CHEMICAL BIOLOGY
Fluorescence Techniques for Nucleic Acids
Ulysse Asseline, Centre de Biophysique Moleculaire UPR 4301 CNRS, Orleans, France
doi: 10.1002/9780470048672.wecb177
The power of fluorescence spectroscopy in the study of nucleic acids relies on the availability of hundreds of different fluorophores that span the visible spectrum of light and the possibility of using them to label oligonucleotides (short fragments of DNA or RNA obtained by chemical synthesis) specifically. Any sequence can be obtained with one or more fluorophores and fluorophore quenchers attached at preselected positions by linkages with suitable parameters to ensure the optimal properties for the fluorescent oligonucleotides. Fluorescent labeling of long DNA or RNA fragments is also possible by enzymatic methods. Fluorescence-based studies of nucleic acids are widely used in different areas such as basic research, molecular and clinical diagnostics, disease monitoring, therapeutic development, food technology, environmental sciences, and biotechnology. After a few words on biological and fluorescence backgrounds, the main labeling strategies will be developed and examples of biological questions that can be addressed with fluorescent oligonucleotides and the major types of fluorescence spectroscopy techniques will be reported.
Fluorescence-based studies of nucleic acids (NAs) rely on two key contributions of synthetic chemistry: the automatization of oligonucleotide (ON) syntheses (1) and the development of efficient methods for site-specific fluorescent labeling of these ONs (2-7). Many applications are based on the complementary hybridization between short fluorescent synthetic ONs (15-25 nucleotides in length) and the NA to be analyzed. ON sequences can also be chosen for assembling into structures that mimic those found in living biological systems, which provides tools for structural, dynamic, and interaction studies. Because the intrinsic fluorescence emission of the major nucleic bases is too weak to be used for detection, fluorescence-based studies have to rely on synthetically modified nucleotides with more desirable emissive characteristics or exogenous fluorescent labels (Fs). It is a challenge for the chemist to develop ON analogs that do not perturb the samples to be analyzed while providing the desired spectroscopic properties. Fluorescence-based techniques can be used both in solution and in solid-phase applications. In the last few years, the development of fluorescent ONs (FONs) that show a modified emission in the presence of the target has been the focus of intense research (2-7). These new FON probes simplify the analyses in vitro and provide the possibility of applications in living organisms. New strategies for specific enzymatic labeling of large NAs are also in development. Many reviews have been published over the last 3 years on the use of fluorescence to study NAs (2-7). After a brief account on biological and fluorescence backgrounds, this article will focus on the main strategies used to label ONs with Fs and examples will be given of biological questions that can be addressed with FON using the major fluorescence parameters.
Background
Biological background
The sequencing of the human genome provides the full genetic map of the NAs of a human being but lacks information as to how they are involved in the processes within cells. Analysis of the interindividual sequence variations in healthy and diseased people can help to identify and understand inherited or acquired pathologies as well as drug side effects (8). The ability to detect, localize, and quantify the different RNAs (pre-mRNA, mRNA, micro RNA) in living cells and tissues, in real time, will offer tremendous opportunities for biological and disease studies, and it will have a significant impact on medical diagnostics and drug discovery (8, 9). To understand RNA functions in the cell, it is also necessary to monitor their conformational changes directly during pre-mRNA splicing, ribosome assembly, and tRNA processing. These different biological questions can be addressed with different ON probe structures conveniently labeled with selected Fs that exploit the main characteristics of fluorescence.
Table 1. The physical properties of the main fluorophores and fluorescent quenchers reported in this article
|
Fluorescent labels |
λexc (nm) |
λem (nm) |
References |
|
Xanthene dyes FAM |
494 |
517 |
3, 4, 15, 16, 17, 18, 19, 20, 21, 22, 23, 25 |
|
TAMRA |
555 |
577 |
15, 17, 18, 26, 20, 23, 27, 24, 25, 33 |
|
Cyanine dyes Cy3.5 |
582 (550) |
593 (570) |
15 |
|
Cy5 |
650 |
670 |
15, 19, 21, 22, 27, 28 |
|
Thiazole orange |
480 |
530 |
29, 30, 31, 32, 33 |
|
Alexa Fluor 546 |
556 |
570 |
34 |
|
Polycarbocyclic dyes Pyrene |
240, 345 |
375, 395, 480 |
2, 4, 5, 7, 16, 35, 36 |
|
Perylene |
444 |
461, 487 |
4, 5, 16 |
|
Triphenyl methane dye Malachite green |
610 nm |
648 nm |
37 |
|
Quenchers |
λabs(nm) |
|
References |
|
TAMRA |
540 |
|
15 |
|
Dabcyl |
474 |
|
15, 36, 18, 19, 20, 22, 25 |
|
BHQ-1 |
534 |
|
21 |
|
Metal |
|
|
26 |
Fluorescence background
Emission of light, which occurs when an F excited to the singlet state relaxes back to the ground state, can be characterized by parameters such as emission spectrum, fluorescence intensity (enhancement or quenching), fluorescence lifetimes, and anisotropy (10). The fluorescence exhibited by a given F depends on its intrinsic properties, its exposure to solvents, as well as its interaction with other environmental elements. In our applications, the F can interact with the labeled ON. Interactions of the FON with its target sequence or with other nucleic acid binding molecules induce additional contacts of the F leading to quenching or enhancement of the fluorescence. The fluorescent signal can also be modified by the use of more than one F (identical or different) or by a suitable fluorophore/fluorescence quencher (F/Q) pairs linked at convenient positions on the ONs. When two different Fs are in close proximity and the emission spectrum of the donor overlaps the excitation spectrum of the acceptor, energy is transferred from donor F to acceptor F in a distance-dependent manner that follows a nonradiative process (Forster). Another strategy to induce changes of the fluorescent signal consists in the use of modified fluorescent ONs that can adopt specific structures prone to changes during interaction with their targets. When two or more aromatic hydrocarbon molecules such as pyrene are in close proximity, an excited-state pyrene monomer and ground-state monomer can form an excimer state that fluoresces at a longer wavelength than the monomer emission (Birks). Fluorescence resonance energy transfer (FRET) and excimer formation coupled with quenching/unquenching strategies are the most frequently used fluorescence parameters because of the ease of their usage. Fluorescence lifetime and anisotropy measurements require more expertise and sophisticated equipment. Fluorescence detection and measurements can be performed with various instruments (spectrofluorometers, micro plate readers, scanners, microscopes, and flow cytometers) that provide different pieces of information. The possibility of detection at different wavelengths associated with the suitable covalent fluorescent labeling of ONs provides the possibility of multiplexing. Finally, progress made in the field of instrumentation associated with the development of very sensitive Fs allows detection at the single-molecule level (11).
Nucleic Acids Labeling Strategies
Fluorescence labeling of NAs relies on chemical or enzymatic methods. However, intrinsic fluorescence emissions of the five most frequent natural nucleobases is too weak to be used for detection, and fluorescent labeling relies on synthetically modified nucleotides or exogenous Fs incorporated at convenient positions inside the ONs or NA sequences to provide the required spectroscopic properties.
Different classes of Fs and F quenchers (Qs)
Fs used to detect and study NAs involve nucleoside analogs, organic dyes, metal complexes, nanoparticles, and quantum dots (2-7). These Fs cover much of the visible spectrum and are available from many suppliers (12-14) (Table 1) (15-37). Several Qs are also commercially available. A very useful study on F/Q pair efficiencies has been reported (15). Researchers are also continuously developing Fs and Qs with spectroscopically tuned properties to fit the requirements of new applications into different formats and new detection instruments (2-5).
Different chemical labeling strategies
Nonspecific labeling
Many organic dyes F bind to NAs by electrostatic and hydrophobic (including intercalation) interactions that are not specific to the NA sequence. They provide information about the amount of NA available in a sample. These applications mainly include staining of the NA in polyacrylamide and agarose gels as well as DNA quantification during real-time PCR or cell labeling. These compounds, which are available from many suppliers, include SYBR Green, ethidium bromide, and YO-Pro-1.
Specific labeling
The selective chemical covalent labeling of ONs (DNA and RNA) can be achieved by either the incorporation of modified fluorescent nucleosides or the covalent attachment of Fs (and Qs) at preselected positions on the ON (2-5, 16). Fluorescent nucleotide analogs include either modified bases (isomorphic base analogs, purine analogs, extended base analogs, natural bases conjugated to Fs, etc.), or base replacement (polycarbo-cyclic Fs...) (2-5, 16, 38) (Fig. 1). These analogs can be incorporated via their phosphoramidite or H-phosphonate derivatives during the ON synthesis on solid phase. The phosphoramidite derivatives of the most common organic Fs (fluorescein, rhodamine, a few cyanines), Q (dabcyl...), and nucleosides with pyrene and perylene pendant groups as well as functionalized supports that allow the incorporation of the Fs and Qs at the 3'-end of the ONs can be obtained from different suppliers (12-14). However, the direct incorporation of the Fs during the ON synthesis requires that the F or nucleoside analogs can withstand the chemical conditions needed for the deprotection step and that they are soluble in organic solvents. Another strategy consists in the postsynthetic attachment of Fs to various positions of ONs by specific reactions between convenient functional groups incorporated at preselected positions of the ONs and in the Fs. Many Fs with groups that can react with the amino or thiol functionalized linker incorporated into ONs are commercially available (12, 13). The preparation of FONs with new specific properties can be achieved by the refinement of the F structures together with the development of new linkage parameters. Commonly used positions are the 5'- or 3'-termini (3, 4, 15, 17, 18, 20-22, 25, 31, 32, 34-36, 39-41), internucleotidic position (29, 30, 32) and the 2'-position of the sugar residue (3 and 4, positioning the F in the minor groove) (Fig. 2). It is also possible to attach Fs to nucleobases typically at the 5-position of T(U) or C (positioning the F in the major groove), the 7-C carbon of 7-deazaadenine or 7-deazaguanine, and the 8-position of adenine or guanine (2-5, 16, 42 and 43). In addition to the amide, thiourea, and thioether bonds, other linkages such as amine, phosphoramidate, oxime, hydrazone, phospho- thiolodiester, triazole, squarate, or disulfide can be used. The coupling reactions can take place in different solvents, aqueous buffers, or aqueous buffers/organic solvent mixtures depending on the F solubility (17, 18, 29-32, 35, 36, 39). It is also possible to perform the coupling steps directly in methanol in the presence of crown-ether to solubilize the deprotected ON (31).
The noncovalent specific labeling of NA probes based on the aptamer strategy has also been reported. The malachite green (37) aptamer, which was designed as a binary probe, assembles only in the presence of the target sequences and induces an increase in the intensity of the signal emitted by the F bound aptamer.
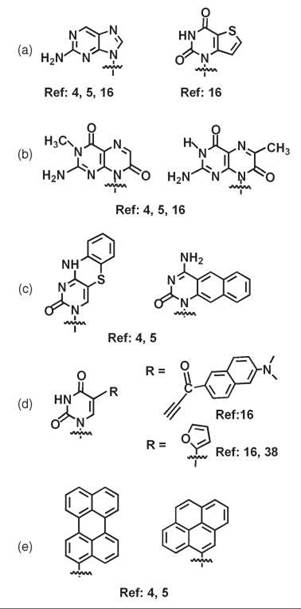
Figure 1. A selection of modified fluorescent nucleoside analogs. (a) Isomorphic base analogs. (b) Purine analogs. (c) Extended base analogs. (d) Natural bases conjugated to Fs. (e) Base replacements.

Figure 2. "The most frequently used positions for the covalent attachment of Fs onto the oligonucleotides."
Multilabeling
The chemical strategy allows the positioning of Fs at many selected sites along the ON. When several Fs are involved, they must be linked to the ON probes in positions that enable the most efficient properties required [FRET (17, 39), excimer (35)]. Tunable intensities of the excimer dependent on the pyrene number can also be observed (36).
Hybridization formats
The design of hybridization formats that enable important changes (fluorescence intensity increase or wavelength shifts) in the presence of the targets is the focus of intense research because their use can simplify the analyses in vitro and provide the possibility of applications in living organisms. The most frequently used involve binary probes (35), competitive hybridization probes (18), linear probes with only one F (2, 5, 16, 29, 31), and molecular beacons (MBs) (39-41), (Fig. 3).

Figure 3. Schematic representation of the main hybridization formats. (a) Binary probes. (b) Competitive hybridization probes. (c) Linear probe with one F. (d) Molecular Beacon. (e) Aptamer.
Enzymatic labeling
The enzymatic incorporation of modified fluorescent nucleosides or analogs via their triphosphate derivatives is also possible using DNA polymerases (3). One of the most important examples of applying this labeling strategy concerns the methods for NAs sequencing (42). More recently, a DNA sequencing method by synthesis on a solid surface has been proposed (43). This method is based on the four nucleotide 5'-triphosphates, which each contain a unique F with a distinct fluorescence emission at the base, modified at their 3' position to be reversible terminators. This method has yielded the Solexa sequencing technology (44). However, the sequence-specific internal labeling of large DNA require the development of other strategies as for example the use of DNA methyltransferase associated with a cofactor labeled with a F (45). A new strategy for the site-specific labeling of DNA and RNA is based on the expansion of the genetic alphabet by unnatural base pairs (46).
Selected Fluorescence Techniques and Applications
Different biological questions can be addressed using different ON probe structures conveniently labeled with a selected F that exploits the main characteristics of fluorescence, which is the most sensitive spectroscopic technique (10). Their definitions are widely reported in companion articles and will not be discussed here. The applications can be divided into different series. One application concerns synthetic FONs that mimic the different structures found in living systems and aims at structural, dynamics, and interaction studies in vitro. Another series includes mainly molecule and clinical diagnostic studies performed with native DNA targets. However, the amount of the DNA sequence available is in general very small, and the amplification of the NA acid target by PCR is necessary prior to detection. A comprehensive review concerning the PCR methods has recently been published (3). The last group involves studies in living cells mainly devoted to the detection of the different RNA families. A selection of examples of applications of the fluorescent oligonucleotides reported in this article is listed in the Table 2.
Table 2. A selection of applications of the fluorescent oligonucleotides reported in this article
|
Applications |
Hybridization formats |
F(s), Q(s), positions |
Fluorescent techniques |
References |
|
SNP analysis in homogeneous solutions |
Monolabeled uniprobes |
Fluorescent nucleosides in opposite position to the interrogation position (internal position of duplexes) |
Fluorescent sensing |
2 |
|
SNP analysis with genomic DNA |
Monolabeled uniprobes |
F at the 5'-end + free minor groove binder F |
FRET |
34 |
|
Improving specificity of |
Binary aptamer |
Noncovalent labeling |
Fluorescent sensing |
37 |
|
sequence detection |
||||
|
Detection of terminal |
Monolabeled uniprobes |
F at the 5'-end |
Fluorescence sensing |
31 |
|
mismatches |
||||
|
Detection of abasic site |
Monolabeled uniprobes |
Modified nucleoside in opposite position to the interrogation position (internal position of duplexes) |
Fluorescence sensing |
38 |
|
Sequencing by synthesis |
Templeted enzymatic incorporation |
F attached to bases of nucleoside triphosphates |
Fluorescence sensing, multicolour detection |
43 |
|
Pathogen detection (multiplex) |
MB on beads |
F/Q pairs at both ends |
Fluorescence sensing multicolour detection |
20, 21 |
|
Virus localisation in living cells |
MB |
F/Q pair at both ends |
Fluorescence sensing |
25 |
|
Detection of RNAs In living cells |
Autoligating Binary probes Uniprobe |
Two F and one Q (at selected positions) MB or MB pair F at internal position |
FRET, flow cytometry and confocal microscopy |
19, 29, 40-42, |
|
Visualization of mRNA in cellular extracts |
Binary probes |
Two Pyrene. One at the 5'-end of one ON probe and the second at the 3'-end of the second ON probe |
Excimer detection Time-resolved microscopy |
35 |
In vitro applications
Single-nucleotide polymorphism (SNP) analysis by fluorescence sensing and FRET
Most SNP genotyping assays detectable by fluorescence can be separated into four groups based on molecular mechanisms: primer extension, oligonucleotide ligation, invasive cleavage, and allele-specific hybridization (3, 47). Examples of specific hybridization applications are reported. Methods based on variations of the emitted light of nucleoside analogs inserted in the position opposite the mutation spot on the NA sequence to be analyzed have attracted a great deal of attention in the past few years. Most of these are based on the use of size-expanded base-discriminating fluorescent nucleosides or pyrene-labeled nucleosides. These FONs are attractive because they are easy to prepare and are cheap (2). A genotyping method without amplification of genomic DNA has been reported (34). It is based on the different conformations of a cationic polythiophene F in the presence of single- and double-stranded DNA that results in a fluorescence signal increase in the presence of the latter. Fluorescence can be enhanced via FRET by convenient labeling of one strand of the duplex with Alexa Fluor 546. Using 20-mer probes, discrimination between the fully matched duplex and the mismatched one has been possible.
Detection of terminal mismatches on DNA duplexes by fluorescence sensing
FON probes labeled at their 5'-ends with thiazole orange can discern perfect duplexes from those that involve a terminal mismatch, as well as those that involve mismatches at the penultimate or last two positions under nonstringent conditions (31).
Increasing the specificity of detection with binary probes
Duplexes of 15-25 base pairs are often too stable to be sensitive to the presence of a mismatched base pair. The division of the ON probes into two parts that lead to good selectivity of the NA recognition has recently been reported. These examples are autoligating probes detectable by FRET (19) or aptamers (37) assembled in the presence of the NA target. The formation of the complex resulted in a great increase in the fluorescence emission of the reporter molecule. A malachite green aptamer can discriminate 41 out of 42 possible single-nucleotide substitutions in a 14-mer DNA target (37).
Detection of DNA abasic site with light-up probes
The detection of an abasic site located at the central position of a 13-base pair duplex by a modified nucleobase inserted at the opposite position was signaled by a 7-fold enhancement of the fluorescence emission when compared with that of the perfectly matched duplex (38).
Detection of mRNA splicing by FRET and single-molecule spectroscopy
Two probes fluorescently labeled to undergo FRET can hybridize to areas flanking the splice sites of an RNA molecule and demonstrate a difference in FRET efficiency between spliced and unspliced mRNA as shown by both bulk solution and single-molecule spectroscopy measurements (33).
Multiplex pathogen detection
The development of a highly accessible and easily adaptable multiplex system for the detection of pathogens remains the ultimate goal in molecular diagnostic laboratories (48). By attaching the MB to bar-coded nanowires, it is possible to detect multiple target sequences with only one F (26), whereas multiplex detection by MBs in a homogeneous solution requires different F/Q pairs (20). Recently, MBs labeled with 6-FAM/BHQ-1 or Cy5/BHQ-2 quencher pairs were linked to microspheres of different sizes by a biotin-streptavidine linkage and used for the multiplex detection by flow cytometry. The average limit of detection for these beads specific for SARS coronavirus, HCV, PIV-3, and RSV was found to be 37 fmol (21).
FISH to size the telomeric sequences
The length of the telomere repeats at individual chromosome ends influences biological functions that range from aging to carcinogenesis (9). Measurements of the telomere length can be achieved by fluorescence in situ hybridization (FISH). Recently, a flow FISH method has been developed (22).
Probing nucleic acid structures, dynamics, and interactions
Studies of the structural dynamics of NA in solution and their complexes are very important for the understanding of their functions in living organisms (4, 49). This research can be easily performed by using fluorescent base analogs that resemble natural ones with respect to their dimensions and hydrogen binding patterns (5). Among them, 2-aminopurine and pteridines have been used in numerous applications (5, 16). Alternatively, the covalent attachment of Fs at convenient positions of the NA to be analyzed can be performed to allow FRET to take place (3, 4). The latter strategy has been used to elucidate complex NA structures, which include the hammerhead ribozyme (23) and the human telometric G-quadruplex (27). Important biological processes such as the stepwise movement of an aminoacyl-tRNA into the ribosome (28) or the structural basis for Flap Endonuclease-1 substrate specificity (24) have also been studied by FRET.
Applications in cellular extracts and living cells
These applications require the use of FON probes resistant to nucleases. ONs with modified backbones such as oligo-2'-O-methyl-2'-deoxyribonucleotides (19), oligo-α-deoxy ribonucleotides (29), peptide nucleic acids (33) and locked nucleic acids (LNA) (50) (Fig. 4) are often used because of the commercially available building blocks. They form, with the RNA targets, hybrids that are not RNAse H substrates unless they can stimulate degradation of the targets rather than report on their presence.

Figure 4. Structures of selected ON backbone modifications resistant to nuclease degradation used in cellular experiments. (a) 2-O-methyl-2'-deoxyribonucleoside. (b) PNA. (c) Oligo-α-deoxyribonucleoside. (d) LNA.
The visualization of RNAs in cellular extracts using time-resolved spectroscopy
An important factor for in vivo studies is the auto fluorescent cellular background. To overcome this problem, pyrene-labeled binary probes have been used to take advantage of the long fluorescence lifetime (>40 ns) of the pyrene excimer, compared with that of the cellular extracts (7 ≈ ns), that allows selective detection of the excimer using time-resolved spectroscopy (35).
Profiling microRNA expression in biological samples
Because of the importance of micro-RNAs in gene regulation in both plants and animals, there is a need for profiling their expression in biological samples (51). Multiplexed detection methods in solution and in solid phase have been developed (52).
Detection of viral sequences by fluorescence sensing
The direct visualization of poliovirus NA in living host cells was achieved with MB, involving TAMRA as F and Dabcyl as Q, targeting the viral plus-strand RNA (25). The poliovirus plus strand was observed to display different distribution patterns at different post-infection time points in living vero cells.
The visualization of RNAs in living cells using FRET, excimer and light-up probes
Competitive hybridization probes (18), binary hybridization probes (35), MBs (40), light-up probes (29), and quenched auto-ligation probes (QUAL) (19) can be used. Among them, the quenched probe strategy offers the advantage of a decreased background signal in the absence of the RNA target. By using QUAL probes, mRNAs as well as 28S ribosomal RNA were detected in HL-60 cells by FC and were visible by confocal microscopy (19). A cell membrane-permeant couple of MBs designed to produce FRET in the presence of the target sequence was successfully used to detect human GAPDH and surviving mRNAs in living cells (41). Oligo-α-thymidylates that involve thiazole orange at the internucleotidic position were used to visualize the mRNAs intracellular distribution in HOS cells (29). The accurate detection of mRNA in living cells with F-ON probes requires homogenous distribution of the F-ONs throughout the cells. To make visible the cytoplasmic mRNAs in living cells, MBs were linked to tRNA (53).
Summary
In this article, we have attempted to show, through a few examples, the power of fluorescence strategies based on fluorescent oligonucleotides to study nucleic acids. This process has been made possible through chemistry that makes the synthesis of fluorescent oligonucleotidic probes possible with the required properties suitable for experiments in varied formats including living cell experiments. We apologize to authors whose work has inadvertently not been cited. It is our hope that this article will stimulate further reading and help the design of new strategies.
References
1. Beaucage SL, Caruthers MH. Deoxynucleoside phosphor amidites. A new class of key intermediates for deoxypolynucleotide synthesis. Tetrahed. Lett. 1981; 22:1859-1862.
2. Okamoto A, Saito Y, Saito I. Design of base-discriminating fluorescent nucleosides. J. Photochem. Photobiol. C: Photochem. Rev. 2005; 6:108-122.
3. Ranasinghe R, Brown T. Fluorescence based strategies for genetics analysis. Chem. Commun. 2005; 5487-5502.
4. Asseline U. Development and applications of fluorescent oligonucleotides. Curr. Org. Chem. 2006; 10:491-518.
5. Wilson JN, Kool ET. Fluorescent DNA base replacements: reporters and sensors for biological systems. Org. Biomol. Chem. 2006; 4:4265-4274.
6. Sapsford KE, Berti L, Medintz IL. Materials for fluorescence resonance energy transfer analysis: beyond traditional donor-acceptor combinations. Angew. Chem. Int. Ed. 2006; 45:4562- 4588.
7. Marti AA, Jockusch S, Stevens N, Ju J, Turro NJ. Fluorescent hybridization probes for sensitive and selective DNA and RNA detection. Acc. Chem. Res. 2007; 40:402-409.
8. Hirschhorn JN, Daly MJ. Genome-wide association studies for common diseases and complex traits. Nat. Rev. Genet. 2005; 6:95-108.
9. Steward SA, Weinberg RA. Telomeres: cancer to human aging. Annua. Rev. Cell. Dev. Biol. 2006; 22:531-557.
10. Lakowicz JR. Principles of Fluorescence Spectroscopy. 1999. Kluwer Academic/Plenum Publishers, New York.
11. Widengren J, Kudryavtsev V, Antonik M, Berger S, Gerken M, Seidel CAM. Single-molecule detection and identification of multiple species by multiparameter fluorescence detection. Anal. Chem. 2006; 78:2039-2050.
12. Haughland RP. Handbook of Fluorescent Probes and Research Products. 9th edition. Molecular Probes
13. Invitrogen. The handbook. A guide to fluorescent Probes and Labeling Technologies.
14. Molecular probes. GE Healthcare. AnaSpec. Pierce. Biosearch Technologies. AttoTec, Glen Research.
15. Marras SA, Kramer FR, Tyagi S. Efficiencies of fluorescence resonance energy transfer and contact-mediated quenching in oligonucleotide probes. Nucleic Acids Res. 2002; 30 e122.
16. Tetrahedron Symposium-in-Print Number 128. Fluorescent nucleoside analogs: synthesis, properties and applications. Tor Y, ed. Tetrahedron 2007; 63:(17).
17. Cardullo RA, Agrawal S, Flores C, Zamecnik PC, Wolf DE. Detection of nucleic acid hybridization by non-radiative fluorescence resonance energy transfer. Proc. Natl. Acad. Sci. U.S.A. 1988; 85:8790-8794.
18. Li Q, Luan G, Guo Q, Liang J. A new class of homogeneous nucleic acid probes based on specific displacement hybridization. Nucleic Acids Res. 2002; 30:e5.
19. Abe H, Kool ET. Flow cytometry detection of specific RNAs in native human cells with quenched autoligating FRET probes. Proc. Natl. Acad. Sci. U.S.A. 2006; 103:263-268.
20. Tyagi S, Bratu DP, Kramer FR. Multicolor molecular beacons for allele discrimination. Nat. Biotechnol. 1998; 16:49-53.
21. Horejsh D, Martini F, Poccia F, Ippolito G, Di Caro A, Capobianchi MR. A molecular beacon, bead-based assay for the detection of nucleic acids by flow cytometry. Nucleic Acids Res. 2005; 33:e13.
22. Baerlocher GM, Vulto I, de Jong G, Lansdorp PM. Flow cytometry and FISH to measure the average length of telomeres Nat. Protoc. 2006; 1:2365-2376.
23. Tuschl T, Gohlke C, Jovin TM, Westhof, E, Eckstein, F. A three-dimensional model for the hammerhead ribozyme based on fluorescence measurements. Science 1994; 266:785-789.
24. Chapados BR, Hosfield DJ, Han S, Qiu J, Yelent B, Shen B, Tainer JA. Structural basis for FEN-1 substrate specificity and PCNA-mediated activation in DNA replication and repair. Cell 2004; 116:39-50.
25. Cui ZQ, Zhang ZP, Zhang XE, Wen JK, Zhou YF, Xie WH. Visualizing the dynamic behaviour of poliovirus plus-strand RNA in living host cells. Nucleic Acids Res. 2005; 33:3245-3252.
26. Stoermer R, Cederquist KB, McFarland SK, Sha MY, Penn SG, Keating CD. Coupling molecular beacons to barcoded metal nanowires for multiplexed, sealed chamber DNA bioassays. J. Am. Chem. Soc. 2006; 128:16892-16903.
27. Ying L, Green JJ, Li H, Klenerman D, Balasubramanian S. Studies on the structure and dynamics of the human telomeric G quadruplex by single-molecule resonance energy transfer. Proc. Natl. Acad. Sci. U.S.A. 2003; 100:14629-14634.
28. Blanchard SC, Gonzalez RL, Kim HD, Chu S, Puglisi JD. tRNA selection and kinetic proofreading in translation. Nat. Struct. Mol. Biol. 2004; 11:1008-1014.
29. Privat E, Melvin T. Asseline U, Vigny P. Oligonucleotide conjugated thiazole orange probes as ‘light-up’ probes for messenge RNA molecules in living cells. Photochem. Photobiol. B. 2001; 74:532-541.
30. Privat E, Asseline U. Synthesis and properties of oligonucleotide-conjugated thiazole orange. Bioconjugate Chem. 2001; 12:757- 769.
31. Asseline U, Chassignol M, Aubert Y, Roig V. Detection of terminal mismatches on DNA duplexes with fluorescent oligonucleotides. Org. Biomol. Chem. 2006; 4:1949-1957.
32. Lartia R, Asseline U. New cyanine-oligonucleotide conjugates: relationships between chemical structures and properties. Chem. Eur. J. 2006; 12:2270-2281.
33. Robertson KL, Yu L, Armitage BA, Lopez AJ, Peteanu LA. Fluorescent PNA probes as hybridization labels for biological RNA. Biochemistry 2006; 45:6066-6074.
34. Ho HA, Dore K, Boissinot M, Bergeron MG, Tanguay RM, Boudreau D, Leclerc M. Direct Molecular detection of nucleic acids by fluorescence signal amplification. J. Am. Chem. Soc. 2005; 127:12673-12676.
35. Marti AA, Li X, Jockusch S, Li Z, Raveendra B, Kalachikov S, Russo JJ., Morozova I, Puthanveettil SV, Ju J, Turro NJ. Pyrene binary probes for unambiguous detection of mRNA using time-resolved fluorescence spectroscopy. Nucleic Acids Res. 2006; 34:3161-3168.
36. Conlon P, Yang CJ, Wu Y, Chen Y, Martinez K, Kim Y, Stevens N, Marti AA, Jockusch S, Turro NJ, Tan W. Pyrene excimer signaling molecular beacons for probing nucleic acids. J. Am. Chem. Soc. 2008; 130:336-342.
37. Kolpashchikov DM. Binary malachite green aptamer for fluorescent detection of nucleic acids. J. Am. Chem. Soc. 2005; 127:12442-12443.
38. Greco NJ, Tor Y. Simple fluorescent pyrimidine analogue detect the presence of DNA abasic sites. J. Am. Chem. Soc. 2005; 127:10784-10785.
39. Tyagi S, Kramer FR. Molecular beacons/Probes that fluoresce upon hybridization. Nat. Biotechnol. 1996; 14:303-308.
40. Bratu DP, Cha BJ, Mhlanga MM, Kramer FR, Tyagi S. Visualizing the distribution and transport of mRNAs in living cells. Proc. Natl. Acad. Sci. U.S.A. 2003; 100:13308-13313.
41. Nitin N, Santangelo PJ, Kim G, Nie S, Bao G. Peptide-linked molecular beacons for efficient delivery and rapid mRNA detection in living cells. Nucleic Acids Res. 2004; 32:e58.
42. Rosenblum BB, Lee LG, Spurgeon SL, Khan SH, Menchen SM, Heiner CR, Chen SM. New dye-labeled terminators for improved DNA sequencing patterns. Nucleic Acids Res. 1997; 25:4500- 4504.
43. Ju J, Kim, DH Bi, L, Meng Q, Bai X, Li Z, Li X, Marma MS, Shi S, Wu J, Edwards JR, Romu A, Turro NJ. Four-color DNA sequencing by synthesis using cleavable fluorescent nucleotide reversible terminators. Proc. Natl Acad. Sci. U.S.A. 2006; 103:19635-19640.
44. Barnes C, Balasubramanian S, Liu X, Swerdlow H, Milton J. Solexa sequencing technology. Illumina Inc. 2006. U.S. Patent 7,057,026.
45. Pljevaljcic G, Pignot M, Weinhold E. Design of a new fluorescent cofactor for DNA methyltransferase and sequence-specific labeling of DNA. J. Am. Chem. Soc. 2003; 125:3486-3492.
46. Kimoto M, Mitsui T, Harada Y, Sato A, Yokoyama S, Hirao I. Fluorescent probing for RNA molecules by an unnatural base-pair system. Nucleic Acids Res. 2007; 35:5360-5369.
47. Sobrino B, Brion M, Carracedo A. SNPs in forensic genetics: a review on SNP typing methodologies. Forensic Sci. Int. 2005; 154:181-194.
48. Sassolas A, Leca-Bouvier BD, Blum LJ. DNA biosensors and microarrays. Chem. Rev. 2008; 108:109-139.
49. Rist MJ, Marino JP. Fluorescent nucleotide base analogs as probes of nucleic acid structure, dynamics and interactions. Curr. Org. Chem. 2002; 6:775-793.
50. Kaur H, Babu BR, Maiti S. Perspectives on chemistry and therapeutic applications of Locked Nucleic Acids. Chem. Rev. 2007; 107:4672-4697.
51. He L, Hannon GJ. MicroRNAs: small RNAs with a big role in gene regulation. Nat. Rev. Gen. 2004; 5:522-531.
52. Wark AW, Lee HJ, Corn RM. Multiplexed detection methods for profiling microRNA expression in biological samples. Angew. Chem. Int. Ed. 2008; 47:644-652.
53. Mhlanga MM, Tyagi S. Using tRNA-linked molecular beacons to image cytoplasmic mRNAs in live cells. Nat. Protoc. 2006; 1:1392-1398.
Further Reading
Forster T. Analyt. Physic. 1948; 2:55.
Clegg RM. Methods Enzymol. 1992; 211:353
Selvin PR. Methods Enzymol. 1995; 246:300
Birks JB. Acta Phys. Polon. 1968; 34:603
Beaucage S, Bergstrom DE, Glick GD, Jones RA, eds. Current Protocols in Nucleic Acid Chemistry. 2000. John Wiley & Sons, New York.
Silverman AP, Kool ET. Oligonucleotide probes for RNA-targeted fluorescence in situ hybridization. Adv. Clin. Chem. 2007; 43:79-115.
See Also
Fluorescence in Living Systems: Overview of Applications in Chemical Biology
Fluorescent Spectroscopy: Overview of Applications in Chemical Biology
Tools to Detect DNA Polymorphisms (SNPs focus)
Peptide Nucleic Acids (PNAs)
Solid-Phase Synthesis of Biomolecules
Single Molecule Studies of Biomolecules