycoproteins
Robert K. Murray, MD, PhD
OBJECTIVES
After studying this chapter, you should be able to:
![]() Have a general appreciation of the importance of glycobiology and glycomics, and in particular of glycoproteins, in health and disease.
Have a general appreciation of the importance of glycobiology and glycomics, and in particular of glycoproteins, in health and disease.
![]() Know the principal sugars found in glycoproteins.
Know the principal sugars found in glycoproteins.
![]() Be aware of the several major classes of glycoproteins (N-linked, O-linked, and GPI-linked).
Be aware of the several major classes of glycoproteins (N-linked, O-linked, and GPI-linked).
![]() Understand the major features of the pathways of biosynthesis and degradation of O- and N-linked glycoproteins.
Understand the major features of the pathways of biosynthesis and degradation of O- and N-linked glycoproteins.
![]() Understand the importance of advanced glycation end-products in causing tissue damage in diabetes mellitus.
Understand the importance of advanced glycation end-products in causing tissue damage in diabetes mellitus.
![]() Be able to indicate the involvement of glycoproteins in inflammation and in a host of conditions including I-cell disease, congenital disorders of glycation, paroxysmal nocturnal hemoglobinuria and cancer.
Be able to indicate the involvement of glycoproteins in inflammation and in a host of conditions including I-cell disease, congenital disorders of glycation, paroxysmal nocturnal hemoglobinuria and cancer.
![]() Be familiar with the concept that many microorganisms, such as influenza virus, attach to cell surfaces via sugar chains.
Be familiar with the concept that many microorganisms, such as influenza virus, attach to cell surfaces via sugar chains.
BIOMEDICAL IMPORTANCE
Glycobiology is the study of the roles of sugars in health and disease. The glycome is the entire complement of sugars, whether free or present in more complex molecules, of an organism. Glycomics, an analogous term to genomics and proteomics, is the comprehensive study of glycomes, including genetic, physiologic, pathologic, and other aspects.
One major class of molecules included in the glycome is glycoproteins. These are proteins that contain oligosaccharide chains (glycans) covalently attached to their polypeptide backbones. It has been estimated that approximately 50% of eukaryotic proteins have sugars attached, so that glycosylation (enzymic attachment of sugars) is the most frequent posttranslational modification of proteins. Nonenzymic attachment of sugars to proteins can also occur, and is referred to as glycation. This process can have serious pathologic consequences (eg, in poorly controlled diabetes mellitus). Glycoproteins are one class of glycoconjugate or complex carbohydrate—equivalent terms used to denote molecules containing one or more carbohydrate chains covalently linked to protein (to form glycoproteins or proteoglycans) or lipid (to form glycolipids). (Proteoglycans are discussed in Chapter 48 and glycolipids in Chapter 15.) Almost all the plasma proteins of humans—with the notable exception of albumin—are glycoproteins. Many proteins of cellular membranes (Chapter 40) contain substantial amounts of carbohydrate. A number of the blood group substances are glycoproteins, whereas others are glycosphingolipids. Certain hormones (eg, chorionic gonadotropin) are glycoproteins. A major problem in cancer is metastasis, the phenomenon whereby cancer cells leave their tissue of origin (eg, the breast), migrate through the bloodstream to some distant site in the body (eg, the brain), and grow there in an unregulated manner, with catastrophic results for the affected individual. Many cancer researchers think that alterations in the structures of glycoproteins and other glycoconjugates on the surfaces of cancer cells are important in the phenomenon of metastasis.
GLYCOPROTEINS OCCUR WIDELY & PERFORM NUMEROUS FUNCTIONS
Glycoproteins occur in most organisms, from bacteria to humans. Many viruses also contain glycoproteins, some of which have been much investigated, in part because they often play key roles in viral attachment to cells (eg, HIV-1 and influenza A virus). Numerous proteins with diverse functions are glycoproteins (Table 47-1); their carbohydrate content ranges from 1% to over 85% by weight.
TABLE 47–1 Some Functions Served by Glycoproteins
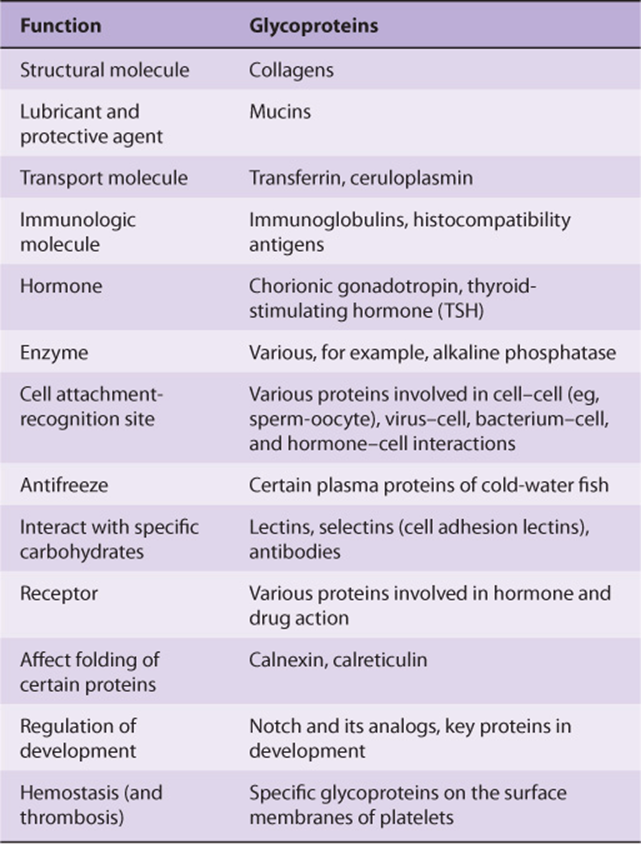
Many studies have been conducted in an attempt to define the precise roles oligosaccharide chains play in the functions of glycoproteins. Table 47-2 summarizes results from such studies. Some of the functions listed are firmly established; others are still under investigation.
TABLE 47–2 Some Functions of the Oligosaccharide Chains of Glycoproteins

OLIGOSACCHARIDE CHAINS ENCODE BIOLOGIC INFORMATION
An enormous number of glycosidic linkages can be generated between sugars. For example, three different hexoses may be linked to each other to form over 1000 different trisaccharides. The conformations of the sugars in oligosaccharide chains vary depending on their linkages and proximity to other molecules with which the oligosaccharides may interact. It is now established that certain oligosaccharide chains encode biologic information and that this depends upon their constituent sugars, their sequences, and their linkages. For instance, man-nose 6-phosphate residues target newly synthesized lysosomal enzymes to that organelle (see later). The biologic information that sugars contain is expressed via interactions between specific sugars, either free or in glycoconjugates, and proteins (such as lectins; see below) or other molecules. These interactions lead to changes of the cellular activity. Thus, deciphering the so-called sugar code of life (one of the principal aims of glycomics) entails elucidating all of the interactions that sugars and sugar-containing molecules participate in, and also the results of these interactions on cellular behavior. This will not be an easy task, considering the diversity of glycans found in cells.
TECHNIQUES ARE AVAILABLE FOR DETECTION, PURIFICATION, STRUCTURAL ANALYSIS & SYNTHESIS OF GLYCOPROTEINS
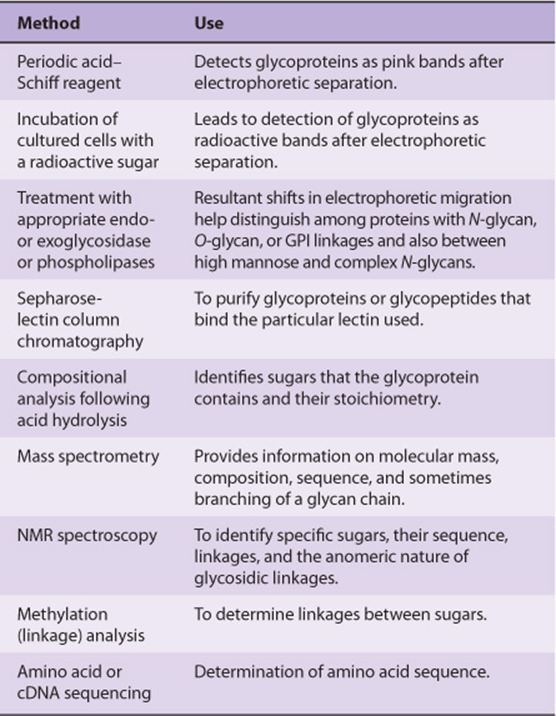
A variety of methods used in the detection, purification, and structural analysis of glycoproteins are listed in Table 47-3. The conventional methods used to purify proteins and enzymes are also applicable to the purification of glycoproteins. Once a glycoprotein has been purified, the use of mass spectrometry and high-resolution NMR spectroscopy can often identify the structures of its glycan chains. Analysis of glycoproteins can be complicated by the fact that they often exist as glycoforms; these are proteins with identical amino acid sequences but somewhat different oligosaccharide compositions. Although linkage details are not stressed in this chapter, it is critical to appreciate that the precise natures of the linkages between the sugars of glycoproteins are of fundamental importance in determining the structures and functions of these molecules.
TABLE 47–3 Some Important Methods Used to Study Glycoproteins
Impressive advances are also being made in synthetic chemistry, allowing synthesis of complex glycans that can be tested for the biologic and pharmacologic activity. In addition, methods have been developed that use simple organisms, such as yeasts, to secrete human glycoproteins of therapeutic value (eg, erythropoietin) into their surrounding medium.
EIGHT SUGARS PREDOMINATE IN HUMAN GLYCOPROTEINS
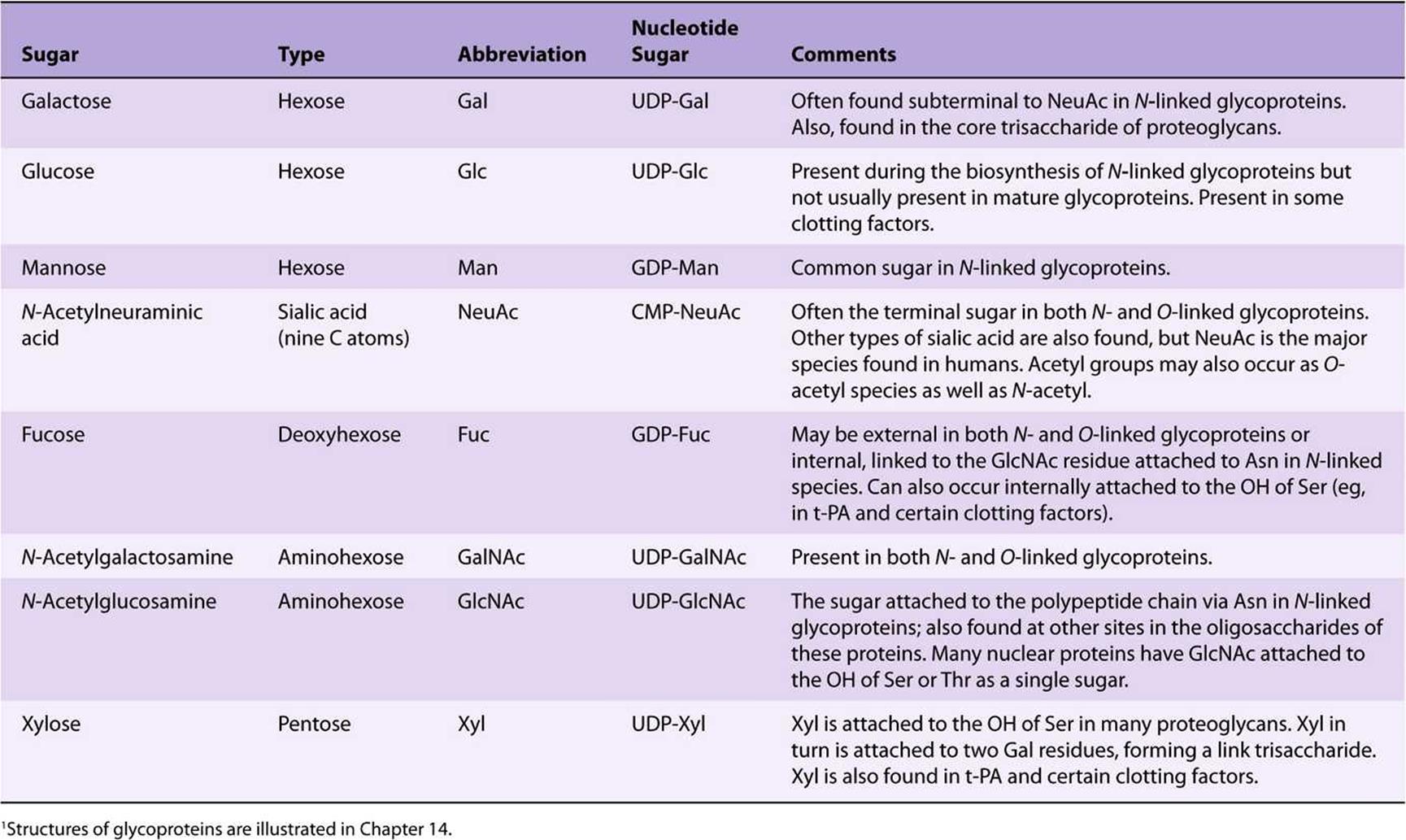
About 200 monosaccharides are found in nature; however, only eight are commonly found in the oligosaccharide chains of glycoproteins (Table 47-4). Most of these sugars were described in Chapter 14. N-acetylneuraminic acid (NeuAc) is usually found at the termini of oligosaccharide chains, attached to subterminal galactose (Gal) or N-acetylgalactosamine (Gal-NAc) residues. The other sugars listed are generally found in more internal positions. Sulfateis often found in glycoproteins, usually attached to Gal, GalNAc, or GlcNAc.
TABLE 47–4 The Principal Sugars Found in Human Glycoproteins1
NUCLEOTIDE SUGARS ACT AS SUGAR DONORS IN MANY BIOSYNTHETIC REACTIONS
It is important to understand that in most biosynthetic reactions, it is not the free sugar or phosphorylated sugar that is involved in such reactions, but rather the corresponding nucleotide sugar. The first nucleotide sugar to be reported was uridine diphosphate glucose (UDP-Glc); its structure is shown in Figure 19–2. The common nucleotide sugars involved in the biosynthesis of glycoproteins are listed in Table 47-4; the reasons some contain UDP and others guanosine diphosphate (GDP) or cytidine monophosphate (CMP) are not clear. Many of the glycosylation reactions are involved in the biosynthesis of glycoproteins utilize these compounds (see below). The anhydro natureof the linkage between the phosphate group and the sugars is of the high energy, high-group-transfer-potential type (Chapter 11). The sugars of these compounds are thus “activated” and can be transferred to suitable acceptors provided appropriate transferases are available.
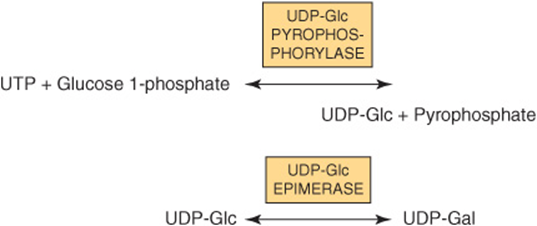
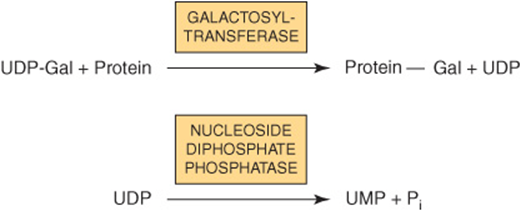
Most nucleotide sugars are formed in the cytosol, generally from reactions involving the corresponding nucleoside triphosphate. CMP-sialic acids are formed in the nucleus. Formation of uridine diphosphate galactose (UDP-Gal) requires the following two reactions in mammalian tissues.
Because many glycosylation reactions occur within the lumen of the Golgi apparatus, carrier systems (permeases, transporters) are necessary to transport nucleotide sugars across the Golgi membrane. Systems transporting UDP-Gal, GDP-Man, and CMP-NeuAc into the cisternae of the Golgi apparatus have been described. They are antiport systems; that is, the influx of one molecule of nucleotide sugar is balanced by the efflux of one molecule of the corresponding nucleotide (eg, UMP, GMP, or CMP) formed from the nucleotide sugars. This mechanism ensures an adequate concentration of each nucleotide sugar inside the Golgi apparatus. UMP is formed from UDP-Gal in the above process as follows.
EXO- & ENDOGLYCOSIDASES FACILITATE STUDY OF GLYCOPROTEINS
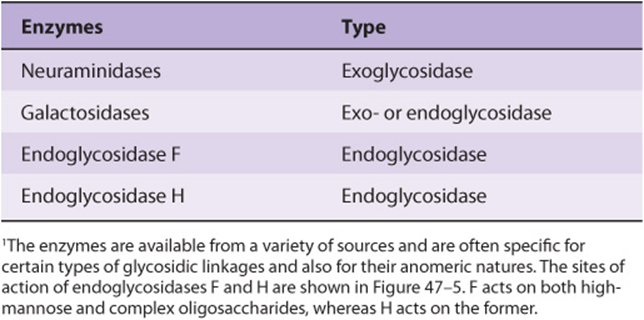
A number of glycosidases of defined specificity have proved useful in examining structural and functional aspects of glycoproteins (Table 47-5). These enzymes act at either external (exoglycosidases) or internal (endoglycosidases) positions of oligosaccharide chains. Examples of exoglycosidases are neuraminidases and galactosidases; their sequential use removes terminal NeuAc and subterminal Gal residues from most glycoproteins. Endoglycosidases Fand H are examples of the latter class; these enzymes cleave the oligosaccharide chains at specific GlcNAc residues close to the polypeptide backbone (ie, at internal sites; Figure 47–5) and are thus useful in releasing large oligosaccharide chains for structural analyses. A glycoprotein can be treated with one or more of the above glycosidases to analyze the effects on its biologic behavior of removal of specific sugars.
TABLE 47–5 Some Glycosidases Used to Study the Structure and Function of Glycoproteins1
THE MAMMALIAN ASIALOGLYCO PROTEIN RECEPTOR IS INVOLVED IN CLEARANCE OF CERTAIN GLYCOPROTEINS FROM PLASMA BY HEPATOCYTES
Experiments performed by Ashwell and his colleagues in the early 1970s played an important role in focusing attention on the functional significance of the oligosaccharide chains of glycoproteins. They treated rabbit ceruloplasmin (a plasma protein; see Chapter 50) with neuraminidase in vitro. This procedure exposed subterminal Gal residues that were normally masked by terminal NeuAc residues. Neuraminidase-treated radioactive ceruloplasmin was found to disappear rapidly from the circulation, in contrast to the slow clearance of the untreated protein. Very significantly, when the Gal residues exposed to treatment with neuraminidase were removed by treatment with a galactosidase, the clearance rate of the protein returned to normal. Further studies demonstrated that liver cells contain a mammalian asialoglycoprotein receptor that recognizes the Gal moiety of many desialylated plasma proteins and leads to their endocytosis. This work indicated that an individual sugar, such as Gal, could play an important role in governing at least one of the biologic properties (ie, time of residence in the circulation) of certain glycoproteins. This greatly strengthened the concept that oligosaccharide chains could contain biologic information.
LECTINS CAN BE USED TO PURIFY GLYCOPROTEINS & TO PROBE THEIR FUNCTIONS
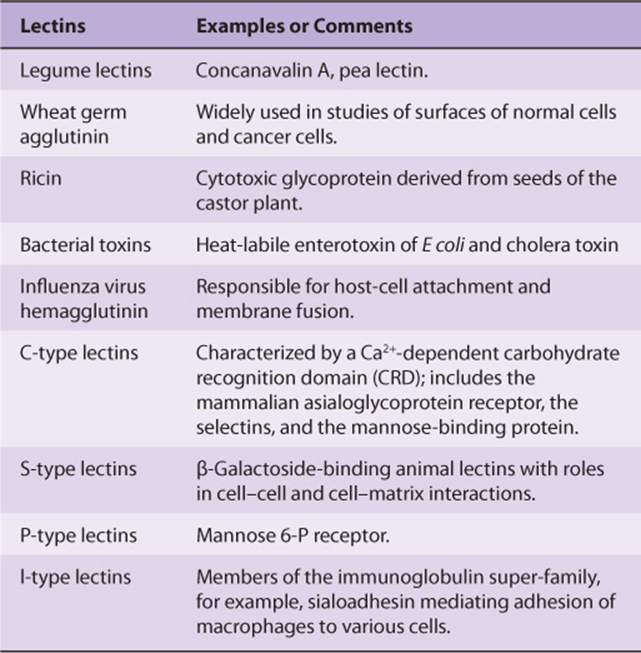
Lectins are carbohydrate-binding proteins that agglutinate cells or precipitate glycoconjugates; a number of lectins are themselves glycoproteins. Immunoglobulins that react with sugars are not considered lectins. Lectins contain at least two sugar-binding sites; proteins with a single sugar-binding site will not agglutinate cells or precipitate glycoconjugates. The specificity of a lectin is usually defined by the sugars that are best at inhibiting its ability to cause agglutination or precipitation. Enzymes, toxins, and transport proteins can be classified as lectins if they bind carbohydrate. Lectins were first discovered in plants and microbes, but many lectins of animal origin are now known. The mammalian asialoglyco-protein receptor described above is an important example of an animal lectin. Some important lectins are listed in Table 47-6. Much current research is centered on the roles of various animal lectins in the mechanisms of action of glycoproteins, some of which are discussed below (eg, with regard to the selectins).
TABLE 47–6 Some Important Lectins
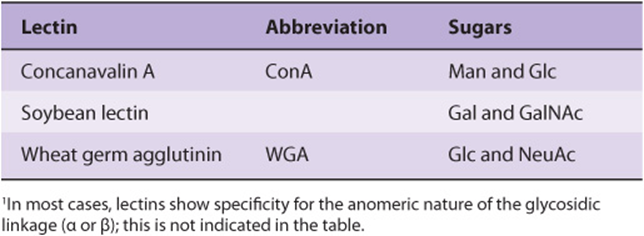
Numerous lectins have been purified and are commercially available; three plant lectins that have been widely used experimentally are listed in Table 47-7. Among many uses, lectins have been employed to purify specific glycoproteins, as tools for probing the glycoprotein profiles of cell surfaces, and as reagents for generating mutant cells deficient in certain enzymes involved in the biosynthesis of oligosaccharide chains.
TABLE 47–7 Three Plant Lectins and the Sugars with Which They Interact1
THERE ARE THREE MAJOR CLASSES OF GLYCOPROTEINS
Based on the nature of the linkage between their polypeptide chains and their oligosaccharide chains, glycoproteins can be divided into three major classes (Figure 47–1). (1) Those containing an O-glycosidic linkage (ie, O-linked), involving the hydroxyl side chain of serine or threonine and a sugar such as N-acetylgalactosamine (GalNAc-Ser[Thr]); (2) those containing an JV-gycosidic linkage (ie, N-linked), involving the amide nitrogen ofasparagine and N-acetylglucosamine(GlcNAc-Asn); and (3) those linked to the carboxyl terminal amino acid of a protein via a phosphoryl-ethanolamine moiety joined to an oligosaccharide (glycan), which in turn is linked via glucosamine to phosphatidylinositol (PI). This latter class is referred to as glycosylphosphatidylinositol-anchored (GPI-anchored, or GPI-linked) glycoproteins. Members of this class, among other functions, are involved in directing certain glycoproteins to the apical or basolateral areas of the plasma membrane (PM) of some polarized epithelial cells (see Chapter 40 and below). Other minor classes of glycoproteins also exist.
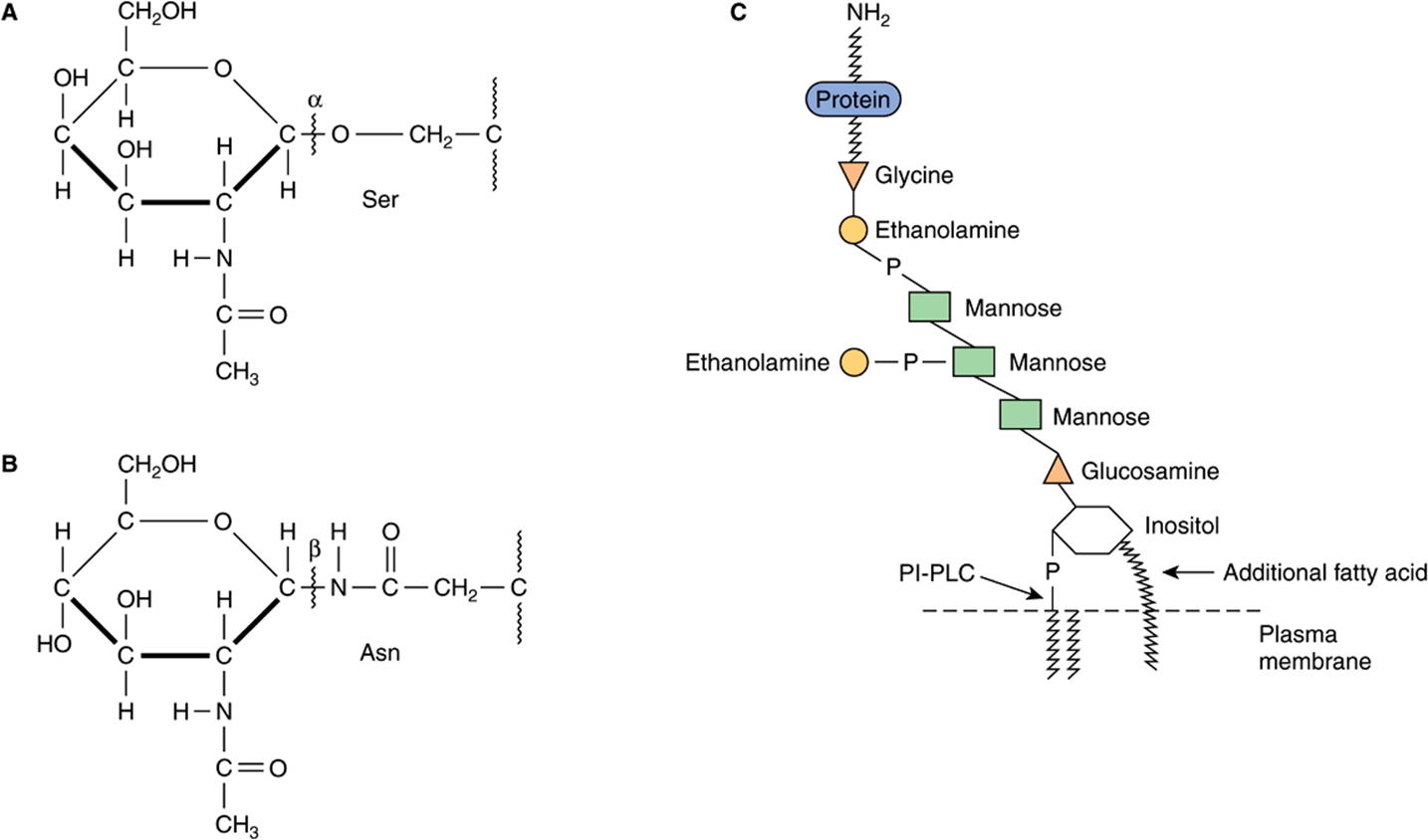
FIGURE 47–1 Depictions of (A) an O-linkage(N-acetylgalactosamine to serine), (B) an N-linkage (N-acetylglucosamine to asparagine), and (C) a glycosylphosphatidylinositol (GPI) linkage. The GPI structure shown is that linking acetylcholinesterase to the plasma membrane of the human red blood cell. The carboxyl terminal amino acid is glycine joined in amide linkage via its COOH group to the NH2 group of phosphorylethanolamine, which in turn is joined to a mannose residue. The core glycan contains three mannose and one glucosamine residues. The glucosamine is linked to inositol, which is attached to the phosphatidic acid. The site of action of PI-phospholipase C (PI-PLC) is indicated. The structure of the core glycan is shown in the text. This particular GPI contains an extra fatty acid attached to inositol and also an extra phosphorylethanolamine moiety attached to the middle of the three mannose residues. Variations found among different GPI structures include the identity of the carboxyl terminal amino acid, the molecules attached to the mannose residues, and the precise nature of the lipid moiety.
The number of oligosaccharide chains attached to one protein can vary from one to 30 or more, with the sugar chains ranging from one or two residues in length to much larger structures. Many proteins contain more than one type of sugar chain; for instance, glycophorin, an important red cell membrane glycoprotein (Chapter 52), contains both O- and N-linked oligosaccharides.
GLYCOPROTEINS CONTAIN SEVERAL TYPES OF O-GLYCOSIDIC LINKAGES
At least four subclasses of O- glycosidic linkages are found in human glycoproteins. (1) The GalNAc-Ser(Thr) linkage shown in Figure 47–1 is the predominant linkage. Two typical oligosaccharide chains found in members of this subclass are shown in Figure 47–2. Usually a Gal or a NeuAc residue is attached to the GalNAc, but many variations in the sugar compositions and lengths of such oligosaccharide chains are found. This type of linkage is found in mucins (see below). (2) Proteoglycans contain a Gal-Gal-Xyl-Ser trisaccharide (the so-called link trisaccharide). (3) Collagens contain a Gal-Hydroxylysine (Hyl) linkage. (Subclasses [2] and [3] are discussed further in Chapter 48.) (4) Many nuclear proteins (eg, certain transcription factors) and cytosolic proteins contain side chains consisting of a single GlcNAc attached to a serine or threonine residue (GlcNAc-Ser[Thr]).
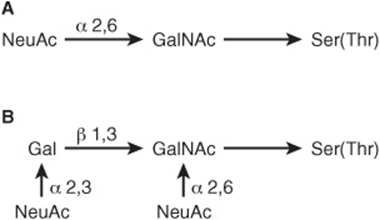
FIGURE 47–2 Structures of two O-linked oligosaccharides found in (A) submaxillary mucins and (B) fetuin and in the sialoglycoprotein of the membrane of human red blood cells. (Modified and reproduced, with permission, from Lennarz WJ: The Biochemistry of Glycoproteins and Proteoglycans. Plenum Press, 1980. Reproduced with kind permission from Springer Science and Business Media.)
Mucins Have a High Content of O-Linked Oligosaccharides & Exhibit Repeating Amino Acid Sequences
Mucins are glycoproteins with two major characteristics: (1) a high content of O-linked oligosaccharides (the carbohydrate content of mucins is generally more than 50%); and (2) the presence of variable numbers of tandem repeats (VNTRs) of peptide sequence in the centre of their polypeptide backbones, to which the O-glycan chains are attached in clusters (Figure 47–3). These sequences are rich in serine, threonine, and proline. Although O-glycans predominate, mucins often contain a number of N-glycan chains. Both secretory and membrane-bound mucins occur. The former are found in the mucus present in the secretions of the gastrointestinal, respiratory, and reproductive tracts. Mucus consists of about 94% water and 5% mucins, with the remainder being a mixture of various cell molecules, electrolytes, and remnants of cells. Secretory mucins generally have an oligomeric structure and thus often have a very high molecular mass. The oligomers are composed of monomers linked by disulfide bonds. Mucus exhibits a high viscosity and often forms a gel. These qualities are functions of its content of mucins. The high content of O-glycans confers an extended structure on mucins. This is in part explained by steric interactions between their GalNAc moieties and adjacent amino acids, resulting in a chain-stiffening effect so that the conformations of mucins often become those of rigid rods. Intermolecular noncovalent interactions between various sugars on neighboring glycan chains contribute to gel formation. The high content of NeuAc and sulfate residues found in many mucins confers a negative charge on them. With regard to their functions, mucins help lubricate and form a protective physical barrier on epithelial surfaces. Membrane-bound mucins participate in various cell-cell interactions (eg, involving selectins; see below). The density of oligosaccharide chains makes it difficult for proteases to approach their polypeptide backbones, so that mucins are often resistant to their action. Mucins also tend to “mask” certain surface antigens. Many cancer cells form excessive amounts of mucins; perhaps the mucins may mask certain surface antigens on such cells and thus protect the cells from immune surveillance. Mucins also carry cancer-specific peptide and carbohydrate epitopes (an epitope is a site on an antigen recognized by an antibody, also called an antigenic determinant). Some of these epitopes have been used to stimulate an immune response against cancer cells.
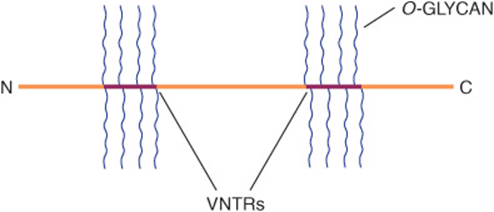
FIGURE 47–3 Much simplified schematic of a mucin. O-glycans (blue) are shown attached to two of many VNTR regions (red). N-glycans may also be present. Mucins generally contain cysteines (not shown) near their N and C termini, which are involved in polymerization via disulfide bridges. Other domains (D) near their N-termini are also involved in polymerization. Membrane-bound mucins contain transmembrane and cytosolic domains, in addition to larger extracellular domains containing O-glycans.
The genes encoding the polypeptide backbones of a number of mucins derived from various tissues (eg, pancreas, small intestine, trachea and bronchi, stomach, and salivary glands) have been cloned and sequenced. These studies have revealed new information about the polypeptide backbones of mucins (size of tandem repeats, potential sites of N-glycosylation, etc) and ultimately should reveal aspects of their genetic control. Some important properties of mucins are summarized in Table 47-8.
TABLE 47–8 SomePropertiesofMucins

The Biosynthesis of O-Linked Glycoproteins Uses Nucleotide Sugars
The polypeptide chains of O-linked and other glycoproteins are encoded by mRNA species; because most glycoproteins are membrane-bound or secreted, they are generally translated on membrane-bound polyribosomes (Chapter 37). Hundreds of different oligosaccharide chains of the O-glycosidic-type exist. These glycoproteins are built up by the stepwise donation of sugars from nucleotide sugars, such as UDP-GalNAc, UDP-Gal, and CMP-NeuAc. The enzymes catalyzing this type of reaction are membrane-bound glycoprotein glycosyltransferases. Generally, synthesis of one specific type of linkage requires the activity of a correspondingly specific transferase. The factors that determine which specific serine and threonine residues are glycosylated have not been identified but are probably found in the peptide structure surrounding the glycosylation site. The enzymes assembling O-linked chains are located in the Golgi apparatus, sequentially arranged in an assembly line with terminal reactions occurring in the trans-Golgi compartments.
The major features of the biosynthesis of O-linked glycoproteins are summarized in Table 47-9.
TABLE 47–9 Summary of Main Features of O-Glycosylation

N-LINKED GLYCOPROTEINS CONTAIN AN Asn-GLcNAc LINKAGE
N-Linked glycoproteins are distinguished by the presence of the Asn-GlcNAc linkage (Figure 47–1). It is the major class of glycoproteins and has been much studied, since the most readily accessible glycoproteins (eg, plasma proteins) mainly belong to this group. It includes both membrane-bound and circulating glycoproteins. The principal difference between this and the previous class, apart from the nature of the amino acid to which the oligosaccharide chain is attached (Asn vs Ser or Thr), concerns their biosynthesis.
Complex, Hybrid & High-Mannose Are the Three Major Classes of N-Linked Oligosaccharides
There are three major classes of N-linked oligosaccharides: complex, hybrid, and high-mannose (Figure 47–4). Each type shares a common pentasaccharide, Man3GlcNAc2—shown within the boxed area in Figure 47–4 and depicted also in Figure 47–5—but they differ in their outer branches. The presence of the common pentasaccharide is explained by the fact that all three classes share an initial common mechanism of biosynthesis. Glycoproteins of the complex type generally contain terminal NeuAc residues and underlying Gal and GlcNAc residues, the latter often constituting the disaccharide N-acetyllactosamine. Repeating N-acetyllactosamine units— [Galβ1-3/4GlcNAcβ1-3]n (poly-N-acetyllactosaminoglycans)—are often found on N-linked glycan chains. I/i blood group substances belong to this class. The majority of complex-type oligosaccharides contain two, three, or four outer branches (Figure 47–4), but structures containing five branches have also been described. The oligosaccharide branches are often referred to as antennae, so that bi-, tri-, tetra-, and penta-antennary structures may all be found. A bewildering number of chains of the complex type exist, and that indicated in Figure 47–4 is only one of many. Other complex chains may terminate in Gal or Fuc. High-mannose oligosaccharides typically have two to six additional Man residues linked to the pentasaccharide core. Hybrid molecules contain features of both of the two other classes.
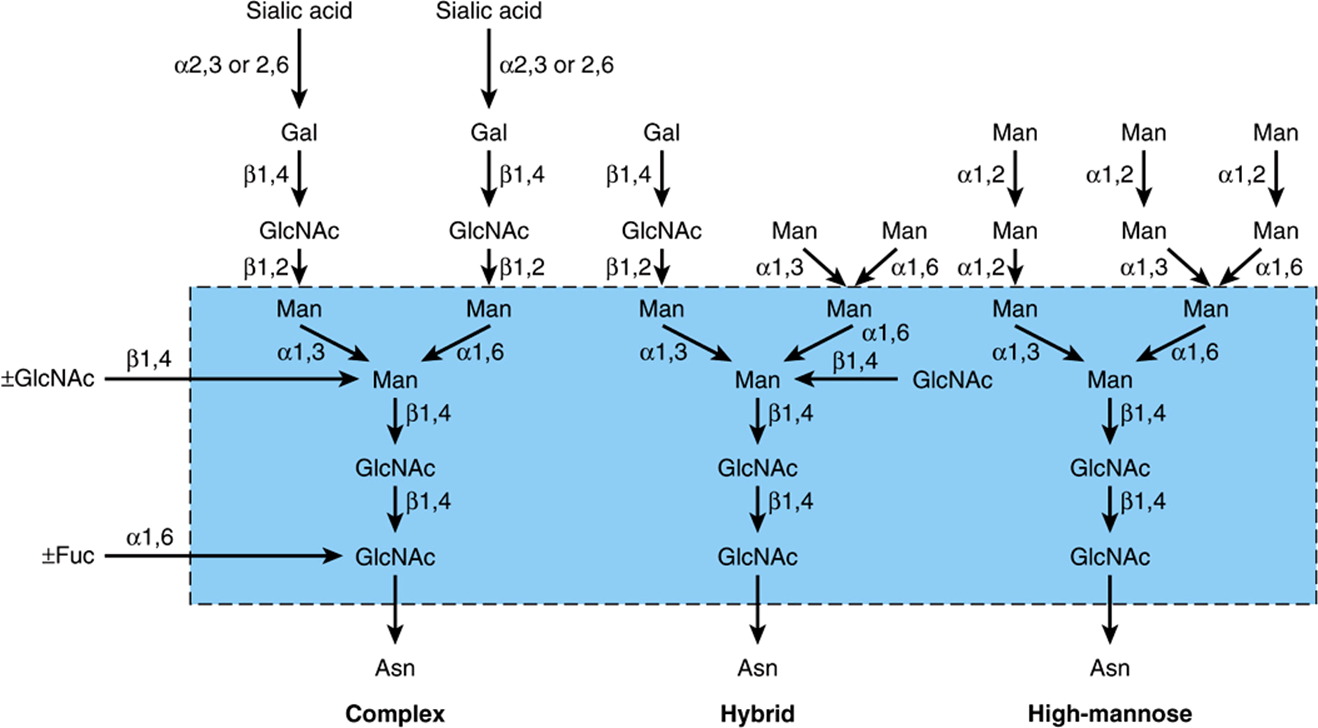
FIGURE 47–4 Structures of the major types of asparagine-linked oligosaccharides. The boxed area encloses the pentasaccharide core common to all N-linked glycoproteins. (Reproduced, with permission, from Kornfeld R, Kornfeld S: Assembly of asparagine-linked oligosaccharides. Annu Rev Biochem 1985;54:631. Copyright © 1985 by Annual Reviews. Reprinted with permission.)
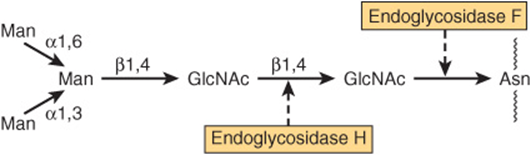
FIGURE 47–5 Schematic of the pentasaccharide core common to all N-linked glycoproteins and to which various outer chains of oligosaccharides may be attached. The sites of action of endoglycosidases F and H are also indicated.
The Biosynthesis of N-Linked Glycoproteins Involves Dolichol-P-P-Oligosaccharide
Leloir and his colleagues described the occurrence of a dolicholpyrophosphate-oligosaccharide (Dol-P-P-oligosaccharide), which subsequent research showed to play a key role in the biosynthesis of N-linked glycoproteins. The oligosaccharide chain of this compound generally has the structure R-Glc-NAc2Man9Glc3 ![]() . The sugars of this compound are first assembled on the Dol-P-P backbone, and the oligosaccharide chain is then transferred en bloc to suitable Asn residues of acceptor apoglycoproteins during their synthesis on membrane-bound polyribosomes. All N--glycans have a common pentasaccharide core structure (Figure 47–5).
. The sugars of this compound are first assembled on the Dol-P-P backbone, and the oligosaccharide chain is then transferred en bloc to suitable Asn residues of acceptor apoglycoproteins during their synthesis on membrane-bound polyribosomes. All N--glycans have a common pentasaccharide core structure (Figure 47–5).
To form high-mannose chains, only the Glc residues plus certain of the peripheral Man residues are removed. To form an oligosaccharide chain of the complex type, the Glc residues and four of the Man residues are removed by glycosidases in the endoplasmic reticulum and Golgi. The sugars characteristic of complex chains (GlcNAc, Gal, and NeuAc) are added by the action of individual glycosyltransferases located in the Golgi apparatus. The phenomenon whereby the glycan chains of N-linked glycoproteins are first partially degraded and then in some cases rebuilt is referred to as oligosaccharide processing. Hybrid chains are formed by partial processing, forming complex chains on one arm and Man structures on the other arm.
Thus, the initial steps involved in the biosynthesis of the N-linked glycoproteins differ markedly from those involved in the biosynthesis of the O-linked glycoproteins. The former involves Dol-P-P-oligosaccharide; the latter, as described earlier, does not.
The process of N-glycosylation can be broken down into two stages: (1) assembly of Dol-P-P-oligosaccharide and transfer of the oligosaccharide; and (2) processing of the oligosaccharide chain.
Assembly & Transfer of Dolichol-P-P-Oligosaccharide
Polyisoprenol compounds exist in both bacteria and eukaryotic cells. They participate in the synthesis of bacterial polysaccharides and in the biosynthesis of N-linked glycoproteins and GPI anchors. The polyisoprenol used in eukaryotic tissues is dolichol, which is, next to rubber, the longest naturally occurring hydrocarbon made up of a single repeating unit. Dolichol is composed of 17-20 repeating isoprenoid units (Figure 47–6).
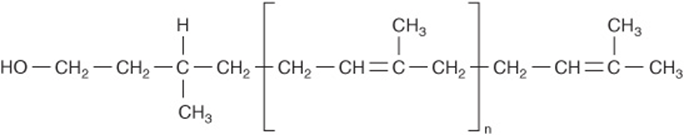
FIGURE 47–6 The structure of dolichol. The phosphate in dolichol phosphate is attached to the primary alcohol group at the left-hand end of the molecule. The group within the brackets is an isoprene unit (![]() isoprenoid units).
isoprenoid units).
Before it participates in the biosynthesis of Dol-P-P-oligosaccharide, dolichol must first be phosphorylated to form dolichol phosphate (Dol-P) in a reaction catalyzed by dolichol kinase and using ATP as the phosphate donor.
Dolichol-P-P-GlcNAc (Dol-P-P-GlcNAc) is the key lipid that acts as an acceptor for other sugars in the assembly of Dol-P-P-oligosaccharide. It is synthesized in the membranes of the endoplasmic reticulum from Dol-P and UDP-GlcNAc in the following reaction, catalyzed by GlcNAc-P transferase.
![]()
The above reaction, which is the first step in the assembly of Dol-P-P-oligosaccharide, and the other later reactions are summarized in Figure 47–7. The essential features of the subsequent steps in the assembly of Dol-P-P-oligosaccharide are as follows:
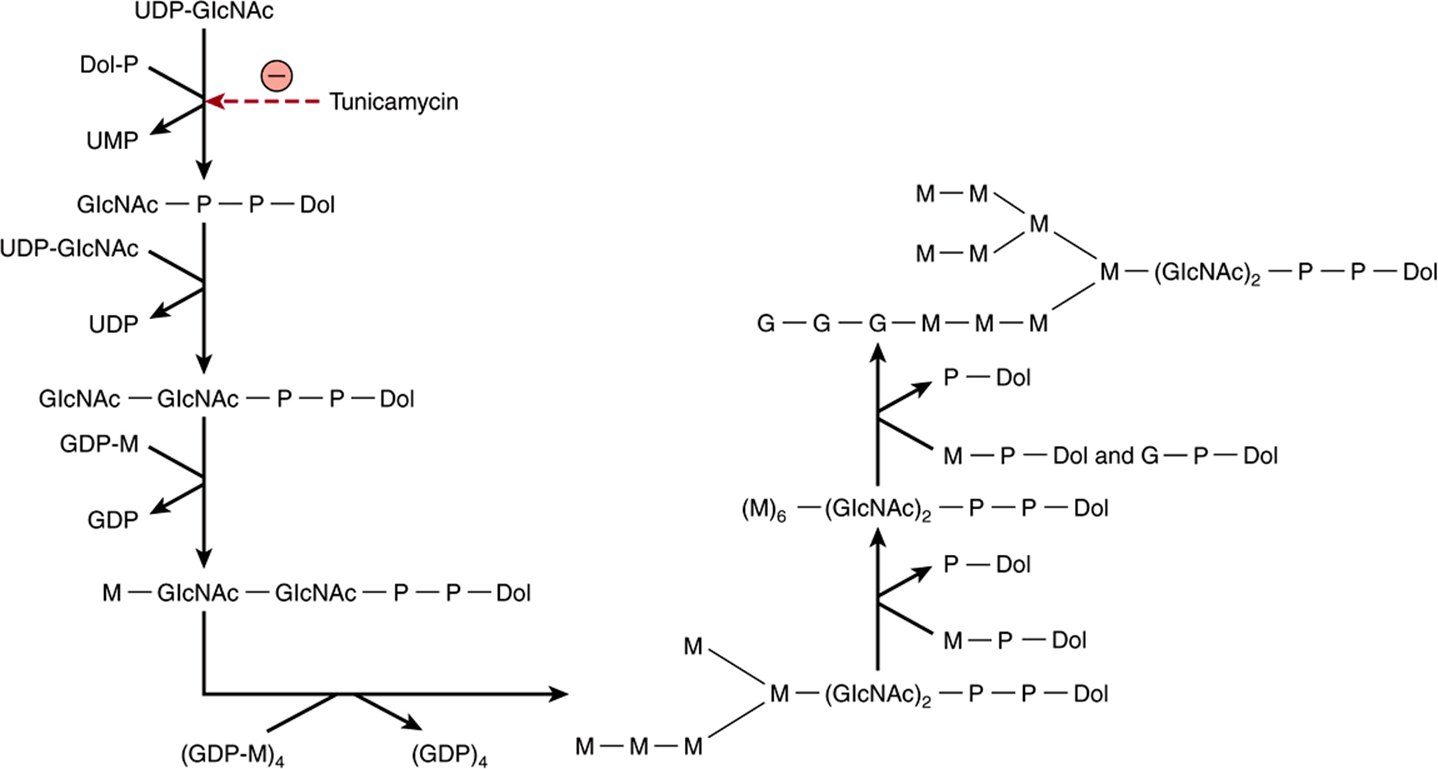
FIGURE 47–7 Pathway of biosynthesis of dolichol-P-P-oligosaccharide. The specific linkages formed are indicated in Figure 47–8. Note that the first five internal mannose residues are donated by GDP-mannose, whereas the more external mannose residues and the glucose residues are donated by dolichol-P-mannose and dolichol-P-glucose. (UDP, uridine diphosphate; Dol, dolichol; P, phosphate; UMP, uridine monophosphate; GDP, guanosine diphosphate.
1. A second GlcNAc residue is added to the first, again using UDP-GlcNAc as the donor.
2. Five Man residues are added, using GDP-mannose as the donor.
3. Four additional Man residues are next added, using Dol-P-Man as the donor. Dol-P-Man is formed by the following reaction.
![]()
4. Finally, the three peripheral glucose residues are donated by Dol-P-Glc, which is formed in a reaction analogous to that just presented except that Dol-P and UDP-Glc are the substrates.
It should be noted that the first seven sugars (two GlcNAc and five Man residues) are donated by nucleotide sugars, whereas the last seven sugars (four Man and three Glc residues) added are donated by dolichol-sugars. The net result is assembly of the compound illustrated in Figure 47–8 and referred to in shorthand as Dol-P-P-GlcNAc2 Man9 Glc3.
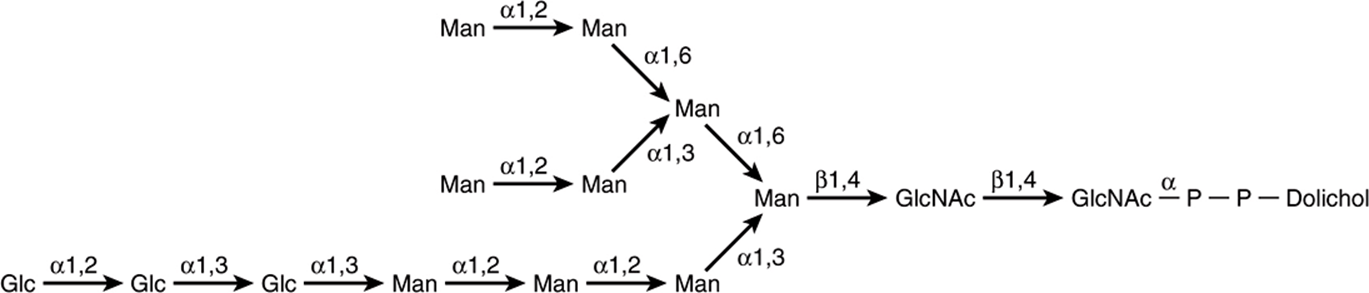
FIGURE 47–8 Structure of dolichol-P-P-oligosaccharide. (With permission from Li E, et al: Structure of the lipid-linked oligosaccharide precursor of the complex-type oligosaccharides of the vesicular stomatitis virus G protein. J Biol Chem 1978;253:7762.)
The oligosaccharide linked to dolichol-P-P is transferred en bloc to form an N-glycosidic bond with one or more specific Asn residues of an acceptor protein emerging from the luminal surface of the membrane of the endoplasmic reticulum. The reaction is catalyzed by oligosaccharide: protein transferase, a membrane-associated enzyme complex. The transferase will recognize and transfer any substrate with the general structure Dol-P-P-(GlcNAc)2-R, but it has a strong preference for the Dol-P-P-GlcNAc2Man9Glc3 structure. Glycosylation occurs at the Asn residue of an Asn-XSer/Thr tripeptide sequence, where X is any amino acid except proline, aspartic acid, or glutamic acid. A tripeptide site contained within a β turn is favored. Only about one-third of the Asn residues that are potential acceptor sites are actually glycosylated, suggesting that factors other than the tripeptide are also important.
The acceptor proteins are of both the secretory and integral membrane class. Cytosolic proteins are rarely glycosylated. The transfer reaction and subsequent processes in the glycosylation of N-linked glycoproteins, along with their subcellular locations, are depicted in Figure 47–9. The other product of the oligosaccharide: protein transferase reaction is dolichol-P-P, which is subsequently converted to dolichol-P by a phosphatase. The dolichol-P can serve again as an acceptor for the synthesis of another molecule of Dol-P-P-oligosaccharide.
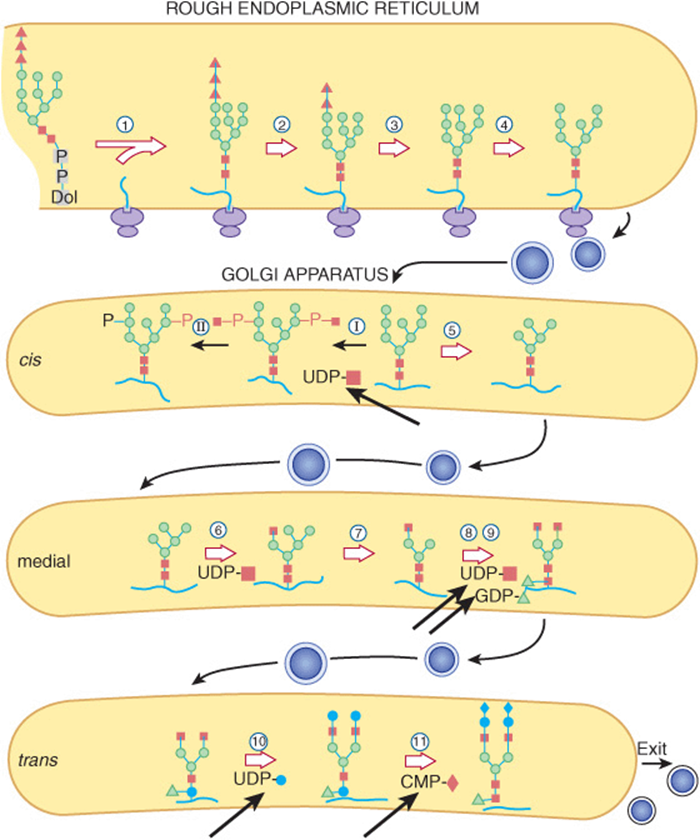
FIGURE 47–9 Schematic pathway of oligosaccharide processing. The reactions are catalyzed by the following enzymes: ![]() oligosaccharide: protein transferase;
oligosaccharide: protein transferase; ![]() α-glucosidase I;
α-glucosidase I; ![]() α-glucosidase II;
α-glucosidase II; ![]() endoplasmic reticulum α 1,2-mannosidase; I N-acetylglucosaminylphosphotransferase; II N-acetylglucosamine-1-phosphodiester α-N-acetylglucosaminidase;
endoplasmic reticulum α 1,2-mannosidase; I N-acetylglucosaminylphosphotransferase; II N-acetylglucosamine-1-phosphodiester α-N-acetylglucosaminidase; ![]() Golgi apparatus α-mannosidase I;
Golgi apparatus α-mannosidase I; ![]() N-acetylglucosaminyltransferase I;
N-acetylglucosaminyltransferase I; ![]() Golgi apparatus α-mannosidase II;
Golgi apparatus α-mannosidase II; ![]() N-acetylglucosaminyltransferase II;
N-acetylglucosaminyltransferase II; ![]() fucosyltransferase;
fucosyltransferase; ![]() galactosyltransferase;
galactosyltransferase; ![]() sialyltransferase. The thick arrows indicate various nucleotide sugars involved in the overall scheme. (Solid square, N-acetylglucosamine; open circle, mannose; solid triangle, glucose; open triangle, fucose; solid circle, galactose; solid diamond, sialic acid.) (Reproduced, with permission, from Kornfeld R, Kornfeld S: Assembly of asparagine-linked oligosaccharides. Annu Rev Biochem 1985;54:631. Copyright © 1985 by Annual Reviews. Reprinted with permission.)
sialyltransferase. The thick arrows indicate various nucleotide sugars involved in the overall scheme. (Solid square, N-acetylglucosamine; open circle, mannose; solid triangle, glucose; open triangle, fucose; solid circle, galactose; solid diamond, sialic acid.) (Reproduced, with permission, from Kornfeld R, Kornfeld S: Assembly of asparagine-linked oligosaccharides. Annu Rev Biochem 1985;54:631. Copyright © 1985 by Annual Reviews. Reprinted with permission.)
Processing ofthe Oligosaccharide Chain
1. Early Phase: the various reactions involved are indicated in Figure 47–9. The oligosaccharide: protein transferase catalyzes reaction 1 (see above). Reactions 2 and 3 involve the removal of the terminal Glc residue by glucosidase I and of the next two Glc residues by glucosidase II, respectively. In the case of high-mannose glycoproteins, the process may stop here, or up to four Man residues may also be removed. However, to form complex chains, additional steps are necessary, as follows. Four external Man residues are removed in reactions 4 and 5 by at least two different mannosidases. In reaction 6, a GlcNAc residue is added to the Man residue of the Man 1-3 arm by GlcNAc transferase I. The action of this latter enzyme permits the occurrence of reaction 7, a reaction catalyzed by yet another mannosidase (Golgi α-mannosidase II) and which results in a reduction of the Man residues to the core number of three (Figure 47–5).
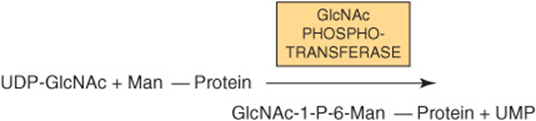
An important additional pathway is indicated in reactions I and II of Figure 47–9. This involves enzymes destined for lysosomes. Such enzymes are targeted to the lysosomes by a specific chemical marker. In reaction I, a residue of GlcNAc-1-P is added to carbon 6 of one or more specific Man residues of these enzymes. The reaction is catalyzed by a GlcNAc phosphotransferase, which uses UDPGlcNAc as the donor and generates UMP as the other product:
In reaction II, the GlcNAc is removed by the action of a phosphodiesterase, leaving the Man residues phosphorylated in the sixth position:
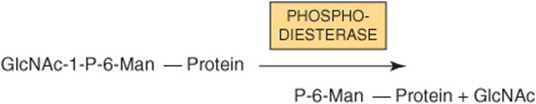
Man 6-P receptors, located in the Golgi apparatus, bind the Man 6-P residues of these enzymes and direct them to the lysosomes. Fibroblasts from patients with I-cell disease (see below) are severely deficient in the activity of the GlcNAc phosphotransferase.
2. Late Phase: to assemble a typical complex oligosaccharide chain, additional sugars must be added to the structure formed in reaction 7. Hence, in reaction 8, a second GlcNAc is added to the peripheral Man residue of the other arm of the bi-antennary structure shown in Figure 47–9; the enzyme catalyzing this step is GlcNAc transferase II. Reactions 9, 10, and 11 involve the addition of Fuc, Gal, and NeuAc residues at the sites indicated, in reactions catalyzed by fucosyl, galactosyl, and sialyl transferases, respectively. The assembly of poly-N-acetyllactosamine chains requires additional GlcNAc transferases.
The Endoplasmic Reticulum & Golgi Apparatus Are the Major Sites of Glycosylation
As indicated in Figure 47–9, the endoplasmic reticulum and the Golgi apparatus are the major sites involved in glycosylation processes. The assembly of Dol-P-P-oligosaccharide occurs on both the cytoplasmic and luminal surfaces of the ER membranes. Addition of the oligosaccharide to protein occurs in the rough endoplasmic reticulum during or after translation. Removal of the Glc and some of the peripheral Man residues also occurs in the endoplasmic reticulum. The Golgi apparatus is composed of cis, medial, and trans cisternae; these can be separated by appropriate centrifugation procedures. Vesicles containing glycoproteins bud off in the endoplasmic reticulum and are transported to the cis-Golgi. Various studies have shown that the enzymes involved in glycoprotein processing show differential locations in the cisternae of the Golgi. As indicated in Figure 47–9, Golgi α-mannosidase I (catalyzing reaction 5) is located mainly in the cis-Golgi, whereas GlcNAc transferase I (catalyzing reaction 6) appears to be located in the medial Golgi, and the fucosyl, galactosyl, and sialyl transferases (catalyzing reactions 9, 10, and 11) are located mainly in the trans-Golgi. The major features of the biosynthesis of N-linked glycoproteins are summarized in Table 47-10 and should be contrasted with those previously listed (Table 47-9) for O-linked glycoproteins.
TABLE 47–10 Summary of Main Features of N-Glycosylation

Some Glycan Intermediates Formed During N-Glycosylation Have Specific Functions
The following are a number of specific functions of N-glycan chains that have been established or are under investigation: (1) The involvement of the mannose 6-P signal in targeting of certain lysosomal enzymes is clear (see above and discussion of I-cell disease, in the following). (2) It is likely that the large N-glycan chains present on newly synthesized glycoproteins may assist in keeping these proteins in a soluble state inside the lumen of the endoplasmic reticulum. (3) One species of N-glycan chains has been shown to play a role in the folding and retention of certain glycoproteins in the lumen of the endoplasmic reticulum. Calnexin is a protein present in the endoplasmic reticulum membrane that acts as a chaperone (Chapter 46) and lectin. Binding to calnexin prevents a glycoprotein from aggregating. It has been found that calnexin will bind specifically to a number of glycoproteins (eg, the influenza virus hemagglutinin [HA]) that possess the monoglycosylated core structure. This species is the product of reaction 2 shown in Figure 47–9, but from which the terminal glucose residue has been removed, leaving only the innermost glucose attached. Calnexin and the bound glycoprotein form a complex with ERp57, a homolog of protein disulfide isomerase (PDI), which catalyzes disulfide bond interchange, facilitating proper folding. The bound glycoprotein is released from its complex with calnexin-ERp57 when the sole remaining glucose is hydrolyzed by glucosidase II and leaves the ER if properly folded. If not properly folded, an ER glucosyltransferase recognizes this and reglucosylates the glycoprotein, which rebinds to the calnexin-Erp57 complex. If now properly folded, the glycoprotein is again deglucosylated and leaves the ER. If not capable of proper folding, it is translocated out of the ER into the cytoplasm, where it is degraded (compare Figure 46–8). This so-called calnexin cycle is illustrated in Figure 47–10. In this way, calnexin retains certain partly folded (or misfolded) glycoproteins and releases them when further folding has occurred. The glucosyltransferase, by sensing the folding of the glycoprotein and only reglucosylating misfolded proteins, is a key component of the cycle. The calnexin cycle is an important component of the quality control systems operating in the lumen of the ER. The soluble ER protein calreticulin performs a similar function.
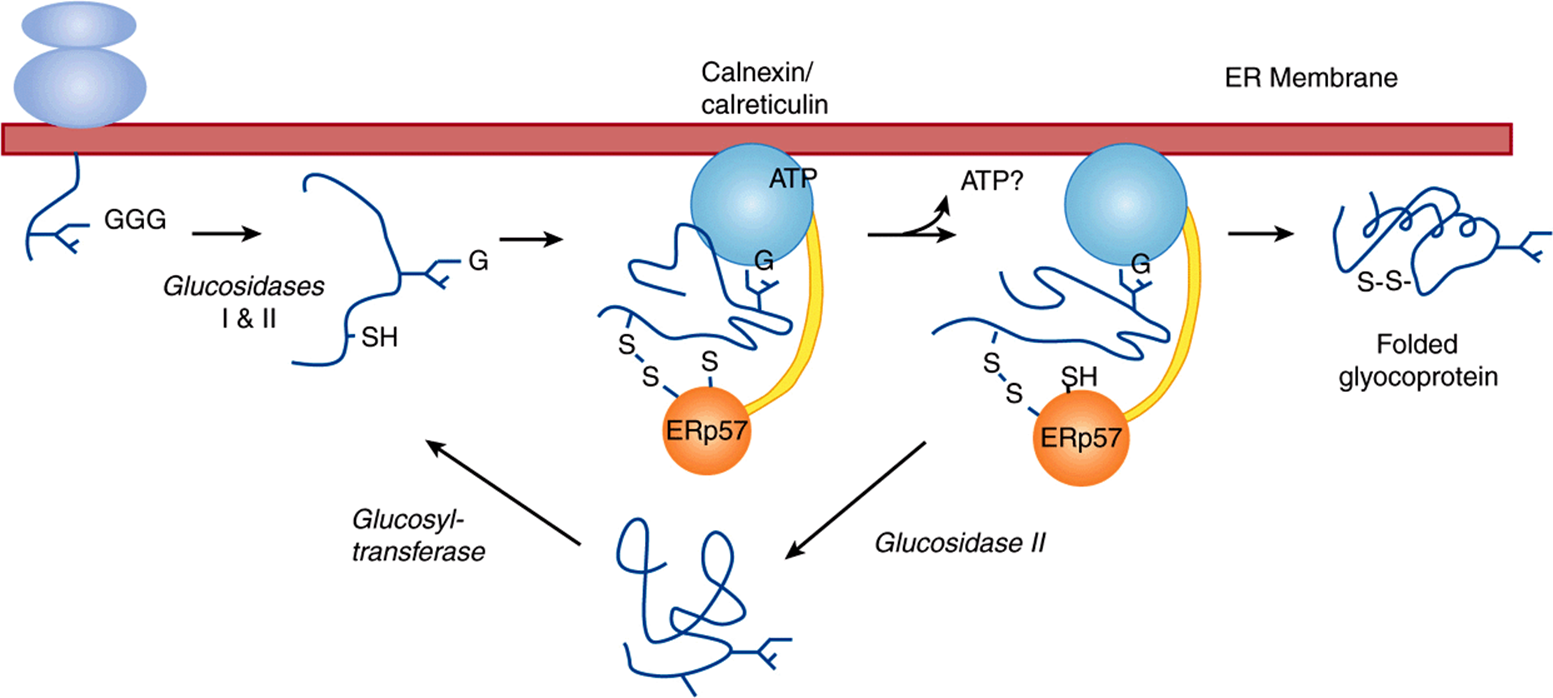
FIGURE 47–10 Model of the calnexin cycle. As a nascent (growing) polypeptide chain enters the ER, certain Asn residues are glycosylated by addition of Glc3Man9GlcNAc2 (see the text). The outermost two molecules of glucose are removed via the actions of glucosidases I and II. This exposes the innermost molecule of glucose, which is recognized by the lectin sites of calnexin and calreticulin. In their ATP-bound state, calnexin and calreticulin bind to the monoglucosylated oligosaccharide (via their lectin sites) as well as to hydrophobic segments of the unfolded glycoprotein (via their polypeptide binding or chaperone sites). Glycoprotein dissociation involves the action of glucosidase II to remove the terminal glucose and also a change in affinity of the polypeptide binding site. After dissociation, if folding does not occur rapidly, the glycoprotein is reglucosylated by an ER glucosyltransferase, which acts only on non-native protein conformers (conformer = a protein in one of several possible conformations). The reglucosylated glycoprotein can then rebind to the ATP form of calnexin/calreticulin. Thus, both the glucosyltransferase and calnexin/calreticulin act as folding sensors. This cycle of binding and release has three functions: it prevents glycoprotein aggregation; it retains non-native conformers in the ER until a native structure is acquired (quality control); and binding to calnexin/calreticulin brings ERp57 into proximity with the non-native glycoprotein. ERp57 catalyzes disulfide bond formation and isomerization within the glycoprotein substrate, assisting it to assume its native conformation. If the glycoprotein is not capable of proper folding, it is translocated out of the ER into the cytoplasm for proteosomal degradation (compare Figure 46–8). Calreticulin, a soluble ER protein, plays a similar role to calnexin. (G, glucose.) (Figure and legend generously supplied by Dr D B Williams, and modified slightly with his permission.)
Several Factors Regulate the Glycosylation of Glycoproteins
It is evident that glycosylation of glycoproteins is a complex process involving a large number of enzymes. It has been estimated that some 1% of the human genome may be involved with glycosylation events. Another index of its complexity is that more than ten distinct GlcNAc transferases involved in glycoprotein biosynthesis have been reported, and others are theoretically possible. Multiple species of the other glycosyl-transferases (eg, sialyltransferases) also exist. Controlling factors of the first stage of N-linked glycoprotein biosynthesis (ie, oligosaccharide assembly and transfer) include (1) the presence of suitable acceptor sites in proteins, (2) the tissue level of Dol-P, and (3) the activity of the oligosaccharide: protein transferase.
Some factors known to be involved in the regulation of oligosaccharide processing are listed in Table 47-11. Two of the points listed merit further comment. First, species variations among processing enzymes have assumed importance in relation to the production of glycoproteins of therapeutic use by means of recombinant DNA technology. For instance, recombinant erythropoietin (epoetin alfa; EPO) is sometimes administered to patients with certain types of chronic anemia in order to stimulate erythropoiesis. The half-life of EPO in plasma is influenced by the nature of its glycosylation pattern, with certain patterns being associated with a short half-life, appreciably limiting its period of therapeutic effectiveness. It is thus important to harvest EPO from host cells that confer a pattern of glycosylation consistent with a normal half-life in plasma. Second, there is great interest in analysis of the activities of glycoprotein-processing enzymes in various types of cancer cells. These cells have often been found to synthesize different oligosaccharide chains (eg, they often exhibit greater branching) from those made in control cells. This could be due to cancer cells containing different patterns of glycosyltransferases from those exhibited by corresponding normal cells, due to specific gene activation or repression. The differences in oligosaccharide chains could affect adhesive interactions between cancer cells and their normal parent tissue cells, contributing to metastasis. If a correlation could be found between the activity of particular processing enzymes and the metastatic properties of cancer cells, this could be important as it might permit synthesis of drugs to inhibit these enzymes and, secondarily, metastasis.
TABLE 47–11 Some Factors Affecting the Activities of Glycoprotein Processing Enzymes
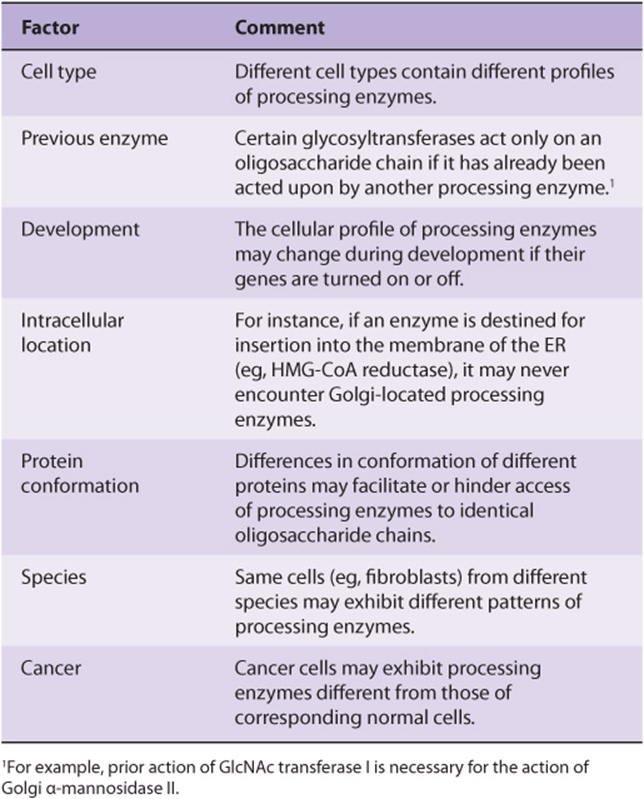
The genes encoding many glycosyltransferases have already been cloned, and others are under study. Cloning has revealed new information on both protein and gene structures. The latter should also cast light on the mechanisms involved in their transcriptional control, and gene knockout studies are being used to evaluate the biologic importance of various glycosyltransferases.
Tunicamycin Inhibits N- But Not O-Glycosylation
A number of compounds are known to inhibit various reactions involved in glycoprotein processing. Tunicamycin, deoxynojirimycin, and swainsonine are three such agents. The reactions they inhibit are indicated in Table 47-12.These agents can be used experimentally to inhibit various stages of glycoprotein biosynthesis and to study the effects of specific alterations upon the process. For instance, if cells are grown in the presence of tunicamycin, no glycosylation of their normally N-linked glycoproteins will occur. In certain cases, lack of glycosylation has been shown to increase the susceptibility of these proteins to proteolysis. Inhibition of glycosylation does not appear to have a consistent effect upon the secretion of glycoproteins that are normally secreted. The inhibitors of glycoprotein processing listed in Table 47-12 do not affect the biosynthesis of O-linked glycoproteins. The extension of O-linked chains can be prevented by GalNAc-benzyl. This compound competes with natural glycoprotein substrates and thus prevents chain growth beyond GalNAc.
TABLE 47–12 Three Inhibitors of Enzymes Involved in the N-Glycosylation of Glycoproteins and Their Sites of Action

SOME PROTEINS ARE ANCHORED TO THE PLASMA MEMBRANE BY GLYCOPHOSPHATIDYL-INOSITOL STRUCTURES
GPI-linked glycoproteins comprise the third major class of glycoprotein. The GPI structure (sometimes called a “sticky foot”) involved in linkage of the enzyme acetylcholinesterase (ACh esterase) to the plasma membrane of the red blood cell is shown in Figure 47–1. GPI-linked proteins are anchored to the outer leaflet of the plasma membrane by the fatty acids of phosphatidylinositol (PI). The PI is linked via a GlcN moiety to a glycan chain that contains various sugars (eg, Man, GlcN). In turn, the oligosaccharide chain is linked via phosphorylethanolamine in an amide linkage to the carboxyl terminal amino acid of the attached protein. The core of most GPI structures contains one molecule of phosphorylethanolamine, three Man residues, one molecule of GlcN, and one molecule of phosphatidylinositol, as follows.
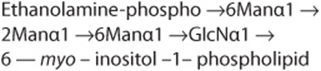
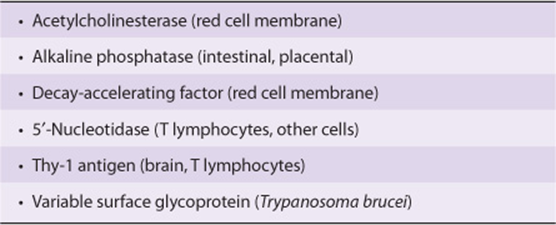
Additional constituents are found in many GPI structures; for example, that shown in Figure 47–1 contains an extra phosphorylethanolamine attached to the middle of the three Man moieties of the glycan and an extra fatty acid attached to GlcN. The functional significance of these variations among structures is not understood. This type of linkage was first detected by the use of bacterial PI-specific phospholipase C (PI-PLC), which was found to release certain proteins from the plasma membrane of cells by splitting the bond indicated in Figure 47–1. Examples of some proteins that are anchored by this type of linkage are given in Table 47-13. At least three possible functions of this type of linkage have been suggested. (1) The GPI anchor may allow greatly enhanced mobility of a protein in the plasma membrane compared with that observed for a protein that contains transmembrane sequences. This is perhaps not surprising, as the GPI anchor is attached only to the outer leaflet of the lipid bilayer, so that it is freer to diffuse than a protein anchored via both leaflets of the bilayer. Increased mobility may be important in facilitating rapid responses to appropriate stimuli. (2) Some GPI anchors may connect with signal transduction pathways. (3) It has been shown that GPI structures can target certain proteins to apical domains and also basolateral domains of the plasma membrane of certain polarized epithelial cells. The biosynthesis of GPI anchors is complex and begins in the endoplasmic reticulum. The GPI anchor is assembled independently by a series of enzyme-catalyzed reactions and then transferred to the carboxyl terminal end of its acceptor protein, accompanied by cleavage of the preexisting carboxyl terminal hydrophobic peptide from that protein. This process is sometimes called glypiation. An acquired defect in an early stage of the biosynthesis of the GPI structure has been implicated in the causation of paroxysmal nocturnal hemoglobinuria (see below).
TABLE 47–13 Some GPI-Linked Proteins
ADVANCED GLYCATION END-PRODUCTS (AGEs) ARE THOUGHT TO BE IMPORTANT IN THE CAUSATION OF TISSUE DAMAGE IN DIABETES MELLITUS
Glycation refers to nonenzymic attachment of sugars (mainly glucose) to amino groups of proteins and also to other molecules (eg, DNA, lipids). Glycation is distinguished from glycosylation because the latter involves enzyme-catalyzed attachment of sugars. When glucose attaches to a protein, intermediate products formed include Schiff bases. These can further be rearranged by the Amadori rearrangement to ketoamines (see Figure 47–11). The overall series of reactions is known as the Maillard reaction. These reactions are involved in the browning of certain foodstuffs that occurs on storage or processing (eg, heating). The end-products of glycation reactions are termed advanced glycation end-products (AGEs).
The major medical interest in AGEs has been in relation to them causing tissue damage in diabetes mellitus, in which the level of blood glucose is often consistently elevated, promoting increased glycation. At constant time intervals, the extent of glycation is more or less proportional to the blood glucose level. It has also been suggested that AGEs are involved in other processes, such as aging.
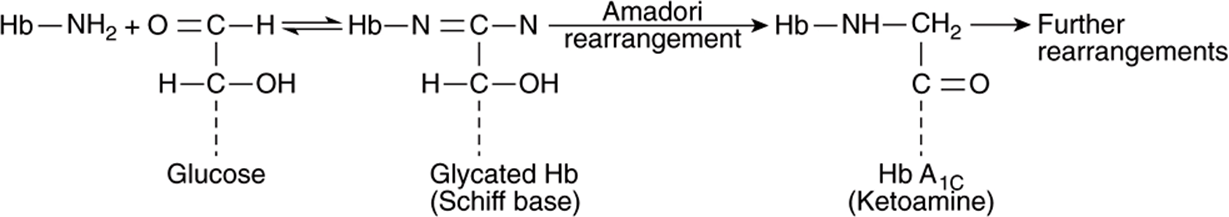
FIGURE 47–11 Formation of AGEs from glucose. Glucose is shown interacting with the amino group of hemoglobin (Hb) forming a Schiff base. This is subject to the Amadori rearrangement, forming a ketoamine. Further rearrangements can occur, leading to other AGEs.
Glycation of collagen and other proteins in the ECM alters their properties (eg, increasing the cross-linking of collagen). Cross-linking can lead to accumulation of various plasma proteins in the walls of blood vessels; in particular, accumulation of LDL can contribute to atherogenesis. AGEs appear to be involved in both microvascular and macrovascular damage in diabetes mellitus (Figure 47–12). Also endothelial cells and macrophages have AGE receptors on their surfaces. Uptake of glycated proteins by these receptors can activate the transcription factor NF-kB (see Chapter 50), generating a variety of cytokines and proinflammatory molecules. It is thus believed that AGEs are one significant contributor to some of the pathologic finding found in diabe. Nonenzymic attachment of glucose to hemoglobin A present in red blood cells (ie, formation of HbA1c) occurs in normal individuals and is increased in patients with diabetes mellitus whose blood sugar levels are elevated. As discussed in Chapter 6, measurement of HbA1 c has become a very important part of the management of patients with diabetes mellitus.
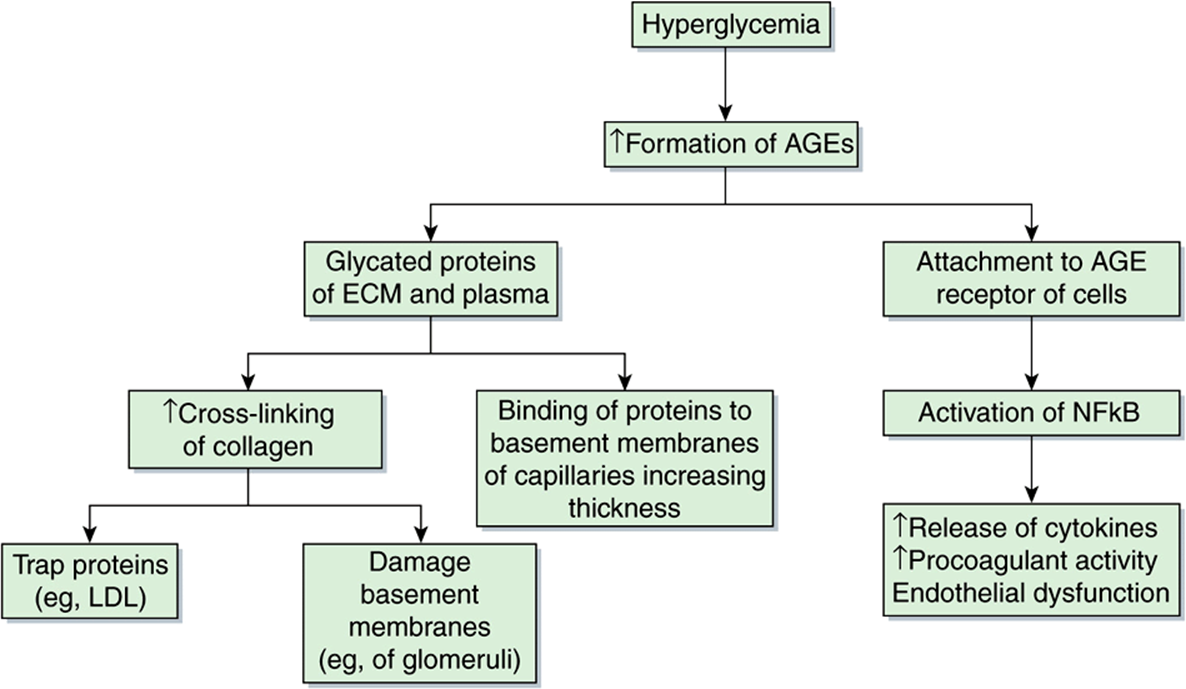
FIGURE 47–12 Some consequences of the formation of AGEs. Hyperglycemia (eg, occurring in poorly controlled diabetes) leads to the formation of AGEs. These can occur in proteins of the ECM or plasma. In the ECM, they can cause increased cross-linking of collagen, which can trap proteins such as LDL (contributing to atherogenesis) and damage basement membranes in the kidneys and other sites. Thickening of basement membranes can also occur by binding of glycated proteins to them. AGEs can attach to AGE receptors on cells, activating NFkB (see Chapter 50), which has several consequences (as shown). Damage to renal basement membranes, thickening of these membranes in capillaries and endothelial dysfunction are found in ongoing uncontrolled diabetes mellitus.
GLYCOPROTEINS ARE INVOLVED IN MANY BIOLOGIC PROCESSES & IN MANY DISEASES
As listed in Table 47-1, glycoproteins have many different functions; some have already been addressed in this chapter and others are described elsewhere in this text (eg, transport molecules, immunologic molecules, and hormones). Here, their involvement in two specific processes—fertilization and inflammation—will be briefly described. In addition, the bases of a number of diseases that are due to abnormalities in the synthesis and degradation of glycoproteins will be summarized.
Glycoproteins Are Important in Fertilization
To reach the plasma membrane of an oocyte, a sperm has to traverse the zona pellucida (ZP), a thick, transparent, noncellular envelope that surrounds the oocyte. The zona pellucida contains three glycoproteins of interest, ZP1-3. Of particular note is ZP3, an O-linked glycoprotein that functions as a receptor for the sperm. A protein on the sperm surface, possibly galactosyl transferase, interacts specifically with oligosaccharide chains of ZP3; in at least certain species (eg, the mouse), this interaction, by transmembrane signaling, induces the acrosomal reaction, in which enzymes such as proteases and hyaluronidase and other contents of the acrosome of the sperm are released. Liberation of these enzymes helps the sperm to pass through the zona pellucida and reach the plasma membrane (PM) of the oocyte. In hamsters, it has been shown that another glycoprotein, PH-30, is important in both the binding of the PM of the sperm to the PM of the oocyte, and also in the subsequent fusion of the two membranes. These interactions enable the sperm to enter and thus fertilize the oocyte. It may be possible to inhibit fertilization by developing drugs or antibodies that interfere with the normal functions of ZP3 and PH-30 and which would thus act as contraceptive agents.
Selectins Play Key Roles in Inflammation & in Lymphocyte Homing
Leukocytes play important roles in many inflammatory and immunologic phenomena. The first steps in many of these phenomena are interactions between circulating leukocytes and endothelial cells prior to passage of the former out of the circulation. Work done to identify specific molecules on the surfaces of the cells involved in such interactions has revealed that leukocytes and endothelial cells contain on their surfaces specific lectins, called selectins, that participate in their intercellular adhesion. Features of the three major classes of selectins are summarized in Table 47-14. Selectins are single-chain Ca2+-binding transmembrane proteins that contain a number of domains (Figure 47–13). Their amino terminal ends contain the lectin domain, which is involved in binding to specific carbohydrate ligands.
TABLE 47–14 Some Molecules Involved in Leukocyte-Endothelial Cell Interactions
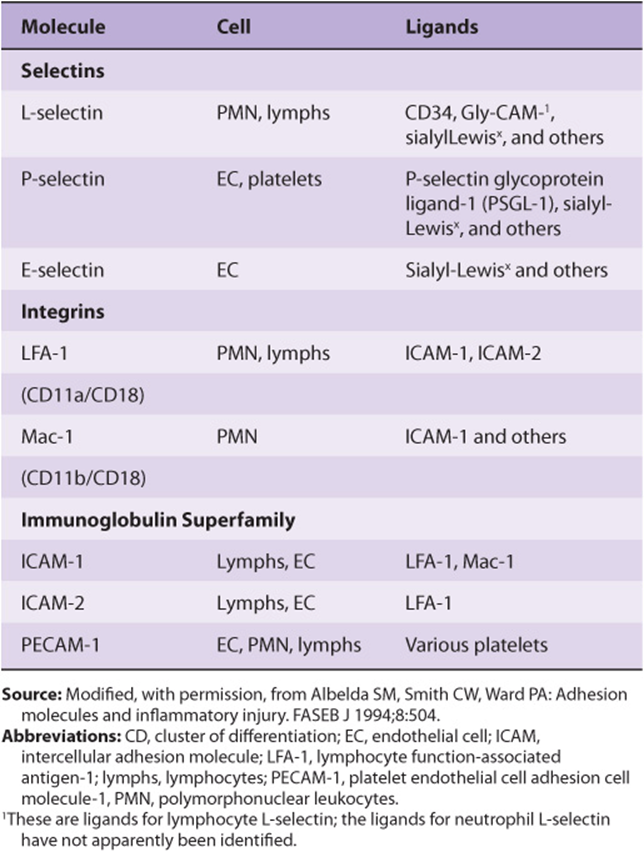
FIGURE 47–13 Schematic of the structure of human L-selectin. The extracellular portion contains an amino terminal domain homologous to C-type lectins and an adjacent epidermal growth factor-like domain. These are followed by a variable number of complement regulatory-like modules (numbered circles) and a trans-membrane sequence (black diamond). A short cytoplasmic sequence (red rectangle) is at the carboxyl terminal. The structures of P- and E-selectin are similar to that shown except that they contain more complement-regulatory modules. The numbers of amino acids in L-, P-, and E- selectins, as deduced from the cDNA sequences, are 385, 789, and 589, respectively. (Reproduced, with permission, from Bevilacqua MP, Nelson RM: Selectins. J Clin Invest 1993;91:370.)
The adhesion of neutrophils to endothelial cells of post-capillary venules can be considered to occur in four stages, as shown in Figure 47–14. The initial baseline stage is succeeded by slowing or rolling of the neutrophils, mediated by selectins. Interactions between L-selectin on the neutrophil surface and CD34 and GlyCAM-1 or other glycoproteins on the endothelial surface are involved. These particular interactions are initially short-lived, and the overall binding is of relatively low affinity, permitting rolling. However, during this stage, activation of the neutrophils by various chemical mediators (discussed below) occurs, resulting in a change of shape of the neutrophils and firm adhesion of these cells to the endothelium. An additional set of adhesion molecules is involved in firm adhesion, namely, LFA-1 and Mac-1 on the neutrophils and ICAM-1 and ICAM-2 on endothelial cells. LFA-1 and Mac-1 are CD11/CD18 integrins (see Chapter 52 for a discussion of integrins), whereas ICAM-1 and ICAM-2 are members of the immunoglobulin superfamily. The fourth stage is transmigration of the neutrophils across the endothelial wall. For this to occur, the neutrophils insert pseudopods into the junctions between endothelial cells, squeeze through these junctions, cross the basement membrane, and then are free to migrate in the extravascular space. Platelet-endothelial cell adhesion molecule-1 (PECAM-1) has been found to be localized at the junctions of endothelial cells and thus may have a role in transmigration. A variety of biomolecules have been found to be involved in activation of neutrophil and endothelial cells, including tumor necrosis factor, various interleukins, platelet activating factor (PAF), leukotriene B4, and certain complement fragments. These compounds stimulate various signaling pathways, resulting in changes in cell shape and function, and some are also chemotactic. One important functional change is recruitment of selectins to the cell surface, as in some cases selectins are stored in granules (eg, in endothe-lial cells and platelets).
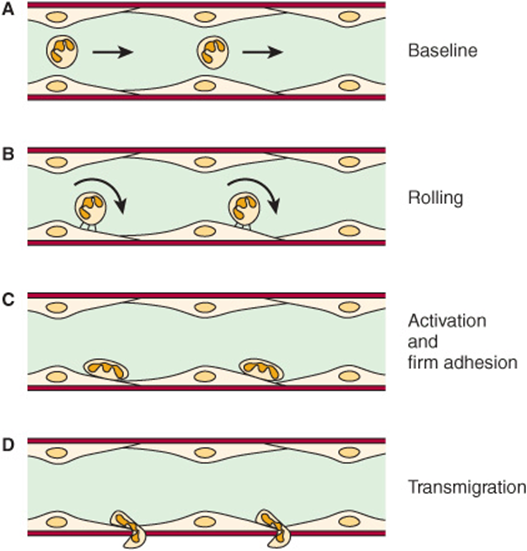
FIGURE 47–14 Schematic of neutrophil-endothelial cell interactions. (A) Baseline conditions: Neutrophils do not adhere to the vessel wall. (B) The first event is the slowing or rolling of the neutrophils within the vessel (venule) mediated by selectins. (C) Activation occurs, resulting in neutrophils firmly adhering to the surfaces of endothelial cells and also assuming a flattened shape. This requires interaction of activated CD18 integrins on neutrophils with ICAM-1 on the endothelium. (D) The neutrophils then migrate through the junctions of endothelial cells into the interstitial tissue; this requires involvement of PECAM-1. Chemotaxis is also involved in this latter stage. (Reproduced, with permission, from Albelda SM, Smith CW, Ward PA: Adhesion molecules and inflammatory injury. FASEB J 1994;8;504.)
The precise chemical nature of some of the ligands involved in selectin-ligand interactions has been determined. All three selectins bind sialylated and fucosylated oligosaccharides, and in particular all three bind sialyl-Lewisx(Figure 47–15), a structure present on both glycoproteins and glycolipids. Whether this compound is the actual ligand involved in vivo is not established. Sulfated molecules, such as the sulfatides (Chapter 15), may be ligands in certain instances. This basic knowledge is being used in attempts to synthesize compounds that block selectin-ligand interactions and thus may inhibit the inflammatory response. Approaches include administration of specific monoclonal antibodies or of chemically synthesized analogs of sialyl-Lewisx, both of which bind selectins. Cancer cells often exhibit sialyl-Lewisx and other selectin ligands on their surfaces. It is thought that these ligands play a role in the invasion and metastasis of cancer cells.
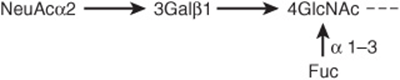
FIGURE 47–15 Schematic representation of the structure of sialyl-Lewisx.
Abnormalities in the Synthesis of Glycoproteins Underlie Certain Diseases
Table 47-15 lists a number of conditions in which abnormalities in the synthesis of glycoproteins are of importance. As mentioned above, many cancer cells exhibit different profiles of oligosaccharide chains on their surfaces, some of which may contribute to metastasis.
The congenital disorders of glycosylation (CDG) are a group of disorders of considerable current interest. The major features of these conditions are summarized in Table 47-16.
TABLE 47–15 Some Diseases Due to or Involving Abnormalities in the Biosynthesis of Glycoproteins
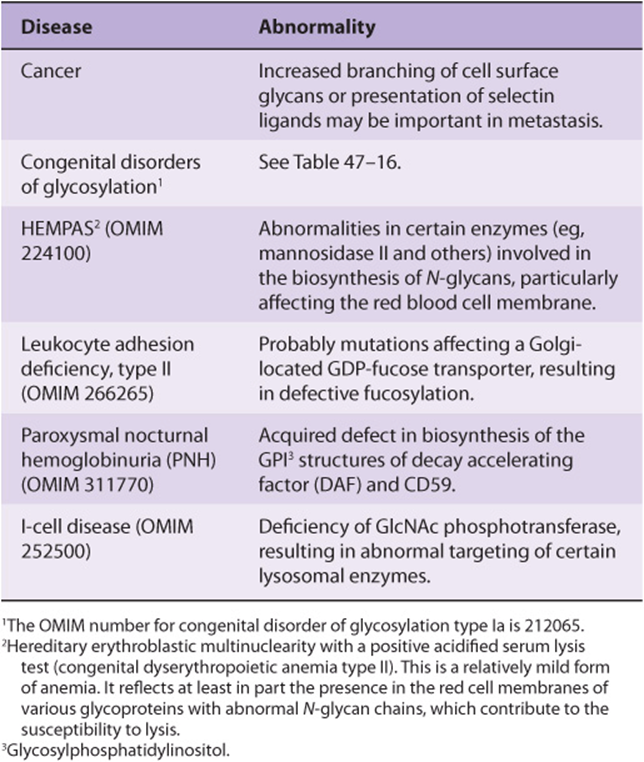
TABLE 47–16 Major Features of the Congenital Disorders of Glycosylation

Leukocyte adhesion deficiency (LAD) II is a rare condition probably due to mutations affecting the activity of a Golgi-located GDP-fucose transporter. It can be considered a congenital disorder of glycosylation. The absence of fucosylated ligands for selectins leads to a marked decrease in neutrophil rolling. Subjects suffer life-threatening, recurrent bacterial infections, and also psychomotor and mental retardation. The condition appears to respond to oral fucose.
Hereditary erythroblastic multinuclearity with a positive acidified lysis test—congenital dyserythropoietic anemia type II—is another disorder in which abnormalities in the processing of N-glycans are thought to be involved. Some cases have been claimed to be due to defects in alpha-mannosidase II.
PNH is an acquired mild anemia characterized by the presence of hemoglobin in urine due to hemolysis of red cells, particularly during sleep. This latter phenomenon may reflect a slight drop in plasma pH during sleep, which increases susceptibility to lysis by the complement system (Chapter 50). The basic defect in PNH is the acquisition of somatic mutations in the PIG-A (for PI glycan class A) gene of certain hematopoietic cells. The product of this gene appears to be the enzyme that links glucosamine to PI in the GPI structure (Figure 47–1). Thus, proteins that are anchored by a GPI linkage are deficient in the red cell membrane. Two proteins are of particular interest: decay accelerating factor and another protein designated CD59. They normally interact with certain components of the complement system (Chapter 50) to prevent the hemolytic actions of the latter. However, when they are deficient, the complement system can act on the red cell membrane to cause hemolysis. A monoclonal antibody to C5, a terminal component of the complement system, has proven useful in the management of PNH by inhibiting the complement cascade. PNH can be diagnosed relatively simply, as the red cells are much more sensitive to hemolysis in normal serum acidified to pH 6.2 (Ham’s test); the complement system is activated under these conditions, but normal cells are not affected. Figure 47–16 summarizes the etiology of PNH.
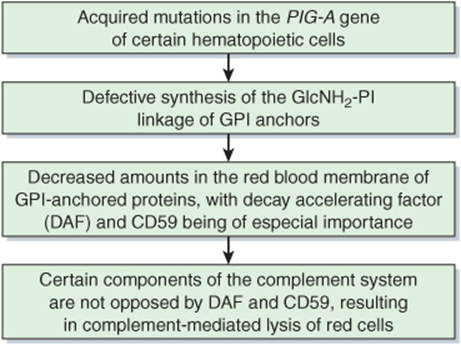
FIGURE 47–16 Scheme of causation of paroxysmal nocturnal hemoglobinuria (OMIM 311770).
Study of the congenital muscular dystrophies (CMDs) has revealed that certain of them (eg, the Walker-Warburg syndrome, muscle-eye-brain disease, Fukuyama CMD) are the result of defects in the synthesis of glycans in the protein α-dystroglycan (α-DG). This protein protrudes from the surface membrane of muscle cells and interacts with laminin-2 (merosin) in the basal lamina (see Figure 49–11). If the glycans of α-DG are not correctly formed (as a result of mutations in genes encoding certain glycosyltransferases), this results in defective interaction of α-DG with laminin, which in turn leads to the development of a CMD.
Rheumatoid arthritis is associated with an alteration in the glycosylation of circulating immunoglobulin G (IgG) molecules (Chapter 50), such that they lack galactose in their Fc regions and terminate in GlcNAc. Mannose-binding protein (MBP, not to be confused with the mannose 6-P receptor), a C-lectin synthesized by liver cells and secreted into the circulation, binds mannose, GlcNAc, and certain other sugars. It can thus bind agalactosyl IgG molecules, which subsequently activate the complement system (see Chapter 50), contributing to chronic inflammation in the synovial membranes of joints.
MBP can also bind the above sugars when they are present on the surfaces of certain bacteria, fungi, and viruses, preparing these pathogens for opsonization or for destruction by the complement system. This is an example of innate immunity, not involving immunoglobulins or T lymphocytes. Deficiency of this protein in young infants as a result of mutation renders them very susceptible to recurrent infections.
I-Cell Disease Results from Faulty Targeting of Lysosomal Enzymes
As indicated above, Man 6-P serves as a chemical marker to target certain lysosomal enzymes to that organelle. Analysis of cultured fibroblasts derived from patients with I-cell (inclusion cell) disease played a large part in revealing the above role of Man 6-P. I-cell disease is an uncommon condition characterized by severe progressive psychomotor retardation and a variety of physical signs, with death often occurring in the first decade. Cultured cells from patients with I-cell disease were found to lack almost all of the normal lysosomal enzymes; the lysosomes thus accumulate many different types of undegraded molecules, forming inclusion bodies. Samples of plasma from patients with the disease were observed to contain very high activities of lysosomal enzymes; this suggested that the enzymes were being synthesized but were failing to reach their proper intracellular destination and were instead being secreted. Cultured cells from patients with the disease were noted to take up exogenously added lysosomal enzymes obtained from normal subjects, indicating that the cells contained a normal receptor on their surfaces for endocytic uptake of lysosomal enzymes. In addition, this finding suggested that lysosomal enzymes from patients with I-cell disease might lack a recognition marker. Further studies revealed that lysosomal enzymes from normal individuals carried the Man 6-P recognition marker described above, which interacted with a specific intracellular protein, the Man 6-P receptor. Cultured cells from patients with I-cell disease were then found to be deficient in the activity of the cis-Golgi-located GlcNAc phosphotransferase, explaining how their lysosomal enzymes failed to acquire the Man 6-P marker. It is now known that there are two Man 6-P receptor proteins, one of high (275 kDa) and one of low (46 kDa) molecular mass. These proteins are -ectins, recognizing Man 6-P. The former is cation-independent and also binds IGF-II (hence it is named the Man 6-P-IGFII receptor), whereas the latter is cation-dependent in some species and does not bind IGF-II. It appears that both receptors function in the intracellular sorting of lysosomal enzymes into clathrin-coated vesicles, which occurs in the trans-Golgi subsequent to synthesis of Man 6-P in the cis-Golgi. These vesicles then leave the Golgi and fuse with a prelysosomal compartment. The low pH in this compartment causes the lysosomal enzymes to dissociate from their receptors and subsequently enter into lysosomes. The receptors are recycled and reused. Only the smaller receptor functions in the endocytosis of extracellular lysosomal enzymes, which is a minor pathway for lysosomal location. Not all cells employ the Man 6-P receptor to target their lysosomal enzymes (eg, hepatocytes use a different but undefined pathway); furthermore, not all lysosomal enzymes are targeted by this mechanism. Thus, biochemical investigations of I-cell disease not only led to elucidation of its basis, but also contributed significantly to knowledge of how newly synthesized proteins are targeted to specific organelles, in this case the lysosome. Figure 47–17 summarizes the causation of I-cell disease.

FIGURE 47–17 Summary of the causation of I-cell disease (OMIM 252500).
Pseudo-Hurler polydystrophy is another genetic disease closely related to I-cell disease. It is a milder condition, and patients may survive to adulthood. Studies have revealed that the GlcNAc phosphotransferase involved in I-cell disease has several domains, including a catalytic domain and a domain that specifically recognizes and interacts with lysosomal enzymes. It has been proposed that the defect in pseudo-Hurler polydystrophy lies in the latter domain, and the retention of some catalytic activity results in a milder condition.
Genetic Deficiencies of Glycoprotein Lysosomal Hydrolases Cause Diseases Such as α-Mannosidosis
Glycoproteins, like most other biomolecules, undergo both synthesis and degradation (ie, turnover). Degradation of the oligosaccharide chains of glycoproteins involves a battery of lysosomal hydrolases, including α-neuraminidase, β-galactosidase, β-hexosaminidase, α- and β-mannosidases, α-N-acetylgalactosaminidase, α-fucosidase, endo-β-N-acetyl-glucosaminidase, and aspartylglucosaminidase. The sites of action of the last two enzymes are indicated in the legend to Figure 47–5. Genetically determined defects of the activities of these enzymes can occur, resulting in abnormal degradation of glycoproteins. The accumulation in tissues of such degraded glycoproteins can lead to various diseases. Among the best recognized of these diseases are mannosidosis, fucosidosis, sialidosis, aspartylglycosaminuria, and Schindler disease, due respectively to deficiencies of α-mannosidase, α-fucosidase, α-neuraminidase, aspartylglucosaminidase, and α-N-acetylgalactosaminidase. These diseases, which are relatively uncommon, have a variety of manifestations; some of their major features are listed in Table 47-17. The fact that patients affected by these disorders all show signs referable to the central nervous system reflects the importance of glycoproteins in the development and normal function of that system.
TABLE 47–17 Major Features of Some Diseases1 Due to Deficiencies of Glycoprotein Hydrolases2

THE GLYCANS OF GLYCOCONJUGATES ARE INVOLVED IN THE BINDING OF VIRUSES, BACTERIA & CERTAIN PARASITES TO HUMAN CELLS
A principal feature of glycans, and one that explains many of their biologic actions, is that they bind specifically to a variety of molecules such as proteins or other glycans. One reflection of this is their ability to bind certain viruses, many bacteria and some parasites.
Influenza virus A binds to cell surface glycoprotein receptor molecules containing NeuAc via a protein named hemagglutinin (H). It also possesses a neuraminidase (N) that plays a key role in allowing elution of newly synthesized progeny from infected cells. If this process is inhibited, spread of the viruses is markedly diminished. Inhibitors of this enzyme (eg, zanamivir, oseltamivir) are now available for use in treating patients with influenza. Influenza viruses are classified according to the type of hemagglutinin and neuraminidase that they possess. There are at least 16 types of hemagglutinin and nine types of neuraminidase. Thus, avian influenza virus is classified as H5N1. There is great interest in how this virus attaches to human cells, in view of the possibility of a pandemic occurring. It has been found that the virus preferentially attaches to glycans terminated by the disaccharide galactose—α 2,3-NeuAc (Figure 47–18). However, the predominant disaccharide terminating glycans in cells of the human respiratory tract is galactose → α 2,6-NeuAc. If a change in the structure of the viral hemagglutinin (due to mutation) occurs that allows it to bind to the latter disaccharide, this could greatly increase the potential infectivity of the virus, possibly resulting in very serious consequences.
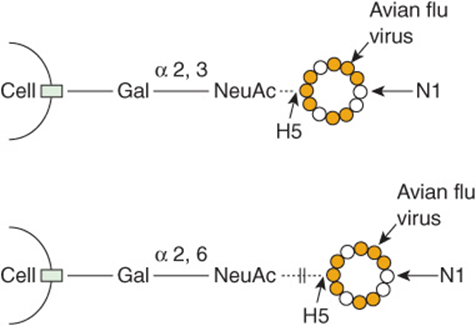
FIGURE 47–18 Schematic representation of binding of the avian influenza virus (H5N1) to a respiratory epithelial cell. The viral hemagglutin (HA) mediates its entry to cells by binding to a glycan on the cell surface that is terminated by the disaccharide galactose → α 2,3-NeuAc. It will not bind to a glycan terminated by galactose → α 2,6-NeuAc, which is the type predominantly found in the human respiratory tract. If the viral HA alters via mutation to be able to bind to the latter disaccharide, this could greatly increase its pathogenicity for humans. (H5, hemagglutinin type 5; N1, neuraminidase type 1.)
Human immunodeficiency virus type 1 (HIV-1), thought by most to be the cause of AIDS, attaches to cells via one of its surface glycoproteins (gp120) and uses another surface glycoprotein (gp 41) to fuse with the host cell membrane. Antibodies to gp 120 develop during infection by HIV-1, and there has been interest in using the protein as a vaccine. One major problem with this approach is that the structure of gp 120 can change relatively rapidly due to mutations, allowing the virus to escape from the neutralizing activity of antibodies directed against it.
Helicobacter pylori is believed to be the major cause of peptic ulcers. Studies have shown that this bacterium binds to at least two different glycans present on the surfaces of epithelial cells in the stomach (see Figure 47–19).This allows it to establish a stable attachment site to the stomach lining, and subsequent secretion of ammonia and other molecules by the bacterium are believed to initiate ulceration.
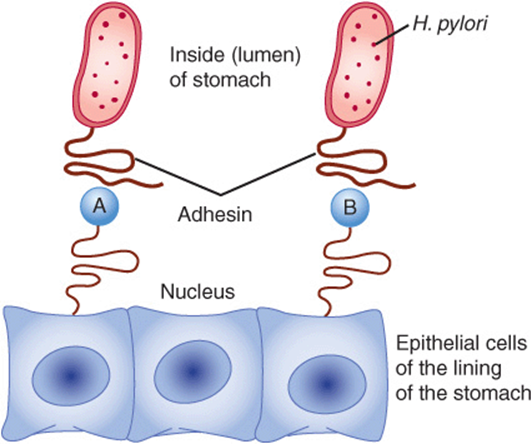
FIGURE 47–19 Attachment of Helicobacterpylori to epithelial cells of the stomach. Adhesin, a protein present in the tail of Hpylori, interacts with two different glycans (structures shown in the figure) present in glycoproteins on the surface of gastric epithelial cells. This provides an attachment site for the bacterium. Subsequently it liberates molecules, such as ammonia, that contribute to initiating peptic ulceration. (A) NeuAcα2,3Galβ1,4—Protein (Neuraminylgalactose); (B) Fucα1,2Galβ1,3GlcNAc—Protein (LewisB substance).
Similarly, many bacteria that cause diarrhea are also known to attach to surface cells of the intestine via glycans present in glycoproteins or glycolipids.
The basic cause of cystic fibrosis (CF) is mutations in the gene encoding CFTR (see Chapters 40 and 57). A major problem in this disease is recurring lung infections by bacteria such as Pseudomonas aeruginosa. In CF, a relative dehydration of respiratory secretions occurs secondary to changes in electrolyte composition in the airway as a result of mutations in CFTR. Bacteria such as P aeruginosa attach to the sugar chains of mucins and find the dehydrated environment in the bronchioles a favorable location in which to multiply.
The attachment of Plasmodium falciparum—one of the types of plasmodia causing malaria—to human cells is mediated by a GPI present on the surface of the parasite.
Various researchers are analyzing the surfaces of viruses, bacteria, parasites and human cells to determine which molecules are involved in attachment. It is important to define the precise nature of the interactions between invading organisms and host cells, as this will hopefully lead to the development of drugs or other agents that will specifically inhibit attachment.
THE PACE OF RESEARCH IN GLYCOMICS IS ACCELERATING
Research on glycoconjugates in the past has been hampered by the lack of availability of suitable technics to determine the structures of glycans. However, appropriate analytical technics are now available (some of which are listed in Table 47-3), as are powerful new genetic technics (eg, knockouts and knockdowns using RNAi molecules). It is certain that research in glycomics will not only provide a wealth of structural information on glyconconjugates, helping to disclose “the sugar code of life,” but will also uncover many new important biologic interactions that are sugar-dependent and will provide targets for drug and other therapies.
SUMMARY
![]() Glycoproteins are widely distributed proteins—with diverse functions—that contain one or more covalently linked carbohydrate chains.
Glycoproteins are widely distributed proteins—with diverse functions—that contain one or more covalently linked carbohydrate chains.
![]() The carbohydrate components of a glycoprotein range from 1% to more than 85% of its weight and may be simple or very complex in the structure. Eight sugars are mainly found in the sugar chains of human glycoproteins: xylose, fucose, galactose, glucose, mannose, N-acetylgalactosamine, N-acetylglucosamine and N-acetylneuraminic acid.
The carbohydrate components of a glycoprotein range from 1% to more than 85% of its weight and may be simple or very complex in the structure. Eight sugars are mainly found in the sugar chains of human glycoproteins: xylose, fucose, galactose, glucose, mannose, N-acetylgalactosamine, N-acetylglucosamine and N-acetylneuraminic acid.
![]() At least certain of the oligosaccharide chains of glycoproteins encode biologic information; they are also important to glycoproteins in modulating their solubility and viscosity, in protecting them against proteolysis, and in their biologic actions.
At least certain of the oligosaccharide chains of glycoproteins encode biologic information; they are also important to glycoproteins in modulating their solubility and viscosity, in protecting them against proteolysis, and in their biologic actions.
![]() The structures of oligosaccharide chains can be elucidated by gas-liquid chromatography, mass spectrometry, and high-resolution NMR spectrometry.
The structures of oligosaccharide chains can be elucidated by gas-liquid chromatography, mass spectrometry, and high-resolution NMR spectrometry.
![]() Glycosidases hydrolyze specific linkages in oligosaccharides and are used to explore both the structures and functions of glycoproteins.
Glycosidases hydrolyze specific linkages in oligosaccharides and are used to explore both the structures and functions of glycoproteins.
![]() Lectins are carbohydrate-binding proteins involved in cell adhesion and many other biologic processes.
Lectins are carbohydrate-binding proteins involved in cell adhesion and many other biologic processes.
![]() The major classes of glycoproteins are O-linked (involving an OH of serine or threonine), N-linked (involving the N of the amide group of asparagine), and GPI-linked.
The major classes of glycoproteins are O-linked (involving an OH of serine or threonine), N-linked (involving the N of the amide group of asparagine), and GPI-linked.
![]() Mucins are a class of O-linked glycoproteins that are distributed on the surfaces of epithelial cells of the respiratory, gastrointestinal, and reproductive tracts.
Mucins are a class of O-linked glycoproteins that are distributed on the surfaces of epithelial cells of the respiratory, gastrointestinal, and reproductive tracts.
![]() The endoplasmic reticulum and Golgi apparatus play a major role in glycosylation reactions involved in the biosynthesis of glycoproteins.
The endoplasmic reticulum and Golgi apparatus play a major role in glycosylation reactions involved in the biosynthesis of glycoproteins.
![]() The oligosaccharide chains of O-linked glycoproteins are synthesized by the stepwise addition of sugars donated by nucleotide sugars in reactions catalyzed by individual specific glycoprotein glycosyltransferases.
The oligosaccharide chains of O-linked glycoproteins are synthesized by the stepwise addition of sugars donated by nucleotide sugars in reactions catalyzed by individual specific glycoprotein glycosyltransferases.
![]() In contrast, the synthesis of N-linked glycoproteins involves a specific dolichol-P-P-oligosaccharide and various glycotransferases and glycosidases. Depending on the enzymes and precursor proteins in a tissue, it can synthesize complex, hybrid, or high-mannose types of N-linked oligosacccharides.
In contrast, the synthesis of N-linked glycoproteins involves a specific dolichol-P-P-oligosaccharide and various glycotransferases and glycosidases. Depending on the enzymes and precursor proteins in a tissue, it can synthesize complex, hybrid, or high-mannose types of N-linked oligosacccharides.
![]() Glycoproteins are implicated in many biologic processes. For instance, they have been found to play key roles in fertilization and inflammation.
Glycoproteins are implicated in many biologic processes. For instance, they have been found to play key roles in fertilization and inflammation.
![]() A number of diseases involving abnormalities in the synthesis and degradation of glycoproteins have been recognized. Glycoproteins are also involved in many other diseases, including influenza, AIDS, rheumatoid arthritis, cystic fibrosis and peptic ulcer.
A number of diseases involving abnormalities in the synthesis and degradation of glycoproteins have been recognized. Glycoproteins are also involved in many other diseases, including influenza, AIDS, rheumatoid arthritis, cystic fibrosis and peptic ulcer.
![]() Developments in the new field of glycomics are likely to provide much new information on the roles of sugars in health and disease and also indicate targets for drug and other types of therapies.
Developments in the new field of glycomics are likely to provide much new information on the roles of sugars in health and disease and also indicate targets for drug and other types of therapies.
REFERENCES
Chandrasekeran A, Srinivasan A, Raman R, et al: Glycan topology determines human adaptation of avian H5N1 virus hemagglutinin. Nat Biotechnology 2008;26:107.
Freeze HH: Congenital disorders of glycosylation: CDG-I, CDG-II, and beyond. Curr Mol Med 2007;7:389.
Kiessling LL, Splain RA: Chemical approaches to glycobiology. Annu Rev Biochem. 2010;79:619.
Kornfeld R, Kornfeld S: Assembly of asparagine-linked oligosaccharides. Annu Rev Biochem 1985;54:631.
Ohtsubo K, Marth JD: Glycosylation in cellular mechanisms of health and disease. Cell 2006;126:855.
Pilobelli KT, Mahal LK: Deciphering the glycocode: the complexity and analytical challenge of glycomics. Curr Opin Chem Biol 2007;11:300.
Ramasamy R, Yan SF, Herold K, et al: Receptor for advanced glycation end products: fundamental roles in the inflammatory response: winding the way to the pathogenesis of endothelial dysfunction and atherosclerosis. Ann NY Acad Sci 2008;1126:7.
Scriver CR, Beaudet AI, Sly WS, et al (editors): The Metabolic and Molecular Bases of Inherited Disease. 8th ed. McGraw-Hill, 2001. (Various chapters in this text and its updated online version [see References for Chapter 1] give in-depth coverage of topics such as I-cell disease and disorders of glycoprotein degradation.)
Taylor ME, Drickamer K: Introduction to Glycobiology. Oxford University Press, 2003.
Varki A, Cummings RD, Esko JD, et al: Essentials of Glycobiology. 2nd ed. Cold Spring Harbor Laboratory Press, 2008.
Von Itzstein M, Plebanski M, Cooke RM, Coppel RL: Hot, sweaty and sticky: the glycobiology of Plasmodium falciparum. Trends Parasitol 2008;24:210.
Werz DB, Seeberger PH: Carbohydrates are the next frontier in pharmaceutical research. Chemistry 2005;11:3194.