Biology of Humans
22. Evolution and Our Heritage
In Chapter 21a we learned how mutations in certain genes lead to loss of control over cell division and the development of cancer. In this chapter, we consider mutations in a different light. We focus on evolution and how mutations, along with processes such as genetic drift, gene flow, and natural selection, cause changes in allele frequencies within populations. We also address questions such as how did life arise and evolve on Earth? How has evolution shaped species, including our own? What were our ancestors like?
Evolution of Life on Earth
Evolution can broadly be defined as descent with modification from a common ancestor. It is the process by which Earth's life-forms have changed from their earliest beginnings to today. But how did life first arise on Earth?
Earth is estimated to be 4.5 billion years old. Evidence from physical and chemical changes in Earth's crust and atmosphere suggests that life has existed on Earth for about 3.8 billion years. The environment of the early Earth was very different from that of today and would have been an extremely hostile place for most organisms (Figure 22.1). Earth's crust was hot and volcanic. Intense lightning and ultraviolet radiation struck Earth's surface, and there was almost no gaseous oxygen (O2) in the atmosphere. Scientists agree on the scarcity of oxygen, but debate the other components of the early atmosphere. Most models suggest that carbon dioxide (CO2), nitrogen (N2), and water (H2O) were present. Other gases including carbon monoxide (CO), hydrogen (H2), methane (CH4), sulfur dioxide (SO2), and hydrogen sulfide (H2S) may have been present. Once the crust cooled, water vapor condensed as rain, and runoff collected in depressions to form early seas.
· The fossil record provides evidence of evolution by documenting that life on Earth has changed over time.

FIGURE 22.1. Representation of the early Earth
How might life have evolved under these conditions? In the following sections, we present a plausible sequence of events that most scientists believe explains the origin of life on Earth. The sequence, known as chemical evolution, suggests that life evolved from chemicals slowly increasing in complexity over a period of perhaps 300 million years.
Small Organic Molecules
Scientists hypothesize that conditions of the early Earth favored the synthesis of small organic molecules from inorganic molecules. Specifically, the low-oxygen atmosphere of the primitive Earth encouraged the joining of simple molecules to form complex molecules. The low-oxygen atmosphere was important because oxygen attacks chemical bonds. Scientists further hypothesize that the energy required for the joining of simple molecules could have come from the lightning and intense ultraviolet (UV) radiation striking the primitive Earth. (UV radiation was likely more intense during those times than it is now because young suns emit more UV radiation than do mature suns, and the early Earth lacked an ozone layer to shield it from UV radiation.)
Stanley Miller and Harold Urey of the University of Chicago tested the hypothesis that organic molecules could be synthesized from inorganic ones in 1953. In their laboratory, these scientists re-created conditions presumed to be similar to those of the early Earth (Figure 22.2). Miller and Urey discharged electric sparks (meant to simulate lightning) through an atmosphere that contained some of the gases thought to be present in the early atmosphere. They generated a variety of small organic molecules, supporting the hypothesis that organic molecules could be synthesized from inorganic ones.
The atmospheric composition used in the Miller-Urey experiment differed somewhat from the composition currently favored by scientists who study characteristics of the early Earth. Even so, many simulations conducted by other scientists using different gas mixtures (and energy sources) have produced organic compounds in varying amounts. Taken as a whole, these experiments demonstrate that organic molecules can be synthesized from inorganic molecules.
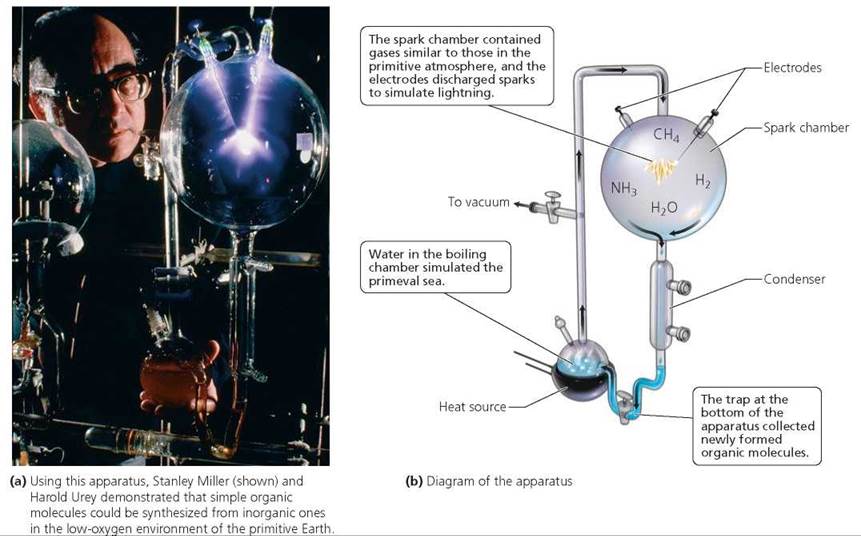
FIGURE 22.2. Testing the hypothesis that organic molecules could be synthesized from inorganic ones
Macromolecules
Scientists hypothesize that these small organic molecules accumulated in the early oceans and over a long period formed a complex mixture. Somehow, perhaps by being washed onto clay or hot sand or lava, the small molecules joined to form macromolecules, such as proteins and nucleic acids. Some scientists suggest instead that deep-sea vents were important locations for the synthesis of small organic molecules and their joining to form larger molecules.
Which macromolecule led to the formation of the first cells? Some scientists suggest that RNA was the critical macromolecule, because it can act as an enzyme and because its self-replicating abilities allowed the transfer of information from one generation to the next. Cells today store their genetic information as DNA. Thus, if RNA was the first genetic material, then DNA is likely to have evolved later from an RNA template. Other scientists suggest that proteins were the macromolecules that led to the first cells. These scientists cite the greater chemical stability of amino acids, peptides, and proteins, as compared to nucleotides and nucleic acids, when exposed to salt (such as would occur in primordial seas) and intense UV radiation (such as that presumed to have struck the early Earth).
Early Cells
Scientists hypothesize that the newly formed organic macromolecules aggregated into droplet-like structures that were the precursors of cells. The early droplets displayed some of the same properties as living cells, such as the ability to maintain an internal environment different from surrounding conditions.
Fossil evidence indicates that the earliest cells were prokaryotes. Recall from Chapter 3 that prokaryotic cells lack membrane- enclosed organelles, such as the nucleus, and are typically smaller than eukaryotic cells. These early prokaryotic cells relied on anaerobic metabolism (metabolism in the absence of oxygen). Eventually, some of these cells captured energy from sunlight and made their own complex organic molecules from CO2 and H2O in their environment. This process, known as photosynthesis, produced oxygen as a by-product. Oxygen began to accumulate in the environment. Next came cells that could use the now abundant oxygen to harness energy from stored organic molecules; these cells used cellular respiration (aerobic metabolism; see Chapter 3).
So the question arises, "How did more complex cells arise from these early prokaryotic cells?" It's possible that some of the organelles within eukaryotic cells appeared when other, smaller organisms became incorporated into the early, primitive cells. Mitochondria, for example, may be descendants of once free-living bacteria. These bacteria either invaded or were engulfed by the ancient cells and formed a mutually beneficial relationship with them. This idea is called the endosymbiont theory. It is generally accepted by the biological community to explain the origin of some features of eukaryotic cells, including mitochondria. Scientists currently are not certain whether endosymbiosis was a factor in the origin of other eukaryotic features such as the membrane-bound nucleus. An additional possibility is that infolding of the plasma membrane of an ancient prokaryote produced some of the organelles (for example, the endoplasmic reticulum or Golgi complex) found in eukaryotic cells of today.
Fossils of prokaryotic cells have been dated at 3.5 billion years old, and scientists estimate that the first prokaryotic cells probably arose around 3.8 billion years ago. As we discuss later in the chapter, fossils are rarely formed and found, so the origin for a group of organisms is usually hypothesized to occur earlier than the oldest fossil evidence for that group. Eukaryotic cells evolved about 1.8 billion years ago. Multicellularity evolved in eukaryotes around 1.5 billion years ago and eventually led to organisms such as plants, fungi, and animals. Figure 22.3 summarizes the possible steps in the origin of life on Earth.
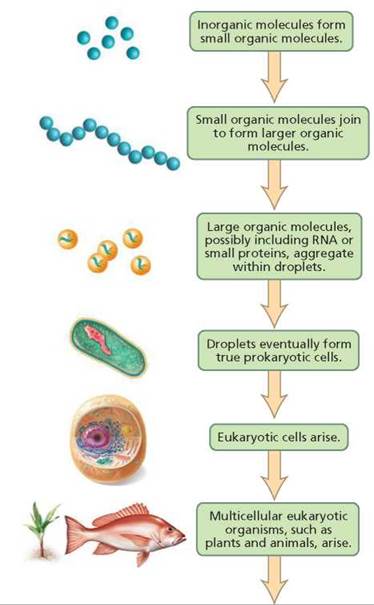
FIGURE 22.3. Possible steps in the origin of life on Earth
Scale of Evolutionary Change
Evolution occurs on two levels. One level, microevolution, is small; and the other, macroevolution, is large. Microevolution occurs through changes in allele frequencies within a population over a few generations. Macroevolution, on the other hand, consists of larger-scale evolutionary change over longer periods of time, such as the origin of groups of species (for example, mammals) and mass extinctions (the catastrophic disappearance of many species). We begin with microevolution, examining genetic variation within populations and describing the causes of microevolution. We then turn to the larger-scale phenomena of macroevolution.
Microevolution
Look around, and you will see variation in almost any group of individuals belonging to the same species. Consider your classmates. They do not all look alike, and, unless you are an identical twin, you probably do not precisely resemble your brothers or sisters. Before looking at what makes individuals in such a group different, let's define some basic terminology. A population is a group of individuals of the same species living in a particular area. A population of bluegill sunfish inhabits a pond, and a population of deer mice inhabits a small tract of forest. A gene pool consists of all the alleles of all the genes of all individuals in a population. (Recall from Chapter 20 that a gene is a segment of DNA on a chromosome that directs the synthesis of a specific protein, whereas alleles are different forms of a gene.) Now let's take a closer look at what makes individuals in a population different and examine some of the ways that variation can appear in populations.
Sexual reproduction and mutation produce variation in populations. Sexual reproduction shuffles alleles already present in the population. As discussed in Chapters 19 and 20, the gametes (eggs or sperm) of any one individual show substantial genetic variation due to crossing over and independent assortment during meiosis. Also, the combination of gametes that unite at fertilization is a chance event. Of the millions of sperm produced by a male, only one fertilizes the egg. This union produces a new individual with a new combination of alleles.
New genes and new alleles originate by mutation, a change in the nucleotide sequence of DNA. Mutations occur at a low rate in any set of genes, so their contribution to genetic diversity in large populations is quite small. Mutations can appear spontaneously from mistakes in DNA replication, or they can be caused by outside sources such as radiation or chemical agents. Only mutations in cell lines that produce eggs or sperm can be passed to offspring.
Recall that microevolution involves changes in the frequency of certain alleles relative to others within the gene pool of a population. Some of the processes that produce those changes are genetic drift, gene flow, mutation, and natural selection.
Genetic drift Genetic drift occurs when allele frequencies within a population change randomly because of chance alone. This process is usually negligible in large populations. However, in populations with fewer than about 100 individuals, chance events can cause allele frequencies to drift randomly from one generation to the next. Two mechanisms that facilitate genetic drift in natural populations are the bottleneck effect and the founder effect.
Sometimes, dramatic reductions in population size occur because of natural disasters that kill many individuals at random. Consider a population that experiences a flood in which most members die. With fewer remaining individuals contributing to the gene pool, the genetic makeup of the survivors may not be representative of the original population. This change in the gene pool is the bottleneck effect, so named because the population experiences a dramatic decrease in size—much as the size of a bottle decreases at the neck. Certain alleles may be more or less common in the flood survivors than in the original population simply by chance. In fact, some alleles may be completely lost, resulting in reduced overall genetic variability among survivors.
Genetic drift also occurs when a few individuals leave their population and establish themselves in a new, somewhat isolated place. By chance alone, the genetic makeup of the colonizing individuals is probably not representative of the full gene pool of the population they left. Genetic drift in new, small colonies is called the founder effect.
Stop and think
Would you expect the founder effect to be associated with a relatively high or a relatively low frequency of inherited recessive disorders?
Gene flow. Another cause of microevolution is gene flow, which occurs when individuals move into and out of populations. As individuals come and go, they carry with them their unique sets of genes. Gene flow occurs if these individuals successfully interbreed (mate and produce offspring) with the resident population, adding to the gene pool.
The cessation of gene flow can be important to the formation of new species. A species is a population or group of populations whose members are capable of successful interbreeding. Such interbreeding must occur under natural conditions and produce fertile offspring. But consider what happens when a population becomes geographically isolated from other populations of the same species. For example, over the course of geologic time, sea level fluctuates. During periods when sea level decreases, a previously offshore island may reconnect with a continent. When sea level increases, the island will redevelop offshore. Suppose that during the period of low sea level a continuous population of frogs extends across the landscape. As sea level rises, however, and the island redevelops, the island population of frogs will be isolated from the one on the mainland. Frogs cannot cross saltwater, so the island population is effectively genetically isolated from the mainland population. The island population may take a separate evolutionary route as distinctly different sets of allele frequencies and mutations accumulate. Eventually, the isolated island population may become so different that it cannot successfully interbreed with the mainland population. At this point, there are two species of frogs instead of one. This process is called speciation.
Mutation. Mutations, you will recall, are rare changes in the DNA of genes; they are the third way that the frequencies of certain alleles can change relative to others within gene pools. Mutations produce new alleles that, when transmitted in gametes, cause an immediate change in the gene pool. Essentially the new (mutant) allele is substituted for another allele. Like gene flow, mutation can introduce new alleles to a population that are then acted upon by natural selection. If the frequency of the mutant allele increases in a population, it is not because mutations are suddenly occurring more frequently. Instead, possession of the mutant allele might confer some advantage that enables individuals with the mutant allele to produce more offspring than can those without it. In other words, the increased frequency in a population of the mutant allele relative to others results from natural selection, our next focus.
Natural selection. In his book On the Origin of Species (1859), Charles Darwin argued that species were not specially created, unchanging forms. He suggested that modern species are descendants of ancestral species. In other words, present-day species evolved from past species. Darwin also proposed that evolution occurred by the process of natural selection. His ideas can be summarized briefly as follows:
1. Individual variation exists within a species. Some of this variation is inherited.
2. Some individuals have more surviving offspring than do others because their particular inherited characteristics make them better suited to their local environment; this is the process of natural selection.
3. Evolutionary change occurs as the traits of individuals that survive and reproduce become more common in the population. Traits of less successful individuals become less common.
According to Darwin's ideas, an individual's evolutionary success can be measured by fitness (sometimes called Darwinian fitness). Fitness compares the number of reproductively viable offspring among individuals. Individuals with greater fitness— that is, producing more successful offspring—have more of their genes represented in future generations. To succeed in terms of evolution, one must reproduce (Figure 22.4). Indeed, you could live more than 100 years, but your individual fitness would be zero if you did not reproduce. Some of the diseases or conditions discussed in earlier chapters are associated with zero fitness because they cause sterility (such as Turner's syndrome) or death before reproductive maturity (such as Tay-Sachs disease).

FIGURE 22.4. Fitness is the number of offspring left by an individual. These parents clearly have high fitness.
One result of natural selection is that populations become better suited to their particular environment. This transformation of the population toward better fitness in their environment is called adaptation. If the environment changes, however, the individuals who have become most finely attuned to the initial environment could lose their advantage, and other individuals with different alleles might be selected by nature to leave more offspring. If the environment once again stabilizes, the population reestablishes itself around a new set of allele frequencies that better meet the new environmental conditions.
The evolution of antibiotic resistance in bacteria provides an example of natural selection in action. Consider what can happen when you take an antibiotic to treat a bacterial infection. Many bacteria are killed by the treatment, but some may survive. Surviving bacteria have genes that confer resistance to the antibiotic. The survivors reproduce and pass on the trait for resistance to future generations of bacteria. Over time, antibiotic resistance becomes more common in bacterial populations than before. The evolution of drug-resistant strains can potentially become a serious threat to humankind (Chapter 13a).
Natural selection does not lead to perfect organisms. It can act on available variation only, and the available variation may not include the traits that would be ideal. Also, natural selection can only modify existing structures; it cannot produce completely new and different structures from scratch. Finally, organisms have to do many different things to survive and reproduce—such as escaping from predators and finding food, shelter, and mates—and it simply is not possible to be perfect at everything. Indeed, adaptations often are compromises between the many competing demands the organism faces.
Macroevolution
Large-scale evolutionary change is macroevolution. Whereas microevolution involves changes in the frequencies of alleles within populations, macroevolution produces changes in groups of species, such as might occur with major changes in climate. Our discussion of macroevolution begins with a description of how species are named. We then consider how their evolutionary histories can be analyzed and diagrammed.
Scientific names. Systematic biology deals with the naming, classification, and evolutionary relationships of organisms. A universal system for naming and classifying organisms is essential for communicating information about them. Scientists use the naming system that Swedish naturalist Carl Linnaeus developed more than 200 years ago. Each organism is given a Latin binomial, or two-part name, consisting of the genus name followed by the specific epithet (the term specific, here, means relating to species). For example, humans are in the genus Homo, and our specific epithet is sapiens, so our binomial is Homo sapiens. By convention, the genus name and specific epithet are italicized, the first letter of the genus name is always capitalized, and the specific epithet is all lowercase. Sometimes, the genus name is abbreviated: H. sapiens.
Linnaeus also developed a system for classifying organisms using a series of increasingly broad categories: species, genus, family, order, class, phylum, and kingdom. Similar species were placed in the same genus; similar genera (the plural of genus) were placed in the same family; similar families in the same order, and so on. Above the kingdom level, scientists have added the category domain to Linnaeus's scheme (the three domains are Archaea, Bacteria, and Eukarya; see Chapter 1). Sometimes the Linnaean categories have subdivisions; for example, subphyla within phyla. Figure 22.5 presents the categories humans belong to.
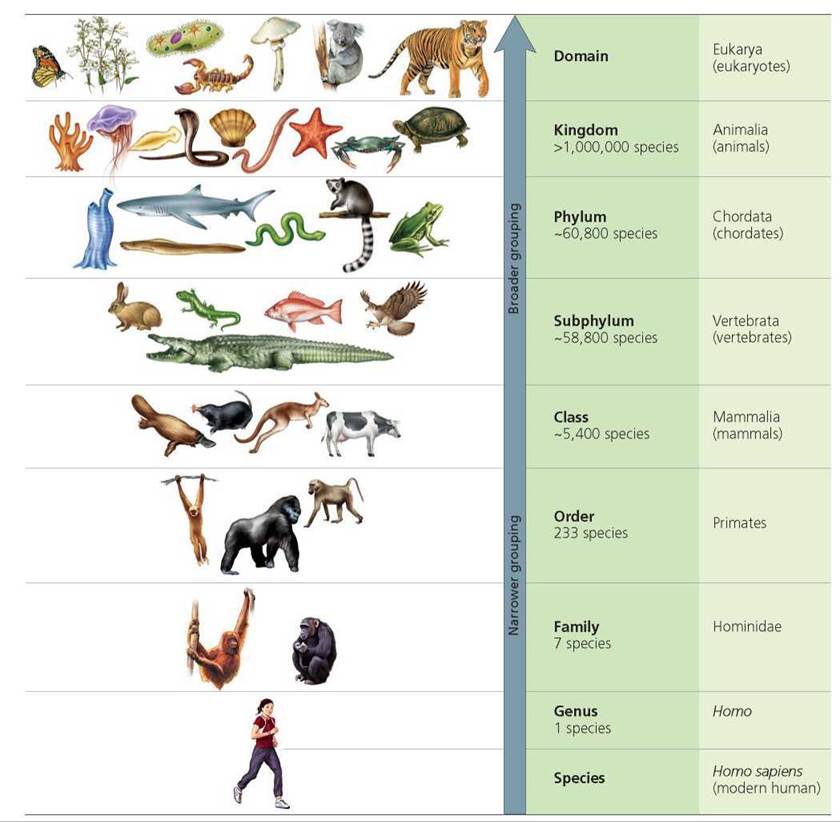
FIGURE 22.5. Categories in the classification of living organisms. Any organism, including a human being, can be classified using a hierarchy of increasingly general categories. Modern humans are called Homo sapiens. We are placed in the family Hominidae, along with gorillas (2 species), chimpanzees (2 species, common chimpanzee and bonobo), and orangutans (2 species). We are the only living species in our genus (Homo).
Phylogenetic trees. Phylogenetic trees are branching diagrams used by scientists to depict hypotheses about evolutionary relationships among species or groups of species. Such trees can illustrate in simple graphic form concepts that are difficult to express in words.
Scientists developing hypotheses about these relationships may begin by creating a character matrix, such as the example in Figure 22.6a. The character matrix can then be used to construct a phylogenetic tree. Typical matrices consist of vertical columns representing species or other groups, and horizontal rows representing "characters" that are either present or absent in those species or groups. Figure 22.6a shows a simple matrix with a shark (fish), frog (amphibian), and human (mammal) being compared regarding the presence or absence of three characters: two paired appendages (fins or limbs), individual digits, and hair. A phylogenetic tree based on the results from the matrix is shown in Figure 22.6b. This tree suggests that humans and frogs have more in common with one another (two paired appendages and individual digits) than either has with sharks (two paired appendages), and thus humans and frogs are likely to share a more recent common ancestor. The tree also shows that humans differ from frogs in having hair.
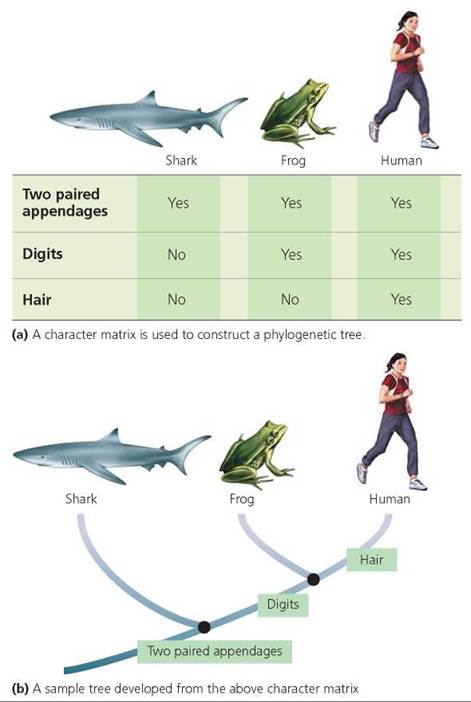
FIGURE 22.6. A phylogenetic tree depicts hypotheses about evolutionary relationships among organisms.
Evidence of Evolution
We know that evolution has occurred throughout Earth's history because the physical evidence of evolution surrounds us. Such evidence comes from many sources, including the fossil record, biogeography, the comparison of anatomical and embryological structures, and molecular biology.
Fossil Record
Earth is littered with silent relics of organisms that lived long ago (Figure 22.7). We find, for example, tiny spiders preserved in resin that dripped as sticky sap from some ancient tree. We find mineralized bones and teeth, hardened remains that tell us much about the ancestry of today's vertebrates (animals with a backbone). We also find impressions, such as footprints, of organisms that lived in the past. These preserved remnants and impressions of past organisms are fossils. Most fossils occur in sedimentary rocks, such as limestone, sandstone, shale, and chalk. These rocks form when sand and other particles settle to the bottoms of rivers, lakes, and oceans; accumulate in layers; and harden. Fossils also occur in volcanic ash, tar pits, and a few other specialized conditions.

FIGURE 22.7. A sampling of past life in fossils
Fossilization is any process by which fossils form (Figure 22.8). In a typical case, an organism dies and settles to the bottom of a body of water. If not destroyed by scavengers, the organism is buried under accumulating layers of sediment. Soft parts usually decay. Hard parts such as bones, teeth, and shells may be preserved if they become impregnated with minerals from surrounding water and sediments. As new layers of sediment are added, the older (lower) layers solidify under the pressure generated by overlying sediments. Eventually, if the sediments are uplifted by geological processes and the water disappears, weather may erode the surface of the rock formation and expose the fossil.
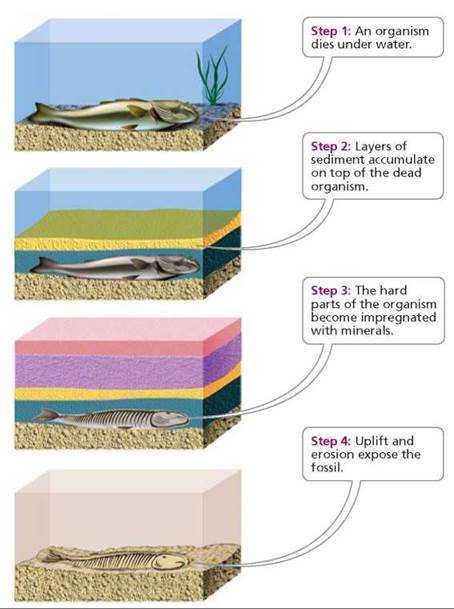
FIGURE 22.8. A typical sequence for fossilization
Fossils provide strong evidence of evolution. Fossils of extinct organisms show both similarities to and differences from living species. Similarities to other fossil and modern species are used to assess degrees of evolutionary relationships. Often, fossils reveal combinations of features not seen in any living forms. Such combinations help us understand how major new adaptations arose. Sometimes we are lucky enough to find transitional forms that closely link ancient organisms to modern species. Consider several fossil whales discovered in the last 30 or so years; these fossils document a progression from terrestrial forms with fore and hind limbs to more aquatic forms with reduced hind limbs to modern whales, which are fully aquatic and have no hind limbs (Figure 22.9a). Additionally, some fossil whale remains have been discovered that have an anklebone with a diagnostic shape; the bone is the astragalus, and the shape is that of a double pulley (Figure 22.9b). The double pulley astragalus is characteristic of artiodactyls (an order of hoofed mammals that includes hippos, deer, cows, and pronghorn). Discovery of this anklebone with a double pulley shape in fossil whales provides strong support for a close phylogenetic relationship between whales and artiodactyls.

FIGURE 22.9. The evolution of whales as revealed by transitional fossils
Radiometric dating can be used to obtain estimates of the absolute ages of rocks and fossils. This technique relies on measuring the proportions of a radioactive isotope and its decay product. For example, radioactive potassium decays at a constant rate to form argon, so the ratio of radioactive potassium to argon in a fossil can be used to estimate the absolute age of the fossil. The relative ages of fossils can be determined because fossils found in deeper layers of rock are typically older than those found in layers closer to the surface. These kinds of observations enable scientists to piece together the chronological emergence of different kinds of organisms. For example, fishes are the first fossil vertebrates to appear in deep (old) layers of rock. Above those layers, fossils of amphibians appear, then reptiles, then mammals, and finally birds. This chronological sequence for the appearance of the major groups of vertebrates has been supported by other lines of evidence, some of which we consider later in this section.
Although the fossil record tells us much about past life, it has limitations. First, fossils are relatively rare. When most animals or plants die, their remains are eaten by predators or scavengers or are broken down by microorganisms, chemicals, or mechanical processes. Even if a fossil should form, the chances are small that it will be exposed by erosion or other forces and not be destroyed by those same forces before it is discovered. Second, the fossil record represents a biased sampling of past life. Aquatic plants and animals have a much higher probability of being buried in deep sediment than do terrestrial organisms. Thus preservation of aquatic organisms is more likely. Large animals with a hard skeleton are far more likely to be preserved than are small animals with soft parts. Organisms from large, enduring populations are more likely to be represented in the fossil record than are those from small, quickly disappearing populations. Despite these limitations, fossils document that life on Earth has not always been the same as it is today. The simple fact of these changes is potent evidence of evolution.
Geographic Distributions
Biogeography is the study of the geographic distribution of organisms. Geographic distributions often reflect evolutionary history and relationships because related species are more likely than are unrelated species to be found in the same geographic area. A careful comparison of the animals in a given place with those occurring elsewhere can yield clues about the relationship of the groups. If groups of animals have been separated, biogeography can tell us how long ago the separation occurred.
For example, today we find many species of marsupials— mammals such as opossums and kangaroos—in Australia but only a few in North and South America. The presence of so many species of marsupials in Australia suggests that they arose from distant ancestors whose descendants were not replaced by animals arriving from other regions. New distributions of organisms occur by two basic mechanisms. In one mechanism, the organisms disperse to new areas. In the other mechanism, the areas occupied by the organisms move or are subdivided. Australia is an island, remote from other major continental landmasses. The evolutionary history of the Australian marsupials involves both dispersal and the movement of continents. Fossil evidence suggests that marsupials evolved in the Northern Hemisphere (probably North America) and that some dispersed southward into South America, then to Antarctica, and later to Australia, to which Antarctica was attached at the time (Figure 22.10). As Australia and other landmasses slowly shifted to form the modern continental arrangement, the ancestors of today's marsupials were carried away from their place of origin to evolve in isolation in Australia.
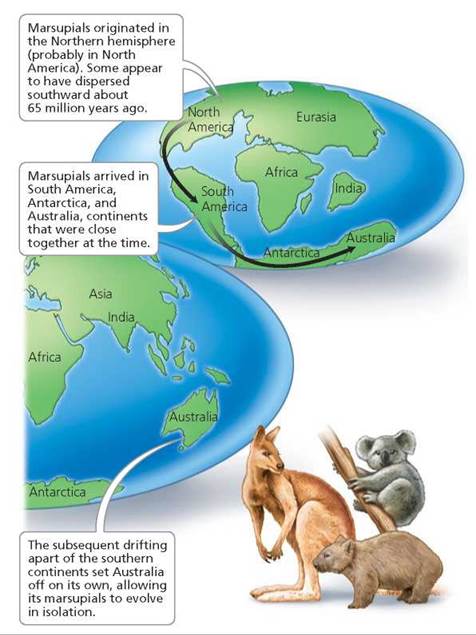
FIGURE 22.10. The story of marsupials and Australia involves both dispersal and movement of continents.
Ethical Issue
Conducting Research on Our Relatives
Chimpanzees look and behave somewhat differently than we do.
Still, it is hard to look into their eyes and not see something of ourselves. Chimpanzees are our closest living relatives, sharing a remarkably high percentage of our DNA sequence. Nevertheless, we use them and other nonhuman primates in invasive scientific research that might benefit us. Is this ethical?
Using nonhuman primates in research is costly and controversial. Even so, they often are preferred as subjects because they are so similar to humans. For example, human and nonhuman primates possess brains with similar organization, develop comparable plaques in their arteries, and experience many of the same changes in anatomy, physiology, and behavior with age. In some cases, Nobel Prize-winning research has resulted from the contributions of research with nonhuman primates, including development of vaccines for yellow fever (1951) and polio (1954) and insight into how visual information is processed in the brain (1981). Research with nonhuman primates has also led to significant advances in our understanding of Alzheimer's disease, AIDS, and severe acute respiratory syndrome (SARS). The care and use of nonhuman primates (and other vertebrate animals) in research is regulated by federal agencies such as the Public Health Service and the U.S. Department of Agriculture. Animal research also is regulated at the local level. Each college, university, or research center has an Institutional
Animal Care and Use Committee whose members include veterinarians, researchers, and members of the public. In addition to federal and local oversight, scientists and animal care personnel are striving to improve housing for captive nonhuman primates and to consider their psychological well-being. Even with such efforts, controversy and questions remain.

Questions to Consider
• Should we ban the use of nonhuman primates in medical research? If we do, will progress in fighting diseases such as AIDS and Alzheimer's be slowed? If you or a loved one had one of these illnesses, would you feel differently about research using nonhuman primates?
• Nonhuman primates represent a fraction of the animals used in research. More than 90% of research animals are rodents, such as rats, mice, and guinea pigs. Where would you draw the line when deciding which (if any) animals are acceptable for use in research that might benefit us?
Comparative Anatomy and Embryology
Comparative anatomy, as its name suggests, is the comparison of the anatomies (physical structures) of different species. Common, or shared, traits among different species have long been considered a measure of relatedness. Put simply, two species with more shared traits are considered more closely related than are two species without shared traits (this principle underlies the use of character matrices to construct phylogenetic trees). For example, many very different vertebrates share similarities in the bones of their forelimbs, showing that they have an ancestor in common (Figure 22.11). Structures that are similar and that probably arose from a common ancestry are called homologous structures. The forelimbs that support bird wings and bat wings are homologous structures. Sometimes, however, similarities are not inherited from a common ancestor. For example, bird wings and insect wings both permit flight, but they are made from entirely different structures. Whereas bird wings consist of forelimbs, insect wings are not true appendages; they are extensions of the insect's cuticle (exoskeleton). Thus, the wings of birds and insects do not reflect common ancestry. Instead, birds and insects independently evolved wings because of similar ecological roles and selection pressures, in a process known as convergent evolution. Structures that are similar because of convergent evolution are called analogous structures.
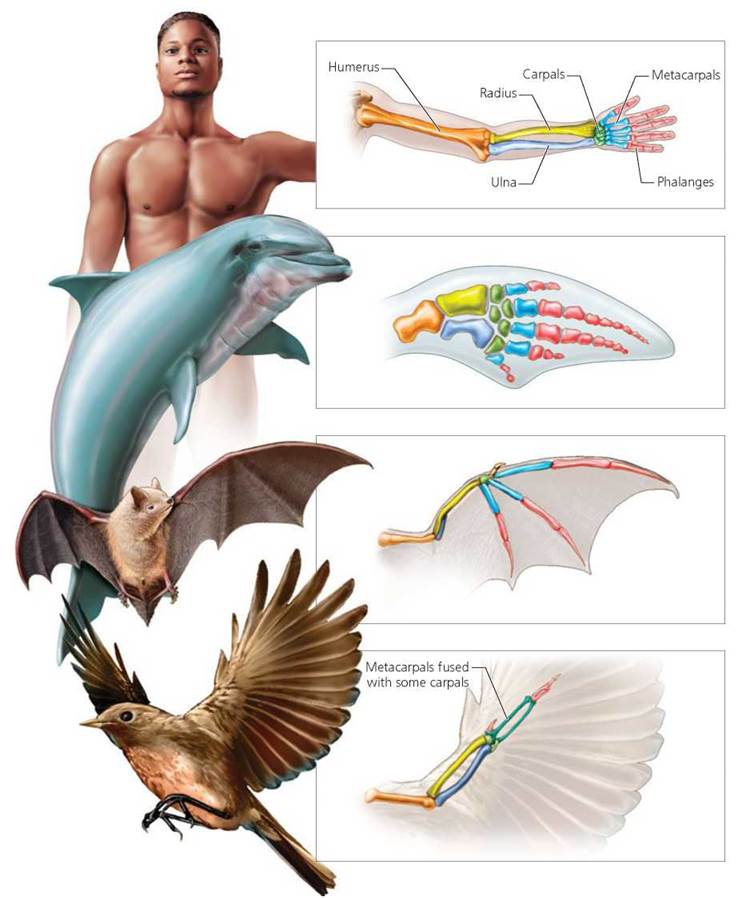
FIGURE 22.11. Homologous structures. The similarity of the forelimb bones of humans, dolphins, bats, and birds suggests that these organisms share a common ancestry.
Homologous structures arise from the same kind of embryonic tissue. Hence, comparative embryology, the comparative study of early development, also can be a useful tool for studying evolution. Common embryological origins can be considered evidence of common descent. For example, 4-week-old human embryos closely resemble embryos of other vertebrates, including fish. Indeed, human embryos at 4 weeks gestation come complete with a tail and gill pouches, as shown in Figure 22.12. As development proceeds, the gill pouches of fish develop into gills. The gill pouches of humans develop into other structures such as the auditory tubes connecting the middle ear and throat. Nevertheless, the fact that human, fish, and all other vertebrate embryos look very similar at early stages of development indicates common descent from an ancient ancestor.
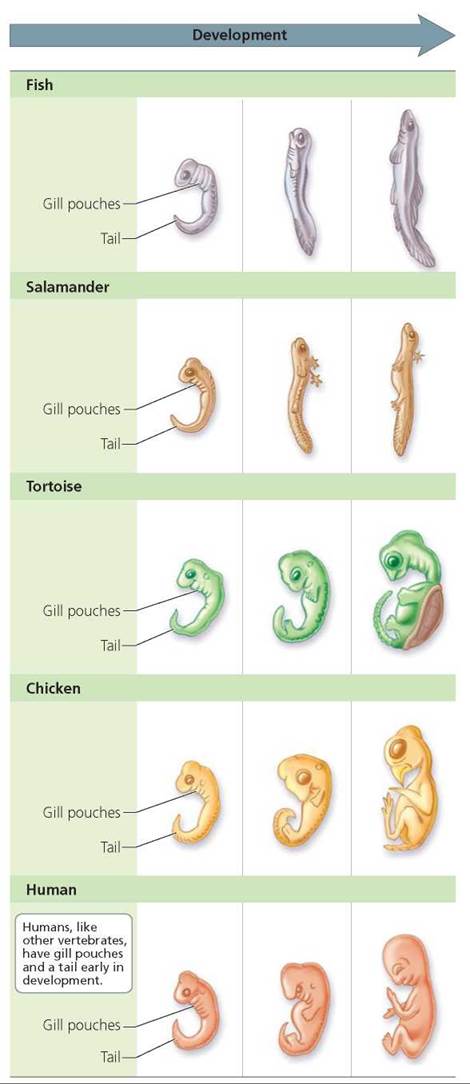
FIGURE 22.12. Resemblance early In development Indicates common descent. Embryos In the different representative vertebrates are not to scale; they have been drawn to the same approximate size to permit comparison.
Comparative Molecular Biology
Just as visible traits such as forelimbs can be compared, so can the molecules that are the basic building blocks of life. For example, scientists can compare the sequences of amino acids in proteins or the nucleotide sequences in DNA. As described in Chapter 21, the Human Genome Project has provided information on the location of genes along our chromosomes and the order of the base pairs that make up our chromosomes. More recently, the chimpanzee's genome has been described and compared with that of humans. Such comparison reveals that the two genomes are strikingly similar; for example, the DNA sequence that can be directly compared between chimp and human genomes is about 99% identical.
Because of background radiation and errors in copying DNA, single-nucleotide changes in DNA, called point mutations, occur constantly over evolutionary time with clocklike regularity. The rates of these changes vary from gene to gene. Once calibrated against the fossil record, these molecular clocks allow scientists to compare DNA sequences in two species as a way of estimating the amount of time that has passed since the two species diverged from a single common ancestor. The more different the sequences, the more time that has elapsed since their common ancestor. For example, comparison of DNA sequences tells us that humans and chimpanzees diverged from a common ancestor about 6 million years ago, making chimpanzees our closest living relatives. (See the Ethical Issue essay, Conducting Research on Our Relatives, on page 478.)
We have learned about microevolution, macroevolution, and the evidence for evolution. Now let's look at our own past and see how humans evolved.
Human Evolution
We begin our discussion of human evolution with the primates, an order of mammals that includes humans, apes, monkeys, and related forms (such as lemurs). Paleontologists (scientists who study fossils) believe that primates evolved about 65 million years ago. This estimate takes into account that the oldest primate fossils are about 55 million years old, and as mentioned previously, organisms are assumed to evolve earlier than when they first appear in the fossil record. In contrast, molecular biologists have suggested that primates evolved about 90 million years ago. Their estimate is based on comparison of DNA sequences. Despite debate over when primates evolved, most scientists agree that the first primates probably arose from an arboreal (tree-living) mammal that ate insects. This ancestor to primates might have looked something like a modern tree shrew (Figure 22.13).

FIGURE 22.13. A large tree shrew. These modern animals resemble the arboreal, insect-eating mammals from which primates probably evolved more than 65 million years ago.
Primate Characteristics
Primates have several distinguishing characteristics. Many of these characteristics reflect an arboreal lifestyle specialized for the visual hunting and manual capture of insects. For example, primates have flexible, rotating shoulder joints and exceptionally mobile digits with sensitive pads on their ends. Flattened nails replace claws. In many primate species, the big toe is separated from the other toes, and thumbs are opposable to other fingers. Thus, primates have grasping feet and hands, features that help in the pursuit and capture of insects along branches. In addition, a complex visual system (forward-facing eyes with stereoscopic vision) and a large brain relative to body size provide the well-developed depth perception, hand-eye coordination, and neuromuscular control needed by arboreal insectivores. The relatively large brain is also associated with complex social behavior (for example, members of social groups may form long-term alliances) and reliance on learned behavior (for example, tool use may be passed from one individual to the next through observation and imitation). Also, most primates give birth to only one infant at a time and provide extensive parental care—these characteristics may reflect the difficulty of carrying and rearing multiple infants in trees.
Modern primates are divided into two main groups (suborders). One suborder contains lemurs, lorises, and pottos, grouped together because they retain ancestral primate features such as small body size and nocturnal habits (Figure 22.14). The other suborder contains monkeys, apes, and humans (Figure 22.15).

FIGURE 22.14. Examples of modern primates from the suborder whose members retain ancestral features. The female loris with young in part (b) illustrates many of the features characteristic of all primates, including grasping hands and feet, forward-facing eyes, and small litter size.

FIGURE 22.15. Monkeys (such as spider monkeys and baboons) and apes (such as orangutans and gorillas) are placed with humans in another suborder.
In the past, the term hominid was used to describe members of the human lineage, such as species in the genera Australopithecus and Homo (discussed later in this chapter). At that time, human and prehuman species were the only members of the family Hominidae. Primate classification has changed, however, and apes now are included in the family Hominidae (refer again to Figure 22.5). Thus, the term hominid now include apes and humans (members of the family Hominidae). The term hominin now is used for the human lineage and its immediate ancestors (members of the subfamily Homininae). As described earlier, molecular evidence suggests that the lines leading to modern humans and chimpanzees diverged about 6 million years ago. Molecular data further indicate that, after chimpanzees, gorillas are our next closest living relatives, followed by orangutans and then gibbons. Figure 22.16 shows these hypothesized relationships among living primates.
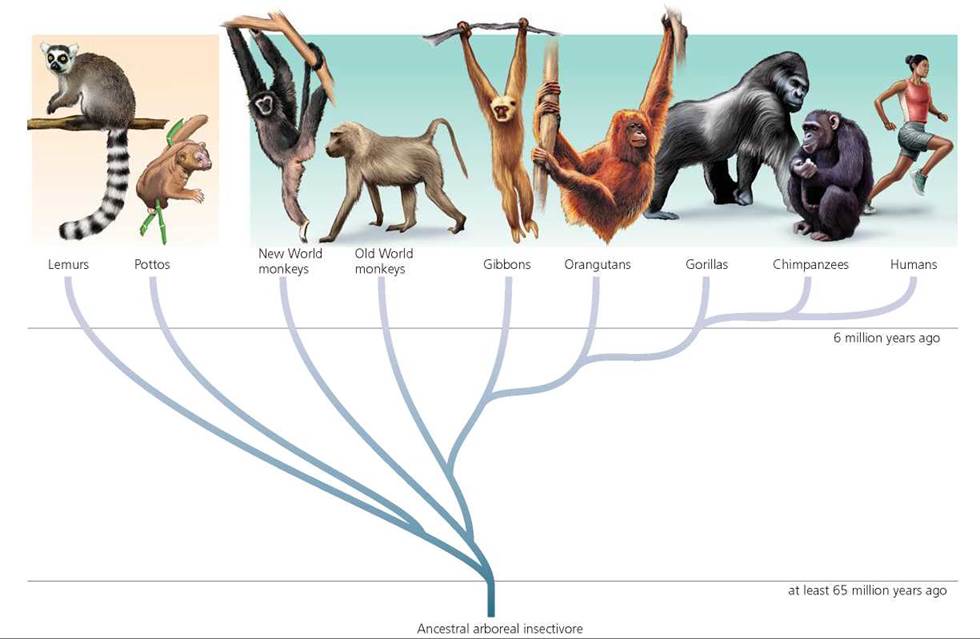
FIGURE 22.16. Hypothesized relationships among extant primates. (Figures are not drawn to scale.)
Comparison of human and chimp skeletal anatomy. Humans are the most terrestrial of primates. Many aspects of our skeletal anatomy reflect this terrestrial lifestyle and our upright stance while walking. Walking on two feet is called bipedalism. Our S-shaped spine and relatively large patella (kneecap) reflect a bipedal gait. Although chimpanzees can walk on two feet, they typically use quadrupedal knuckle-walking when moving on the ground. Their hand bones are more robust than ours because they bear weight during terrestrial locomotion. Chimps also spend time in trees, where they use their arms in locomotion. Thus, the arms of chimps are long and exceptionally strong; the strength is reflected in the extensive areas on the scapula (shoulder blade) for the attachment of large arm muscles. Unlike humans, chimps have opposable big toes, another adaptation for climbing. Anatomical differences between chimpanzees and humans also are apparent in the skull and teeth. Chimps have a smaller braincase than humans, but more pronounced brow ridges and jutting jaws. The degree of sexual dimorphism (difference in appearance between the sexes) in canine teeth also is more pronounced in chimps than in humans; specifically, the canines are larger in males than in females in both species, but the sex difference is greater in chimps. Figure 22.17 shows some of the major differences between chimps and humans in their teeth and skeleton.
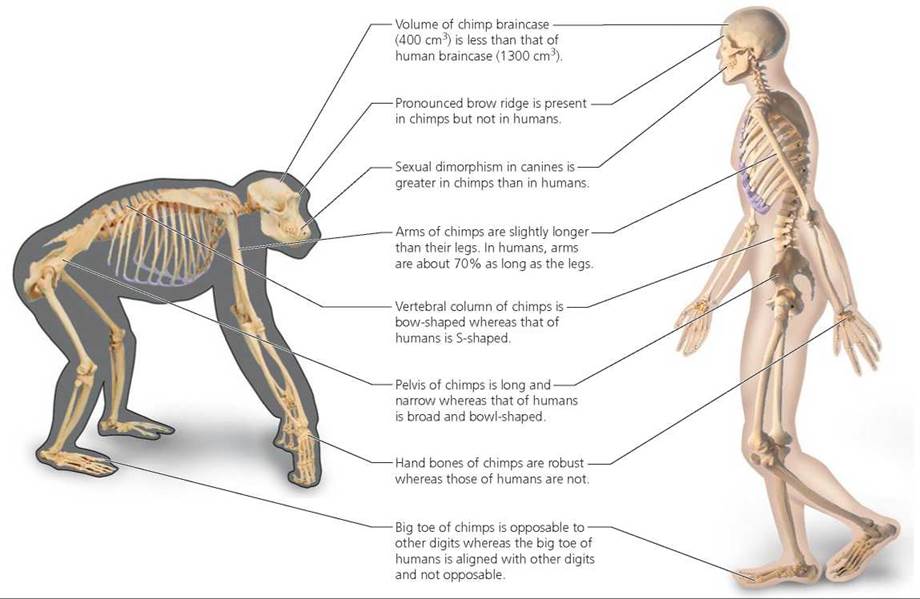
FIGURE 22.17. Some major differences in skeletal anatomy and teeth between chimpanzees and humans. Many of these traits reflect differences in locomotion and stance. Whereas chimpanzees are quadrupedal knuckle-walkers, humans are bipedal with an upright stance.
Stop and think
In the following discussion of our ancestry, we describe the species that led to us. At some point the line became us. Before you start reading about our ancestry, what qualities would you now say are necessary for a species to qualify as human?
Misconceptions
Several popular misconceptions about human evolution exist among nonscientists. One misconception is the idea that we descended from chimpanzees or any of the other modern apes. Humans and chimpanzees represent separate phylogenetic branches that diverged about 6 million years ago. Thus the common ancestor of humans and chimpanzees was different from any modern species of ape.
Another misconception is that modern humans evolved in an orderly, stepwise fashion. We often see such a stepwise progression depicted in drawings, sometimes humorously, and its appeal lies in its simplicity. However, as is so often the case, the real story is far more complex. The path to modern humans has been fraught with unsuccessful phenotypes leading to dead end after dead end. In fact, the path looks more like a family "bush" than an orderly progression from primitive to modern.
A final misconception is that, over the course of human evolution, the various bones and organ systems evolved simultaneously and at the same rate. They did not. There is no reason to believe that the human brain evolved at the same rate as, say, the appendix or the foot. Instead, different traits evolved at different times and rates, by a phenomenon known as mosaic evolution.
Trends in Hominin Evolution
Several evolutionary trends are apparent in the history of hominins. Bipedalism evolved early and probably set the stage for the evolution of other characteristics, such as increases in brain size. Cultural developments, such as tool use and language, are linked to increases in brain size. Once the hands were freed from the requirements of locomotion, they could be used for tasks such as making tools. Evidence that bipedalism preceded increases in brain size and cultural developments comes from fossilized hominin footprints found in Tanzania, Africa. These footprints, estimated to be about 3.6 million years old, were apparently made by two adults and a child (Figure 22.18). The footprints clearly predate the oldest stone tools from 2.6 million years ago. Other changes associated with upright posture include the S-shaped curvature of the vertebral column (the lumbar curve); modifications to the bones and muscles of the pelvis, legs, and feet; and positioning of the skull on top of the vertebral column (refer again to the human skeleton in Figure 22.17).

FIGURE 22.18. These hominin footprints from Laetoli, Tanzania, predate the oldest known tools and thus provide evidence that bipedalism preceded increases in brain size and cultural trends such as toolmaking. The larger prints were made by two individuals, one following in the other’s footsteps. The smaller prints may have been made by a child walking with the two individuals.
The faces of hominins also changed. For example, the forehead changed from sloping to vertical, and sites of muscle attachment, such as the brow ridges and crests on the skull, became smaller. The jaws became shorter, and the nose and chin more prominent. The overall size difference between males and females decreased. Males of our early ancestors appear to have been 1.5 times as heavy as females. Modern human males weigh about 1.2 times as much as females.
In the following discussion of our ancestors, we focus on the hominins that we know the most about—those in the genus Australopithecus and the genus Homo. We also mention some recent finds of apparently older hominins. Keep in mind that as new hominin fossils are found and genetic studies conducted, the dates for the origin of some species may be pushed back.
Australopithecines. The first hominin remains discovered were given the genus name Australopithecus, meaning "southern ape." Species within Australopithecus are sometimes collectively termed australopithecines. Australopithecus anamensis, considered the earliest species of the australopithecine line, is known from a small number of fossils found in Kenya and Ethiopia and dated between 4.2 and 3.9 million years old. The most spectacular australopithecine fossil found to date is that of a young adult female of the species Australopithecus afarensis (because she was found in the Afar region of Ethiopia). She was named Lucy by the scientists who discovered her in 1974. More than 60 pieces of Lucy's bones were found; when the bones were arranged, scientists estimated that, at death, she was about 1 m (3 ft) tall and weighed about 30 kg (66 lb; Figure 22.19). Her bones were determined to be 3.2 million years old. As more remains were found, it became apparent that the males of Lucy's species were somewhat taller (about 1.5 m, or about 5 ft) and heavier (about 45 kg, or 99 lb). The brain of A. afarensis was similar in size to that of modern chimpanzees or gorillas—about 430 cm3 (26 in.3). Although many aspects of the anatomy of A. afarensis suggest adaptations for living in trees, the remains also indicate bipedal- ism. In A. afarensis, we see an example of mosaic evolution— bipedalism evolving before substantial increases in brain size.
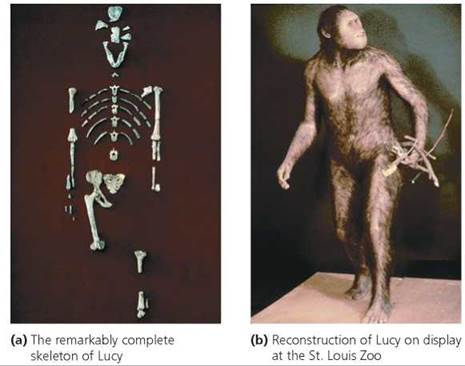
FIGURE 22.19. Fossilized remains and reconstruction of Lucy, a young female of the hominin species Australopithecus afarensis. She was named Lucy because the Beatles’ song “Lucy in the Sky with Diamonds" was playing the night Donald Johanson and his coworkers celebrated her discovery.
In 1994, researchers found hominin remains in Ethiopia that were older than those of A. anamensis. Dated at 4.4 million years old, the fossils were assigned to the species Ardipithecus ramidus. The remains, which included parts from more than 30 individuals, took 15 years to fully excavate and analyze. The most striking find was a partial skeleton (125 pieces) of a female now called Ardi. Eleven papers published in 2009 detailed the anatomy of A. ramidus and its implications for human evolution. Because the shape of Ardi's pelvis indicated that it was good for both climbing and upright walking, the research team suggested that A. ramidus displayed facultative bipedalism. In other words, when moving along tree branches Ardi may have walked upright atop the branches as well as climbed with all four limbs. They further propose that she walked upright on the ground, but not as well as later hominins. It is possible that Ardipithecus gave rise to the genus Australopithecus, which most scientists think led to our own genus, the genus Homo. It also is possible that Ardipithecus was a side branch, not along the path that led to us.
Stop and think
Brain size and shape can be estimated by making an endocast, a model of the inside of the skull. Such reconstructions are possible even when only a few parts of the skull are discovered and reassembled. Endocasts reveal general brain anatomy and size, allowing scientists to draw tentative conclusions about the cognitive capabilities of the skull's owner. Endocasts do not, however, capture the folds or convolutions on the surface of a brain. Do you think endocasts and brain volumes make reliable indicators of intelligence? Why or why not?
About 3 million years ago, when A. afarensis had been in existence for nearly 1 million years, several new hominin species appeared in the fossil record. Scientists believe that Australopithecus africanus, one of these new species, was a hunting-and-gathering omnivore. A. africanus, like A. afarensis, was a gracile (or slender) hominin. Three more "robust" hominins (previously within Australopithecus, but now placed in a separate genus, Paranthropus) also appeared and are thought to have been savanna-dwelling vegetarians. The robust hominins had massive skulls, heavy facial bones, pronounced brows, and huge teeth. Whatever they ate, it required a lot of chewing. It is unclear whether Lucy's species, A. afarensis, gave rise to these other species or simply lived at the same time as they did. While the robust hominins appear to have been evolutionary dead ends, descendants of A. afarensis may have led to genus Homo.
Homo habilis. Homo habilis ("handy man"), the first member of the modern genus of humans, appeared in the fossil record about 2.5 million years ago. The remains classified as H. habilis are highly variable, causing some researchers to question their classification. Some researchers believe the remains are varied enough to represent more than one species. H. habilis differed from A. afarensis primarily in brain size. The cranial capacity of H. habilis has been estimated at between 500 and 800 cm3 (30 and 49 in.3). Some scientists hypothesize that H. habilis was the first hominin to use stone tools. Simple stone tools, dating from 2.5 million to 2.7 million years ago, have been found in Africa. Whether these tools were used by H. habilis or one of the species of robust hominins is unclear. H. habilis may have been capable of rudimentary speech. Casts of one brain made from reassembled skull fragments indicate a bulge in the area of the brain important in speech (see Chapter 8).
Homo ergaster and Homo erectus. a new hominin, Homo ergaster ("working man"), appeared in the fossil record about 1.9 million years ago. The name reflects the many tools found with the remains. Traditionally, scientists had classified these remains as Homo erectus ("upright man"), but they now differentiate them from H. erectus. H. ergaster appears to have originated in East Africa and coexisted there for several thousand years with some of the robust hominins.
H. erectus is thought to have diverged from H. ergaster around 1.6 million years ago. H. erectus was a wanderer, believed by many to be the first hominin to migrate out of Africa, spreading to Asia. H. erectus differed from earlier hominins in being larger (up to 1.85 m, or 6 ft tall, and weighing at least 65 kg, or 143 lb) and less sexually dimorphic. H. erectus had a brain volume of about 1000 cm3(61 in.3). Evidence indicates that H. erectus used sophisticated tools and weapons and may have used fire. H. erectus disappeared from most locations about 400.000 years ago, but some remains from Java have been dated at only 50,000 years. The Java remains suggest that at least one population of H. erectus existed at the same time as modern humans (Homo sapiens).
Homo heidelbergensis, Homo sapiens, and Homo neanderthalensis. The origins of anatomically modern humans over the last 500,000 years are difficult to trace with certainty, and different interpretations exist. Traditionally, fossils that did not quite resemble modern humans were classified as Archaic Homo sapiens. Scientists now place these fossils in the species Homo heidelbergensis; the name refers to Heidelberg, Germany, where a fossil lower jaw intermediate to those of earlier forms and H. sapiens was found. Many scientists now believe that H. heidelbergensis evolved from H. ergaster and not H. erectus. H. heidelbergensis ranges from about 800,000 years ago to about 130,000 years ago, which is when the first anatomically modern human remains show up in the fossil record. Thus, scientists postulate that H. sapiens and H. neanderthalensis evolved from H. heidelbergensis.
The oldest fossil evidence for modern humans (H. sapiens or "thinking man") comes from Africa and is about 130,000 years old. H. sapiens differs from earlier humans in having a larger brain (1300 cm3, or 79 in.3), flat forehead, absent or very small brow ridges, prominent chin, and a very slender body form.
Neanderthals, close evolutionary relatives of ours, are known from Europe and Asia from about 200,000 years ago to 30.000 years ago. Neanderthals had distinct features apparently adapted for life in a cold climate. Some Neanderthals lived in caves (Figure 22.20a). Neanderthal burial sites have been discovered, making them the first hominins known to have buried their dead. Also, the discovery of 50,000-year-old remains of sick, injured, and elderly individuals suggests that Neanderthals cared for the less fortunate among them.
Interestingly, Neanderthals had a larger braincase than do H. sapiens and a slightly larger brain volume (about 1450 cm3 or 88 in.3). However, these features may not correlate with intelligence but rather with the Neanderthals' more massive body. They had larger bones, suggesting heavier musculature, and rather short legs. They also had a thick brow ridge, large nose, broad face, and well-developed incisors and canines. Some anthropologists consider Neanderthals to be a subspecies of H. sapiens and call them H. sapiens neanderthalensis (according to this scheme, modern humans are known as H. sapiens sapiens). Most anthropologists, however, assign Neanderthals species status and call them Homo neanderthalensis.

FIGURE 22.20. Relatively recent representatives of the genus Homo. These photographs were taken at a museum display.
Neanderthals vanished from the fossil record some 30,000 years ago for still mysterious reasons. Some scientists suggest they were outcompeted or simply killed outright by a form of H. sapiens called Cro-Magnons (Figure 22.20b). Other scientists suggest that interbreeding between anatomically modern humans and Neanderthals might have resulted in the loss of the Neanderthal phenotype. Scientists now have a first version of the Neanderthal genome, developed from DNA recovered from the bones of three female Neanderthals whose remains were estimated at 38,000 years old. A comparison of the Neanderthal genome with those of present-day humans from different parts of the world suggests that following their migration from Africa 50,000 to 80,000 years ago, modern humans bred with Neanderthals in the Middle East before extending their range into Eurasia. The scientists estimate that 1% to 4% of the genes of present-day non-Africans came from Neanderthals.
In 2003, hominin remains were discovered in a cave on the island of Flores in Indonesia. The remains display a peculiar mix of primitive features (for example, small brain size) and derived features (for example, small canines). Some researchers suggest the remains represent a new hominin species, Homo floresiensis, estimated to have lived between 95,000 and 17,000 years ago (note that this species would have overlapped in time with H. sapiens). Others suggest the remains are from individuals of H. sapiens who suffered from a pathological condition such as microcephaly (a neurodevelopmental disorder characterized by a small head) or hypothyroidism (reduced growth and development due to undersecretion of thyroid hormone). Debate continues, as does the study of stone artifacts from the cave and nearby sites.
About 12,000 years ago, H. sapiens changed from a nomadic lifestyle to a more sedentary one. Associated with this change were two major milestones in human history: the domestication of animals and the cultivation of crops. For example, mammals were domesticated for protection (dogs), food (cattle, pigs, and goats), transport (horses, camels, donkeys), wool (llamas and alpacas), and rodent control (cats and ferrets). Agriculture began with the cultivation of cereal grains about 9000 years ago.
Major milestones in human evolution are summarized in Table 22.1, and the major hominin species are shown in Figure 22.21.
What would you do?
Human remains can tell scientists much about the diets, diseases, lifestyles, and genetic relationships of our ancestors. Such information can help piece together our evolutionary past. Sometimes, however, keeping human remains for scientific study conflicts with the wishes of modern-day descendants who wish to have their ancestors' remains returned to them for reburial. In the United States, many museums and universities are developing policies for the treatment and disposition of Native American and Native Hawaiian remains. How should human remains be treated, and who should get them? If you were a museum director developing a policy for the treatment of human remains, what would you do?
Highlighting the Concepts
Evolution of Life on Earth (pp. 469-471)
• Earth is estimated to be 4.5 billion years old, and life is thought to have originated about 3.8 billion years ago. A plausible sequence of events known as chemical evolution suggests that life arose from small organic molecules, synthesized from inorganic molecules. The small organic molecules then grew increasingly complex over a long period of time. A critical step in this process was the origin of genetic material (possibly RNA and not DNA), which allowed the transfer of information from one generation to the next. Eventually, large organic molecules aggregated into droplets that became the precursors to cells.
• The earliest cells were prokaryotic. According to the endosym- biont theory, more complex cells formed as smaller organisms were incorporated into the cells, forming organelles such as mitochondria. Multicellularity evolved, leading to the appearance on Earth of organisms such as plants and animals.
Scale of Evolutionary Change (pp. 471-475)
• Microevolution involves change in the frequencies of alleles within populations over a few generations. Macroevolution is large-scale evolution such as the origin or extinction of groups of species over long periods of time. Speciation is the formation of new species.
• Populations are groups of individuals of the same species that live in a particular area. Sexual reproduction and mutation produce variation in populations. Regarding sexual reproduction, sources of variation include crossing over and independent assortment during meiosis. Also, fertilization produces a zygote with new combinations of alleles. Mutations, rare changes in the DNA of genes, produce new genes and new alleles.
• A gene pool is a collection of all the alleles of all the genes of all the individuals in a population.
• Causes of microevolution include genetic drift (random change in allele frequencies due to chance within a small population), gene flow (movement of alleles as individuals move into and out of populations), mutation (change in the DNA of genes), and natural selection (mechanism of evolutionary change by which traits of successful individuals—those that survive and reproduce— become more common in a population).
• Systematic biology deals with the naming, classification, and evolutionary relationships of organisms. Each organism is given a two-part name consisting of the genus name followed by the specific epithet. Organisms also may be classified using a hierarchy of increasingly general categories: species, genus, family, order, class, phylum, kingdom, and domain. A phylogenetic tree is a schematic diagram depicting hypothesized relationships among species or higher categories.
Evidence of Evolution (pp. 475-480)
• Fossils are the preserved remnants and impressions of past organisms. The fossil record provides evidence of evolution by documenting that life on Earth has not always been the same as it is today.
• Biogeography is the study of the geographic distribution of organisms. New distributions of organisms occur either when the organisms disperse to a new location or when the areas they occupy move or are subdivided. Related species are more likely than are unrelated species to be found in the same geographic area.
• Comparative anatomy and embryology also provide evidence of evolution. Species with more shared traits are considered more likely to be related. Structures that have arisen from common ancestry are called homologous structures and usually arise from the same embryonic tissue. Thus, common embryological origins can also be viewed as evidence of common descent.
• Molecules that are the basic building blocks of life can be compared for evidence of evolution. For example, scientists compare the sequences of amino acids in proteins or the nucleotide sequences in DNA of different species to gauge relatedness and to estimate the time of divergence from a most recent common ancestor.
Human Evolution (pp. 480-487)
• Humans are primates, an order of mammals that also includes lemurs, monkeys, and apes. Primates have forward-facing eyes with stereoscopic vision, flexible shoulder joints, and grasping hands and feet. Flattened nails—rather than claws—cover their sensitive digits. Primates provide extensive parental care to a small number of offspring.
• There are two suborders of modern primates. One includes lemurs, lorises, and pottos, and the other includes monkeys, apes, and humans. The term hominin refers to the human lineage and its immediate ancestors, such as species within the genera Australopithecus and Homo.
• Humans did not descend from chimpanzees; rather, humans and chimps represent separate branches that diverged from a common ancestor. Human evolution did not occur in an orderly progression from ancient to modern forms; during several periods two or more species of hominins lived at the same time, and some of those species lines were evolutionary dead ends. Traits of humans did not evolve at the same rate; instead, evidence indicates that traits of humans evolved at different rates, by a phenomenon known as mosaic evolution.
• Bipedalism evolved early on in hominins. Walking on two feet set the stage for the evolution of other characteristics such as an increase in brain size, which in turn was associated with cultural trends such as tool use and language.
• The oldest hominin remains found to date are those of Ardipithecus ramidus, estimated to be at least 4.4 million years old. Few fossils have been found of Australopithecus anamensis, the most primitive australopithecine; this species is believed to have existed between 4.2 and 3.9 million years ago. Excellent specimens have been found of Australopithecus afarensis, a species about 3.8 million years old. About 3 million years ago, several new hominin species appeared (Australopithecus africanus and three more robust species in the genus Paranthropus).
• About 2.5 million years ago, Homo habilis remains appear in the fossil record; these were the first hominins to use tools. About 1.9 million years ago, Homo ergaster arose, and Homo erectus diverged from this species 1.6 million years ago. Fossils of
H. erectus are not restricted to Africa, indicating that this was the first hominin to disperse from Africa; this species may have used fire. The oldest remains of modern humans, Homo sapiens, are dated at 130,000 years old. Homo neanderthalensis was a cold- adapted species that coexisted with modern humans until about 30,000 years ago. Two major milestones of H. sapiens are the domestication of animals and the cultivation of crops.
• The multiregional hypothesis suggests that H. sapiens evolved independently in different regions from distinctive populations of early humans. The Out of Africa hypothesis suggests that modern humans evolved from early humans in Africa and then dispersed to other regions, where they replaced existing hominin species.
Reviewing the Concepts
1. How might life have evolved from inorganic molecules to complex cells? pp. 469-471
2. Distinguish between microevolution and macroevolution. p. 471
3. What are four sources of variation within populations? p. 472
4. How does genetic drift lead to microevolution? p. 472
5. Define speciation and relate it to gene flow. pp. 472-473
6. Define natural selection. How can variation within populations be maintained in the face of natural selection? p. 473
7. Describe the binomial system by which organisms are named and the hierarchical system by which they are classified. pp. 473-474
8. What is a phylogenetic tree? p. 475
9. What is a fossil? Describe the process of fossilization and relate it to limitations of the fossil record. pp. 475-476
10. How do new distributions of organisms arise? p. 477
11. Distinguish between homologous and analogous structures. р. 478
12. Describe how comparative embryology provides evidence of evolution. pp. 478-480
13. What is a molecular clock? p. 480
14. What characteristics set primates apart from other mammals? pp. 480-481
15. Name four skeletal differences between chimpanzees and humans. pp. 481-483
16. What are three popular misconceptions about human evolution? p. 483
17. Describe the major species of hominins and trends in hominin evolution. pp. 483-487
18. Which of the following does not produce variation in populations?
a. Bottleneck effect
b. Mutation
с. Crossing over
d. Independent assortment
19. Which of the following occurs when fertile individuals move into and out of populations?
a. Genetic drift
b. Speciation
c. Mutation
d. Gene flow
20. Which of the following was in very short supply in the environment of the early Earth?
a. Lightning
b. Volcanoes
c. Gaseous oxygen (O2)
d. UV radiation
21. The earliest cells
a. were prokaryotic.
b. were eukaryotic.
c. evolved over 10 billion years ago.
d. were part of multicellular organisms.
22. Which of the following primates is our closest living relative?
a. Gorilla
b. Orangutan
c. Chimpanzee
d. Gibbon
23. Which of the following features of an organism would promote fossilization?
a. Terrestrial existence
b. Member of a small population
c. Soft body parts
d. Aquatic existence
24. Which hominin was probably the first to migrate out of Africa?
a. Australopithecus afarensis
b. Homo sapiens
c. Homo erectus
d. Australopithecus africanus
25. The major events of human evolution occurred in
a. Africa.
b. South America.
c. Europe.
d. North America.
26. Which of the following hominin characters evolved the earliest?
a. Bipedalism
b. Large brain
c. Language
d. Use of fire
27. _____ is differential survival and reproduction.
28. _____ is when different traits evolve at different rates.
29. The _____ hypothesis of human evolution suggests that modern humans evolved independently in several places from local populations of earlier humans.
Applying the Concepts
1. A friend of yours believes that all organisms were specially created. You believe in evolution and want to present your case to your friend. What would you say?
2. Can life arise on Earth from inorganic material today? Why or why not?
3. You are a biologist exploring the dense tropical rain forests of Brazil. You notice an unfamiliar medium-sized mammal moving above you in the trees. What characteristics should this animal display to be classified as a primate?
4. Tay-Sachs disease has zero fitness because it causes death before the individual reaches reproductive age. How does a trait with zero fitness persist in a population?
5. Did modern humans evolve from chimpanzees? Explain.
6. Apes have a fairly easy time giving birth, compared with humans. A female ape has a relatively large pelvis, and the small head of her infant passes easily through a roomy birth canal (the channel formed by the cervix, vagina, and vulva). Birth in humans is decidedly more difficult and risky. With its large head, the typical human fetus may spend hours making its way through a narrow birth canal. What trends in human evolution might have produced the tight squeeze of human birth?
Becoming Information Literate
Develop a PowerPoint presentation on tool use in extant vertebrates. Include the following in your presentation: a definition of tool use, the species in which tool use has been reported, the type of tool use described and the conditions under which it was observed, and any anatomical or behavioral traits associated with species that make and use tools. Use at least three sources; list these sources in your last slide, and briefly explain why you found them to be reliable.