CONCEPTS IN BIOLOGY
PART II. CORNERSTONES: CHEMISTRY, CELLS, AND METABOLISM
4. Cell Structure and Function
4.4. Organelles Composed of Membranes
Although all cells have membranes, eukaryotic cells have many more organelles composed of membranes than do Bacteria and Archaea. Organelles are involved in specialized metabolic activities, the movement of molecules from one side of the membrane to the other, the identification of molecules, and many other activities. In the following section about the plasma membrane, many of these special properties will be discussed in detail.
Plasma Membrane
The outer limiting boundary of all cells is known as the plasma membrane, or cell membrane. It is composed of a phospholipid bilayer and serves as a barrier between the cell contents and the external environment. However, it is not just a physical barrier. The plasma membrane has many different functions. In many ways, it acts in a manner analogous to a border between countries, separating but also allowing controlled movement from one side to the other. The plasma membrane performs several important activities.
Metabolic Activities
Because the plasma membrane is part of a living unit, it is metabolically active. Many important chemical reactions take place within the membrane or on its inside or outside surface. Many of these chemical reactions involve transport of molecules.
Movement of Molecules Across the Membrane
Cells must continuously receive nutrients and rid themselves of waste products—one of the characteristics of life. There is constant traffic of molecules from the external matrix, or environment, across the plasma membrane. The surrounding matrix is rich in many kinds of important compounds including nutrients, growth factors, and hormones. See section 4.7 for a detailed discussion of the many ways by which molecules enter and leave cells. Many of the proteins that are associated with the plasma membrane are involved in moving molecules across the membrane. Some proteins are capable of moving from one side of the plasma membrane to the other and shuttle certain molecules across the membrane. Others extend from one side of the membrane to the other and form channels through which substances can travel. Some of these channels operate like border checkpoints, which open and close when circumstances dictate. Some molecules pass through the membrane passively, whereas others are assisted by metabolic activities within the membrane.
Inside and Outside
The inside of the plasma membrane is different from its outside. The carbohydrates that are associated with the plasma membrane are usually found on the outside of the membrane, where they are bound to proteins or lipids. Many important activities take place on only one of the surfaces of the plasma membrane because of the way the two sides differ.
Identification
The outside surface of the plasma membrane has many proteins, which act as recognition molecules. Each organism has a unique combination of these molecules. Thus, the presence of these molecules enables one cell or one organism to recognize cells that are like it and those that are different. For example, if a disease organism enters your body, the cells of your immune system use the proteins on the invader’s surface to identify it as being foreign. Immune system cells can then destroy the invader (How Science Works 4.2).
Attachment Sites
Some molecules on the outside surface of the plasma membrane serve as attachment sites for specific chemicals, bacteria, protozoa, white blood cells, and viruses. Many dangerous agents cannot stick to the surface of cells and therefore do not cause harm. For this reason, cell biologists explore the exact structure and function of these cell surface molecules. They are also attempting to identify molecules that can interfere with the binding of viruses and bacteria to cells in the hope of controlling infections. For example, human immunodeficiency virus (HIV) attaches to specific molecules on the surface of certain immune system cells and nerve cells. If these attachment sites could be masked, the virus would not be able to attach to the cells and cause disease. Drugs that function this way are called “blockers.”
Signal Transduction
Another way in which attachment sites are important is in signal transduction. Signal transduction is the process by which cells detect specific signals from the surrounding intercellular matrix and transmit these signals to the cell’s interior. These signals can be physical (electrical or heat) or chemical. Some chemicals are capable of passing directly through the membrane of specific target cells. Once inside, they can pass on their message to regulator proteins. These proteins then enter into chemical reactions, which result in a change in the cell’s behavior. For example, estrogen produced in one part of the body travels through the bloodstream and passes through the tissue to make direct contact with specific target cells. Once the hormone passes through the plasma membrane of the target cells, the message is communicated to begin the process of female sex organ development. This is like a person smelling the cologne of his or her date through a curtain. The aroma molecules pass through the curtain to the person’s nose and stimulate a response.
However, most signal molecules are not capable of entering cells in such a direct manner but remain in the external environment (i.e., outside their target cells). When they arrive at the cell, they attach to a receptor site molecule embedded in the membrane. The signal molecule is often called the primary messenger. The receptor-signal molecule combination initiates a sequence of events within the membrane that transmits information through the membrane to the interior, generating internal signal molecules, called secondary messengers.
The secondary messengers are molecules or ions that begin a cascade of chemical reactions causing the target cell to change how it functions (figure 4.8). This is like your mother sending your little brother to tell you it is time for dinner. Your mother provides the primary message, your little brother provides the secondary message, and you respond by going home. In a cell, such signal transduction results in a change in the cell’s chemical activity. Often, this is accomplished by turning genes on or off. For example, when a signal molecule called epidermal growth factor (EGF) attaches to the receptor protein of skin cells, it triggers a chain of events inside the plasma membrane of the cells. These changes within the plasma membrane produce secondary messengers, ultimately leading to gene action, which in turn causes cell growth and division.
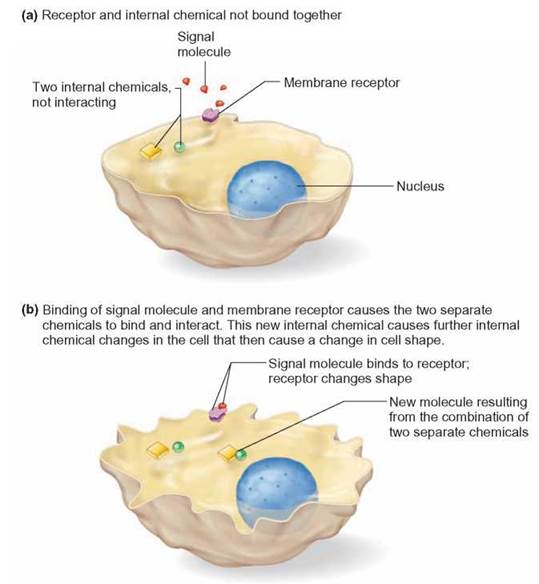
FIGURE 4.8. Signal Transduction and Secondary Messengers
Signal transduction results in chemical changes within the cell and is the result of cell membrane receptors binding with signal molecules from outside the cell. Secondary messengers inside the cell then communicate this information to appropriate molecules, sometimes to DNA.
HOW SCIENCE WORKS 4.2
Cell Membrane Structure and Tissue Transplants
In humans, there is a group of protein molecules, collectively known as histocompatibility antigens (histo = tissue), that are located on the cell surface. Each person has a specific combination of these proteins. It is the presence of these antigens that is responsible for the rejection of transplanted tissues or organs from donors that are "incompatible." In large part, a person's pattern of histocompatibility antigens is hereditary; for instance, in identical twins, the cells of both individuals have a very high percentage of similar proteins. Therefore, in transplant situations, the cells of the immune system would see the cells of the donor twin to be the same as those on the cell surfaces of the recipient twin. When closely related donors are not available, physicians try to find donors whose histocompat-ability antigens are as similar as possible to those of recipients.
Endoplasmic Reticulum
There are many other organelles in addition to the plasma membrane, that are composed of membranes. Each of these membranous organelles has a unique shape or structure associated with its particular functions. One of the most common organelles found in cells, the endoplasmic reticulum (ER), consists of folded membranes and tubes throughout the cell (figure 4.9). This system of membranes provides a large surface on which chemical activities take place. Because the ER has an enormous surface area, many chemical reactions can be carried out in an extremely small space. Picture the vast surface area of a piece of newspaper crumpled into a tight little ball. The surface contains hundreds of thousands of tidbits of information in an orderly arrangement, yet it is packed into a very small volume.
Proteins on the surface of the ER are actively involved in controlling and encouraging chemical activities—whether they are reactions involving cell growth and development or reactions resulting in the accumulation of molecules from the environment. The arrangement of the proteins allows them to control the sequences of metabolic activities, so that chemical reactions can be carried out very rapidly and accurately.
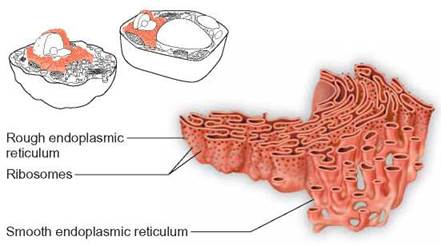
FIGURE 4.9. Endoplasmic Reticulum
The endoplasmic reticulum consists of folded membranes located throughout the cytoplasm of the cell. Some endoplasmic reticulum has ribosomes attached and appears rough. Many kinds of molecules are manufactured on the surfaces of endoplasmic reticulum.
On close examination with an electron microscope, it is apparent that there are two types of ER—rough and smooth. The rough ER appears rough because it has ribosomes attached to its surface. Ribosomes are nonmembranous organelles that are associated with the synthesis of proteins from amino acids. They are “protein-manufacturing machines.” Therefore, cells with an extensive amount of rough ER—for example, human pancreas cells—are capable of synthesizing large quantities of proteins. Smooth ER lacks attached ribosomes but is the site of many other important cellular chemical activities. Fat metabolism and detoxification reactions involved in the destruction of toxic substances, such as alcohol and drugs occur on this surface. Human liver cells are responsible for detoxification reactions and contain extensive smooth ER.
In addition, the spaces between the folded membranes serve as canals for the movement of molecules within the cell. This system of membranes allows for the rapid distribution of molecules within a cell.
Golgi Apparatus
Another organelle composed of membrane is the Golgi apparatus. Animal cells contain several such structures and plant cells contain hundreds. The typical Golgi apparatus consists of 5 to 20 flattened, smooth membranous sacs, which resemble a stack of flattened balloons (figure 4.10). The Golgi apparatus has several functions:
1. it modifies molecules shipped to it from elsewhere in the cell,
2. it manufactures some polysaccharides and lipids, and
3. it packages molecules within sacs.
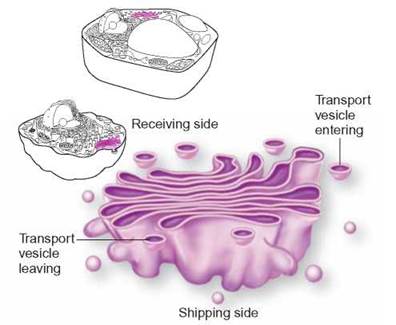
FIGURE 4.10. Golgi Apparatus
The Golgi apparatus is a series of membranous sacs that accept packages of materials and produce vesicles containing specific molecules. Some packages of materials are transported to other parts of the cell. Others are transported to the plasma membrane and release their contents to the exterior of the cell.
There is a constant traffic of molecules through the Golgi apparatus. Tiny, membranous sacs called vesicles deliver molecules to one surface of the Golgi apparatus. Many of these vesicles are formed by the endoplasmic reticulum and contain proteins. These vesicles combine with the sacs of the Golgi apparatus and release their contents into it. Many kinds of chemical reactions take place within the Golgi apparatus. Ultimately, new sacs, containing “finished products,” are produced from the surface of the Golgi apparatus.
The Golgi apparatus produces many kinds of vesicles. Each has a different function. Some are transported within the cell and combine with other membrane structures, such as the endoplasmic reticulum. Some migrate to the plasma membrane and combine with it. These vesicles release molecules such as mucus, cellulose, glycoproteins, insulin, and enzymes to the outside of the cell. In plant cells, cellulose-containing vesicles are involved in producing new cell wall material. Finally, some of the vesicles produced by the Golgi apparatus contain enzymes that can break down the various molecules of the cell, causing its destruction. These vesicles are known as lysosomes.
Lysosomes
Lysosomes are tiny vesicles that contain enzymes capable of digesting carbohydrates, nucleic acids, proteins, and lipids. Because cells are composed of these molecules, these enzymes must be controlled in order to prevent the destruction of the cell. This control is accomplished very simply. The enzymes of lysosomes function best at a pH of about 5. The membrane, which is the outer covering of the lysosome, transports hydrogen ions into the lysosome and creates the acidic conditions these enzymes need. Since the pH of a cell is generally about 7, these enzymes will not function if released into the cell cytoplasm.
The functions of lysosomes are basically digestion and destruction. For example, in many kinds of protozoa, such as Paramecium and Amoeba, food is taken into the cell in the form of a membrane-enclosed food vacuole. Lysosomes combine with food vacuoles and break down the food particles into smaller molecules, which the cell can use.
In a similar fashion, lysosomes destroy disease-causing microorganisms, such as bacteria, viruses, and fungi. The microorganisms become surrounded by membranes from the endoplasmic reticulum. Lysosomes combine with the membranes surrounding these invaders and destroy them. This kind of activity is common in white blood cells that engulf and destroy disease-causing organisms.
Lysosomes are also involved in the breakdown of worn-out cell organelles by fusing with them and destroying them (figure 4.11).
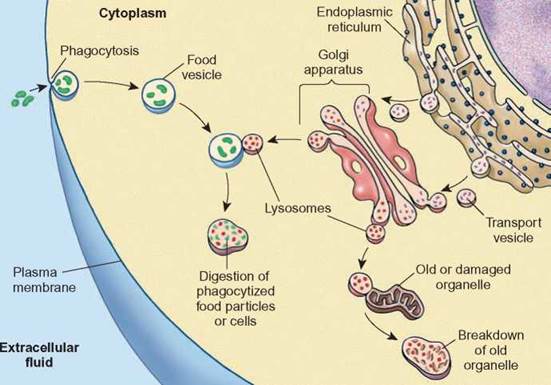
FIGURE 4.11. Lysosome Function
Lysosomes contain enzymes that are capable of digesting many kinds of materials. They are involved in the digestion of food vacuoles, harmful organisms, and damaged organelles.
Peroxisomes
Another organelle that consists of many kinds of enzymes surrounded by a membrane is the peroxisome. Peroxisomes were first identified by the presence of an enzyme, catalase, that breaks down hydrogen peroxide (H2O2). Peroxisomes differ from lysosomes in that peroxisomes are not formed by the Golgi apparatus and they contain different enzymes. It appears that the membrane surrounding peroxisomes is formed from the endoplasmic reticulum and the enzymes are imported into this saclike container. The enzymes of peroxisomes have been shown to be important in many kinds of chemical reactions. These include the breakdown of long-chain fatty acids, the synthesis of cholesterol, and the synthesis of plasma membrane lipids used in nerve cells.
Vacuoles and Vesicles
There are many kinds of membrane-enclosed containers in cells known as vacuoles and vesicles. Vacuoles are the larger structures and vesicles are the smaller ones. They are frequently described by their function. In most plants, there is one huge, centrally located, water-filled vacuole. Many kinds of protozoa have specialized water vacuoles called contractile vacuoles which are able to forcefully expel excess water that has accumulated in the cytoplasm. The contractile vacuole is a necessary organelle in cells that live (figure 4.12) in freshwater because water constantly diffuses into the cell. Animal cells typically have many small vacuoles and vesicles throughout the cytoplasm.
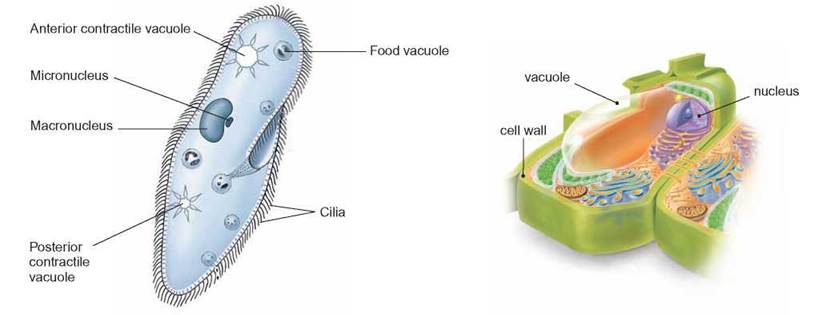
FIGURE 4.12. Vacuoles
Vacuoles are membrane-enclosed sacs that contain a variety of materials. Often, in many kinds of protozoa, food is found inside vacuoles. Plant cells have a large central vacuole filled with water. Some freshwater organisms have contractile vacuoles that expel water from the cell.
Nuclear Membrane
Just as a room is a place created by walls, a floor, and a ceiling, a cell’s nucleus is a place created by the nuclear membrane. If the nuclear membrane were not formed around the cell’s genetic material, the organelle called the cellular nucleus would not exist. This membrane separates the genetic material (DNA) from the cytoplasm. Because they are separated, the cytoplasm and the nuclear contents can maintain different chemical compositions. The nuclear membrane is composed of two layers and has openings called nuclear pore complexes (figure 4.13). The nuclear pore complexes consist of proteins, which collectively form barrel-shaped pores. These pores allow relatively large molecules, such as RNA, to pass through the nuclear membrane. Thousands of molecules move in and out through these pores each second.
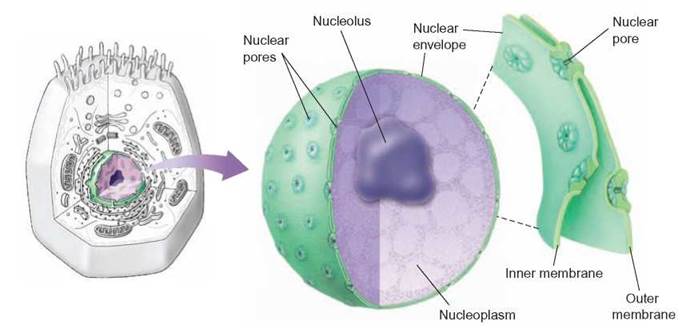
FIGURE 4.13. Nuclear Membrane
The nuclear membrane is a double membrane separating the nuclear contents from the cytoplasm. Pores in the nuclear membrane allow molecules as large as proteins to pass through.
The Endomembrane System—Interconversion of Membranes
It is important to remember that all membranous structures in cells are composed of two layers of phospholipid with associated proteins and other molecules. Furthermore, all of these membranous organelles can be converted from one form to another (figure 4.14). For example, the plasma membrane is continuous with the endoplasmic reticulum; as a cell becomes larger, some of the endoplasmic reticulum moves to the surface to become plasma membrane. Similarly, the nuclear membrane is connected to the endoplasmic reticulum. Remember also that the Golgi apparatus receives membrane- enclosed packages from the endoplasmic reticulum and produces lysosomes that combine with other membrane-enclosed structures and secretory vesicles that fuse with the plasma membrane. Thus, this entire set of membranes is constantly swapping pieces.
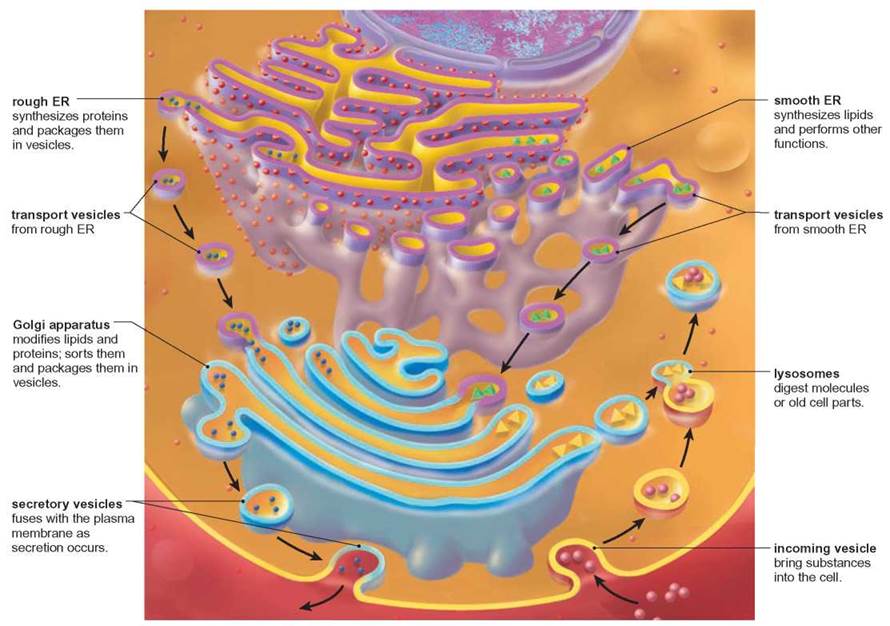
FIGURE 4.14. The Endomembrane System
Eukaryotic cells contain a variety of organelles composed of membranes that consist of two layers of phospholipids and associated proteins. Each organelle has a unique shape and function. Many of these organelles are interconverted from one to another as they perform their essential functions.
Energy Converters—Mitochondria and Chloroplasts
Two other organelles composed of membranes are mitochondria and chloroplasts. Both types of organelles are associated with energy conversion reactions in the cell. Mitochondria and chloroplasts are different from other kinds of membranous structures in four ways. First, their membranes are chemically different from those of other membranous organelles; second, they are composed of two layers of membrane—an inner and an outer membrane; third, both of these structures have ribosomes and DNA that are similar to those of bacteria; fourth, these two structures have a certain degree of independence from the rest of the cell—they have a limited ability to reproduce themselves but must rely on DNA from the cell nucleus for assistance. It is important to understand that cells cannot make mitochondria or chloroplasts by themselves. The DNA of the organelle is necessary for their reproduction.
Mitochondrion
The mitochondrion is an organelle that contains the enzymes responsible for aerobic cellular respiration. It consists of an outer membrane and an inner folded membrane. The individual folds of the inner membrane are known as cristae (figure 4.15a). Aerobic cellular respiration is the series of enzyme-controlled reactions involved in the release of energy from food molecules and requires the participation of oxygen molecules.
![]()
Some of the enzymes responsible for these reactions are dissolved in the fluid inside the mitochondrion and are made using DNA in the mitochondria (mDNA). Others are incorporated into the structure of the membranes and are arranged in an orderly sequence.
The number of mitochondria per cell varies from less than 10 to over 1,000 depending on the kind of cell. Cells involved in activities that require large amounts of energy, such as muscle cells, contain the most mitochondria. When cells are functioning aerobically, the mitochondria swell with activity. When this activity diminishes, though, they shrink and appear as threadlike structures. The details of the reactions involved in aerobic cellular respiration and their relationship to the structure of mitochondria will be discussed in chapter 6.
Chloroplast
The chloroplast is a membranous saclike organelle responsible for the process of photosynthesis. Chloroplasts contain the green pigment, chlorophyll, and are found in cells of plants and other eukaryotic organisms that carry out photosynthesis. The cells of some organisms contain one large chloroplast; others contain hundreds of smaller chloroplasts. Photosynthesis is a metabolic process in which light energy is converted to chemical bond energy. Chemical-bond energy is found in food molecules.
![]()
A study of the ultrastructure—that is, the structures seen with an electron microscope—of a chloroplast shows that the entire organelle is enclosed by a membrane. Inside are other membranes throughout the chloroplast, forming networks and structures of folded membrane. As shown in figure 4.15b, in some areas, these membranes are stacked up or folded back on themselves. Chlorophyll molecules are attached to these membranes and are called thylakoids. Thylakoids that are stacked on top of one another form the grana of the chloroplast. The space between the grana, which has no chlorophyll, is known as the stroma. The details of how photosynthesis occurs and how this process is associated with the structure of the chloroplast will be discussed in chapter 7.
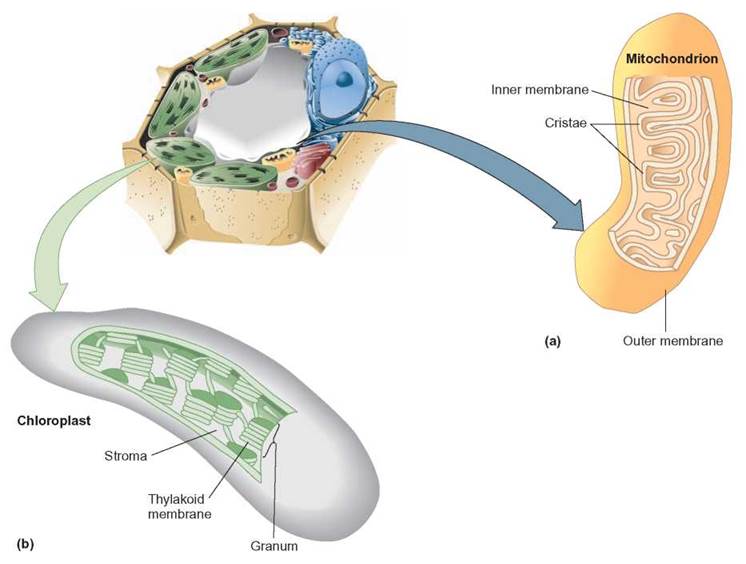
FIGURE 4.15. Energy-Converting Organelles
(a) Mitochondria, with their inner folds called cristae, are the site of aerobic cellular respiration, where food energy is converted to usable cellular energy. (b) Chloroplasts, containing the pigment chlorophyll, are the site of photosynthesis. The chlorophyll, located in the grana, captures light energy, which is used to construct organic, sugarlike molecules in the stroma.
4.4. CONCEPT REVIEW
7. List the membranous organelles of a eukaryotic cell and describe the function of each.
8. Define the following terms: stroma, grana, cristae.
9. Describe the functions of the plasma membrane.