CHEMICAL BIOLOGY
Chemistry of Neoglycoproteins
Yuan C. Lee, Biology Department, Johns Hopkins University, Baltimore, Maryland
doi: 10.1002/9780470048672.wecb379
Natural glycoproteins often contain heterogeneous oligosaccharide chains, which make it difficult to elucidate the structure-activity relationship of the carbohydrates of these glycoproteins. Neoglycoproteins were developed to overcome such a problem. Neoglycoproteins are proteins modified with carbohydrates of defined structures. The modification can be random or specific and can use chemical or enzymatic methods. The protein functional groups most often used are α/ε-amino groups, β/γ-carboxyl groups, γ-carboxylamides, sulfhydrils, and phenols. In some cases, existing glycans on glycoproteins can be modified to make the glycan structures homogeneous. Alternatively, glycopeptides or glycoproteins can be synthesized totally de novo, without using any natural proteins, peptides, or oligosaccharides. In addition to providing homogeneous carbohydrate structures on a single protein, neoglycoproteins also provide a multivalency effect (glycoside clustering effect). The use of neoglycoproteins greatly promoted the understanding of the roles of carbohydrates in biologic systems, which led to some clinically important development.
Glycosylation is one of the most complex posttranslational modification of proteins (1). It is well known that glycans attached to glycoproteins are heterogeneous, even at a single glycosy- lation site of a glycoprotein from a single type of cells. Two reasons for this heterogeneity are that multiple possibilities of linking two monomeric units of carbohydrates exist and that carbohydrates have the ability to form branched structure routinely. Branched structures are especially pervasive on the cell surface glycoconjugates. These factors cause great difficulties in elucidating the biologic roles of carbohydrates in glycoproteins and have hampered considerably the progress in this area of research in comparison with proteins and nucleic acids. To partially alleviate such a problem, neoglycoproteins (2) were devised. In neoglycoproteins, carbohydrates attached to proteins are of a known structure and the level of attachment can be varied more broadly than that found in natural glycoproteins. The latter aspect actually is very important in manifesting the “cluster effect” (multivalency effect) (3, 4), for example, a linear increase in valency that produces logarithmically increasing effects. Production of neoglycoproteins needs not start from natural proteins or glycoproteins but can be designed totally de novo. The de novo approach often is used to attain the desired “cluster effect” more precisely than the random attachment of sugars to proteins. Most of methodologies for the preparation of neoglycoproteins are chemical, but incorporation of enzymatic and molecular biologic methods increasingly are gaining popularity. In this article, the emphasis is placed primarily on protein derivatives (neoglycoproteins), but some examples of peptide derivatives and microarrays also are included.
Historical Background
The first neoglycoproteins (without being referred to as such) were prepared by Avery and Goebel, who used sugar-derivatized proteins to raise antibodies directed toward carbohydrates (5, 6). Typically, carbohydrates by themselves are poor antigens. Avery and Goebel demonstrated that by conjugating carbohydrates to proteins, antibodies against carbohydrates can be raised more efficiently than by carbohydrates alone. Avery and Goebel used diazotized p-aminophenyl or p-aminobenzyl derivatives of sugars for conjugation to tyrosyl side chains of protein (diazo-coupling). Using a similar approach, Monsigny and colleagues (7, 8) prepared (GlcNAc)n-BSA to determine the binding specificity of wheat germ agglutinin. Iyer and Goldstein (9) also used the diazo-coupling technique, but Buss and Goldstein (10) improved the coupling efficiency by first converting the anilino-group of the glycoside aglycon into the isothiocyanate group by reacting it with thiophosgene so that the sugar derivatives will react with the amino groups of proteins rather than with the phenolic groups of proteins. Lee and colleagues (11-14) used thioglycosides that contained various functional groups to conjugate to proteins, and the term “neoglycoprotein” was introduced at this time. Thioglycosides were chosen to improve the stability of the carbohydrate-protein links in biologic systems because most glycosidases are incapable of cleaving thioglycosidic links. In addition, unlike earlier neoglycoproteins, the new type of neoglycoproteins did not contain the hydrophobic aryl group, which often causes undesirable nonspecific binding, and the retention of the positive charges of amino groups avoided a disturbance of the charge distribution of the protein. These neoglycoproteins were used for the identification of the sugar binding specificities of plant and animal lectins. Synthesis of neoglycoprotein also was stimulated greatly by the advent of sodium cyanoborohydride (NaCNBH3) to conduct reductive amination (15-17). Contrary to sodium borohydride, NaCNBH3 preferentially reduces the Schiff base (aldimine) so that the modifying carbohydrate reagents, either reducing oligosaccharides or ω-aldehydo glycosides, would not be wasted and much higher yields could be attained. Reducing oligosaccharides were attached conveniently to amino groups of proteins just by mixing the reagents in a suitable buffer, although the reducing terminal sugar would become acyclic and the overall reaction time could be a week or longer. Later on, several amine-borane complexes were used in lieu of NaCNBH3 (18).
Neoglycoproteins have been used as ligands in numerous studies. More recent uses include: substrates for glycosyltransferases, lectin isolation media, and probes for lectins in biologic systems. The techniques for preparation of neoglycoprotein have been advancing steadily, and they now approach the level of the complete de novo synthesis of glycoproteins (19). This type of neoglycoproteins with precise glycosylation sites allows much more focused applications of glycoconjugates.
Preparation of Neoglycoprotein From Natural Proteins
Most neoglycoproteins are prepared using natural proteins— either nonglycosylated or glycosylated. A summary of chemical principles of various conjugation methods is shown in Fig. 1.
Via tyrosyl groups of proteins
The classic method of Avery and Goebel (5, 6) and Tillett et al. (20) used diazotized aminobenzyl derivatives of carbohydrate to conjugate to Tyr groups of globulins and albumins. Aminophenyl glycosides had been used in a similar fashion to prepare BSA derivatives as mentioned above (8). Such products can be very colorful because of the highly conjugated bonding system. However, usually only a limited number of Tyr groups exist in typical proteins, and the diazo coupling is not totally specific to phenolic side chains. Moreover, the chemical stability of products also is a problem. It should be noted that the modification of Tyr often results in the inactivation of enzymes and other biologically active proteins. The introduction of an aromatic ring for each glycosylation site also considerably increases the hydrophobicity of the product, which results in decreased solubility and increased undesirable nonspecific interaction.
A unique approach of carbohydrate-protein conjugation was reported in the case of chitosan. The target protein first is oxidized with tyrosinase to yield o-quinone groups, which can be conjugated readily to chitosan (21). This approach should be applicable to amino-terminated glycosides also.
Via amino groups of proteins
This approach is by far the most popular approach for the preparation of neoglycoproteins because of the ready surface availability of the ε-amino group of Lys side chains and the higher reactivity of ε-amino groups in most proteins.
Reducing oligosaccharides can be conjugated directly to the amino groups by the reductive alkylation (or reductive amination) method (15) between the carbonyl group of sugars and the amino groups of proteins if a prolonged reaction time is no objection. The slow reaction rate in the reductive amination is because of the extremely low concentration of the acyclic form of sugars in solution that provides the reactive aldehyde group. In the case of oligosaccharides, a compounding effect is a decrease in reactivity as the molecular weight increases. This problem can be overcome by providing the ω-aldehydo group in the aglycon (13, 18), which allows the completion of the reuctive alkylation overnight. Another alternative is to reduce the reducing terminus and then use mild periodate oxidation to generate aldehyde group(s) from the acyclic sugar. When oligosaccharide lactones (prepared from bromine oxidation of the reducing oligosaccharides) were used to conjugate to BSA, many days also were required to complete the reaction (22).
Many other reactive derivatives of mono- or oligosaccharides for reactions other than reductive alkylation have been devised (Fig. 1). For example, isothiocarbamate was derived from p-aminophenyl glycosides (10). Glyconate was used to conjugate to the amino groups of proteins (23). For more complex reactions, methyl imidate (derived from cyanomethyl thioglycosides), for example, would react with amino groups to form an amidino link, which has an even higher pKa than the ε-amino group (12). Various activated form of ω-carboxyl glycosides have been used (24-26) to modify amino groups. It should be noted that although amidation converts the amino group into a neutral amide group, reductive alkylation and amidination retain the positive charge of the original amino group, and the protein integrity is better preserved. When the number of amino groups is high (such as in BSA), it is possible to attach a greater number of glycan chains than normally observed in natural glycoproteins.

Figure 1. Various types of linking arms between glycans and proteins.
Via sulfhydryl groups
If a protein contains a surface-oriented free side chain of cysteine, ω-haloacetyl that contains glycoside or glycopeptide can be reacted to form thioether (19, 27). ω-Maleimidoalkyl glycosides or maleimidated glycopeptides (28) also can be used in the same way. However, a free surface-oriented SH in natural proteins/glycoproteins is rather rare in number, and hence the scope of this type of neoglycoprotein preparation also is limited. If glycosylation is desired at a specific site and if a Cys can be introduced in the peptide sequence by genetic engineering, it may allow a custom-designed neoglycoprotein with site-specific glycosylation. It also is possible to modify the amino groups in protein with haloacetyl or maleimide groups that then can react with thiolated carbohydrate derivatives (e.g., 1-thio-oligosaccharides) to form a thioether link. In this case, the number of glycan attachments (on the amino groups) can be far greater compared with the naturally available number of SH side chains.
An alternative to thioether formation is to form a disulfide bond between thioglycoside and the Cys-SH group of protein (29) or by disulfide exchange between disulfide glycoside and Cys-SH (30). The glycan attached via the disulfide bond, however, has a real risk of being detached by a reductive/oxidative cleavage or a sulfide exchange in biologic systems and, thus, is less desirable.
Via special affinity
Chen and Wold (31, 32) used the unique affinity of avidin/streptavidin for biotin to prepare neoglycoproteins. Glycosides that contain a biotin terminus could be bound so tightly to avidin (Kd < 10-13M) that they could be considered virtually covalently bound. The limitation of this method, however, is that the maximum number of glycan attachments is dependent on the valency of avidin, which is four, and the attached glycans face diverse directions. Interestingly, the fourth binding site of avidin turned out to be very difficult to fulfill. The model of biotinylated glycan and avidin also was used to study glycosylation processing (33). It should be noted that avidin itself is a glycoprotein that contains heterogeneous oligosaccharides and that its nonglycosylated recombinant version (NeutralLite) (34) is more suitable for the preparation of definitive neoglycoproteins.
Via enzymatic action
A transglutaminase reaction was found to be useful in the preparation of neoglycoproteins. For example, M-amino glycosides were used as a substrate for transglutaminase for the addition of carbohydrates to glutamine side chains (35, 36). The success of this reaction, however, seems somewhat unpredictable.
Some endo-β-hexosaminidase were found to be useful in neoglycoprotein preparation. Although the normal function of these enzymes is to cleave the bond between the two GlcNAc residues next to asparagines of the N-glycan attachment site, they can perform an en block transfer of oligosaccharides (e.g., Man9-GlcNAc) to a GlcNAc-terminated acceptor quite effectively under certain conditions (Fig. 2). Endo-A and endo-M have been used to transfer Man9-GlcNAc (endo-A) or a biantennary complex-type oligosaccharide (endo-M) to the GlcNAc-Asn site of glycoproteins, which can be generated by the hydrolytic action of the endo-β-hexosaminidase (37-39). Whereas endo-A is active only on high-mannose-type glycans, endo-M accepts complex-type glycans as a substrate as well, more effectively for biantennary than triantennary structures. Endo-M has been used for modification of eel calcitonin effectively (38, 39). In either case, the acceptor GlcNAc need not be attached to the Asn. GlcNAc itself and its glycosides are known good acceptors.
A very interesting and useful application of endo-A was reported recently by Wang and coworkers (40, 41). Based on the knowledge that some endo -glycosidase effectively can transfer oxazoline derivatives of GlcNAc-oligosaccharides, Wang and coworkers used endo-A on the oxazoline derivative of Man9-GlcNAc (and other similar oligosaccharides) to transfer the oligosaccharide in 80-90% yield (Fig. 2).

Figure 2. Illustration of an en block transfer of oligosaccharides by endo-A.
De Novo or Semisynthetic Preparation of Neoglycoprotein
The advancement in the techniques of the chemical synthesis of oligosaccharides and peptides (including molecular engineering) elevated the neoglycoprotein preparation to the de novo synthesis level, which opened a wide range of application possibilities.
Via incorporated Cys in the peptide
When a Cys can be incorporated into the peptide sequence by means of bioengineering, thioaldoses can be attached via a disulfide bond with the newly introduced SH group in the protein. In a recent example (29), asparagine (N-297) of IgG-Fc was replaced with Cys and its SH group was used to form a disulfide bond with thio-oligosaccharide (29). When 1-thio-derivative of Man3GlcNAc2 was attached to the thio-modified IgG by such a scheme, the Fc-mediated IgG potency increased several fold over that of the IgG that contains the natural Fc. The distance of glycan to the peptide backbone attached by this method is approximately the same distance as in the natural N-glycan.
Via native chemical ligation
The technique of native chemical ligation (NCL) or expressed protein ligation (EPL) (42) tremendously expanded the scope of peptide/protein synthesis. This approach also has been adopted for the construction of neoglycoproteins. For complete synthetic approaches to the preparation of defined glycoproteins, several excellent reviews (19, 43, 44) exist. The basic principle of NCL is illustrated in Fig. 3 (45). In this article, only a few examples are mentioned. Macmillan and Bertozzi (46) used Escherichia coli-expressed peptides and chemically synthesized glycopeptides to construct GlyCAM-1, a natural ligand for L-selectin. Similarly, Tolbert et al. reported chemical ligation of synthetic glycopeptides with E. coli expressed and TEV protease-cleaved peptides (19).

Figure 3. Illustration of ''native chemical ligation'' (45).
Chemoenzymatic synthesis
One prototypic example of chemoenzymatic synthesis is the preparation of “glycotentacles” (36). First, cyclic peptides that contain several glutamine residues were synthesized. Gal-terminated glycan chains were attached to the glutamine residues by transglutaminase reaction, which then was sialylated enzymatically with sialyltransferase to obtain NeuAc(α-2,3-)Gal terminated glycans. This approach provides flexibility in the size of the cyclic peptide and the choice of glycan structures. As mentioned earlier, the transglycosylation by endo-A was facilitated greatly when the reducing terminal GlcNAc was converted chemically to its oxazoline derivative first (41, 47). A synthetic mucin based on a poly (α-L-glutamic acid) backbone decorated with p-aminophenyl-glycoside of LacNAc followed by sialylation with sialyl transferase was prepared as an anti-influenza agent (48).
Although the use of glycosyl transferases require more expensive sugar nucleotides, the exquisite specificities offered by these enzymes sometimes override the cost concerns. Glyco- syltransferase often are used in terminal modifications such as sialylation and fucosylation.
Other techniques and reagents
The ease of “click chemistry” (49) has attracted many applications in the area of neoglycoconjugates. The principle of “click chemistry” is illustrated in Fig. 4. The conjugation based on click chemistry can be performed between either the ω-azido glycoside and an ω-alkyn group on a protein or vice versa (50). The popularity of “click chemistry” resides in its ease of operation, its indifference to reaction conditions (solvent, pH), and its high yields. A traceless Staudinger ligation was reported for a carbohydrate-protein conjugation (47).
A related reaction using the Diels-Adler cycloaddition of a diene-containing glycoside and a maleimide-equipped protein gives neoglycoproteins at ambient temperature in good yield (51). Preparation of “diene” glycosides may be the limiting step in this application.
Squaric acid esters (Fig. 5a) sometimes are used to link glycans to proteins (52, 54-56). N-Acetyl-chito-oligosaccharides were linked to silk fibroin by reacting solubilized silk fibroin with cyanuric chloride-activated oligosaccharides (Fig. 5b).
NMR analysis of the product indicate that Tyr and Lys were modified (53). A homobifunctionalp-nitrophenyl ester was proposed for the preparation of neoglycoproteins (26). In this method, one of the active ester groups is used to react with the ω-aminoalkyl glycoside and the product thereof then will be conjugated to a protein or other amino-bearing material.
A clever approach to prepare (by one-pot synthesis) pyroglutaminylated glycosylamine useful in constructing neoglycoconjugates was reported (57). The reducing end of an oligosaccharide first reacts with an a-amino-group of a glutaminyl derivative; then, the glycosylamine formed is stabilized by cyclization that involves the γ-carboxyl group. The reaction scheme is shown in Fig. 6.
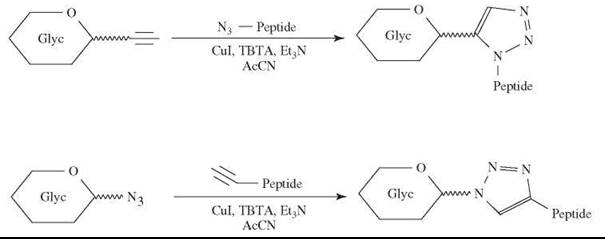
Figure 4. Principle of ''click chemistry'' applied to neoglycoprotein preparation.

Figure 5. Squaric acid (52) and cyanuric acid (53) as linkers in neoglycoproteins.
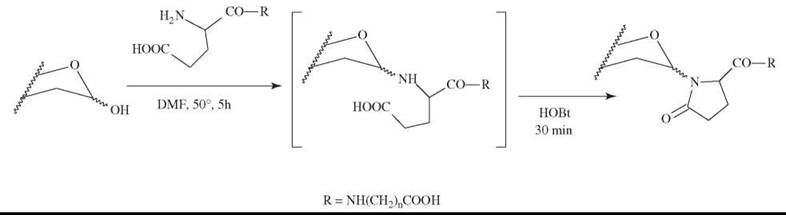
Figure 6. Preparation of glycoconjugate by one-pot reaction (57).
Applications of Neoglycoproteins
Cluster effect (or multivalency effect)
In addition to providing structurally well-defined glycans on a protein carrier, neoglycoproteins offer a platform for presenting multiple copies of glycans in a limited confine. The first indication of the effectiveness of the clustering of sugars on neoglycoprotein was observed in the study of neoglycoproteins binding by rabbit liver membrane (11). The binding of BSA-based neoglycoproteins that carry several different sugars by rabbit liver membrane revealed that Gal-BSA was bound very effectively but neoglycoproteins of other sugars (GlcNAc, Man, L-Fuc) were totally ineffective, which indicates that the sugar binding is specific for Gal. The more astounding finding was that when the number of Gal on BSA increased linearly, the binding affinity increased logarithmically, which resulted in a subnanomolar binding affinity (as KI) at the Gal content of about 40 residues per molecule of BSA. The monomeric Gal derivative had an affinity of only ca. mM in KI. This effect was demonstrated more dramatically by the synthetic oligosaccharide mimetics of natural Gal-terminated N-glycans of mono-, di-, and tri-antennary structures (58), in which a 2- and 3-fold increase in valency brought about a ca. 1000- and 1,000,000-fold increase, respectively, in the binding affinity. Other examples of cluster effect are binding intricately synthesized pentameric and decameric clusters of galabiose-(Galα4Gal-)containing structures to Shiga-like toxin (59) and Man-containing dendritic ligand to DC-SIGN (60). The cluster effect (or multivalency effect) now is applied widely to many carbohydrate-binding systems. A term AVIDITY (originally a serological term) sometimes is used to express the overall affinity of a ligand formed by the clustering of a monomeric ligand (61). The cluster effect is assessed quantitatively in a few publications (61-63).
Vaccines
Avery and Goebel (5, 6) and Tillett et al. (20) constructed their neoglycoproteins for raising vaccines against carbohydrate groups. The attachment of carbohydrates to proteins apparently made the carbohydrates more antigenic. The preparation and the use of neoglycoproteins for antibody generation is a very important endeavor (64). In fact, many vaccines currently in use (Heamophilus b conjugate, meningococcal 4-valent conjugate, and pneumococcal 7-valent conjugate) are neoglycoproteins in one form or another (65, 66). The antigenicity of dextran-chicken serum albumin was studied with respect to glycan size and density (67).
Lectin specificity and glycosyltransferase substrates
The groups of Monsigny and colleagues (7, 8) and Iyer and Goldstein (9) prepared neoglycoproteins for specificity studies of plant lectins (7, 8). The specificity studies later were extended to animal lectins (4, 68-70) with excellent results.
Neoglycoproteins can serve as substrates for glycosyltransferases in the chemo-enzymatic preparation of more complex glycan structures. In addition, neoglycoproteins are useful as substrates in the assay for glycosyltransferases. Examples are β-1,4-galactosyltransferases (71, 72) and a-fucosyltransferase (73).
Isolation Media and Probes
Many animal lectins have been isolated and purified using neoglycoprotein conjugated to agarose beads. C-type lectins are especially amenable to purification using neoglycoprotein-affinity gel because of the ease of elution simply by using EDTA-containing buffers. The examples include the use of GlcNAc-BSA-Sepharose for chicken hepatic lectin (74) and Man-BSA-Sepharose for alligator hepatic lectin (70).
Neoglycoproteins also are useful as probes for sugar-binding proteins. Fucose-specific adhesins on germ tubes of Candida albicans were demonstrated with fluorescein-labeled BSA derivatives bearing different sugars (75). More recent and elegant application was the use of de novo-designed glycoproteins to study the ER-quality control process as related to the activity of glucosyltransferase (76). T-antigen bearing neoglycoprotein was found to be useful as probes for breast carcinoma (77).
Glycoarrays
Microarrays of glycans (glycoarrays), which can be regarded as a special form of neoglycoconjugates, are proving to be a powerful tool for glycomics (78-81). Many above-mentioned conjugation methods and some other methods have been used in the construction of glycoarrays (82). For example, amino-terminated glycosides were reacted with carboxyl derivatives on a glass slide (amide formation) (83, 84) and reducing oligosaccharides were reacted with hydrazide (13, 85) or aminoxy groups (82) on glass. In the case of neoglycolipid (NGL), oligosaccharides were conjugating to 1,2-dihexadecyl-sn-glycero-3-phosphoethanolamine (DHPE) and the resulting NGL was adsorbed directly on a solid surface (86). A modified version of NGL uses aminoxy derivatives of DHPE (87). In a recent report, sulfation specificities of glycosaminoglycan interactions with growth factors and chemotactic proteins were probed with microarrays (88).
References
1. Spiro RG. Protein glycosylation: nature, distribution, enzymatic formation, and disease implications of glycopeptide bonds. Glycobiology 2002; 12:43R-56R.
2. Lee RT, Lee YC. Neoglycoproteins. In: Glycoproteins II. Montreuil J, Vliegenthart JFG, Schachter H, eds. 1997. Elsevier, Amsterdam, The Netherlands. pp. 601-620.
3. Kawaguchi K, Kuhlenschmidt M, Roseman S, Lee YC. Synthesis of some cluster galactosides and their effect on the hepatic galactose-binding system. Arch. Biochem. Biophys. 1980; 205: 388-395.
4. Connolly DT, Townsend RR, Kawaguchi K, Bell WR, Lee YC. Binding and endocytosis of cluster glycosides by rabbit hepatocytes. Evidence for a short-circuit pathway that does not lead to degradation. J. Biol. Chem. 1982; 257:939-945.
5. Avery OT, Goebel WF. Chemo-immunological studies on conjugated carbohydrate-proteins. II. Immunological specificity of synthetic sugar-protein antigens. J. Experim. Med. 1929; 50:533- 550.
6. Goebel WF, Avery OT. Chemo-immunological studies on conjugated carbohydrate-proteins. I. The synthesis of p-aminophenol b-glucoside, p-aminophenol b-galactoside, and their coupling with serum globulin. J. Experim. Med. 1929; 50:521-531.
7. Privat JP, Delmotte F, Monsigny M. Protein-sugar interactions. Association of wheat germ agglutinin (lectin) and O-(4-methyl-umbelliferyl)-glycosides. FEBS Lett. 1974; 46:229-232.
8. Privat JP, Delmotte F, Monsigny M. Protein-sugar interactions. Association of beta-(1 leads to 4) linked N-acetyl-D-glucosamine oligomer derivatives with wheat germ agglutinin (lectin). FEBS Lett. 1974; 46:224-228.
9. Iyer RN, Goldstein IJ. Quantitative studies on the interaction of concanavalin A, the carbohydrate-binding protein of the jack bean, with model carbohydrate-protein conjugates. Immunochemistry 1973; 10:313-322.
10. Buss DH, Goldstein IJ. Protein-carbohydrate interaction. XIV. Carbohydrates containing groups for the alkylation of proteins. J. Chem. Soc. 1968; 12:1457-1461.
11. Krantz MJ, Holtzman NA, Stowell CP, Lee YC. Attachment of thioglycosides to proteins: enhancement of liver membrane binding. Biochemistry. 1976; 15:3963-3968.
12. Lee YC, Stowell CP, Krantz MJ. 2-Imino-2-methoxyethyl 1-thioglycosides: new reagents for attaching sugars to proteins. Biochemistry 1976; 15:3956-3963.
13. Lee RT, Lee YC. Preparation of neoglycoproteins using w-aldehydoalkyl 1-thioglycosides. Meth. Enzymol. 1982; 83:289-294.
14. Stowell CP, Lee YC. Preparation of neoglycoproteins using 2-imino-2-methoxyethyl 1-thioglycosides. Meth. Enzymol. 1982; 83:278-288.
15. Gray GD, Zerylnick M, Davis HN, Dewsbury DA. Effects of variations in the male copulatory behavior on ovulation and implantation in prairie voles, Microtus ochrogaster. Horm. Behav. 1974; 5:389-396.
16. Gray GR, Schwartz BA, Kamicker BJ. Proteins containing reductively aminated disaccharides: chemical and immunochemical characterization. Prog. Clin. Biol. Res. 1978; 23:583-594.
17. Baues RJ, Gray GR. Lectin purification on affinity columns containing reductively aminated disaccharides. J. Biol. Chem. 1977; 252:57-60.
18. Lee RT, Wong TC, Lee R, Yue L, Lee YC. Efficient coupling of glycopeptides to proteins with a heterobifunctional reagent. Biochemistry 1989; 28:1856-1861.
19. Tolbert TJ, Franke D, Wong CH. A new strategy for glycoprotein synthesis:ligation of synthetic glycopeptides with truncated proteins expressed in E. coli as TEV protease cleavable fusion protein. Bioorg. Med. Chem. 2005; 13:909-915.
20. Tillett WS, Avery OT, Goebel WF. Chemo-immunological studies on conjugated carbohydrate-proteins. III. Active and passive anaphylaxis with synthetic sugar-proteins. J. Experimen. Med. 1929; 50:551-567.
21. Chen T, Vazquez-Duhalt R, Wu CF, Bentley WE, Payne GF. Combinatorial screening for enzyme-mediated coupling. Tyrosinase-catalyzed coupling to create protein-chitosan conjugates. Biomacromolecules 2001; 2:456-462.
22. Arakatsu Y, Ashwell G, Kabat EA. Immunochemical studies on dextrans. V. Specificity and cross-reactivity with dextrans of the antibodies formed in rabbits to isomaltonic and isomaltotrionic acids coupled to bovine serum albumin. J. Immunol. 1966; 97:858-866.
23. Lonngren J, Goldstein IJ, Niederhuber JE. Aldonate coupling, a simple procedure for the preparation of carbohydrate-protein conjugates for studies of carbohydrate-binding proteins. Arch. Biochem. Biophys. 1976; 175:661-669.
24. Lemieux RU, Bundle DR, Baker DA. The properties of a “synthetic” antigen related to the human blood-group Lewis a. J. Am. Chem. Soc. 1975; 97:4076-4083.
25. Porro M, Costantino P, Viti S, Vannozzi F, Naggi A, Torri G. Specific antibodies to diphtheria toxin and type 6A pneumococcal capsular polysaccharide induced by a model of semi-synthetic glycoconjugate antigen. Mol. Immunol. 1985; 22:907-919.
26. Wu X, Ling C-C, Bundle DR. A new homobifunctional p-nitrophenyl ester coupling reagent for the preparation of neoglycoproteins. Organic Lett. 2004; 6:4407-4410.
27. Watt GM, Boons GJ. A convergent strategy for the preparation of N-glycan core di-, tri-, and pentasaccharide thioaldoses for the site-specific glycosylation of peptides and proteins bearing free 44. cysteines. Carbohydr Res. 2004; 339:181-193.
28. Caramelo JJ, Castro OA, Alonso LG, De Prat-Gay G, Parodi AJ. UDP-Glc:glycoprotein glucosyltransferase recognizes structured and solvent accessible hydrophobic patches in molten globule-like folding intermediates. Proc. Natl. Acad. Sci. U.S.A. 46. 2003; 100:86-91.
29. Watt GM, Lund J, Levens M, Kolli VS, Jefferis R, Boons GJ. Site-specific glycosylation of an aglycosylated human IgG1-Fc antibody protein generates neoglycoproteins with enhanced function. Chem. Biol. 2003; 10:807-814.
30. Macindoe WM, van Oijen AH, Boons G-J. A unique and highly facile method for synthesizing disulfide linked neoglycoconjugates: a new approach for remodeling of peptides and proteins. Chem. Commun. 1998: 847-848.
31. Chen VJ, Wold F. Neoglycoproteins: preparation of noncovalent glycoproteins through high-affinity protein-(glycosyl) ligand complexes. Biochemistry 1984; 23:3306-3311.
32. Chen VJ, Wold F. Neoglycoproteins: preparation and properties of complexes of biotinylated asparagine-oligosaccharides with avidin and streptavidin. Biochemistry 1986; 25:939-944.
33. Shao MC, Wold F. The use of avidin-biotinylglycan as the model for in vitro glycoprotein processing. J Biol Chem. 1987; 262:2968-2672.
34. Marttila AT, Laitinen OH, Airenne KJ, Kulik T, Bayer EA, Wilchek M, Kulomaa MS. Recombinant NeutraLite avidin: a non-glycosylated, acidic mutant of chicken avidin that exhibits high affinity for biotin and low non-specific binding properties. FEBS Lett. 2000; 467:31-36.
35. Yan SB, Wold F. Neoglycoproteins: in vitro introduction of glycosyl units at glutamines in beta-casein using transglutaminase. Biochemistry 1984; 23:3759-3765.
36. Ohta T, Miura N, Fujitani N, Nakajima F, Niikura K, Sadamoto R, Guo CT, Suzuki T, Suzuki Y, Monde K, Nishimura S. Glycotentacles: synthesis of cyclic glycopeptides, toward a tailored blocker of influenza virus hemagglutinin. Angew Chem Int Ed Engl. 2003; 42:5186-5189.
37. Takegawa K, Tabuchi M, Yamaguchi S, Kondo A, Kato I, Iwahara S. Synthesis of neoglycoproteins using oligosaccharide-transfer activity with endo-beta-N-acetylglucosaminidase. J Biol Chem. 1995; 270:3094-3099.
38. Haneda K, Inazu T, Mizuno M, Iguchi R, Yamamoto K, Kumagai H, Aimoto S, Suzuki H, Noda T. Chemo-enzymatic synthesis of calcitonin derivatives containing N-linked oligosaccharides. Bioorg. Med. Chem. Lett. 1998; 8:1303-1306.
39. Haneda K, Inazu T, Yamamoto K, Kumagai H, Nakahara Y, Kobata A. Transglycosylation of intact sialo complex-type oligosaccharides to the N-acetylglucosamine moieties of glycopeptides by Mucor hiemalis endo-beta-N-acetylglucosaminidase. Carbohydr Res. 1996; 292:61-70.
40. Li B, Song H, Hauser S, Wang LX. A highly efficient chemoenzymatic approach toward glycoprotein synthesis. Org. Lett. 2006; 8:3081-3084.
41. Zeng Y, Wang J, Li B, Hauser S, Li H, Wang LX. Glycopeptide synthesis through endo-glycosidase-catalyzed oligosaccharide transfer of sugar oxazolines: probing substrate structural requirement. Chemistry. 2006; 12:3355-3364.
42. Mills KV, Perler FB. The mechanism of intein-mediated protein splicing: variations on a theme. Protein Pept. Lett. 2005; 12:751- 755.
43. Pratt MR, Bertozzi CR. Synthetic glycopeptides and glycoproteins as tools for biology. Chem.Soc. Rev. 2005; 34:58-68.
44. Brik A, Ficht S, Wong CH. Strategies for the preparation of homogenous glycoproteins. Curr. Opin. Chem. Biol. 2006; 10:638- 644.
45. Buskas T, Ingale S, Boons GJ. Glycopeptides as versatile tools for glycobiology. Glycobiology 2006; 16:113R-136R.
46. Macmillan D, Bertozzi CR. Modular Assembly of Glycoproteins: Towards the Synthesis of GlyCAM-1 by using expressed protein ligation. Angewandte Chemie Int. Ed. 2004; 43:1355-1359.
47. Grandjean C, Boutonnier A, Guerreiro C, Fournier JM, Mulard LA. On the preparation of carbohydrate-protein conjugates using the traceless Staudinger ligation. J. Org. Chem. 2005; 70:7123-7132.
48. Totani K, Kubota T, Kuroda T, Murata T, Hidari KI, Suzuki T, Suzuki Y, Kobayashi K, Ashida H, Yamamoto K, Usui T. Chemoenzymatic synthesis and application of glycopolymers containing multivalent sialyloligosaccharides with a poly(L-glutamic acid) backbone for inhibition of infection by influenza viruses. Glycobiology 2003; 13:315-326.
49. Lewis WG, Green LG, Grynszpan F, Radic Z, Carlier PR, Taylor P, Finn MG, Sharpless KB. Click chemistry in situ: acetylcholinesterase as a reaction vessel for the selective assembly of a femtomolar inhibitor from an array of building blocks. Angewandte Chemie Int. Ed. 2002; 41:1053-1057.
50. Groothuys S, Kuijpers BHM, Quaedflieg PJLM, Roelen HCPF, Wiertz RW, Blaauw RH, van Delft FL, Rutjes FPJT. Chemoenzymatic synthesis of triazole-linked glycopeptides. Synthesis 2006: 3146-3152.
51. Pozsgay V, Vieira NE, Yergey A. A method for bioconjugation of carbohydrates using Diels-Alder cycloaddition. Org. Lett. 2002; 4:3191-3194.
52. Saksena R, Ma X, Kovac P. One-pot preparation of a series of glycoconjugates with predetermined antigen-carrier ratio from oligosaccharides that mimic the O-PS of Vibrio cholerae O:1, serotype Ogawa. Carbohydr Res. 2003; 338:2591-2603.
53. Ma X, Saksena R, Chernyak A, Kovac P. Neoglycoconjugates from synthetic tetra- and hexasaccharides that mimic the terminus of the O-PS of Vibrio cholerae O:1, serotype Inaba. Org. Biomol. Chem. 2003; 1:775-784.
54. Chernyak A, Karavanov A, Ogawa Y, Kovac P. Conjugating oligosaccharides to proteins by squaric acid diester chemistry: rapid monitoring of the progress of conjugation, and recovery of the unused ligand. Carbohydr. Res. 2001; 330:479-486.
55. Nagahori N, Lee RT, Nishimura S, Page D, Roy R, Lee YC. Inhibition of adhesion of type 1 fimbriated Escherichia coli to highly mannosylated ligands. ChemBioChem. 2002;3: 836-844.
56. Gotoh Y, Tsukada M, Aiba S, Minoura N. Chemical modification of silk fibroin with N-acetyl-chito-oligosaccharides. Int. J. Biol. Macromol. 1996; 18:19-26.
57. Quetard C, Bourgerie S, Normand-Sdiqui N, Mayer R, Strecker G, Midoux P, Roche AC, Monsigny M. Novel glycosynthons for glycoconjugate preparation: oligosaccharylpyroglutamylanilide derivatives. Bioconjug. Chem. 1998; 9:268-76.
58. Lee YC, Townsend RR, Hardy MR, Lonngren J, Arnarp J, Haraldsson M, Lonn H. Binding of synthetic oligosaccharides to the hepatic Gal/GalNAc lectin. Dependence on fine structural features. J. Biol. Chem. 1983; 258:199-202.
59. Kitov PI, Sadowska JM, Mulvey G, Armstrong GD, Ling H, Pannu NS, Read RJ, Bundle DR. Shiga-like toxins are neutralized by tailored multivalent carbohydrate ligands. Nature 2000; 403:669-672.
60. Frison N, Taylor ME, Soilleux E, Bousser MT, Mayer R, Monsigny M, Drickamer K, Roche AC. Oligolysine-based oligosaccharide clusters: selective recognition and endocytosis by the mannose receptor and dendritic cell-specific intercellular adhesion molecule 3 (ICAM-3)-grabbing nonintegrin. J. Biol. Chem. 2003; 278:23922-237929.
61. Mammen M, Chio S-K, Whitesides GM. Polyvalent interactions in biological systems: implications for design and use of multivalent ligands and inhibitors. 1998; 37:2754-2794.
62. Lundquist JJ, Toone EJ. The cluster glycoside effect. Chem. Rev. 2002; 102:555-578.
63. Kitov PI, Bundle DR. On the nature of the multivalency effect: a thermodynamic model. J. Am. Chem. Soc. 2003; 125:16271-16284.
64. Willats WG, Marcus SE, Knox JP. Generation of monoclonal antibody specific to (1 → 5)-alpha-L-arabinan. Carbohydr. Res. 1998; 308:149-152.
65. Benaissa-Trouw B, Lefeber DJ, Kamerling JP, Vliegenthart JF, Kraaijeveld K, Snippe H. Synthetic polysaccharide type 3-related di-, tri-, and tetrasaccharide-CRM(197) conjugates induce protection against Streptococcus pneumoniae type 3 in mice. Infect Immun. 2001; 69:4698-4701.
66. Ada G, Isaacs D. Carbohydrate-protein conjugate vaccines. Clin. Microbiol. Infect. 2003; 9:79-85.
67. Seppala I, Makela O. Antigenicity of dextran-protein conjugates in mice. Effect of molecular weight of the carbohydrate and comparison of two modes of coupling. J. Immunol. 1989; 143:1259-1264.
68. Shepherd VL, Lee YC, Schlesinger PH, Stahl PD. L-Fucoseterminated glycoconjugates are recognized by pinocytosis receptors on macrophages. Proc. Natl. Acad. Sci. U.S.A. 1981; 78:1019- 1022.
69. Lehrman MA, Pizzo SV, Imber MJ, Hill RL. The binding of fucose-containing glycoproteins by hepatic lectins. Re-examination of the clearance from blood and the binding to membrane receptors and pure lectins. J. Biol. Chem. 1986; 261:7412-7418.
70. Lee RT, Yang GC, Kiang J, Bingham JB, Golgher D, Lee YC. Major lectin of alligator liver is specific for mannose/L-fucose. J. Biol. Chem. 1994; 269:19617-19625.
71. Zatta PF, Nyame K, Cormier MJ, Mattox SA, Prieto PA, Smith DF, Cummings RD. A solid-phase assay for beta-1,4-galactosyltransferase activity in human serum using recombinant aequorin.Anal. Biochem. 1991; 194:185-191.
72. Abdul-Rahman B, Ailor E, Jarvis D, Betenbaugh M, Lee YC. Beta-(1 → 4)-Galactosyltransferase activity in native and engineered insect cells measured with time-resolved europium fluorescence. Carbohydr Res. 2002; 337:2181-2186.
73. Rabina J, Smithers N, Britten CJ, Renkonen R. A time-resolved immunofluorometric method for the measurement of sialyl Lewis x-synthesizing alpha1,3-fucosyltransferase activity. Anal. Biochem. 1997; 246:71-78.
74. Kuhlenschmidt TB, Lee YC. Specificity of chicken liver carbohydrate binding protein. Biochemistry 1984; 23:3569-3575.
75. Vardar-Unlu G, McSharry C, Douglas LJ. Fucose-specific adhesins on germ tubes of Candida albicans. FEMS Immunol Med. Microbiol. 1998; 20:55-67.
76. Caramelo JJ, Castro OA, de Prat-Gay G, Parodi AJ. The endoplasmic reticulum glucosyltransferase recognizes nearly native glycoprotein folding intermediates. J. Biol. Chem. 2004; 279:46280- 46285.
77. Gabius HJ, Schroter C, Gabius S, Brinck U, Tietze LF. Binding of T-antigen-bearing neoglycoprotein and peanut agglutinin to cultured tumor cells and breast carcinomas. J. Histochem. Cytochem. 1990; 38:1625-1631.
78. Raman R, Raguram S, Venkataraman G, Paulson JC, Sasisekharan R. Glycomics: an integrated systems approach to structure-function relationships of glycans. Nat. Methods. 2005; 2:817-824.
79. Turnbull JE, Field RA. Emerging glycomics technologies. Nat. Chem. Biol. 2007; 3:74-77.
80. Horlacher T, Seeberger PH. The utility of carbohydrate microarrays in glycomics. Omics. 2006; 10:490-498.
81. Alvarez RA, Blixt O. Identification of ligand specificities for glycan-binding proteins using glycan arrays. Methods Enzymol. 2006; 415:292-310.
82. Zhou X, Zhou J. Oligosaccharide microarrays fabricated on aminooxyacetyl functionalized glass surface for characterization of carbohydrate-protein interaction. Biosensors Bioelectron. 2006; 21:1451-1458.
83. Blixt O, Head S, Mondala T, Scanlan C, Huflejt ME, Alvarez R, Bryan MC, Fazio F, Calarese D, Stevens J, Razi N, Stevens DJ, Skehel JJ, van Die I, Burton DR, Wilson IA, Cummings R, Bovin N, Wong CH, Paulson JC. Printed covalent glycan array for ligand profiling of diverse glycan binding proteins. Proc. Natl. Acad. Sci. U.S.A. 2004; 101:17033-17038.
84. Noti C, de Paz JL, Polito L, Seeberger PH. Preparation and use of microarrays containing synthetic heparin oligosaccharides for the rapid analysis of heparin-protein interactions. Chem. A Euro. J. 2006; 12:8664-8686.
85. Zhi Z-L, Powell AK, Turnbull JE. Fabrication of carbohydrate microarrays on gold surfaces: direct attachment of nonderivatized oligosaccharides to hydrazide-modified self-assembled monolayers. Anal. Chem. 2006; 78:4786-4793.
86. Fukui S, Feizi T, Galustian C, Lawson AM, Chai W. Oligosaccharide microarrays for high-throughput detection and specificity assignments of carbohydrate-protein interactions. Nat. Biotechnol. 2002; 20:1011-1017.
87. Liu Y, Chai W, Childs RA, Feizi T. Preparation of neoglycolipids with ring-closed cores via chemoselective oxime-ligation for microarray analysis of carbohydrate-protein interactions. Methods Enzymol. 2006; 415:326-340.
88. Shipp EL, Hsieh-Wilson LC. Profiling the sulfation specificities of glycosaminoglycan interactions with growth factors and chemotactic proteins using microarrays. Chem. Biol. 2007; 14:195-208.
Further Reading
Lee RT, Lee YC. Neoglycoproteins. In: Glycoproteins II. Montreuil J, Vliegenthart JFG, Schachter H, eds. 1997. Elsevier, Amsterdam, The Netherlands. pp. 601-620.
Lee YC, Lee RT. Neoglycoconjugates. Preparation and Applications. 1994. Academic Press, San Diego, CA.
Lee YC, Lee RT. Neoglycoproteins. In: Lennarz W, Lane D, eds. Encyclopedia of Biochemistry. Vol. 2. 2004. Elsevier, Amsterdam, The Netherlands. pp. 11-15.
Monsigny M, Roche A-C, Duverger E, Srinivas O. Multivalent presentation: Neoglycoproteins in Comprehensive Glycoscience, Section D. Kamerlng A, Suguzi S, Taniguchi YC, Lee Boons GJ, Voragen F, eds. In press.
Pratt MR, Bertozzi CR. Synthetic glycopeptides and glycoproteins as tools for biology. Chem. Soc. Rev. 2005; 34:58-68.
Doores KJ, Gamblin DP, Davis BG. Exploring and exploiting the therapeutic potential of glycoconjugates. Chem. Eur. J. 2006; 12:656-665.
See Also
Glycan Synthesis, Key Strategies for
Glycan Therapeutics, Engineering of
Glycans, Chemistry of
Glycoengineering
Glycopeptides and Glycoproteins, Synthesis of