CHEMICAL BIOLOGY
Chemistry of Bacteriorhodopsin
Janos K. Lanyi, University of California, Los Angeles, California
doi: 10.1002/9780470048672.wecb035
Thirty years of research with bacteriorhodopsin has provided answers to many questions about how protons are transported by transmembrane pumps. In this small seven-transmembrane protein, absorption of light by the retinal chromophore initiates a reaction cycle in which the initial state recovers through multiple conformational changes of the retinal and the protein, and a proton is translocated stepwise from one side of the membrane to the other. Spectroscopy, extensive use of site-specific mutations, and crystallography have defined the photocycle reactions in atomic detail and provide a step-by-step description of the proton transfers, the transient local and global perturbations in the protein and how they arise, and the energy flow through the system, which add up to the mechanism of the pump.
Membrane proteins perform some of the most topologically complex and important functions in biology, such as translocating small and large molecules into and out of cells and organelles, coupling energetic functions through the generation and the discharge of transmembrane ion gradients, initiating signaling cascades in cells, and inserting and folding of polypeptides into the lipid bilayer. Bacteriorhodopsin belongs to the heptahelical membrane-protein family that also includes a wide variety of receptors for biological signals. As a light-driven pump for protons in halophilic archaea (1, 2), it generates the inwardly oriented transmembrane electrochemical proton gradient used for synthesis of adenosine triphosphate (ATP) and the uptake of nutrients by the cells. Illumination causes isomerization of the purple retinal chromophore from all-trans to 13-cis,15-anti, and the ensuing sequence of thermal reactions regenerates the initial state in ca. 10 ms, through protein conformational changes and internal proton transfers that translocate a proton across the membrane. Interest in bacteriorhodopsin has been renewed in the last few years because many bacteriorhodopsin-like photoreactive proteins are now found also in eubacteria and eukaryotes, and thus, it is becoming clear that the use of retinal for light-sensing and energetics is more widespread in biology than ever imagined. In addition to the already well-known chloride pump of the archaea, halorhodopsin, these newly discovered proteins include the proton pumps proteorhodopsin and xanthorhodopsin, and various sensory rhodopsins.
Background
As an active transporter, bacteriorhodopsin functions as a biological energy transducer. Proteins of this kind move ions against their electrochemical potential across the membrane barrier, using energy generated by a chemical reaction or from light. Such ion pumps are necessary for muscle contraction (Ca2+ transport), nerve conduction (Na+ and K+ transport), as well as ATP synthesis in mitochondria and photosynthetic systems (H+ transport). Because of their large size and in most cases complex multi-subunit structure, progress in understanding the mechanism of the transport and how it is coupled to the driving reaction in the pumps has been slow.
The conceptual and methodological advantages of bacteriorhodopsin have made it possible to gain insights into its structure, reaction cycle, and function as an ion pump in far more detail than for other membrane proteins. The outstanding questions concern the interhelical interactions that fold the protein and confer stability to the seven-helical bundle immersed in the lipid bilayer, the identity and location of functional groups in the protein interior, the nature of the perturbation of the retinal in its binding site when photoisomerized, the propagation of conformational changes from retinal to near and far regions of the protein, the kinetics and energetics of the reaction cycle in which a proton is transferred from group to group inside the protein, the proton release and uptake at the membrane surfaces, and the nature of the protonation switch that confers directionality on the pump. Answers to such questions are sought for many ion pumps, but in bacteriorhodopsin, the large amount of information now available gives a vivid picture of the molecular events that accomplish the transport.
Structure and Mechanism
The protein (26kDa) is a bundle of seven helices, A through G, with three helices normal to the membrane plane and the rest inclined at small angles to the normal. The retinal is covalently bound through a protonated Schiff base to Lys-216 at the middle of helix G, and it lies nearly parallel to the membrane in the space surrounded by the seven helices. The protein forms in vivo a homotrimer, and the trimers are assembled into an extended hexagonal lattice (P3 symmetry), the “purple membrane.” Protein-protein contact in these patches is through a continuous boundary layer of lipids no more than one lipid wide, at the monomer periphery. The interhelical loops as well as the N- and C-termini are short, with the exception of the connection of helices B and C through a structured P-turn on the extracellular side, and the E-F interhelical loop with a twist, on the cytoplasmic side.
At the active site, the positively charged protonated retinal Schiff base donates a hydrogen bond to a water molecule (wat402) that donates in turn hydrogen bonds to two anionic aspartate residues, Asp-85 and Asp-212 (3). This stable arrangement of a potential proton donor (Schiff base) and a potential acceptor (Asp85) is the active center, which divides the protein into extracellular and cytoplasmic regions. The extracellular region contains a pathway to the surface through charged and polar side chains and seven bound water molecules (Fig. 1). The most important parts of this network are the positively charged Arg-82 and the hydrogen-bonded aqueous network from which the transported proton is released to the surface, coordinated by Glu-194 and Glu-204 (3-5). In contrast, the cytoplasmic region mostly lacks polar groups and water and represents the hydrophobic barrier to prevent leakage of protons and other ions across the protein and thereby across the membrane (3, 6). This region contains the proton donor to the Schiff base after it is deprotonated in the cycle, Asp-96, which is hydrogen-bonded to Thr-46 (Fig. 2).
Absorption of a photon by the purple all-trans retinal chromophore (with a single broad absorption band with a maximum at 568 nm) initiates the reaction sequence BR-hv ⇒ K ⇔ L ⇔ M1 ⇔ M2 ⇔ N⇔ N’ ⇒ O ⇒ BR (7, 8), where each state and substate is well defined by spectroscopic and crystallographic means. Although a kinetic scheme that rigorously fits all data into a linear sequence has not yet been produced, the proton transport mechanism can be understood by the molecular properties of the intermediate states and by their interconversions.
The first stable state, the K-intermediate, contains a highly twisted 13-cis,15-anti retinal, with prominent hydrogen-out-ofplane (HOOP) bands, particularly at C15. Anomalous Schiff base C=N stretch frequency, lesser 15N chemical shift, and the X-ray diffraction structures of K indicate that rotation of the C13=C14 bond disconnects the Schiff base N-H bond from wat402. At this stage, the overall contour of the all-trans retinal is maintained despite the isomerization. In the states that follow K, the retinal gradually relaxes, and deprotonation of the Schiff base as well as an increasingly accommodating binding site allow it to assume the bent configuration expected for a 13-cis ,15-anti retinal.
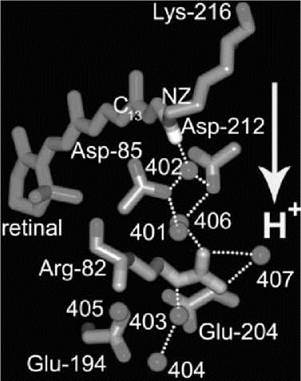
Figure 1. Functional residues and bound water in the extracellular region of bacteriorhodopsin. The all-trans retinal is shown in purple and the hydrogen bonds in gold. The direction of overall proton translocation is indicated with an arrow. The Schiff base (NZ) and the C13 atom of the retinal are labeled. Coordinates from Reference 3.
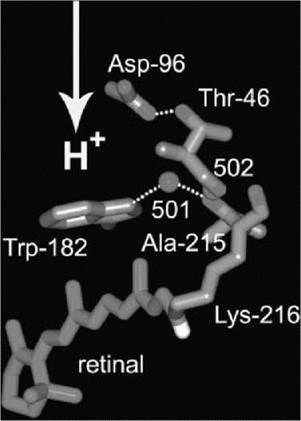
Figure 2. Functional residues and bound water in the cytoplasmic region of bacteriorhodopsin. The all-trans retinal is shown in purple and the hydrogen bonds in gold. The direction of overall proton translocation is indicated with an arrow. Coordinates from Reference 3.
In the L-state, the retinal is still somewhat twisted but the Schiff base regains a hydrogen bond. There is disagreement over the partner in this hydrogen bond. In the highest resolution X-ray structure (9), it is wat402 (still to the extracellular direction), but in others, it is wat402 moved to the cytoplasmic side (10). In the latter case, rotation of the C15=NZ-CE segment will have reoriented the Schiff base in the L-state already. Other evidence suggests that this occurs only in the M-state(s), permitted by loss of the Schiff base—counterion interaction upon transfer of the Schiff base proton to Asp-85. The two alternatives conduct the Schiff base proton to Asp-85 by different pathways. In the first case, the protonation switch in the pump is primarily in the changing proton affinities of donor and acceptor because the critical proton transfer is directly over a water molecule. In the second, the donor and acceptor are separated from one another, and their geometry also plays an important role.
The Arg-82 side chain moves away from the now protonated Asp-85 (11), toward the Glu-194/Glu-204 pair, and a proton delocalized in the aqueous network is released to the surface. The rise of the M-state occurs with multi-exponential kinetics that originates partly from the additional step of proton re-equilibration within the Schiff base-Asp-85 pair as the proton is released to the bulk. At this stage, the pKa of Asp-85 will have risen considerably as the result of its long-range coupling (5) to the proton release site through Arg-82, by about five units, and if the Schiff base is to be reprotonated, it must be by a proton donor other than Asp-85. In the next steps, a suitable proton donor will be generated on the cytoplasmic side.
As the retinal relaxes, C13 and the 13-methyl group moves in the cytoplasmic direction toward Trp-182. The resulting displacements of side chains between helices F and G, and the breaking of the connection between helices F and G through the hydrogen bonds of wat501 to Trp-182 and Ala-215 (Fig. 2), separate helices F and G. The link of Lys216 to wat502 and thereby to the peptide segment of Thr-46 propagates perturbation from the retinal to the Asp-96/Thr-46 pair. Entry of water and growth of water clusters creates a hydrogen-bonded chain of four water molecules between Asp-96 and deprotonated Schiff base (6). The increase of hydration at Asp-96 lowers its initially high pKa, and the aqueous chain conducts the proton to the Schiff base. The M-to-N reaction corresponds to this proton transfer, followed by reprotonation of Asp-96 from the cytoplasmic surface that produces N’ from N. Crystallographic evidence and much non-crystallographic data indicate that these photocycle steps are accompanied by the outward and inward tilts of the cytoplasmic ends of helices F and G, respectively. The tilt of helix G arises from rearrangement of hydrogen bonds that eliminate, partly, the n-bulge in this helix at Lys-216. The tilt of helix F is a consequence of the separation of helices F and G and the repacking of side chains between them. There is reason to believe that it is the tilts that loosen the structure and allow the entry of water.
Reprotonation of Asp-96 allows the thermal re-isomerization of the retinal to all-trans, through another long-range coupling mechanism (12), which probably involves the hydrogen-bonded chain of water that bridges them. This produces the red-shifted O-state, where the retinal is, once again, twisted. Transition to the initial relaxed all-trans isomeric state occurs as the protonated Asp-85 loses its proton to the vacant proton release site at Glu-194/Glu-204, thereby recovering the initial electrostatic environment of the extracellular region and completing the cycle.
Key Experimental Methodology
The current understanding of how bacteriorhodopsin transports protons is the result of the use of an unprecedented variety of biophysical methods in many laboratories. Indeed, in many cases, the methods were developed with bacteriorhodopsin as the test subject. There is room to discuss only the most important of these methods.
Transient and low temperature spectroscopy in the ultra- violet/visible and infrared [Fourier transform infrared (FTIR) and resonance Raman] have yielded spectra of the intermediate states and provided many insights, even if not a fully satisfactory solution, to the kinetics of their interconversions. Transient spectroscopy in the visible (7) makes use of following absorption changes at selected single wavelengths, over as many as six orders of magnitude of time after flash photoexcitation, as well as an optical multichannel analyzer to obtain complete spectra at selected times after the flash. Alternatively, the intermediates can be trapped at cryogenic temperatures, after producing photostationary states (13). Both methods produce characteristic difference spectra and are well suited for identifying which of the various states is produced, as their single absorption maxima for the most part are well defined. Quantitation of these states and calculation of a kinetic model from the observed succession of time-resolved spectra is more difficult, because the spectra are broad and several overlap one another. Furthermore, the reaction scheme is more complex than what is expected from a linear sequence in which each state arises by unidirectional transition from the preceding one. Multiple rise and decay components have been interpreted as evidence for back-reactions that lead to transient equilibration of states, and as multiple parallel photocycles. If present, the latter could arise from conformational heterogeneity, interactions within the bacteriorhodopsin trimer, and heterogenous protonation states near the pKa for the proton release group. Infrared and Raman spectroscopy is performed along the same lines, but the vibrational spectra (14-16) provide essential molecular level information about changes in the isomeric state of the retinal, the protonation states of dissociable residues, the strength of hydrogen bonds, and backbone conformation. Additional information on these spectra comes from the use of D2O, H218O, as well as 13C-, 18O-, and 15N-labeled amino acids, and polarized infrared beam for measuring linear dichroism, as well as single-site mutations to change the photocycle and to assign the positive and negative difference bands to specific amino acid residues.
Solid-state nuclear magnetic resonance (NMR) spectroscopy of bacteriorhodopsin is with the membranes either uniformly oriented relative to the magnetic field (17) or nonoriented but with “magic angle” spinning (18). In either case, the samples are enriched with 15N or 13C isotopes in the examined parts of the molecule. The chemical shifts identify the protonation states of acidic groups, their hydrogen-bonding status, and the electrostatic environment. Dipolar interactions provide accurate interatomic distance and dihedral angle. Importantly, like FTIR, this method does not average structural information in the case of conformational heterogeneity. Difficulties include a requirement for large amounts of specifically labeled bacteriorhodopsin and problems of thoroughly illuminating a nearly opaque sample to trap the photocycle intermediates.
The purple membrane is isolated as an extended two-dimensional array of bacteriorhodopsin and lipids and, after some manipulation to increase the size of the patches, is an ideal material for cryo-electron microscopy. The maps produced to 3.5-7-A resolution (19) identified the positions of the seven-transmembrane helices, the retinal, and some bulky side chains. The latter helped to establish the rotational position of most helices in the seven-helical bundle, and thus, it yielded the first useful structural model. In the last decade, it has been possible to grow well-diffracting three-dimensional crystals. Bacteriorhodopsin crystals are usually thin (10-15 μm) hexagonal plates, 100-150 μm in diameter. X-ray diffraction of crystals produced reflections complete to 1.4-1.5-A resolution, and the resulting model (3) defines the locations of water molecules bound in the protein interior and the hydrogen bonds relevant to the function and the stability of the structure. The highest crystallographic order is in crystals grown with the cubic phase method (20). The protein forms trimers in the same extended P3 sheet as in the native purple membrane, but the stacking of these sheets to produce the P63 symmetry is not with uniform orientation and the crystals usually show merohedral twinning. This problem is absent in crystals grown by other methods (10), but they are less ordered and the resolution is poorer. Many mutations of functional interest apparently disrupt the structure in subtle ways and make the protein unstable in detergent or do not allow the growth of crystals.
The cubic phase for the crystallization (20) is formed by a roughly 1:1 mixture of mono-olein and bacteriorhodopsin solubilized in octylglucoside. In a process not well understood, salts such as KCl or Na phosphate cause the conversion of the cubic phase with the incorporated bacteriorhodopsin into a lamellar phase and condensation of the protein into crystals. Another method, based on bicelles, adds phospholipids to the mixture. Conventional vapor phase crystallization in the presence of detergent yields bacteriorhodopsin crystals with a somewhat different packing arrangement. The crystals in general are suitable for the trapping of most (K-, L-, various M-states, and N’), but not all (N and O), intermediate states (9). The M-state(s) can be accumulated with greatest ease because its absorption band is far removed from the initial state and its decay can be slowed by suitable mutations. Depending on their spectral overlap with the non-illuminated state, the quantum yields for the forward and back photoreactions, the wavelength of illumination, temperature, and mutations (if any), the other intermediates accumulate up to 60% in the photostationary states. Diffraction is measured at 100 K, where the intermediates are thermally stable and damage from the X rays is minimized.
Some movements at specific locations in the protein, for example, at the cytoplasmic surface in the M- and N-states, can be determined with site-specific spin-labels. This method is of particular value for intermediate states that do not accumulate in illuminated crystals. The paramagnetic probes are covalently linked to engineered cysteines, and the erythrocyte sedimentation rate (ESR) spectra of single or pairs of labels, determined statically in photostationary states or dynamically during the photocycle initiated by flash photoexcitation, reveal conformational shifts as changes in environmental restrictions on motion or changes in inter-spin distance with two labeled sites (21, 22). As in other proteins, a complementary method is to follow the reactivity of cysteine residues, placed one by one, along appropriate segments of the protein, such as interhelical loops.
Energetics of the Pump
The quantum yield of the photoisomerization from all-trans to 13-cis,15-anti is 0.60. From calorimetric measurements, the enthalpy gain in the first ground-state intermediate, K, is ca. 45 kJ/mol (23). Part of this will be used for proton transport, the rest is dissipated in the photocycle. One proton is translocated in each cycle across the membrane dielectric, against a proton gradient as high as 200 mV. The pump efficiency (the free energy of the transported proton vs. the energy gain in the K-state) is ca. 50%, and the overall efficiency of use of the energy input (the free energy of the transported proton vs. the energy of the absorbed photon) is ca. 10%.
In the K-state, the energy gain is conserved in bond torsions of the retinal from steric constraints, and electrostatic potential from loss of the Schiff base—counterion interaction and the resulting rearrangement of charges in the extracellular network. As the retinal Schiff base protonates Asp-85 in the L-to-M reaction so as to lose their electrostatic interaction, the binding site relaxes, stepwise, to accommodate the changed shape of the retinal, and free energy is passed from the chromophore to the protein. Most photocycle steps occur without a net free energy change, i.e., near equilibrium (8). The physiological pH (7.5) is well above the pKa of the extracellular proton release complex (ca. 5.0), and the proton release constitutes one of the two steps where energy is dissipated, here as dilution entropy. In contrast, the pH where proton is taken up at the cytoplasmic surface is about the same as the pKa of Asp-96 (ca. 7). The last step in the cycle is where the greatest part of the free energy gain is lost, resulting in a strongly unidirectional reaction that fully recovers the initial state from O. Loss of free energy in this step occurs most likely from proton transfer from Asp-85 to the proton release complex after these regain their low and high proton affinities, respectively (where the pKas are 2.5 and ca. 9.5).
References
1. Oesterhelt D, Stoeckenius W. Functions of a new photoreceptor membrane. Proc. Natl. Acad. Sci. U.S.A. 1973; 70:2853-2857.
2. Lanyi JK. Bacteriorhodopsin. Annu. Rev. Physiol. 2004; 66:665- 688.
3. Luecke H, Schobert B, Richter HT, Cartailler JP, Lanyi JK. Structure of bacteriorhodopsin at 1.55 A resolution. J. Mol. Biol. 1999; 291:899-911.
4. Garczarek F, Brown LS, Lanyi JK, Gerwert K. Proton binding within a membrane protein by a protonated water cluster. Proc. Natl. Acad. Sci. U.S.A. 2005; 102:3633-3638.
5. Balashov SP, Imasheva ES, Govindjee R, Ebrey TG. Titration of 19. aspartate-85 in bacteriorhodopsin: what it says about chromophore isomerization and proton release. Biophys. J. 1996; 70:473-481.
6. Schobert B, Brown LS, Lanyi JK. Crystallographic structures of 20. the M and N intermediates of bacteriorhodopsin: assembly of a hydrogen-bonded chain of water molecules between Asp96 and the retinal Schiff base. J. Mol. Biol. 2003; 330:553-570.
7. Ludmann K, Gergely C, Varo G. Kinetic and thermodynamic 21. study of the bacteriorhodopsin photocycle over a wide pH range. Biophys. J. 1998; 75:3110-3119.
8. Varo G, Lanyi JK. Thermodynamics and energy coupling in the bacteriorhodopsin photocycle. Biochemistry 1991; 30:5016-5022.22.
9. Lanyi JK, Schobert B. Local-global conformational coupling in a heptahelical membrane protein: transport mechanism from crystal structures of the nine states in the bacteriorhodopsin photocycle. 23. Biochemistry 2004; 43:3-8.
10. Kouyama T, Nishikawa T, Tokuhisa T, Okumura H. Crystal structure of the L intermediate of bacteriorhodopsin: evidence for vertical translocation of a water molecule during the proton pumping cycle. J. Mol. Biol. 2004; 335:531-546.
11. Luecke H, Schobert B, Richter HT, Cartailler JP, Lanyi JK. Structural changes in bacteriorhodopsin during ion transport at 2 Angstrom resolution. Science 1999; 286:255-261.
12. Dioumaev AK, Brown LS, Needleman R, Lanyi JK. Partitioning of free energy gain between the photoisomerized retinal and the protein in bacteriorhodopsin. Biochemistry 1998; 37:9889-9893.
13. Balashov SP, Ebrey TG. Trapping and spectroscopic identification of the photointermediates of bacteriorhodopsin at low temperatures. Photochem. Photobiol. 2001; 73:453-462.
14. Maeda A, Kandori H, Yamazaki Y, Nishimura S, Hatanaka M, Chon YS, Sasaki J, Needleman R, Lanyi JK. Intramembrane signaling mediated by hydrogen-bonding of water and carboxyl groups in bacteriorhodopsin and rhodopsin. J. Biochem. (Tokyo) 1997; 121:399-406.
15. Gerwert K. Molecular reaction mechanisms of proteins monitored by time-resolved FTIR-spectroscopy. Biol. Chem. 1999; 380:931-935.
16. Mathies RA, Lin SW, Ames JB, Pollard WT. From femtoseconds to biology: mechanism of bacteriorhodopsin’s light-driven proton pump. Annu. Rev. Biophys. Biophys. Chem. 1991; 20:491-518.
17. Moltke S, Wallat I, Sakai N, Nakanishi K, Brown MF, Heyn MP. The angles between the Q, C5, and C9 methyl bonds of the retinylidene chromophore and the membrane normal increase in the M intermediate of bacteriorhodopsin: direct determination with solid-state 2H NMR. Biochemistry 1999; 38:11762-11772.
18. Herzfeld J, Lansing JC. Magnetic resonance studies of the bacteriorhodopsin pump cycle. Annu. Rev. Biophys. Biomol. Struct. 2002; 31:73-95.
19. Grigorieff N, Ceska TA, Downing KH, Baldwin JM, Henderson R. Electron-crystallographic refinement of the structure of bacteriorhodopsin. J. Mol. Biol. 1996; 259:393-421.
20. Rummel G, Hardmeyer A, Widmer C, Chiu ML, Nollert P, Locher KP, Pedruzzi I, Landau EM, Rosenbusch JP. Lipidic cubic phases: new matrices for the three-dimensional crystallization of membrane proteins. J. Struct. Biol. 1998; 121:82-91.
21. Rink T, Pfeiffer M, Oesterhelt D, Gerwert K, Steinhoff HJ. Unraveling photoexcited conformational changes of bacteriorhodopsin by time resolved electron paramagnetic resonance spectroscopy. Biophys. J. 2000; 78:1519-1530.
22. Thorgeirsson TE, Xiao W, Brown LS, Needleman R, Lanyi JK, Shin YK. Transient channel-opening in bacteriorhodopsin: an EPR study. J. Mol. Biol. 1997; 273:951-957.
23. Birge RR, Cooper TM, Lawrence AF, Masthay MB, Zhang CF, Zidovetzki R. Revised assignment of energy storage in the primary photochemical event in bacteriorhodopsin. J. Am. Chem. Soc. 1991; 113:4327-4328.
Further Reading
Bondar AN, Fischer S, Smith JC, Elstner M, Suhai S. Key role of electrostatic interactions in bacteriorhodopsin proton transfer. J. Am. Chem. Soc. 2004; 126:14668-14677.
Haupts U, Tittor J, Oesterhelt D. Closing in on bacteriorhodopsin: progress in understanding the molecule. Annu. Rev. Biophys. Biomol. Struct. 1999; 28:367-399.
Lanyi JK. A structural view of proton transport in bacteriorhodopsin. In: Biophysical and Structural Aspects of Bioenergetics. Wikstrom M, ed. 2005. Royal Society of Chemistry, Cambridge, UK.
Spudich JL, Yang CS, Jung KH, Spudich EN. Retinylidene proteins: structures and functions from archaea to humans. Annu. Rev. Cell Dev. Biol. 2000; 16:365-392.
See Also
Membrane Proteins, Chemistry of
Photoreceptors, Chemistry of
Proton Translocation, Bioenergetics of
Crystallization of Proteins: Overview of
Spectroscopic Techniques: Overview of